Abstract
Background/Objectives: Parkinson’s disease (PD) is the second-most prevalent neurodegenerative disorder, characterized by the progressive loss of dopaminergic neurons in the Substantia Nigra pars compacta (SNpc). Experimental models that replicate core features of PD are critical to investigate underlying mechanisms and therapeutic strategies. Here we evaluated the effects of an acute unilateral intrastriatal lesion induced by 6-hydroxydopamine (6-OHDA) on neuronal loss and the associated inflammatory response. Methods: Adult male Wistar rats received an injection of 6-OHDA into the right striatum, while the contralateral side received vehicle. Motor behavior was assessed by cylinder and open field tests on post-lesion days (PLDs) 7 and 14. Brains were analyzed by immunohistochemistry for tyrosine hydroxylase (TH), glial response (GFAP and Iba1), and caspase-3 at PLD +14. Results: A marked reduction in TH-immunoreactivity in the lesioned striatum was observed, with ~40% loss of TH-positive neurons in the ipsilateral SNpc. Surviving neurons displayed a 28% increase in soma size compared to the contralateral side. The lesion was accompanied by robust astrocytic and microglial activation at the injection site, as well as enhanced GFAP immunoreactivity in the ipsilateral SN pars reticulata. Apoptotic profiles emerged in the SNpc at PLD +14. Functionally, these alterations were reflected in significant motor asymmetry and decreased locomotor activity. Conclusions: Our findings demonstrate that neuroinflammation accompanies early dopaminergic degeneration following intrastriatal 6-OHDA administration, contributing to motor deficits. Future studies with older animals and broader behavioral and anatomical assessments—including regions such as the ventral tegmental area and motivational or anxiety-related paradigms—may enhance translational relevance.
1. Introduction
Parkinson’s disease (PD) is a prevalent and progressive neurodegenerative disorder that currently affects approximately 10 million individuals worldwide [1]. It is the second most common neurodegenerative condition after Alzheimer’s disease, with incidence increasing notably with age: about 0.1% in individuals aged 50–59, 0.4% between 60 and 69, 1.1% in the 70–79 group, and nearly 1.9% in those over 80 years old [2,3].
Classically, PD is characterized by the progressive loss of dopaminergic neurons in the Substantia Nigra pars compacta (SNpc) and the resulting depletion of striatal dopaminergic innervation. This neurodegeneration leads to cardinal motor symptoms such as resting tremor, bradykinesia, muscular rigidity, and postural instability [4,5,6]. In addition to these motor features, patients often experience non-motor manifestations including cognitive impairment, mood disorders, and autonomic dysfunction, all of which significantly impair quality of life [7,8].
A growing body of evidence indicates that neuroinflammation plays a pivotal role in the pathophysiology and progression of PD [9]. Activated microglia and reactive astrocytes are consistently observed in affected brain regions, accompanied by elevated levels of pro-inflammatory mediators such as tumor necrosis factor alpha (TNF-α), interleukin-1 beta (IL-1β), inducible nitric oxide synthase (iNOS), and cyclooxygenase-2, as reported in both postmortem PD brains and in vivo models [10,11,12]. These neuroinflammatory responses are believed to be related to the onset and progressive loss of dopaminergic neurons by amplifying oxidative stress and aggravating the neurodegenerative cascade.
Owing to its multifactorial etiology and complex pathophysiology, PD has been extensively studied using experimental models that replicate core features of the disease, which are crucial for elucidating pathogenic mechanisms and evaluating therapeutic strategies. Parkinsonism can be induced via genetic or pharmacological methods, with the latter typically relying on neurotoxins that selectively impair the dopaminergic system [13,14]. Among these, 6-hydroxydopamine (6-OHDA) remains one of the most widely used and well-established neuropharmacological or neurotoxic models. First described by Ungerstedt in 1968 [15], 6-OHDA induces selective degeneration of dopaminergic neurons due to its structural similarity to dopamine. It is taken up by dopaminergic fibers and retrogradely transported to the neurons through the dopamine transporter and generates reactive oxygen species (ROS), including hydrogen peroxide, leading to oxidative stress and subsequent neurotoxicity [16]. This oxidative insult initiates a cascade of neuroinflammatory responses [17,18], which further aggravate neuronal damage via the activation of apoptotic pathways [19].
