Interacting Controls of Pyrolysis Temperature and Plant Taxa on the Degradability of PyOM in Fire-Prone Northern Temperate Forest Soil

 , , , ,
, , , ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Physcial and Chemical Characterization of 13C-Enriched PyOM and Source Wood
2.2. Soil Sampling and Laboratory Incubation Experiments
2.3. Stable Isotope Measurements
2.4. Curve Fitting, Correlative and Statistical Analyses
3. Results
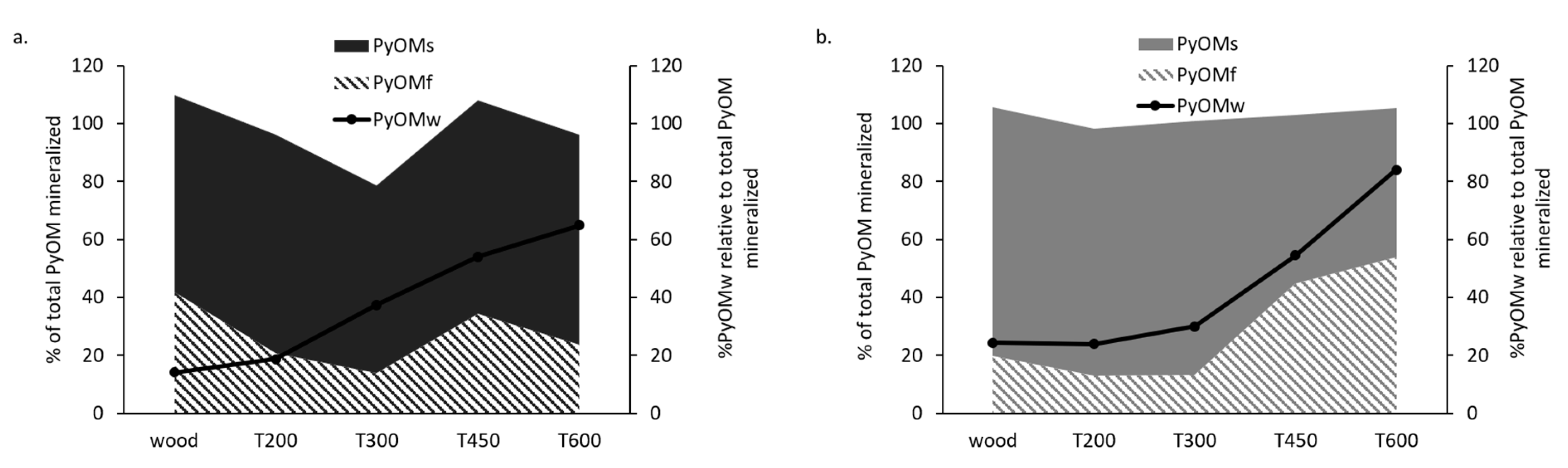
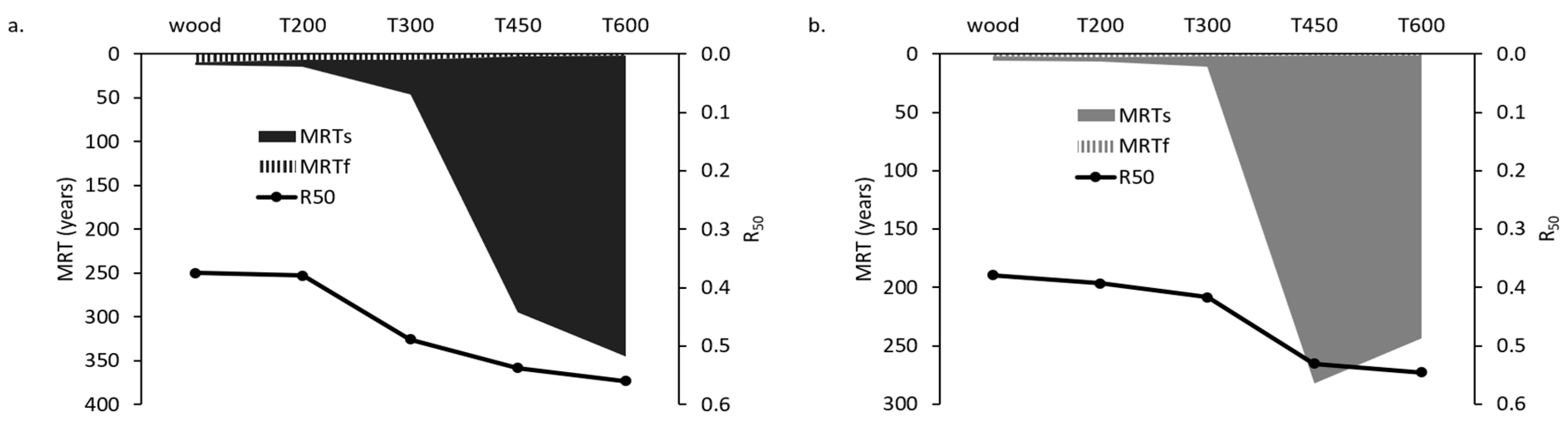
3.1. PyOM Production Temperature Controls Mineralization
3.2. Linking Physiochemical Structure and PyOM Mineralization
3.3. Influence of Source Taxa on PyOM Mineralization
3.4. Influence of Sucrose on PyOM Mineralization
4. Discussion
4.1. Changes in Chemical Composition with Increased Pyrolysis Temperature Controls PyOM Mineralization Rate and MRT
4.2. Plant Source Taxa Mediates MRT and Mineralization at Intermediate PyOM Pyrolysis Temperatures
4.3. PyOMw Content and Estimated PyOM Recalcitrance Correlate with PyOM Decomposition
4.4. Labile C Addition has a Minor Effect on PyOM Decomposition
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- IPCC Climate Change. The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University: Cambridge, UK; New York, NY, USA, 2013. [Google Scholar]
- Santín, C.; Doerr, S.H.; Kane, E.S.; Masiello, C.A.; Ohlson, M.; Rosa, J.M.; Preston, C.M.; Dittmar, T. Towards a global assessment of pyrogenic carbon from vegetation fires. Glob. Chang. Biol. 2016, 22, 76–91. [Google Scholar] [CrossRef] [PubMed]
- Santín, C.; Doerr, S.H.; Preston, C.M.; González-Rodríguez, G. Pyrogenic organic matter production from wildfires: A missing sink in the global carbon cycle. Glob. Chang. Biol. 2015, 21, 1621–1633. [Google Scholar] [CrossRef] [PubMed]
- Santos, F.; Fraser, M.P.; Bird, J.A. Atmospheric black carbon deposition and characterization of biomass burning tracers in a northern temperate forest. Atmos. Environ. 2014, 95, 383–390. [Google Scholar] [CrossRef]
- Hatton, P.-J.; Chatterjee, S.; Filley, T.R.; Dastmalchi, K.; Plante, A.F.; Abiven, S.; Gao, X.; Masiello, C.A.; Leavitt, S.W.; Nadelhoffer, K.J.; et al. Tree taxa and pyrolysis temperature interact to control the efficacy of pyrogenic organic matter formation. Biogeochemistry 2016, 130, 103–116. [Google Scholar] [CrossRef]
- Reich, P.B.; Sendall, K.M.; Rice, K.; Rich, R.L.; Stefanski, A.; Hobbie, S.E.; Montgomery, R.A. Geographic range predicts photosynthetic and growth response to warming in co-occurring tree species. Nat. Clim. Chang. 2015, 5, 148–152. [Google Scholar] [CrossRef]
- Lehmann, J.; Abiven, S.; Kleber, M.; Pan, G.; Singh, B.P.; Sohi, S.P.; Zimmerman, A.R.; Lehmann, J.; Joseph, S. Persistence of biochar in soil. Biochar Environ. Manag. Sci. Technol. Implement. 2015, 2, 233–280. [Google Scholar]
