Effect of Cover Crop on Carbon Distribution in Size and Density Separated Soil Aggregates

 , , , , , ,
, , , , , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Field Setting and Sample Collection
2.2. Aggregate Size Separation
2.3. Aggregate Density Separation
2.4. Elemental and Isotopic Analysis
2.5. X-ray Absorption Spectroscopy
2.6. Chemical Extractions
2.7. Statistics
3. Results
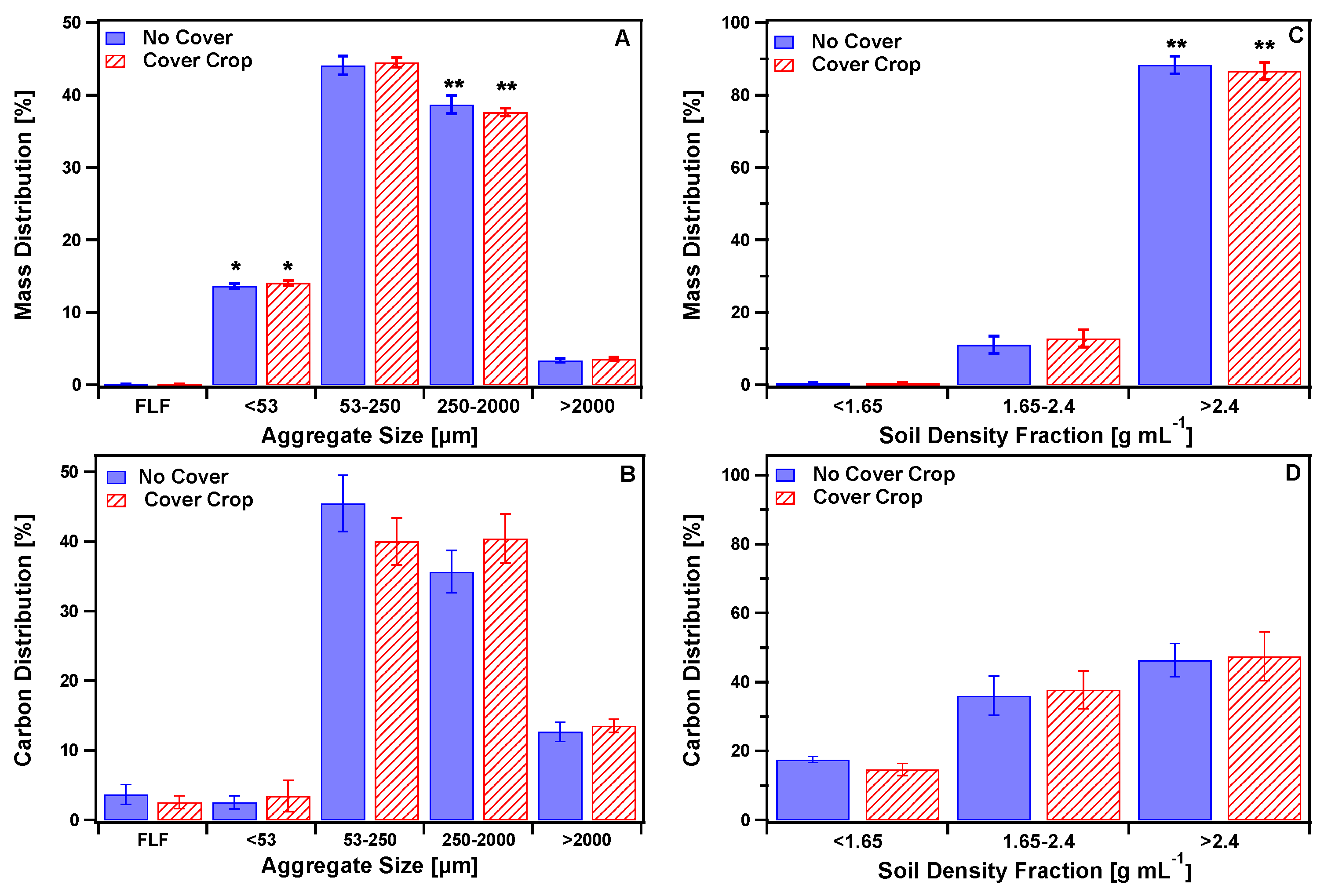
3.1. Mass, Carbon, and Nitrogen Distribution in Aggregates
3.1.1. Size Separated Aggregates
3.1.2. Density Separated Aggregates
3.1.3. Sequential Size-Density Separation
3.2. Carbon and Nitrogen Isotopes
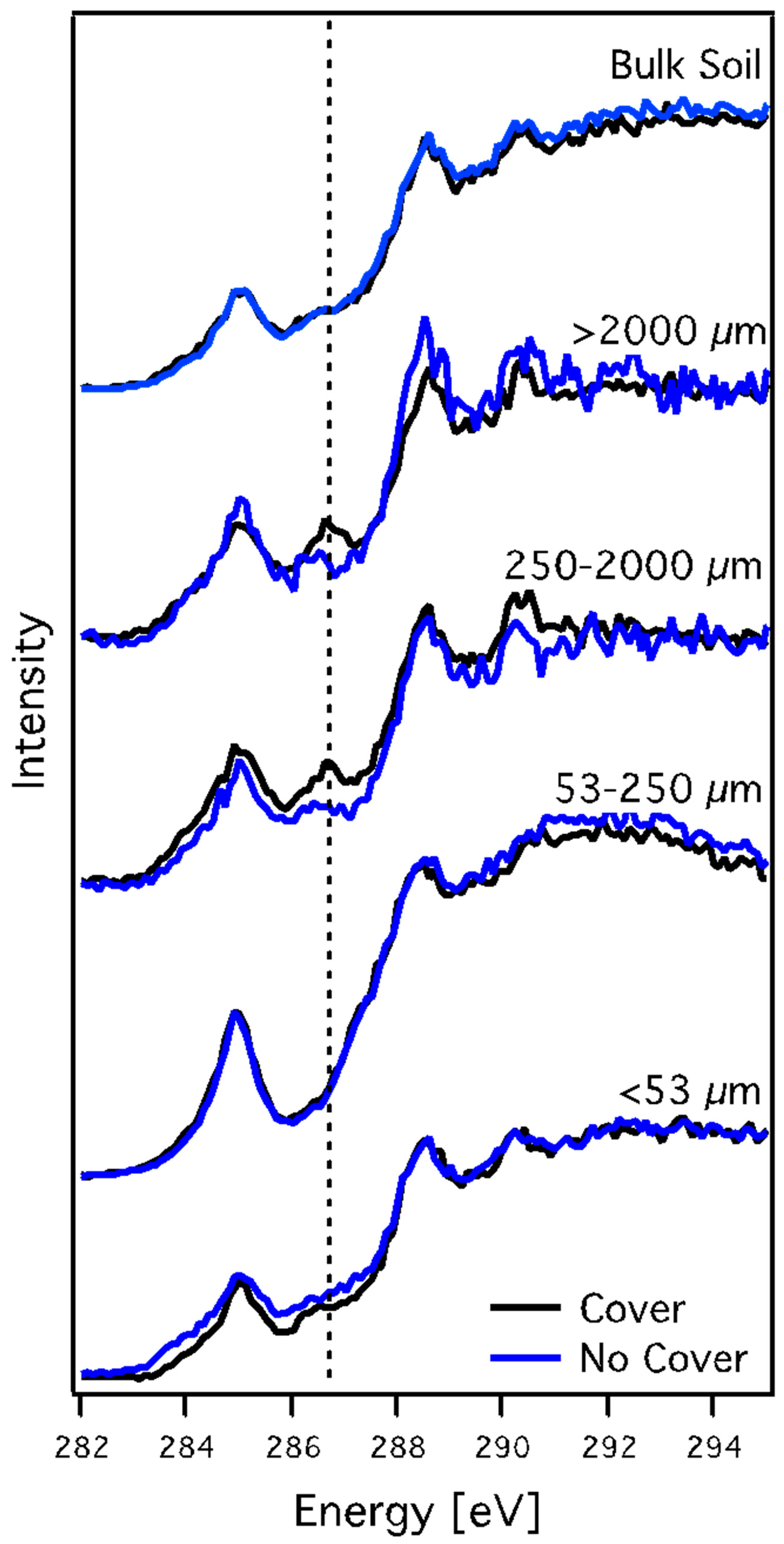
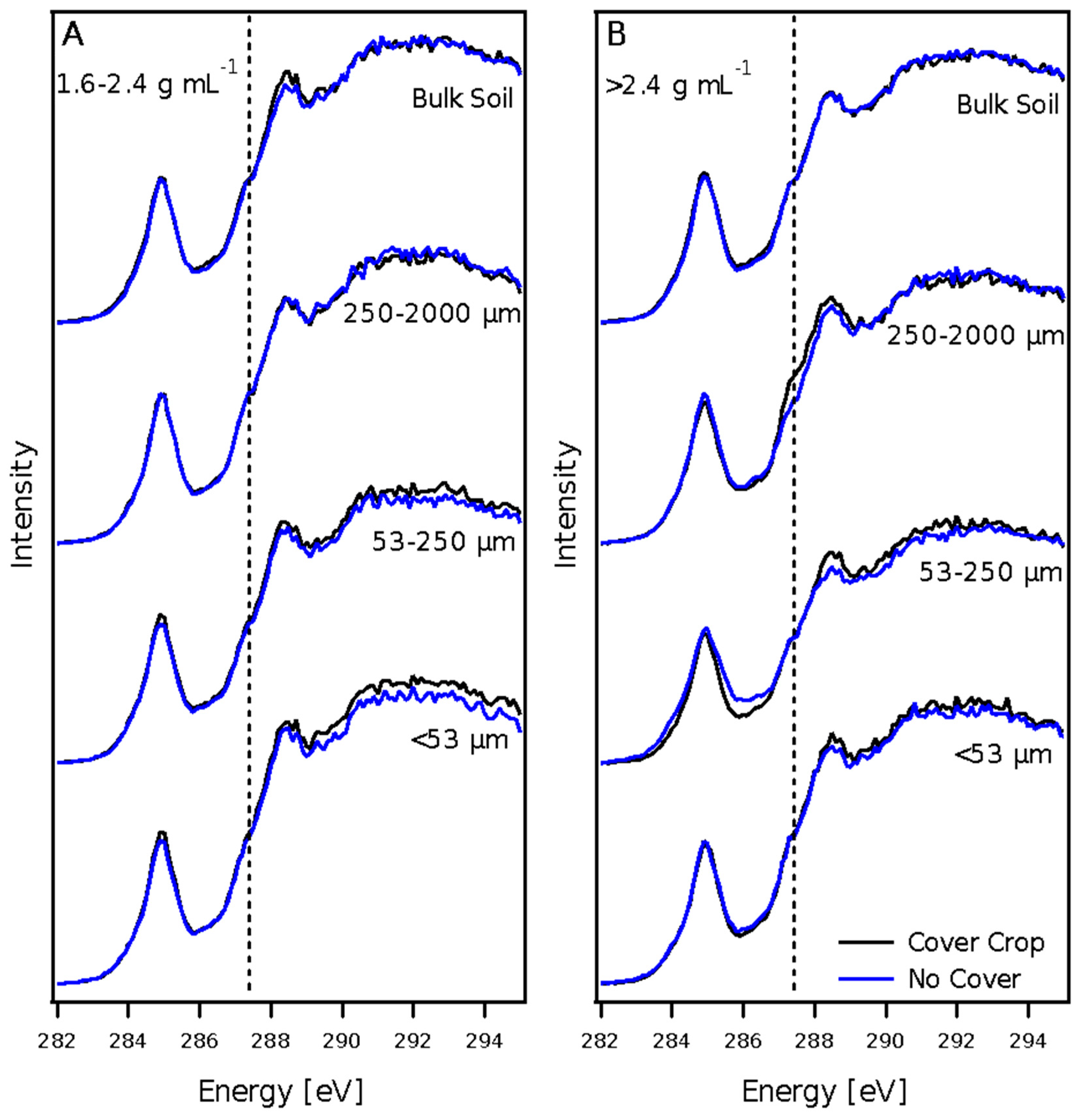
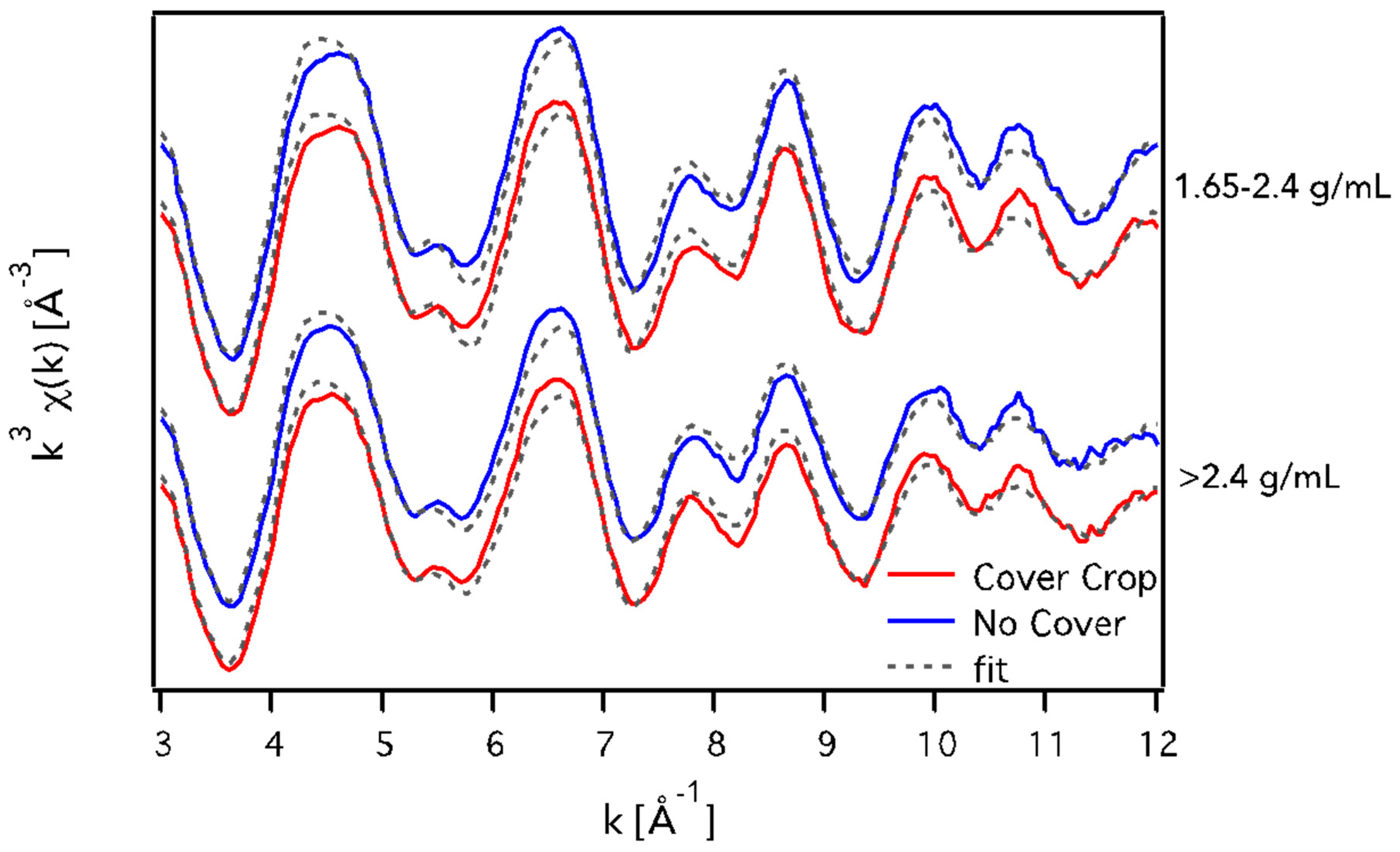
3.3. X-ray Absorption Spectroscopy
3.3.1. Carbon NEXAFS Spectroscopy
3.3.2. Fe K-Edge EXAFS Spectroscopy
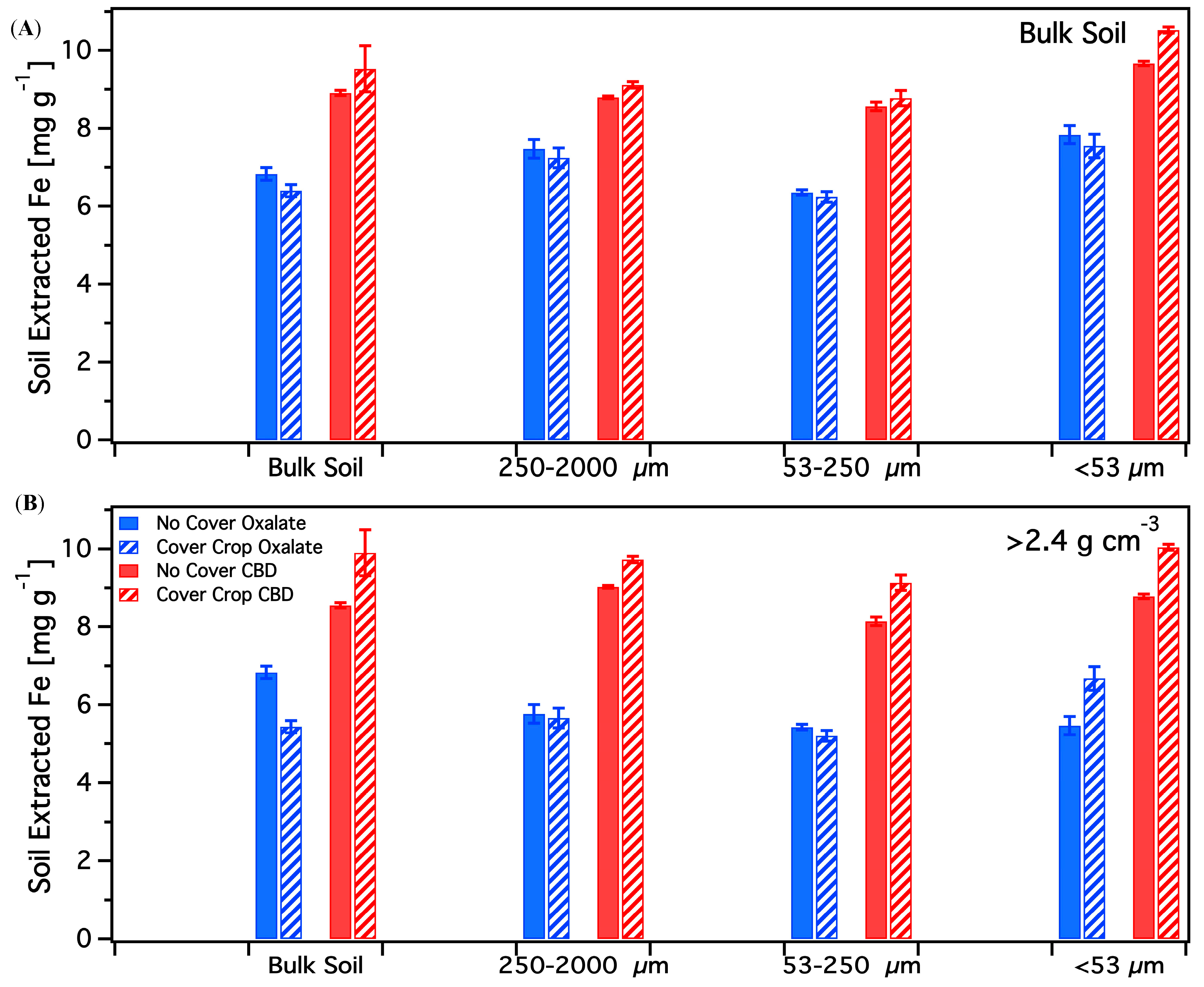
3.4. Chemical Extractions
3.4.1. Acid Ammonium Oxalate
3.4.2. Citrate-Bicarbonate-Dithionite
4. Discussion
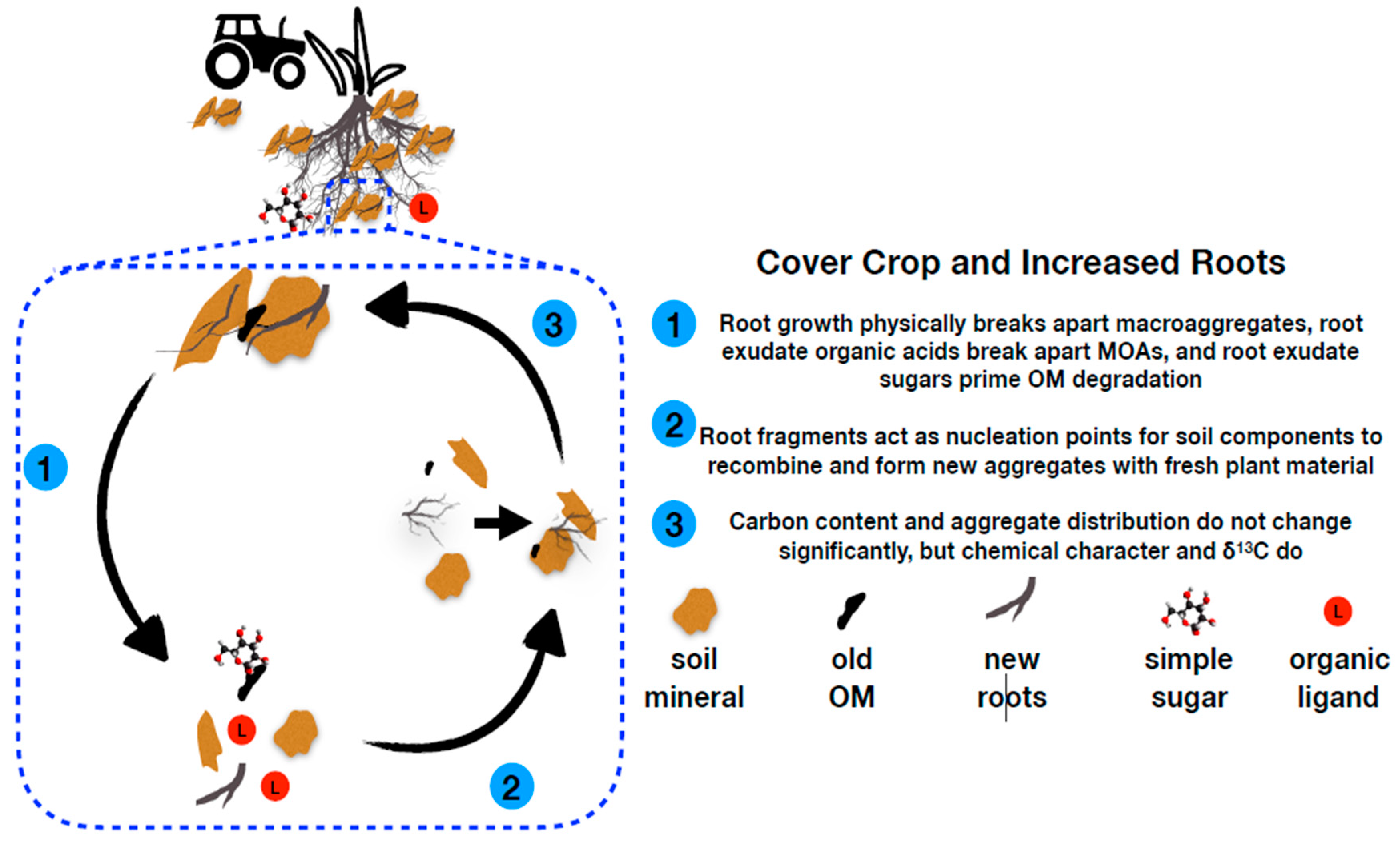
4.1. Cover Crop Effect on Carbon Inputs
4.2. Aggregate Dynamics
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Minasny, B.; Malone, B.P.; McBratney, A.B.; Angers, D.A.; Arrouays, D.; Chambers, A.; Chaplot, V.; Chen, Z.-S.; Cheng, K.; Das, B.S.; et al. Soil carbon 4 per mille. Geoderma 2017, 292, 59–86. [Google Scholar] [CrossRef]
- Poeplau, C.; Don, A. Carbon sequestration in agricultural soils via cultivation of cover crops—A meta-analysis. Agric. Ecosyst. Environ. 2015, 200, 33–41. [Google Scholar] [CrossRef]
- Mayer, A.; Hausfather, Z.; Jones, A.D.; Silver, W.L. The potential of agricultural land management to contribute to lower global surface temperatures. Sci. Adv. 2018, 4, eaaq0932. [Google Scholar] [CrossRef] [PubMed]