Typically, unilateral injections of 6-OHDA into the medial forebrain bundle (MFB), Substantia Nigra (SN) or striatum produce a reliable and reproducible lesion that mimics the motor deficits observed in PD [16,20,21]. Behavioral assessments such as the cylinder test are commonly employed to evaluate motor asymmetry, in which a reduced use of the forelimb contralateral to the lesion serves as an indicator of dopaminergic dysfunction [22,23,24].
In this sense, the present study aimed to investigate the inflammatory response in the nigrostriatal pathway following a focal 6-OHDA injection into the dorsal striatum of rats, and to assess tyrosine hydroxylase (TH) immunoreactivity and apoptotic cell death. To evaluate the functional consequences of the lesion, behavioral testing was performed using the cylinder test to quantify spontaneous forelimb use asymmetry and the open field test to assess general locomotor activity. While the unilateral intrastriatal 6-OHDA model is well-established [16,20,21], our study integrates temporal behavioral assessments with neuronal apoptosis and glial (astrocytic and microglial) responses within the same experimental design. This approach sought to provides novel insights into the interplay between neurodegeneration, inflammation, and functional impairment, which remain underexplored in previous studies, offering a clearer perspective on the progression and impact of striatal 6-OHDA lesions.
2. Materials and Methods
2.1. Animals and Ethics Statement
Twelve adult male Wistar rats (6–7 months-old; 310 ± 25 g) were obtained from Animal Facility at the Federal University of Sergipe (UFS). Animals were housed in standard polypropylene cages (33 × 40 × 17 cm) (4 animals per cage), under controlled conditions (22 ± 2 °C, 65–75% relative humidity, 12 h light/dark cycle with lights on at 7:00 a.m.), with free access to drinking water and food.
All experimental procedures were approved by the Institutional Review Board (UFS Ceua 2315030919) and conducted in accordance with the NIH Guide for the Care and Use of Laboratory Animals (NIH Publications No. 80–23, revised 1985). All efforts were made to minimize the number of animals used and to avoid discomfort and pain.
2.2. Experimental Procedures
One day prior to surgery, rats were premedicated with vitamin K (1.0 mg/kg, intramuscular; Kanakion, Roche Pharmaceuticals, Nutley, NJ, USA) to prevent excessive bleeding. On the following day, the animals were anesthetized with an intraperitoneal (i.p.) injection of ketamine hydrochloride (100 mg/kg; Ketalar, Parker-Davis, Detroit, MI, USA) and xylazine hydrochloride (10 mg/kg; Rompun, Bayer, Leverkusen, Germany). They were then positioned in a stereotaxic apparatus (Insight Scientific Equipments Ltd., Ribeirão Preto, SP, Brazil). After trichotomy, a small incision was made to expose the skull, followed by a craniotomy performed using a dental drill. Four microliters of 6-OHDA solution (4 mg/mL in 0.05% ascorbic acid in saline) (Sigma Company, St Louis, MO, USA) were slowly injected into the right striatum over 3 min using a Hamilton syringe (Hamilton Company, Reno, NV, USA), according to the following stereotaxic coordinates (in millimeters relative to bregma): anteroposterior (AP) +1.0; mediolateral (ML) +3.0; dorsoventral (DV) −4.0 [25]. The contralateral striatum served as an intrinsic control and received an equivalent volume of vehicle solution (0.05% ascorbic acid, saline). Following surgery, the animals were returned to their home cages, with free access to water and food. Paracetamol (1 mg/mL; Neo Química Ltd., São Paulo, SP, Brazil) was added to the drinking water to provide postoperative analgesia.
The concentration of 6-OHDA was selected based on well-established protocols in the literature that reliably induce a partial striatal lesion without significant mortality [26,27].