- Wang, J.; Xiong, Z.; Kuzyakov, Y. Biochar stability in soil: Meta-analysis of decomposition and priming effects. Gcb Bioenergy 2016, 8, 512–523. [Google Scholar] [CrossRef]
- Weng, Z.; Van Zwieten, L.; Singh, B.P.; Kimber, S.; Morris, S.; Cowie, A.; Macdonald, L.M. Plant-biochar interactions drive the negative priming of soil organic carbon in an annual ryegrass field system. Soil Biol. Biochem. 2015, 90, 111–121. [Google Scholar] [CrossRef]
- Schmidt, M.W.; Torn, M.S.; Abiven, S.; Dittmar, T.; Guggenberger, G.; Janssens, I.A.; Kleber, M.; Kögel-Knabner, I.; Lehmann, J.; Manning, D.A.; et al. Persistence of soil organic matter as an ecosystem property. Nature 2011, 478, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.P.; Cowie, A.L.; Smernik, R.J. Biochar carbon stability in a clayey soil as a function of feedstock and pyrolysis temperature. Environ. Sci. Technol. 2012, 46, 11770–11778. [Google Scholar] [CrossRef] [PubMed]
- Hammes, K.; Torn, M.S.; Lapenas, A.G.; Schmidt, M.W. Centennial black carbon turnover observed in a Russian steppe soil. Biogeosciences 2008, 5, 1339–1350. [Google Scholar] [CrossRef]
- Keiluweit, M.; Nico, P.S.; Johnson, M.G.; Kleber, M. Dynamic molecular structure of plant biomass-derived black carbon (biochar). Environ. Sci. Technol. 2010, 44, 1247–1253. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, S.; Santos, F.; Abiven, S.; Itin, B.; Stark, R.E.; Bird, J.A. Elucidating the chemical structure of pyrogenic organic matter by combining magnetic resonance, mid-infrared spectroscopy and mass spectrometry. Org. Geochem. 2012, 51, 35–44. [Google Scholar] [CrossRef]
- Brodowski, S.; John, B.; Flessa, H.; Amelung, W. Aggregate-occluded black carbon in soil. Eur. J. Soil Sci. 2006, 57, 539–546. [Google Scholar] [CrossRef]
- Liang, B.; Lehmann, J.; Solomon, D.; Sohi, S.; Thies, J.E.; Skjemstad, J.O.; Luizao, F.J.; Engelhard, M.H.; Neves, E.G.; Wirick, S. Stability of biomass-derived black carbon in soils. Geochim. Cosmochim. Acta 2008, 72, 6069–6078. [Google Scholar] [CrossRef]
- Farrell, M.; Kuhn, T.K.; Macdonald, L.M.; Maddern, T.M.; Murphy, D.V.; Hall, P.A.; Singh, B.P.; Baumann, K.; Krull, E.S.; Baldock, J.A. Microbial utilisation of biochar-derived carbon. Sci. Total Environ. 2013, 465, 288–297. [Google Scholar] [CrossRef] [PubMed]
- Santos, F.; Torn, M.S.; Bird, J.A. Biological degradation of pyrogenic organic matter in temperate forest soils. Soil Biol. Biochem. 2012, 51, 115–124. [Google Scholar] [CrossRef]
- Wang, X.; Song, D.; Liang, G.; Zhang, Q.; Ai, C.; Zhou, W. Maize biochar addition rate influences soil enzyme activity and microbial community composition in a fluvo-aquic soil. Appl. Soil Ecol. 2015, 96, 265–272. [Google Scholar] [CrossRef]