- Tisdall, J.M.; Oades, J.M. Organic matter and water-stable aggregates in soils. J. Soil Sci. 1982, 33, 141–163. [Google Scholar] [CrossRef]
- Rorick, J.D.; Kladivko, E.J. Cereal rye cover crop effects on soil carbon and physical properties in southeastern Indiana. J. Soil Water Conserv. 2017, 72, 260–265. [Google Scholar] [CrossRef]
- Kabir, Z.; Koide, R.T. The effect of dandelion or a cover crop on mycorrhizal inoculum potential, soil aggregation and yield of maize. Agric. Ecosyst. Environ. 2000, 78, 167–174. [Google Scholar] [CrossRef]
- Roberson, E.B.; Firestone, M.K.; Sarig, S. Cover Crop Management of Polysaccharide-Mediated Aggregation in an Orchard Soil. Soil Sci. Soc. Am. J. 1991, 55, 734–739. [Google Scholar] [CrossRef]
- Kong, A.Y.Y.; Six, J. Tracing Root vs. Residue Carbon into Soils from Conventional and Alternative Cropping Systems. Soil Sci. Soc. Am. J. 2010, 74, 1201–1210. [Google Scholar] [CrossRef]
- Kong, A.Y.Y.; Six, J. Microbial community assimilation of cover crop rhizodeposition within soil microenvironments in alternative and conventional cropping systems. Plant Soil 2012, 356, 315–330. [Google Scholar] [CrossRef]
- Berhe, A.A.; Suttle, K.B.; Burton, S.D.; Banfield, J.F. Contingency in the direction and mechanics of soil organic matter responses to increased rainfall. Plant Soil 2012, 358, 371–383. [Google Scholar] [CrossRef]
- Hall, S.J.; Berhe, A.A.; Thompson, A. Order from disorder: Do soil organic matter composition and turnover co-vary with iron phase crystallinity? Biogeochemistry 2018, 140, 93–110. [Google Scholar] [CrossRef]
- Eusterhues, K.; Neidhardt, J.; Hädrich, A.; Küsel, K.; Totsche, K.U. Biodegradation of ferrihydrite-associated organic matter. Biogeochemistry 2014, 119, 45–50. [Google Scholar] [CrossRef]
