Soil Oxygen Limits Microbial Phosphorus Utilization in Humid Tropical Forest Soils


{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Methods
2.1. Site Description
2.2. Incubation Experiment
2.3. Iron Measurements
2.4. Soil CO2 Measurements
2.5. Phosphorus Measurements
2.6. Single Point Sorption Test
2.7. Statistical Analysis
3. Results
3.1. Iron Species
3.2. Soil CO2 Production
3.3. Phosphorus Pools
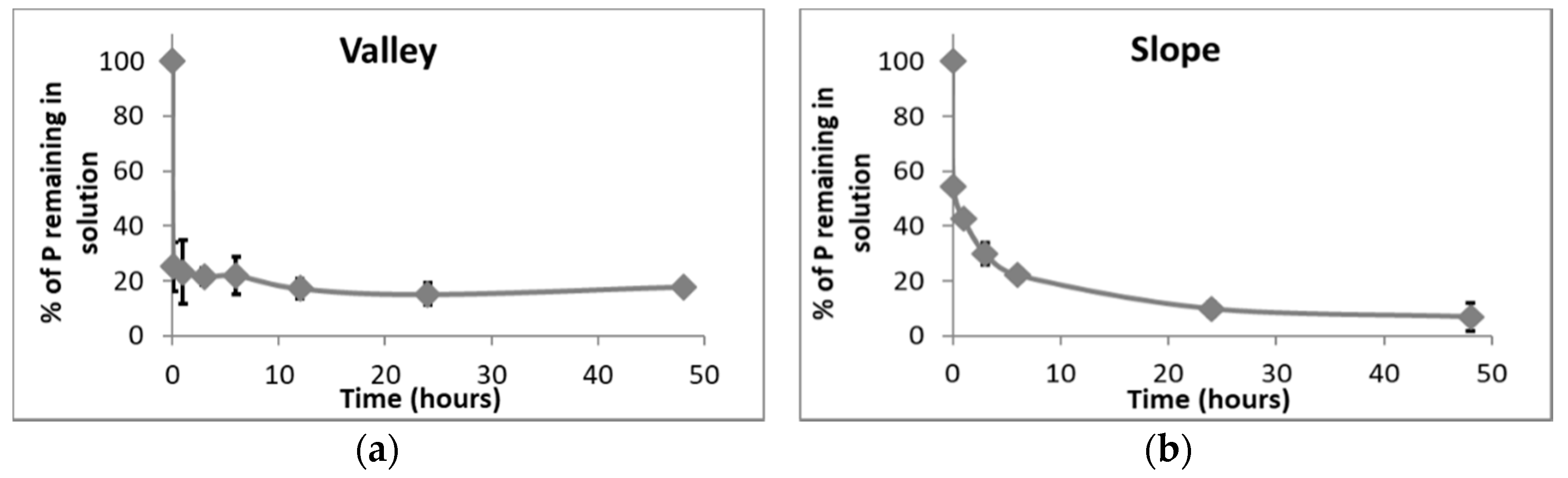
3.4. Single Point P Sorption Test
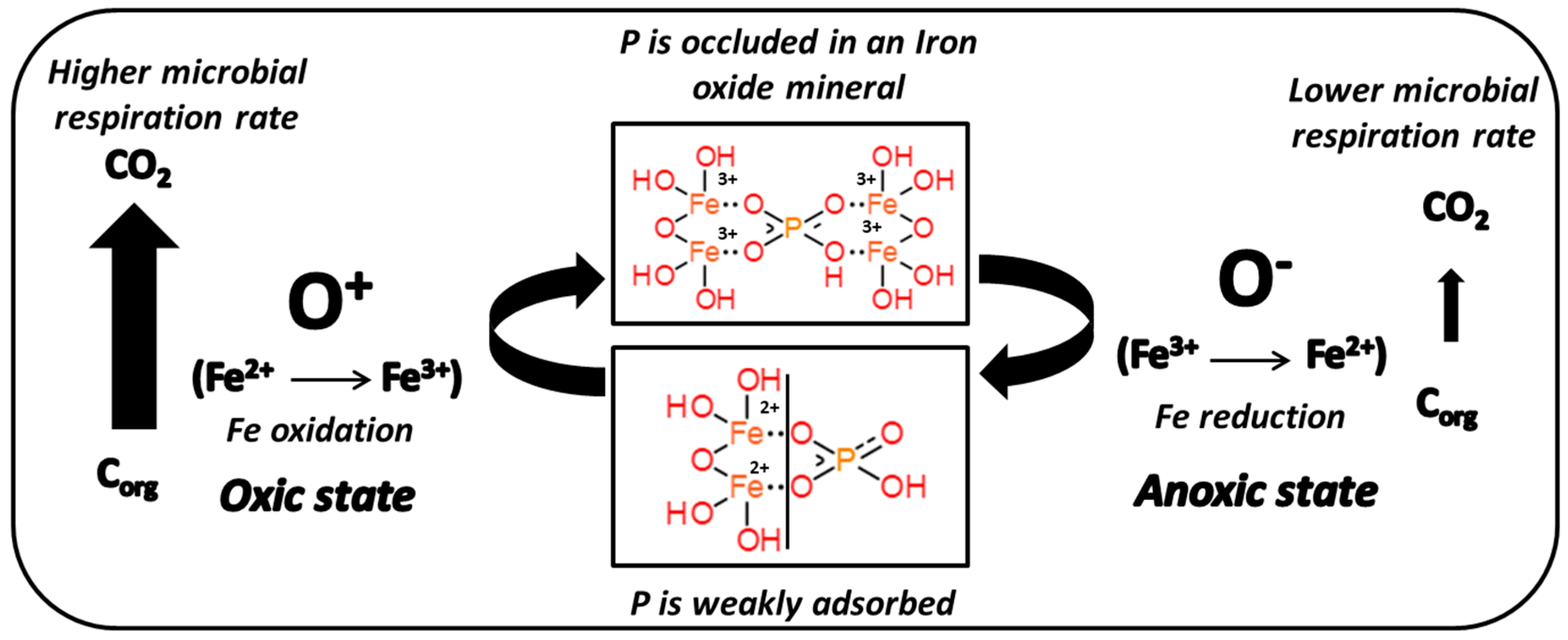
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cleveland, C.C.; Townsend, A.R.; Taylor, P.; Alvarez-Clare, S.; Bustamante, M.M.; Chuyong, G.; Dobrowski, S.Z.; Grierson, P.; Harms, K.E.; Houlton, B.Z.; et al. Relationships among net primary productivity, nutrients and climate in tropical rain forest: A pan-tropical analysis. Ecol. Lett. 2011, 14, 1313–1317. [Google Scholar] [CrossRef]
- Vitousek, P.M.; Porder, S.; Houlton, B.Z.; Chadwick, O.A. Terrestrial phosphorus limitation: Mechanisms, implications, and nitrogen-phosphorus interactions. Ecol. Appl. 2010, 20, 5–15. [Google Scholar] [CrossRef] [PubMed]
- Nottingham, A.T.; Turner, B.L.; Stott, A.W.; Tanner, E.V. Nitrogen and phosphorus constrain labile and stable carbon turnover in lowland tropical forest soils. Soil Boil. Biochem. 2015, 80, 26–33. [Google Scholar] [CrossRef]
- Cleveland, C.C.; Townsend, A.R. Nutrient additions to a tropical rain forest drive substantial soil carbon dioxide losses to the atmosphere. Proc. Natl. Acad. Sci. USA 2006, 103, 10316–10321. [Google Scholar] [CrossRef] [PubMed]
- Kaspari, M.; Garcia, M.N.; Harms, K.E.; Santana, M.; Wright, S.J.; Yavitt, J.B. Multiple nutrients limit litterfall and decomposition in a tropical forest. Ecol. Lett. 2008, 11, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Cleveland, C.C.; Townsend, A.R.; Schmidt, S.K. Phosphorus limitation of microbial processes in moist tropical forests: Evidence from short-term laboratory incubations and field studies. Ecosystems 2002, 5, 0680–0691. [Google Scholar] [CrossRef]
- Pan, Y.; Birdsey, R.A.; Phillips, O.L.; Jackson, R.B. The structure, distribution, and biomass of the world’s forests. Annu. Rev. Ecol. Evol. Syst. 2013, 44, 593–622. [Google Scholar] [CrossRef]
- Jobbágy, E.G.; Jackson, R.B. The vertical distribution of soil organic carbon and its relation to climate and vegetation. Ecol. Appl. 2000, 10, 423–436. [Google Scholar] [CrossRef]
- Reed, S.C.; Yang, X.; Thornton, P.E. Incorporating phosphorus cycling into global modeling efforts: A worthwhile, tractable endeavor. New Phytol. 2015, 208, 324–329. [Google Scholar] [CrossRef] [PubMed]