- Cheng, C.-H.; Lehmann, J.; Thies, J.E.; Burton, S.D.; Engelhard, M.H. Oxidation of black carbon by biotic and abiotic processes. Org. Geochem. 2006, 37, 1477–1488. [Google Scholar] [CrossRef]
- Liu, Z.; Demisie, W.; Zhang, M. Simulated degradation of biochar and its potential environmental implications. Environ. Pollut. 2013, 179, 146–152. [Google Scholar] [CrossRef] [PubMed]
- Gibson, C.; Berry, T.D.; Wang, R.; Spencer, J.A.; Johnston, C.T.; Jiang, Y.; Bird, J.A.; Filley, T.R. Weathering of pyrogenic organic matter induces fungal oxidative enzyme response in single culture inoculation experiments. Org. Geochem. 2016, 92, 32–41. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Gibson, C.D.; Berry, T.D.; Jiang, Y.; Bird, J.A.; Filley, T.R. Photooxidation of pyrogenic organic matter reduces its reactive, labile C pool and the apparent soil oxidative microbial enzyme response. Geoderma 2017, 293, 10–18. [Google Scholar] [CrossRef]
- Du, Z.; Wang, Y.; Huang, J.; Lu, N.; Liu, X.; Lou, Y.; Zhang, Q. Consecutive biochar application alters soil enzyme activities in the winter wheat–growing season. Soil Sci. 2014, 179, 75–83. [Google Scholar] [CrossRef]
- Gibson, C.; Hatton, P.-J.; Bird, J.A.; Nadelhoffer, K.; Le Moine, J.; Filley, T. Tree taxa and pyrolysis temperature interact to control pyrogenic organic matter induced native soil organic carbon priming. Soil Biol. Biochem. 2018, 119, 174–183. [Google Scholar] [CrossRef]
- Bruun, E.W.; Hauggaard-Nielsen, H.; Ibrahim, N.; Egsgaard, H.; Ambus, P.; Jensen, P.A.; Dam-Johansen, K. Influence of fast pyrolysis temperature on biochar labile fraction and short-term carbon loss in a loamy soil. Biomass Bioenergy 2011, 35, 1182–1189. [Google Scholar] [CrossRef]
- Cross, A.; Sohi, S.P. The priming potential of biochar products in relation to labile carbon contents and soil organic matter status. Soil Biol. Biochem. 2011, 43, 2127–2134. [Google Scholar] [CrossRef]
- Pereira, R.C.; Kaal, J.; Arbestain, M.C.; Lorenzo, R.P.; Aitkenhead, W.; Hedley, M.; Macías, F.; Hindmarsh, J.; Maciá-Agulló, J.A. Contribution to characterisation of biochar to estimate the labile fraction of carbon. Org. Geochem. 2011, 42, 1331–1342. [Google Scholar] [CrossRef]
- Hamer, U.; Marschner, B.; Brodowski, S.; Amelung, W. Interactive priming of black carbon and glucose mineralisation. Org. Geochem. 2004, 35, 823–830. [Google Scholar] [CrossRef]
- Kuzyakov, Y.; Bogomolova, I.; Glaser, B. Biochar stability in soil: Decomposition during eight years and transformation as assessed by compound-specific 14 C analysis. Soil Biol. Biochem. 2014, 70, 229–236. [Google Scholar] [CrossRef]