- Duiker, S.W.; Rhoton, F.E.; Torrent, J.; Smeck, N.E.; Lal, R. Iron (Hydr)Oxide Crystallinity Effects on Soil Aggregation. Soil Sci. Soc. Am. J. 2003, 67, 606–611. [Google Scholar] [CrossRef]
- Oades, J.M. Soil organic matter and structural stability: Mechanisms and implications for management. Plant Soil 1984, 76, 319–337. [Google Scholar] [CrossRef]
- Keiluweit, M.; Bougoure, J.J.; Nico, P.S.; Pett-Ridge, J.; Weber, P.K.; Kleber, M. Mineral protection of soil carbon counteracted by root exudates. Nat. Clim. Chang. 2015, 5, 588–595. [Google Scholar] [CrossRef]
- Six, J.; Paustian, K.; Elliott, E.T.; Combrink, C. Soil Structure and Organic Matter I. Distribution of Aggregate-Size Classes and Aggregate-Associated Carbon. Soil Sci. Soc. Am. J. 2000, 64, 681–689. [Google Scholar] [CrossRef]
- Sollins, P.; Swanston, C.; Kleber, M.; Filley, T.; Kramer, M.; Crow, S.; Caldwell, B.A.; Lajtha, K.; Bowden, R. Organic C and N stabilization in a forest soil: Evidence from sequential density fractionation. Soil Biol. Biochem. 2006, 38, 3313–3324. [Google Scholar] [CrossRef]
- Moni, C.; Derrien, D.; Hatton, P.J.; Zeller, B.; Kleber, M. Density fractions versus size separates: Does physical fractionation isolate functional soil compartments? Biogeosciences 2012, 9, 5181–5197. [Google Scholar] [CrossRef]
- Hatton, P.-J.; Kleber, M.; Zeller, B.; Moni, C.; Plante, A.F.; Townsend, K.; Gelhaye, L.; Lajtha, K.; Derrien, D. Transfer of litter-derived N to soil mineral–organic associations: Evidence from decadal 15N tracer experiments. Org. Geochem. 2012, 42, 1489–1501. [Google Scholar] [CrossRef]
- Golchin, A.; Oades, J.; Skjemstad, J.; Clarke, P. Study of free and occluded particulate organic matter in soils by solid state 13C Cp/MAS NMR spectroscopy and scanning electron microscopy. Soil Res. 1994, 32, 285–309. [Google Scholar] [CrossRef]