- Miller, A.J.; Schuur, E.A.; Chadwick, O.A. Redox control of phosphorus pools in Hawaiian montane forest soils. Geoderma 2001, 102, 219–237. [Google Scholar] [CrossRef]
- Sanchez, P.A. Properties and Management of Soils in the Tropics. Soil Sci. 1977, 124, 187. [Google Scholar] [CrossRef]
- Ayaga, G.; Todd, A.; Brookes, P.C. Enhanced biological cycling of phosphorus increases its availability to crops in low-input sub-Saharan farming systems. Soil Boil. Biochem. 2006, 38, 81–90. [Google Scholar] [CrossRef]
- Olander, L.P.; Vitousek, P.M. Biological and geochemical sinks for phosphorus in soil from a wet tropical forest. Ecosystems 2004, 7, 404–419. [Google Scholar] [CrossRef]
- Hall, S.J.; Silver, W.L. Reducing conditions, reactive metals, and their interactions can explain spatial patterns of surface soil carbon in a humid tropical forest. Biogeochemistry 2015, 125, 149–165. [Google Scholar] [CrossRef]
- Silver, W.L.; Lugo, A.E.; Keller, M. Soil oxygen availability and biogeochemistry along rainfall and topographic gradients in upland wet tropical forest soils. Biogeochemistry 1999, 44, 301–328. [Google Scholar] [CrossRef]
- Liptzin, D.; Silver, W.L.; Detto, M. Temporal dynamics in soil oxygen and greenhouse gases in two humid tropical forests. Ecosystems 2011, 14, 171–182. [Google Scholar] [CrossRef]
- Chacon, N.; Silver, W.L.; Dubinsky, E.A.; Cusack, D.F. Iron reduction and soil phosphorus solubilization in humid tropical forests soils: The roles of labile carbon pools and an electron shuttle compound. Biogeochemistry 2006, 78, 67–84. [Google Scholar] [CrossRef]
- Liptzin, D.; Silver, W.L. Effects of carbon additions on iron reduction and phosphorus availability in a humid tropical forest soil. Soil Boil. Biochem. 2009, 41, 1696–1702. [Google Scholar] [CrossRef]
- Peretyazhko, T.; Sposito, G. Iron(III) reduction and phosphorous solubilization in humid tropical forest soils. Geochim. Cosmochim. Acta 2005, 69, 3643–3652. [Google Scholar] [CrossRef]
- Lin, Y.; Bhattacharyya, A.; Campbell, A.N.; Nico, P.S.; Pett-Ridge, J.; Silver, W.L. Phosphorus fractionation responds to dynamic redox conditions in a humid tropical forest soil. J. Geophys. Res. Biogeosci. 2018, 123, 3016–3027. [Google Scholar] [CrossRef]
- Parfitt, R.L.; Atkinson, R.J.; Smart, R.S. The mechanism of phosphate fixation by iron oxides. Soil Sci. Soc. Am. J. 1975, 39, 837–841. [Google Scholar] [CrossRef]