- Wang, J.; Dokohely, M.E.; Xiong, Z.; Kuzyakov, Y. Contrasting effects of aged and fresh biochars on glucose-induced priming and microbial activities in paddy soil. J. Soils Sediments 2016, 16, 191–203. [Google Scholar] [CrossRef]
- Ryan, K.C. Dynamic interactions between forest structure and fire behavior in boreal ecosystems. Silva Fenn. 2002, 36, 13–39. [Google Scholar] [CrossRef]
- Santín, C.; Doerr, S.H.; Merino, A.; Bryant, R.; Loader, N.J. Forest floor chemical transformations in a boreal forest fire and their correlations with temperature and heating duration. Geoderma 2016, 264, 71–80. [Google Scholar] [CrossRef]
- Kleber, M.; Hockaday, W.; Nico, P.S. Characteristics of biochar: Macro-molecular properties. In Biochar for Environmental Management; Routledge: London, UK, 2015. [Google Scholar]
- Cong, Z.; Kang, S.; Gao, S.; Zhang, Y.; Li, Q.; Kawamura, K. Historical Trends of Atmospheric Black Carbon on Tibetan Plateau As Reconstructed from a 150-Year Lake Sediment Record. Environ. Sci. Technol. 2013, 47, 2579–2586. [Google Scholar] [CrossRef] [PubMed]
- Socher, S.A.; Prati, D.; Boch, S.; Müller, J.; Baumbach, H.; Gockel, S.; Hemp, A.; Schöning, I.; Wells, K.; Buscot, F. Others Interacting effects of fertilization, mowing and grazing on plant species diversity of 1500 grasslands in Germany differ between regions. Basic Appl. Ecol. 2013, 14, 126–136. [Google Scholar] [CrossRef]
- Cumming, S.G.; Drever, C.R.; Houle, M.; Cosco, J.; Racine, P.; Bayne, E.; Schmiegelow, F.K. A gap analysis of tree species representation in the protected areas of the Canadian boreal forest: Applying a new assemblage of digital Forest Resource Inventory data 1. Can. J. For. Res. 2014, 45, 163–173. [Google Scholar] [CrossRef]
- Sun, Y.; Gao, B.; Yao, Y.; Fang, J.; Zhang, M.; Zhou, Y.; Chen, H.; Yang, L. Effects of feedstock type, production method, and pyrolysis temperature on biochar and hydrochar properties. Chem. Eng. J. 2014, 240, 574–578. [Google Scholar] [CrossRef]
- Wang, S.; Gao, B.; Zimmerman, A.R.; Li, Y.; Ma, L.; Harris, W.G.; Migliaccio, K.W. Physicochemical and sorptive properties of biochars derived from woody and herbaceous biomass. Chemosphere 2015, 134, 257–262. [Google Scholar] [CrossRef] [PubMed]
- Shortle, W.C.; Smith, K.T.; Jellison, J.; Schilling, J.S. Potential of decaying wood to restore root-available base cations in depleted forest soils. Can. J. For. Res. 2012, 42, 1015–1024. [Google Scholar] [CrossRef]
- McFarlane, K.J.; Torn, M.S.; Hanson, P.J.; Porras, R.C.; Swanston, C.W.; Callaham, M.A.; Guilderson, T.P. Comparison of soil organic matter dynamics at five temperate deciduous forests with physical fractionation and radiocarbon measurements. Biogeochemistry 2012, 112, 457–476. [Google Scholar] [CrossRef]