2.3. Behavioral Tests
2.3.1. Cylinder Test
The cylinder test is a behavioral assay designed to assess forelimb use asymmetry during spontaneous vertical exploration, typically induced by unilateral depletion of the nigrostriatal pathway [28]. Animals were placed individually in a transparent, open-top glass cylinder (20 cm in diameter, 40 cm in height), with a mirror positioned behind the apparatus to allow 360° observation of limb movements. Behavior was video-recorded for 5 min using a CCD Nikon DS Fi1c digital camera (Nikon Inc., Tokyo, Japan). Forelimb wall contacts were quantified separately for the ipsilateral and contralateral limbs. The preference for using the forelimb contralateral to the lesion was calculated using the formula: contralateral forelimb contacts/(contralateral + ipsilateral forelimb contacts) × 100 [29], with the result being expressed as percentage. The test was conducted 1 day before surgery (baseline), and again at 7 and 14 days after the 6-OHDA injection, with each session lasting 5 min. After each test session, the cylinder was thoroughly cleaned with 70% ethanol to eliminate residual olfactory cues.
2.3.2. Open Field Test
The open field test was employed to assess general locomotor activity and exploratory behavior in rats, as previously described [30]. The apparatus consisted of a circular arena (84 cm in diameter, 40 cm in height) constructed of wood and painted black. Animals were gently placed in the center of the arena and allowed to move freely for 5 min. The sessions were video recorded using a CCD Nikon DS-Fi1c digital camera (Nikon Inc., Tokyo, Japan) positioned above the apparatus, and the total distance traveled (in centimeters) and average locomotor speed were then calculated using Any-maze video tracking system (version 4.3; Stoelting Co., Wood Dale, IL, USA). The test was conducted 1 day before surgery (baseline), and again at 7 and 14 days after the 6-OHDA injection, with each session lasting 5 min. As described above, after each test session the arena was thoroughly cleaned with 70% ethanol to eliminate residual olfactory cues.
2.4. Perfusion and Immunohistochemical Procedures
After 14 post-lesion days (PLDs) the animals were deeply anesthetized with a mixture of ketamine hydrochloride and xylazine hydrochloride (100 mg/kg and 10 mg/kg, respectively, i.p.). Following the complete abolishment of both corneal and paw withdrawal reflexes, the animals were transcardially perfused with 0.9% warm heparinized-saline (~30 °C), followed by 4% cold paraformaldehyde (Sigma Company, St Louis, MO, USA) in 0.1 M phosphate buffer (PB), pH 7.4.
Brains were carefully removed from the skull and immersed in 20% sucrose diluted in 0.1 M PB for 24 h. Coronal sections (40 µm) were obtained using a freezing microtome (Leica Biosystems Inc., Buffalo Grove, IL, USA) at −20 °C. The sections were sequentially collected into four compartments containing antifreeze solution (30% ethylene glycol and 20% glycerol in 0.2 M PB, pH 7.4). The approximate distance between sections within the same compartment was 200 μm. All sections were stored at 4 °C in the antifreeze solution for up to two weeks before immunohistochemical processing.
The sections underwent immunohistochemical processing to detect dopaminergic (TH) immunoreactivity, inflammatory cells (Iba-1 and GFAP), and apoptotic profiles (caspase-3). In brief, free-floating sections were pre-treated with 1% hydrogen peroxide (H2O2) solution in 0.1 M PB for 20 min to block endogenous peroxidase activity, followed by incubation in a blocking solution containing 0.5% fresh skim milk and 0.3% Triton X-100 in 0.1 M PB for 20 min. Thereafter, sections were incubated overnight with primary antibodies diluted in 0.3% Triton X-100 and 0.1 M PB, pH 7.4 (Table 1). After washing, sections were incubated with biotinylated secondary antibodies for 2 h at room temperature (Table 1), washed again in 0.1 M PB (3×, 5 min each), and then incubated with avidin-biotin-peroxidase complex (Vectastain Standard ABC kit, Vector Labs, Burlingame, CA, USA) for 90 min. Following further washes in 0.1 M PB (3×, 5 min each), the sections were exposed to a chromogen solution containing 0.03% 3,3′ diaminobenzidine tetrahydrochloride (DAB; Sigma Company, St Louis, MO, USA) and 0.001% H2O2 in 0.1 M PB, pH 7.4. After the DAB reaction, sections were mounted on silanized glass slides (StarFrost, Waldemar Knittel Glasbearbeitungs GmbH, Braunschweig, Germany), dried overnight, dehydrated through a graded alcohol series, cleared in xylol, and coverslipped with Entellan (Merck, Darmstadt, Germany). To minimize variability due to background staining, all sections were processed simultaneously. Specificity of labeling was confirmed by substituting the primary antibodies with 0.1 M PB in randomly selected control sections.