- McGroddy, M.; Silver, W.L. Variations in belowground carbon storage and soil CO2 flux rates along a wet tropical climate gradient. Biotropica 2000, 32, 614–624. [Google Scholar] [CrossRef]
- Menezes-Blackburn, D.; Giles, C.; Darch, T.; George, T.S.; Blackwell, M.; Stutter, M.; Shand, C.; Lumsdon, D.; Cooper, P.; Wendler, R.; et al. Opportunities for mobilizing recalcitrant phosphorus from agricultural soils: A review. Plant Soil 2018, 427, 5–16. [Google Scholar] [CrossRef]
- Liptzin, D.; Silver, W.L. Spatial patterns in oxygen and redox sensitive biogeochemistry in tropical forest soils. Ecosphere 2015, 6, 1–14. [Google Scholar] [CrossRef]
- Harris, N.L.; Lugo, A.E.; Brown, S.; Heartsill-Scalley, T. Luquillo experimental forest: Research history and opportunities. In Experimental Forest and Range EFR-1; USDA Forest Service: Washington, DC, USA, 2012. [Google Scholar]
- Wadsworth, F.H. Ordenación forestal en las montañas de LuquilloForest management in the Luquillo mountains. Caribb. For. 1951, 12, 115–132. [Google Scholar]
- O’Connell, C.S.; Ruan, L.; Silver, W.L. Drought drives rapid shifts in tropical rainforest soil biogeochemistry and greenhouse gas emissions. Nat. Commun. 2018, 9, 1348. [Google Scholar] [CrossRef] [PubMed]
- Hall, S.J.; Silver, W.L. Iron oxidation stimulates organic matter decomposition in humid tropical forest soils. Glob. Chang. Boil. 2013, 19, 2804–2813. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharyya, A.; Campbell, A.N.; Tfaily, M.M.; Lin, Y.; Silver, W.L.; Nico, P.S.; Pett-Ridge, J. Redox fluctuations control the coupled cycling of iron and carbon in tropical forest soils. BioRxiv 2018. [Google Scholar] [CrossRef] [PubMed]
- Pett-Ridge, J.; Firestone, M.K. Redox fluctuation structures microbial communities in a wet tropical soil. Appl. Environ. Microbiol. 2005, 71, 6998–7007. [Google Scholar] [CrossRef] [PubMed]
- Pett-Ridge, J.; Petersen, D.G.; Nuccio, E.; Firestone, M.K. Influence of oxic/anoxic fluctuations on ammonia oxidizers and nitrification potential in a wet tropical soil. FEMS Microbiol. Ecol. 2013, 85, 179–194. [Google Scholar] [CrossRef] [PubMed]
- Pett-Ridge, J.; Silver, W.L.; Firestone, M.K. Redox Fluctuations Frame Microbial Community Impacts on N-cycling Rates in a Humid Tropical Forest Soil. Biogeochemistry 2006, 81, 95–110. [Google Scholar] [CrossRef]
- Hall, S.J.; Silver, W.L.; Timokhin, V.I.; Hammel, K.E. Lignin decomposition is sustained under fluctuating redox conditions in humid tropical forest soils. Glob. Chang. Boil. 2015, 21, 2818–2828. [Google Scholar] [CrossRef] [PubMed]