- De Groot, W.J.; Flannigan, M.D.; Cantin, A.S. Climate change impacts on future boreal fire regimes. For. Ecol. Manag. 2013, 294, 35–44. [Google Scholar] [CrossRef]
- Bird, J.A.; Torn, M.S. Fine Roots vs. Needles: A Comparison of 13C and 15N Dynamics in a Ponderosa Pine Forest Soil. Biogeochemistry 2006, 79, 361–382. [Google Scholar] [CrossRef]
- Hammes, K.; Smernik, R.J.; Skjemstad, J.O.; Herzog, A.; Vogt, U.F.; Schmidt, M.W. Synthesis and characterisation of laboratory-charred grass straw (Oryza sativa) and chestnut wood (Castanea sativa) as reference materials for black carbon quantification. Org. Geochem. 2006, 37, 1629–1633. [Google Scholar] [CrossRef]
- Fernández, J.M.; Peltre, C.; Craine, J.M.; Plante, A.F. Improved characterization of soil organic matter by thermal analysis using CO2/H2O evolved gas analysis. Environ. Sci. Technol. 2012, 46, 8921–8927. [Google Scholar] [CrossRef] [PubMed]
- Ward, C.P.; Sleighter, R.L.; Hatcher, P.G.; Cory, R.M. Insights into the complete and partial photooxidation of black carbon in surface waters. Environ. Sci. Process. Impacts 2014, 16, 721–731. [Google Scholar] [CrossRef] [PubMed]
- Weishaar, J.L.; Aiken, G.R.; Bergamaschi, B.A.; Fram, M.S.; Fujii, R.; Mopper, K. Evaluation of specific ultraviolet absorbance as an indicator of the chemical composition and reactivity of dissolved organic carbon. Environ. Sci. Technol. 2003, 37, 4702–4708. [Google Scholar] [CrossRef] [PubMed]
- Gough, C.M.; Vogel, C.S.; Schmid, H.P.; Su, H.-B.; Curtis, P.S. Multi-year convergence of biometric and meteorological estimates of forest carbon storage. Agric. For. Meteorol. 2008, 148, 158–170. [Google Scholar] [CrossRef]
- Gough, C.M.; Vogel, C.S.; Schmid, H.P.; Curtis, P.S. Controls on annual forest carbon storage: Lessons from the past and predictions for the future. Bioscience 2008, 58, 609–622. [Google Scholar] [CrossRef]
- Žifčáková, L.; Větrovský, T.; Howe, A.; Baldrian, P. Microbial activity in forest soil reflects the changes in ecosystem properties between summer and winter. Environ. Microbiol. 2016, 18, 288–301. [Google Scholar] [CrossRef] [PubMed]
- Zimmerman, A.R.; Gao, B.; Ahn, M.-Y. Positive and negative carbon mineralization priming effects among a variety of biochar-amended soils. Soil Biol. Biochem. 2011, 43, 1169–1179. [Google Scholar] [CrossRef]
- Santos, F.; Nadelhoffer, K.; Bird, J.A. Rapid fine root C and N mineralization in a northern temperate forest soil. Biogeochemistry 2016, 128, 187–200. [Google Scholar] [CrossRef]
- Zimmerman, A.R. Abiotic and microbial oxidation of laboratory-produced black carbon (biochar). Environ. Sci. Technol. 2010, 44, 1295–1301. [Google Scholar] [CrossRef] [PubMed]