Table 1.
Primary and secondary antibodies utilized.
Additionally, some sections were immunostained for TH using fluorescent labeling. In these cases, the secondary biotinylated antibody was replaced by an Alexa Fluor 555-conjugated goat anti-rabbit secondary antibody (1:700, diluted in 0.1 M PB; Invitrogen, Grand Island, NY, USA) (Table 1), applied for 2 h. Slides were then mounted with Vectashield mounting medium for fluorescence (Antifade solution; Vector Labs, Burlingame, CA, USA) and sealed with Entellan (Merck, Darmstadt, Germany).
2.5. Qualitative and Quantitative Analysis
Tissue sections were examined under brightfield illumination using an Olympus BX-41 microscope (Olympus Corporation, Tokyo, Japan) and images were acquired with a Nikon DXM-1200 digital camera (Nikon Inc., Tokyo, Japan) mounted on the microscope. Image contrast and brightness were adjusted using Adobe Photoshop CS6 software, version 13.0 (Adobe Systems Inc., San José, CA, USA).
Quantitative analysis of TH-immunoreactive neuropil in the dorsal striatum was performed by densitometry from digital images captured using the aforementioned equipment, as previously described [31]. Briefly, mean densitometric values were obtained using the NIH ImageJ software, version 1.53 (http://rsb.info.nih.gov/ij/, accessed on 11 June 2025). Measurements were taken within a 0.5 mm2 square sampling window placed across the striatum (three samples per section, three sections per animal). A contrast index (C) was then calculated using the equation C = (St − W)/(St + W), where St corresponds to optical density (OD) of the striatal region and W corresponds to the OD of the non-reactive cortical white matter, which was used as an internal reference.
TH-immunoreactive cells and caspase-3-reactive cells were quantified (five sections per animal) as cell density (cells/mm2) using Image J software, and values were compared to assess differences between the 6-OHDA-lesioned SNpc and the contralateral counterpart. To avoid biased overestimation due to split nuclei in sectioned profiles, Abercrombie’s correction factor was applied [32]. The corrected count (C) was calculated using the equation C = R (ST/ST + D), where R is the raw cell count, ST is the section thickness (in micrometers), and D is the main diameter of cell bodies along the axis perpendicular to the plane of section.
2.6. Statistical Analysis
Normality of the data was assessed using the Shapiro–Wilk test (scipy.stats.shapiro) to determine whether parametric or non-parametric tests should be used. Data collected from experimental animals were initially organized in Excel spreadsheets and imported into Python, version 3.11.12 as Pandas DataFrames for analysis.
When the data followed a normal distribution, parametric tests were applied. For non-parametric variables, the Kruskal–Wallis test (scipy.stats.kruskal) was applied to assess group differences. When significant, this was followed by Dunn’s post hoc test using scikit-posthocs to identify specific group differences.
All statistical analyses were performed using the Python programming language (version 3.11.12, main, 9 April 2025, 08:55:54) [GCC 11.4.0]. The following libraries and their respective versions were used: Pandas v.2.2.2—data handling and manipulation; NumPy v.2.0.2—numerical operations and covariance computation; SciPy v.1.14.1—statistical tests (Shapiro–Wilk, Kruskal–Wallis, Pearson correlation, Friedman, paired t-test); Scikit-Posthocs v.0.11.4—post hoc analyses (Dunn’s and Nemenyi tests); Statsmodels v.0.14.4—repeated measures ANOVA and multiple comparisons correction; Matplotlib v.3.10.0—data visualization
To ensure biologically meaningful analyses, we employed planned comparisons instead of performing all possible pairwise tests.
For repeated measures of parametric data, the paired Student’s t-test (scipy.stats.ttest_rel) was applied between specific conditions. For repeated measures of non-parametric data, the Friedman chi-square test (scipy.stats.friedmanchisquare) was used. When significant, this was followed by post hoc comparisons using the Nemenyi test from scikit-posthocs.
In cases where three or more repeated parametric measurements were present, a repeated-measures, one-way ANOVA was performed using statsmodels.api. Significant main effects were followed by pairwise comparisons, with Bonferroni correction applied for multiple testing.