- Golchin, A.; Oades, J.M.; Skjemstad, J.O.; Clarke, P.; Golchin, A.; Oades, J.M.; Skjemstad, J.O.; Clarke, P. Soil structure and carbon cycling, Soil structure and carbon cycling. Soil Res. Soil Res. 1994, 32, 1043–1068. [Google Scholar] [CrossRef]
- Regier, T.; Krochak, J.; Sham, T.K.; Hu, Y.F.; Thompson, J.; Blyth, R.I.R. Performance and capabilities of the Canadian Dragon: The SGM beamline at the Canadian Light Source. Nucl. Instrum. Methods Phys. Res. Sect. A Accel. Spectrometers Detect. Assoc. Equip. 2007, 582, 93–95. [Google Scholar] [CrossRef]
- Gillespie, A.W.; Phillips, C.L.; Dynes, J.J.; Chevrier, D.; Regier, T.Z.; Peak, D. Chapter One—Advances in Using Soft X-Ray Spectroscopy for Measurement of Soil Biogeochemical Processes. In Advances in Agronomy; Sparks, D.L., Ed.; Academic Press: Cambridge, MA, USA, 2015; Volume 133, pp. 1–32. [Google Scholar]
- Ravel, B.; Newville, M. Athena, Artemis, Hephaestus: Data analysis for X-ray absorption spectroscopy using IFEFFIT. J. Synchrotron Radiat. 2005, 12, 537–541. [Google Scholar] [CrossRef] [PubMed]
- McKeague, J.A.; Day, J.H. Dithionite and oxalate extractable Fe and Al as in differentiating various classes of soils. Can. J. Soil. Sci. 1966, 46, 13–22. [Google Scholar] [CrossRef]
- Degens, B.P. Macro-aggregation of soils by biological bonding and binding mechanisms and the factors affecting these: A review. Soil Res. 1997, 35, 431–460. [Google Scholar] [CrossRef]
- Angers, D.A.; Caron, J. Plant-induced Changes in Soil Structure: Processes and Feedbacks. Biogeochemistry 1998, 42, 55–72. [Google Scholar] [CrossRef]
- Six, J.; Bossuyt, H.; Degryze, S.; Denef, K. A history of research on the link between (micro)aggregates, soil biota, and soil organic matter dynamics. Soil Tillage Res. 2004, 79, 7–31. [Google Scholar] [CrossRef]