- Ippolito, J.A.; Spokas, K.A.; Novak, J.M.; Lentz, R.D.; Cantrell, K.B. Biochar elemental composition and factors influencing nutrient retention. In Biochar for Environmental Management: Science, Technology and Implementation, 2nd ed.; Routledge: New York, NY, USA, 2015; pp. 139–163. [Google Scholar]
- Pignatello, J.J.; Uchimiya, M.; Abiven, S.; Schmidt, M.W. Evolution of biochar properties in soil. Biochar Environ. Manag. 2015, 1, 195–234. [Google Scholar]
- Jiang, X.; Haddix, M.L.; Cotrufo, M.F. Interactions between biochar and soil organic carbon decomposition: Effects of nitrogen and low molecular weight carbon compound addition. Soil Biol. Biochem. 2016, 100, 92–101. [Google Scholar] [CrossRef]
- Hopkins, F.M.; Filley, T.R.; Gleixner, G.; Lange, M.; Top, S.M.; Trumbore, S.E. Increased belowground carbon inputs and warming promote loss of soil organic carbon through complementary microbial responses. Soil Biol. Biochem. 2014, 76, 57–69. [Google Scholar] [CrossRef]
- Cotrufo, M.F.; Soong, J.L.; Horton, A.J.; Campbell, E.E.; Haddix, M.L.; Wall, D.H.; Parton, W.J. Formation of soil organic matter via biochemical and physical pathways of litter mass loss. Nat. Geosci. 2015, 8, 776–779. [Google Scholar] [CrossRef]
- Whitman, T.; Zhu, Z.; Lehmann, J. Carbon Mineralizability Determines Interactive Effects on Mineralization of Pyrogenic Organic Matter and Soil Organic Carbon. Environ. Sci. Technol. 2014, 48, 13727–13734. [Google Scholar] [CrossRef] [PubMed]
- Whitman, T.; Singh, B.P.; Zimmerman, A.R.; Lehmann, J.; Joseph, S. Priming effects in biochar-amended soils: Implications of biochar-soil organic matter interactions for carbon storage. Biochar Environ. Manag. Sci. Technol. Implement. 2015, 2, 455–488. [Google Scholar]
- Skjemstad, J.O.; Clarke, P.; Taylor, J.A.; Oades, J.M.; McClure, S.G. The chemistry and nature of protected carbon in soil. Soil Res. 1996, 34, 251–271. [Google Scholar] [CrossRef]
- Kawamoto, K.; Ishimaru, K.; Imamura, Y. Reactivity of wood charcoal with ozone. J. Wood Sci. 2005, 51, 6672. [Google Scholar] [CrossRef]
- Plante, A.F.; Fernández, J.M.; Haddix, M.L.; Steinweg, J.M.; Conant, R.T. Biological, chemical and thermal indices of soil organic matter stability in four grassland soils. Soil Biol. Biochem. 2011, 43, 1051–1058. [Google Scholar] [CrossRef]
- Harvey, O.R.; Kuo, L.-J.; Zimmerman, A.R.; Louchouarn, P.; Amonette, J.E.; Herbert, B.E. An index-based approach to assessing recalcitrance and soil carbon sequestration potential of engineered black carbons (biochars). Environ. Sci. Technol. 2012, 46, 1415–1421. [Google Scholar] [CrossRef] [PubMed]
- Goodell, B.; Jellison, J.; Liu, J.; Daniel, G.; Paszczynski, A.; Fekete, F.; Krishnamurthy, S.; Jun, L.; Xu, G. Low molecular weight chelators and phenolic compounds isolated from wood decay fungi and their role in the fungal biodegradation of wood. J. Biotechnol. 1997, 53, 133–162. [Google Scholar] [CrossRef]