All statistical tests considered a significance level of p < 0.05 unless otherwise stated.
3. Results
3.1. Cylinder Test
Assessment of spontaneous forelimb use in the cylinder test revealed a significant asymmetry in limb preference following the unilateral 6-OHDA-induced lesion. At baseline, animals exhibited a balanced use of both forelimbs (contralateral: 50.54% vs. ipsilateral: 49.46%). However, at PLD +7, there was a marked reduction in the contralateral forelimb use (31.04%), accompanied by a corresponding increase in ipsilateral forelimb use (68.96%) (Figure 1).
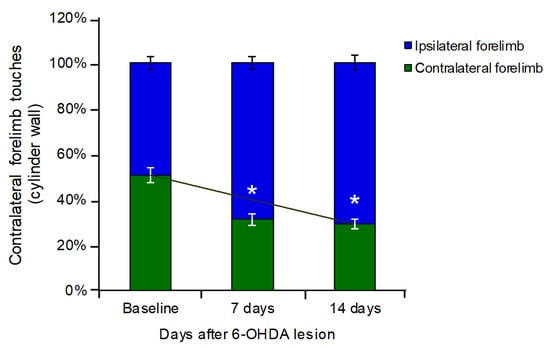
Figure 1.
Effect of unilateral striatal 6-OHDA lesion on forelimb use asymmetry in the cylinder test. At PLD −1, animals exhibited a balanced use of both forelimbs. At PLD +7, a marked reduction in the use of the forelimb contralateral to the lesion was observed (31.04% of total touches), accompanied by a corresponding increase in ipsilateral forelimb use. This asymmetrical pattern persisted at PLD +14 (contralateral: 29.09% of total touches). Values expressed in percent. * p < 0.05; Student’s t-test, contralateral vs. lesioned striatum.
This asymmetry persisted at PLD +14, with animals continuing to favor the ipsilateral forelimb (70.91%) and showing reduced use of the contralateral forelimb (29.09%), indicating sustained motor impairment resulting from the dopaminergic depletion (Figure 1).
3.2. Open Field Test
Quantitative analysis of locomotor activity in the open field test demonstrated that animals evaluated at PLD +7- and +14 exhibited a significant reduction in the total distance traveled and in the average locomotor speed compared to PLD −1 (** p < 0.01; one-way ANOVA followed by Tukey’s post hoc test) (Figure 2A,B). Representative trajectory plots revealed restricted exploratory behavior in both post-lesion time points (Figure 2C), further supporting the presence of motor impairments.
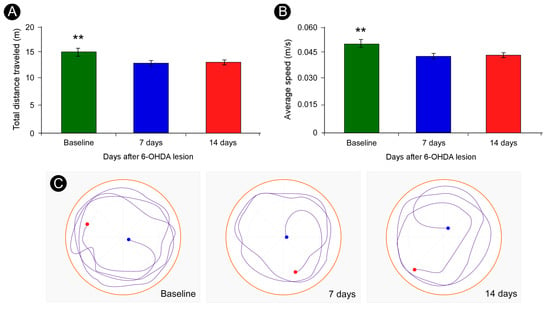
Figure 2.
Quantification of total distance traveled and average locomotor speed in the open-field test. Animals assessed at PLD + 7- and +14 spent less time moving across the arena as compared with the baseline (A). In addition, their mean locomotor speeds were reduced (B). Representative trajectories covered during open field exploration (C). Blue dots indicate starting points; red dots indicate endpoints. Data are presented as mean ± SD. ** p < 0.01, corrected for multiple comparisons; one-way ANOVA, followed by Tukey’s post hoc test.
3.3. Pattern of TH-Immunoreactivity
The 6-OHDA-induced lesion in the dorsal striatum resulted in a marked reduction in TH-immunoreactivity on the lesioned side compared to the contralateral striatum, characterized by a reduced density of TH-immunopositive fibers in the affected region (Figure 3A), quantitatively corroborated by densitometric analysis (lesioned: 0.279 ± 0.011; contralateral: 0.486 ± 0.036) (Figure 3B).