- Helal, H.M.; Sauerbeck, D.R. Influence of plant roots on C and P metabolism in soil, Developments in Plant and Soil Sciences. In Biological Processes and Soil Fertility; Tinsley, J., Darbyshire, J.F., Eds.; Springer: Dordrecht, The Netherlands, 1984; pp. 175–182. [Google Scholar]
- Gale, W.J.; Cambardella, C.A.; Bailey, T.B. Root-Derived Carbon and the Formation and Stabilization of Aggregates. Soil Sci. Soc. Am. J. 2000, 64, 201–207. [Google Scholar] [CrossRef]
- Daynes, C.N.; Field, D.J.; Saleeba, J.A.; Cole, M.A.; McGee, P.A. Development and stabilisation of soil structure via interactions between organic matter, arbuscular mycorrhizal fungi and plant roots. Soil Biol. Biochem. 2013, 57, 683–694. [Google Scholar] [CrossRef]
- Denef, K.; Six, J.; Bossuyt, H.; Frey, S.D.; Elliott, E.T.; Merckx, R.; Paustian, K. Influence of dry–wet cycles on the interrelationship between aggregate, particulate organic matter, and microbial community dynamics. Soil Biol. Biochem. 2001, 33, 1599–1611. [Google Scholar] [CrossRef]
- Denef, K.; Six, J. Clay mineralogy determines the importance of biological versus abiotic processes for macroaggregate formation and stabilization. Eur. J. Soil Sci. 2005, 56, 469–479. [Google Scholar] [CrossRef]
- Banfield, J.F.; Barker, W.W.; Welch, S.A.; Taunton, A. Biological impact on mineral dissolution: Application of the lichen model to understanding mineral weathering in the rhizosphere. Proc. Natl. Acad. Sci. USA 1999, 96, 3404–3411. [Google Scholar] [CrossRef] [PubMed]
- Easterwood, G.W.; Sartain, J.B. Clover Residue Effectiveness in Reducing Orthophosphate Sorption on Ferric Hydroxide Coated Soil. Soil Sci. Soc. Am. J. 1990, 54, 1345–1350. [Google Scholar] [CrossRef]
- Gerke, J. Phosphate adsorption by humic/Fe-oxide mixtures aged at pH 4 and 7 and by poorly ordered Fe-oxide. Geoderma 1993, 59, 279–288. [Google Scholar] [CrossRef]