- Zapanta, L.S.; Tien, M. The roles of veratryl alcohol and oxalate in fungal lignin degradation. J. Biotechnol. 1997, 53, 93–102. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Treatment | C (g kg−1) | N (g kg−1) | 13C (atom %) | 15N (atom %) | Porosity (%) | BET-N2 Surface Area (m2 g−1) | % Long-Chain Aliphatics | % Non-Lignin Phenols | % Carbohydrates | Recalcitrance Index (R50) | Specific UV Absorption (254 nm) | % Leachable PyOM of Initial PyOM or Wood (PyOMw) | Energy Density (J mg−1 C) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| JP wood | 464 | 2.8 | 2.2 | 19.2 | 53 | 2.6 | 10.7 | 21.2 | 58.7 | 0.4 | 1.1 <0.01 | 7.4 | 21.1 |
| JP T200 | 507 | 3.4 | 2.4 | 18.8 | 60 | 2.8 | 10.6 | 18.3 | 67.6 | 0.4 | 1.7 <0.01 | 4.2 | 20.2 |
| JP T300 | 669 | 5.4 | 2.4 | 18.8 | 75 | 4.3 | 22.1 | 5.9 | 15.8 | 0.5 | 1.7 <0.01 | 1.4 | 26.4 |
| JP T450 | 786 | 5.4 | 2.3 | 18.9 | - | 13 | 7.7 | 0.09 | 6.8 | 0.5 | 3.2 0.10 | 0.3 | 27.2 |
| JP T600 | 831 | 5.3 | 2.3 | 18.8 | - | 212 | 0.0 | 3.3 | 0.0 | 0.6 | 3.7 0.10 | 0.4 | 26.9 |
| RM wood | 451 | 6.4 | 3.8 | 8.9 | 60 | 0.5 | 13.7 | 63.1 | 65.1 | 0.4 | 2.6 0.10 | 11.8 | 20.0 |
| RM T200 | 495 | 6.9 | 3.8 | 8.9 | 61 | 1.4 | 12.3 | 36.9 | 65.1 | 0.4 | 3.1 <0.01 | 8.1 | 22.2 |
| RM T300 | 574 | 9.0 | 3.7 | 8.9 | 72 | 2.3 | 19.4 | 14.3 | 41.4 | 0.4 | 2.4 <0.01 | 4.4 | 23.4 |
| RM T450 | 761 | 9.4 | 3.9 | 9.1 | 78 | 3.3 | 5.7 | 0.62 | 4.5 | 0.5 | 3.0 0.10 | 0.4 | 30.5 |
| RM T600 | 765 | 9.5 | 3.9 | 9.2 | 81 | 132 | 0.0 | 0.01 | 0.0 | 0.5 | 4.0 0.10 | 0.4 | 29.6 |
| Treatment | Total PyOM-C Mineralized (mg C g −1 Added C) | Fast-Cycling C Pool (mg C g −1 Added C) | Slow-Cycling C Pool (mg C g −1 Added C) | MRTf (days) | MRTs (years) | PyOMw (mg C) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| JP wood | 560.66 | 75.4 | a | 214.1 | 4.8 | a | 403.9 | 31.7 | a | 10.0 | 1.1 | a | 2.7 | 0.9 | a | 4.1 | 0.3 | b |
| JP T200 | 302.1 | 65.9 | b | 45.5 | 1.1 | c * | 249.0 | 7.4 | c * | 6.1 | 1.3 | b | 8.0 | 4.3 | b * | 2.3 | 0.2 | d |
| JP T300 | 41.3 | 6.0 | d | 6.3 | 0.8 | e * | 29.2 | 1.2 | e * | 6.6 | 1.2 | b | 40.0 | 4.2 | c | 0.8 | 0.1 | f |
| JP T450 | 6.6 | 0.5 | e | 2.3 | 1.8 | f | 5.1 | 0.8 | f | 2.2 | 0.4 | c | 292.3 | 45.6 | d | 0.2 | 0.0 | g |
| JP T600 | 6.6 | 1.7 | e | 1.4 | 0.0 | f | 5.0 | 1.0 | f | 1.7 | 1.0 | c | 343.3 | 72.1 | d | 0.2 | 0.0 | g |
| RM wood | 488.3 | 20.7 | a | 99.4 | 4.9 | b * | 432.3 | 22.0 | a * | 2.4 | 0.3 | c | 3.4 | 0.7 | a | 6.5 | 0.4 | a |
| RM T200 | 351.2 | 40.4 | b | 46.1 | 1.2 | c * | 309.6 | 23.0 | b * | 2.5 | 0.5 | c | 4.2 | 0.9 | a | 4.5 | 0.3 | c |
| RM T300 | 150.8 | 16.5 | c | 19.7 | 0.5 | d | 136.6 | 9.1 | d * | 1.8 | 0.2 | c | 8.9 | 1.1 | b | 2.4 | 0.2 | e |