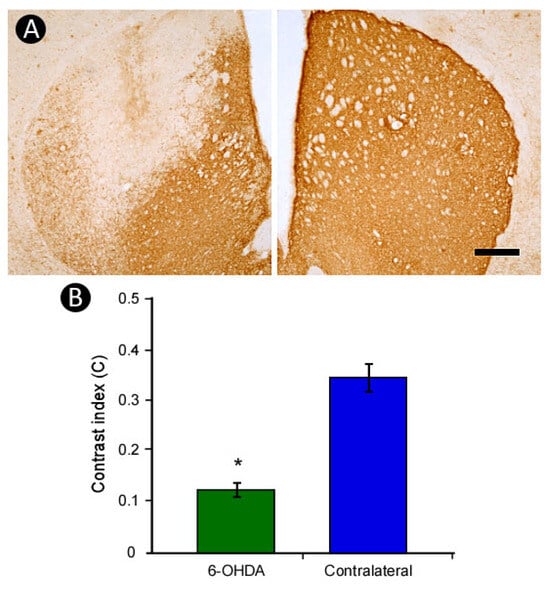
Figure 3.
Striatal lesion induced by 6-OHDA. A marked loss of TH-immunoreactivity was observed in the lesioned striatum compared to the contralateral side (A), as confirmed by densitometric analysis (B). Scale bar: 500 µm. * p < 0.05; Student’s t-test, contralateral vs. lesioned striatum.
This loss of TH-immunostaining was accompanied by an approximately 40% reduction in the number of TH-positive neurons in the SNpc ipsilateral to the lesion, an effect not observed on the no n-lesioned side (lesioned: 29.42 ± 1.22 cells/mm2; contralateral: 48.09 ± 2.58 cells/mm2; p < 0.01, Student’s t-test) (Figure 4A–C). These findings are consistent with a retrograde degenerative process, likely triggered by the toxic action of 6-OHDA on dopaminergic terminals in the striatum. In addition, TH-positive neurons on the lesioned side exhibited a 28% increase in soma size compared to the contralateral side (p < 0.05, Student’s t test) (Figure 4D,E), suggesting cellular hypertrophy as a possible compensatory or pathological response.
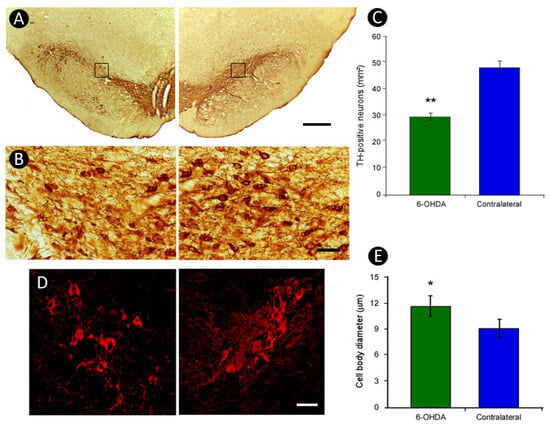
Figure 4.
Effect of unilateral striatal 6-OHDA lesion on TH-positive neurons of Substantia Nigra pars compacta (SNpc). A reduction in TH-positive neurons and fibers was detected in the Substantia Nigra ipsilateral to the lesion, with no changes observed on the contralateral side (A,B), as further demonstrated by quantitative analysis (C). Additionally, TH-positive neurons on the lesioned side showed an increase of 28% in the size of their cell bodies as compared to the contralateral side (D,E). The squares in (A) indicates the region shown at higher magnification. Scale bars: 500 µm (A); 100 µm (B); 40 µm (D). * p < 0.05, ** p < 0.01; Student’s t-test, contralateral vs. lesioned striatum.
3.4. Pattern of Inflammatory Response
The inflammatory response, assessed through Iba-1-immunohistochemistry, revealed pronounced microglial activation within the lesioned striatum. Notably, morphological alterations such as enlarged cell bodies and retracted processes were observed in microglia surrounding the 6-OHDA injection site. In contrast, microglial cells in the contralateral striatum maintained a resting phenotype, characterized by smaller somas and elongated, ramified processes (Figure 5).