- Miller, R.M.; Jastrow, J.D. Mycorrhizal Fungi Influence Soil Structure. In Arbuscular Mycorrhizas: Physiology and Function; Kapulnik, Y., Douds, D.D., Eds.; Springer: Dordrecht, The Netherlands, 2000; pp. 3–18. [Google Scholar]
- Schwertmann, U. Solubility and dissolution of iron oxides. Plant Soil 1991, 130, 1–25. [Google Scholar] [CrossRef]
- Schwertmann, U.; Friedl, J.; Stanjek, H. From Fe(III) Ions to Ferrihydrite and then to Hematite. J. Colloid Interface Sci. 1999, 209, 215–223. [Google Scholar] [CrossRef]
- Hansel, C.M.; Benner, S.G.; Fendorf, S. Competing Fe(II)-induced mineralization pathways of ferrihydrite. Environ. Sci. Technol. 2005, 39, 7147–7153. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| C | δ13C | N | δ15N | C/N | Mg | Al | Si | P | S | K | Ca | Mn | Fe | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| % | ‰ | % | ‰ | % | % | % | % | Ppm | % | % | Ppm | % | |||
| Bulk Soil | NCC 1 | 1.00(2) 3 | −25.2 ** | 0.11 | 4.1(2) | 11.1 | 3.4 | 5.3 | 24.1 | 0.0 | 270 | 1.3 | 0.8 | 1010 | 5.2 |
| CC 2 | 1.04(4) | −25.6 ** | 0.11 | 3.8 | 11.3(5) | 3.6 | 5.4 | 24.6 | 0.1 | 280 | 1.3 | 0.8 | 1030 | 5.2 | |
| >2000 µm | NCC | 1.28(6) | −25.7(4) | 0.15 | 4.2(6) | 9.8(2) | 2.9 | 4.8 | 21.7 | 0.0 | 250 | 1.2 | 0.8 | 920 | 5.0 |
| CC | 1.81(29) | −26.7(4) | 0.19(2) | 4.1(3) | 11.1(6) | 3.1 | 4.7 | 21.0 | 0.0 | 260 | 1.1 | 0.7 | 900 | 4.8 | |
| 250–2000 µm | NCC | 1.24(5) | −25.1 ** | 0.12 | 6.5(6) | 12.2(2) | 1.8 | 4.0 | 19.5 | 0.0 | 289 | 1.1 | 0.8 | 951 | 5.4 |
| CC | 1.25(4) | −25.4 ** | 0.12 | 5.9(3) | 12.6(2) | 3.2 | 5.0 | 22.3 | 0.0 | 250 | 1.2 | 0.8 | 949 | 5.5 | |
| 53–250 µm | NCC | 0.94(2) | −25.1 | 0.09 | 6.8(5) | 11.7(2) | 3.1 | 5.0 | 22.2 | 0.0 | 419 | 1.2 | 0.8 | 918 | 4.9 |