| RM T450 | 7.4 | 0.9 | e | 3.3 | 0.2 | f | 4.5 | 0.4 | f | 1.2 | 0.3 | c | 281.1 | 54.2 | d * | 0.2 | 0.0 | g |
| RM T600 | 6.0 | 1.4 | e | 2.5 | 0.1 | f | 3.9 | 0.2 | f | 1.3 | 0.0 | c | 242.4 | 21.3 | e * | 0.2 | 0.0 | g |
| JPS wood | 686.2 | 58.5 | a | 174.0 | 10.1 | a | 533.8 | 44.6 | a | 9.9 | 1.1 | a | 1.6 | 0.3 | a | 4.1 | 0.3 | b |
| JPS T200 | 345.4 | 25.4 | b | 62.9 | 0.2 | c * | 293.3 | 8.2 | b * | 6.1 | 1.3 | b | 4.1 | 0.6 | a * | 2.3 | 0.2 | d |
| JPS T300 | 31.0 | 1.9 | d | 13.4 | 1.5 | e * | 18.5 | 1.8 | d * | 8.0 | 2.0 | b | 64.0 | 7.7 | c | 0.8 | 0.1 | f |
| JPS T450 | 7.0 | 0.6 | e | 1.7 | 0.2 | f | 5.5 | 0.4 | e | 2.2 | 0.4 | c | 246.0 | 29.8 | d | 0.2 | 0.0 | g |
| JPS T600 | 6.3 | 2.0 | e | 1.8 | 1.0 | f | 4.7 | 0.2 | e | 2.3 | 0.8 | c | 333.4 | 73.2 | d | 0.2 | 0.0 | g |
| RMS wood | 611.5 | 29.8 | a | 71.3 | 2.6 | b * | 559.3 | 30.2 | a * | 2.3 | 0.3 | c | 2.0 | 0.6 | a | 6.5 | 0.4 | a |
| RMS T200 | 328.5 | 24.8 | b | 52.7 | 1.6 | c * | 286.2 | 6.6 | b * | 2.4 | 0.5 | c | 3.9 | 0.5 | a | 4.5 | 0.3 | c |
| RMS T300 | 102.2 | 11.6 | c | 18.9 | 1.0 | d | 86.5 | 5.3 | d * | 2.0 | 0.2 | c | 12.7 | 2.0 | b | 2.4 | 0.2 | e |
| RMS T450 | 6.2 | 0.3 | e | 3.1 | 0.1 | f | 3.3 | 0.4 | e | 1.6 | 0.2 | c | 437.3 | 27.0 | e * | 0.2 | 0.0 | g |
| RMS T600 | 6.5 | 1.0 | e | 2.4 | 0.1 | f | 4.3 | 0.6 | e | 1.4 | 0.0 | c | 332.9 | 50.9 | e * | 0.2 | 0.0 | g |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gibson, C.D.; Hatton, P.-J.; Bird, J.A.; Nadelhoffer, K.; Ward, C.P.; Stark, R.E.; Filley, T.R. Interacting Controls of Pyrolysis Temperature and Plant Taxa on the Degradability of PyOM in Fire-Prone Northern Temperate Forest Soil. Soil Syst. 2018, 2, 48. https://doi.org/10.3390/soilsystems2030048
Gibson CD, Hatton P-J, Bird JA, Nadelhoffer K, Ward CP, Stark RE, Filley TR. Interacting Controls of Pyrolysis Temperature and Plant Taxa on the Degradability of PyOM in Fire-Prone Northern Temperate Forest Soil. Soil Systems. 2018; 2(3):48. https://doi.org/10.3390/soilsystems2030048
Chicago/Turabian StyleGibson, Christy D., Pierre-Joseph Hatton, Jeffrey A. Bird, Knute Nadelhoffer, Collin P. Ward, Ruth E. Stark, and Timothy R. Filley. 2018. "Interacting Controls of Pyrolysis Temperature and Plant Taxa on the Degradability of PyOM in Fire-Prone Northern Temperate Forest Soil" Soil Systems 2, no. 3: 48. https://doi.org/10.3390/soilsystems2030048
APA StyleGibson, C. D., Hatton, P.-J., Bird, J. A., Nadelhoffer, K., Ward, C. P., Stark, R. E., & Filley, T. R. (2018). Interacting Controls of Pyrolysis Temperature and Plant Taxa on the Degradability of PyOM in Fire-Prone Northern Temperate Forest Soil. Soil Systems, 2(3), 48. https://doi.org/10.3390/soilsystems2030048