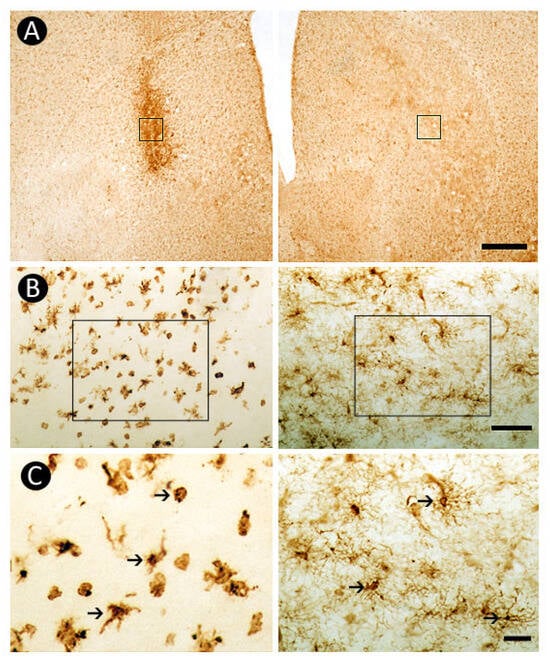
Figure 5.
Inflammatory response revealed by Iba-1-immunoreactivity in the dorsal striatum. Marked microglial activation is observed at the 6-OHDA-injected site (A), evidenced by morphological changes in Iba-1-positive cells (B, left side) when compared to the non-lesioned contralateral side (B, right side). The squares in (A) indicates the regions shown in (B). The higher magnification in (C) highlights the distinct morphological features between non-reactive (right side) and reactive microglial cells (left side). The squares in (B) indicate the region shown at higher magnification in (C). Arrows in (C) point to Iba-1-immunoreactive cells. Scale bars: 500 µm (A), 50 µm (B,C).
3.5. Pattern of Astrocytic Response
A striking pattern of gliosis was observed with GFAP immunohistochemistry, showing pronounced astrocytic activation in the lesioned striatum (Figure 6A, left side) compared with the non-lesioned side (Figure 6A, right side). In the lesioned striatum, astrocytes exhibited marked morphological changes, acquiring a hypertrophic and globular appearance with retracted processes (Figure 6B, left side) relative to the non-activated morphology on the contralateral side (Figure 6B, right side). Consistent astrocytic activation was also detected in the ipsilateral Substantia Nigra pars reticulata (SNpr) (Figure 6C), with prominent reactivity (Figure 6D, left side) compared to the contralateral counterpart (Figure 6D, right side).
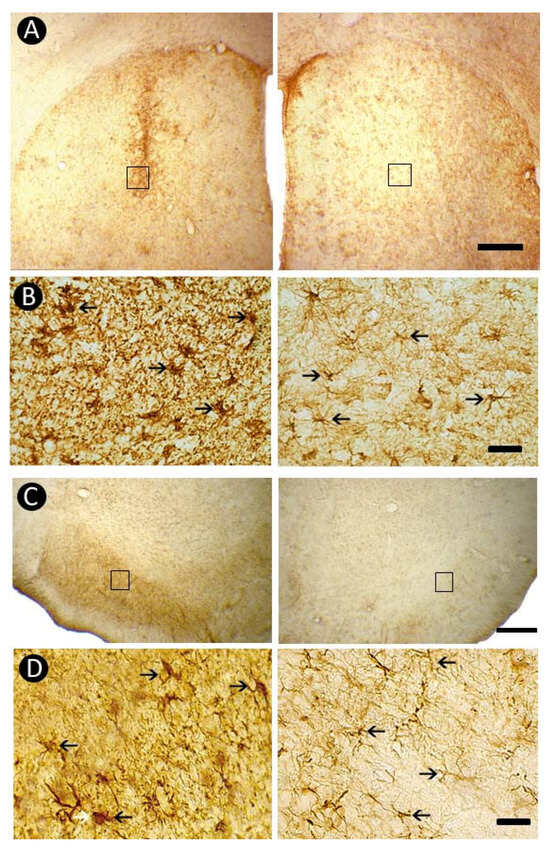
Figure 6.