- Fredrickson, J.K.; Zachara, J.M.; Kennedy, D.W.; Dong, H.; Onstott, T.C.; Hinman, N.W.; Li, S.M. Biogenic iron mineralization accompanying the dissimilatory reduction of hydrous ferric oxide by a groundwater bacterium. Geochim. Cosmochim. Acta 1998, 62, 3239–3257. [Google Scholar] [CrossRef]
- Viollier, E.; Inglett, P.W.; Hunter, K.; Roychoudhury, A.N.; Van Cappellen, P. The ferrozine method revisited: Fe (II)/Fe (III) determination in natural waters. Appl. Geochem. 2000, 15, 785–790. [Google Scholar] [CrossRef]
- Cooperband, L.R.; Gale, P.M.; Comerford, N.B. Refinement of the anion exchange membrane method for soluble phosphorus measurement. Soil Sci. Soc. Am. J. 1999, 63, 58–64. [Google Scholar] [CrossRef]
- Qian, P.; Schoenau, J.J. Practical applications of ion exchange resins in agricultural and environmental soil research. Can. J. Soil Sci. 2002, 82, 9–21. [Google Scholar] [CrossRef]
- Gross, A.; Turner, B.L.; Wright, S.J.; Tanner, E.V.; Reichstein, M.; Weiner, T.; Angert, A. Oxygen isotope ratios of plant available phosphate in lowland tropical forest soils. Soil Boil. Biochem. 2015, 88, 354–361. [Google Scholar] [CrossRef]
- Turner, B.L.; Engelbrecht, B.M.J. Soil organic phosphorus in lowland tropical rain forests. Biogeochemistry 2010, 103, 297–315. [Google Scholar] [CrossRef]
- Tiessen, H.; Moir, J.O. Characterization of available P by sequential extraction. In Soil Sampling and Methods of Analsis; CRC Press: Baca Ratoon, FL, USA, 1993; pp. 75–86. [Google Scholar]
- Kouno, K.; Tuchiya, Y.; Ando, T. Measurement of soil microbial biomass phosphorus by an anion-exchange membrane method. Soil Boil. Biochem. 1995, 27, 1353–1357. [Google Scholar] [CrossRef]
- Brookes, P.; Powlson, D.S.; Jenkinson, D.S. Measurement of microbial biomass phosphorus in soil. Soil Boil. Biochem. 1982, 14, 319–329. [Google Scholar] [CrossRef]
- Bilyera, N.; Blagodatskaya, E.; Yevdokimov, I.; Kuzyakov, Y. Towards a conversion factor for soil microbial phosphorus. Eur. J. Soil Biol. 2018, 87, 1–8. [Google Scholar] [CrossRef]
- Murphy, J.; Riley, J.P. A modified single solution method for the determination of phosphate in natural waters. Anal. Chim. Acta 1962, 27, 31–36. [Google Scholar] [CrossRef]
- Henry, P.; Smith, M.F. Two-step approach to determining some useful phosphorus characteristics of South African soils: A review of work done at the ARC-Institute for Soil, Climate and Water. South Afr. J. Plant Soil 2006, 23, 64–72. [Google Scholar] [CrossRef]