| CC | 1.01(2) | −25.3 | 0.1 | 6.1(4) | 12.3(3) | 3.1 | 4.8 | 21.7 | 0.0 | 403 | 1.2 | 0.8 | 909 | 4.8 | |
| <53 µm | NCC | 1.09(4) | −25.3 | 0.13 | 5.1(4) | 9.9(2) | 3.4 | 5.9 | 24.1 | 0.1 | 621 | 1.2 | 0.8 | 1168 | 5.5 |
| CC | 1.07 | −25.4 | 0.13 | 4.6(5) | 10.0(2) | 3.1 | 5.5 | 22.1 | 0.1 | 601 | 1.2 | 0.8 | 1163 | 5.5 | |
| FLF | NCC | 27.3 | −22.4 | 1.38 | 1.2 | 23.2 | |||||||||
| CC | 26.0 | −27.2 | 1.22 | −0.2 | 25.8 |
| Starting Material | Density 1 g cm−3 | C % | δ13C ‰ | N % | δ15N ‰ | C/N | |
|---|---|---|---|---|---|---|---|
| NCC 2 | Bulk Soil | <1.65 | 27(2) 4 | −24.6(5) * | 1.4 | 1.6(4) | 21.8 * |
| 1.65–2.4 | 4.6(6) | −24.8 ** | 0.4(2) | 2.8 | 12.4(2) | ||
| >2.4 | 0.5 | −24.9 ** | 0.1 | 4.1(2) ** | 7.7 * | ||
| CC 3 | Bulk Soil | <1.65 | 26(2) | −25.7(2) * | 1.6 | 1.2(2) | 19.5(6) * |
| 1.65–2.4 | 4.3(7) | −25.3 ** | 0.4 | 2.9 | 12.6 | ||
| >2.4 | 0.5 | −25.0 ** | 0.1 | 4.3 ** | 7.7 * | ||
| NCC | 250–2000 µm | <1.65 | 26 | 23.7 | 1.5 | 0.8 | 20.5 |
| 1.65–2.4 | 2.6 | −24.9 | 0.2 | 3.0 | 12.8 | ||
| >2.4 | 0.6 | −24.8 | 0.1 | 4.5 | 9.9 | ||
| CC | 250–2000 µm | <1.65 | 32 | −24.9 | 1.9 | 0.2 | 19.4 |
| 1.65–2.4 | 2.5 | −25.5 | 0.2 | 2.6 | 12.4 | ||
| >2.4 | 0.6 | −25.0 | 0.1 | 4.0 | 9.8 | ||
| NCC | 53–250 µm | <1.65 | 29 | −24.1 | 1.6 | 1.9 | 21.5 |
| 1.65–2.4 | 2.1 | −24.9 | 0.2 | 3.1 | 12.1 | ||
| >2.4 | 0.4 | −24.7 | 0.1 | 4.7 | 8.6 | ||
| CC | 53–250 µm | <1.65 | 38 | −24.0 | 2.0 | −0.8 | 22.2 |
| 1.65–2.4 | 2.3 | −25.2 | 0.2 | 2.9 | 12.3 | ||
| >2.4 | 0.4 | −24.9 | 0.1 | 3.4 | 8.3 | ||
| NCC | <53 µm | <1.65 | 36 | −25.3 | 1.7 | 2.4 | 21.7(4) |
| 1.65–2.4 | 5.0(7) | −25.1 ** | 0.4 | 3.0 ** | 13.7(2) ** | ||
| >2.4 | 0.5 | −24.9 | 0.1 | 4.2 | 7.2 | ||
| CC | <53 µm | <1.65 | 28(6) | −25.6(4) | 1.3(3) | 1.3(8) | 25.3(4) |
| 1.65–2.4 | 7(1) | −25.4 ** | 0.6 | 3.1 ** | 14.8(2) ** | ||
| >2.4 | 0.5 | −24.9 | 0.1 | 4.4(2) | 7.3 |
| Treatment | Density (g cm−3) | Goethite | Hornblende | Nontronite | Phlogopite | Vermiculite | x2 |
|---|---|---|---|---|---|---|---|
| No Cover | 1.65–2.4 | 17% | 23% | 43% | 12% | 6% | 0.43 |
| Cover Crop | 1.65–2.4 | 18% | 25% | 40% | 14% | 4% | 0.38 |
| No Cover | >2.4 | 21% | 32% | 27% | 4% | 16% | 0.33 |
| Cover Crop | >2.4 | 19% | 28% | 31% | 3% | 19% | 0.31 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schaefer, M.V.; Bogie, N.A.; Rath, D.; Marklein, A.R.; Garniwan, A.; Haensel, T.; Lin, Y.; Avila, C.C.; Nico, P.S.; Scow, K.M.; et al. Effect of Cover Crop on Carbon Distribution in Size and Density Separated Soil Aggregates. Soil Syst. 2020, 4, 6. https://doi.org/10.3390/soilsystems4010006
Schaefer MV, Bogie NA, Rath D, Marklein AR, Garniwan A, Haensel T, Lin Y, Avila CC, Nico PS, Scow KM, et al. Effect of Cover Crop on Carbon Distribution in Size and Density Separated Soil Aggregates. Soil Systems. 2020; 4(1):6. https://doi.org/10.3390/soilsystems4010006
Chicago/Turabian StyleSchaefer, Michael V., Nathaniel A. Bogie, Daniel Rath, Alison R. Marklein, Abdi Garniwan, Thomas Haensel, Ying Lin, Claudia C. Avila, Peter S. Nico, Kate M. Scow, and et al. 2020. "Effect of Cover Crop on Carbon Distribution in Size and Density Separated Soil Aggregates" Soil Systems 4, no. 1: 6. https://doi.org/10.3390/soilsystems4010006
APA StyleSchaefer, M. V., Bogie, N. A., Rath, D., Marklein, A. R., Garniwan, A., Haensel, T., Lin, Y., Avila, C. C., Nico, P. S., Scow, K. M., Brodie, E. L., Riley, W. J., Fogel, M. L., Berhe, A. A., Ghezzehei, T. A., Parikh, S., Keiluweit, M., & Ying, S. C. (2020). Effect of Cover Crop on Carbon Distribution in Size and Density Separated Soil Aggregates. Soil Systems, 4(1), 6. https://doi.org/10.3390/soilsystems4010006