- Weber, K.A.; Achenbach, L.A.; Coates, J.D. Coates, Microorganisms pumping iron: Anaerobic microbial iron oxidation and reduction. Nat. Rev. Microbiol. 2006, 4, 752. [Google Scholar] [CrossRef] [PubMed]
- Thompson, A.; Rancourt, D.G.; Chadwick, O.A.; Chorover, J. Iron solid-phase differentiation along a redox gradient in basaltic soils. Geochim. Cosmochim. Acta 2011, 75, 119–133. [Google Scholar] [CrossRef]
- Keiluweit, M.; Gee, K.; Denney, A.; Fendorf, S. Anoxic microsites in upland soils dominantly controlled by clay content. Soil Boil. Biochem. 2018, 118, 42–50. [Google Scholar] [CrossRef]
- Quantin, C.; Becquer, T.; Rouiller, J.H.; Berthelin, J. Redistribution of metals in a New Caledonia Ferralsol after microbial weathering. Soil Sci. Soc. Am. J. 2002, 66, 1797–1804. [Google Scholar] [CrossRef]
- Hall, S.J.; Treffkorn, J.; Silver, W.L. Breaking the enzymatic latch: Impacts of reducing conditions on hydrolytic enzyme activity in tropical forest soils. Ecology 2014, 95, 2964–2973. [Google Scholar] [CrossRef]
- Turner, B.L.; Lambers, H.; Condron, L.M.; Cramer, M.D.; Leake, J.R.; Richardson, A.E.; Smith, S.E. Soil microbial biomass and the fate of phosphorus during long-term ecosystem development. Plant Soil 2013, 367, 225–234. [Google Scholar] [CrossRef]
- Turner, B.L.; Wright, J.S. The response of microbial biomass and hydrolytic enzymes to a decade of nitrogen, phosphorus, and potassium addition in a lowland tropical rain forest. Biogeochemistry 2014, 117, 115–130. [Google Scholar] [CrossRef]
- Wood, T.E.; Matthews, D.; Vandecar, K.; Lawrence, D. Short-term variability in labile soil phosphorus is positively related to soil moisture in a humid tropical forest in Puerto Rico. Biogeochemistry 2016, 127, 35–43. [Google Scholar] [CrossRef]



© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gross, A.; Pett-Ridge, J.; Silver, W.L. Soil Oxygen Limits Microbial Phosphorus Utilization in Humid Tropical Forest Soils. Soil Syst. 2018, 2, 65. https://doi.org/10.3390/soilsystems2040065
Gross A, Pett-Ridge J, Silver WL. Soil Oxygen Limits Microbial Phosphorus Utilization in Humid Tropical Forest Soils. Soil Systems. 2018; 2(4):65. https://doi.org/10.3390/soilsystems2040065
Chicago/Turabian StyleGross, Avner, Jennifer Pett-Ridge, and Whendee L. Silver. 2018. "Soil Oxygen Limits Microbial Phosphorus Utilization in Humid Tropical Forest Soils" Soil Systems 2, no. 4: 65. https://doi.org/10.3390/soilsystems2040065
APA StyleGross, A., Pett-Ridge, J., & Silver, W. L. (2018). Soil Oxygen Limits Microbial Phosphorus Utilization in Humid Tropical Forest Soils. Soil Systems, 2(4), 65. https://doi.org/10.3390/soilsystems2040065