Astrocytic response revealed by GFAP-immunoreactivity in the dorsal striatum and Substantia Nigra. Marked astrocytic activation is observed in the 6-OHDA–injected striatum (A), with hypertrophic GFAP-positive cells (B, left side) compared to the non-lesioned contralateral side (B, right side). A similar pattern of gliosis is evident in the ipsilateral Substantia Nigra pars reticulata (SNpr) (C), where prominent astrocytic activation (D, left side) contrasts with the contralateral counterpart (D, right side). The squares in (A,C) indicate the regions shown at higher magnification in (B,D), respectively. Arrows in (B,D) point to GFAP-immunoreactive cells. Scale bars: 500 µm (A,C), 50 µm (B,D).
3.6. Pattern of Caspase-3-Immunolabeling
Caspase-3-immunohistochemistry confirmed the presence of apoptotic cell death following 6-OHDA administration. Apoptotic profiles were clearly identified within the neurotoxin-injected striatal region, a pattern not observed in the contralateral side (Figure 7A,B). In addition, caspase-3-positive cells were detected in the SNpc ipsilateral to the lesion, a pattern absent in the contralateral hemisphere (Figure 7C), reinforcing that 6-OHDA-induced injury leads to cell death in the nigrostriatal pathway.
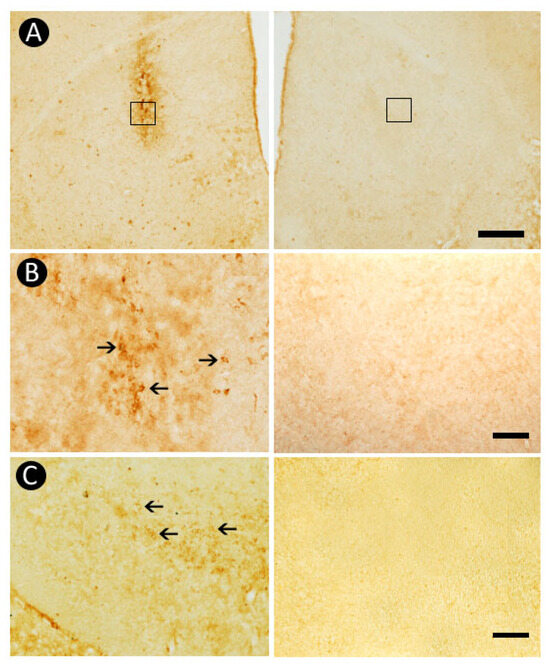
Figure 7.
Apoptotic cell death induced by 6-OHDA. Caspase-3-positive profiles were identified within the neurotoxin-injected region of the striatum, a pattern not identified in the contralateral counterpart (A,B). Moreover, apoptotic cells were detected within the Substantia Nigra pars compacta (SNpc) ipsilateral to the lesion, a pattern absent in the contralateral hemisphere (C). The squares in (A) indicates the region shown at higher magnification in (B). Arrows in (B,C) point to caspase-3-immunoreactive cells. Scale bars: 500 µm (A), 100 µm (B,C).
4. Discussion
In this study, we demonstrated that unilateral injection of 6-OHDA into the dorsal striatum of rats induces marked dopaminergic degeneration in the nigrostriatal pathway, accompanied by a marked inflammatory response and apoptotic cell death. These histopathological alterations were associated with significant motor impairments, as evidenced by the asymmetrical use of forelimbs in the cylinder test, along with decreased locomotor activity.
While the unilateral intrastriatal 6-OHDA model is a well-established paradigm in PD research [16,20,21], our study sought to integrates temporal behavioral assessments with cellular analyses of both apoptotic neuronal loss and glial responses within the same experimental framework. This integrative approach enables a direct correlation between histopathological changes and functional impairments, offering novel insights into the progression and impact of striatal 6-OHDA lesions.
Behaviorally, the results of the cylinder test provided functional evidence of the lesion-induced motor deficits. While baseline assessments revealed symmetrical forelimb use, lesioned animals exhibited a progressive shift toward increased ipsilateral reliance and reduced contralateral support at both PLD +7- and +14. These findings are consistent with the partial depletion of dopaminergic innervation on the lesioned side, resulting in motor asymmetry, a hallmark of unilateral models of parkinsonism [21,28]. Similar behavioral alterations have been reported in studies employing the cylinder test to assess motor impairments following 6-OHDA lesions in the striatum, MFB, or SNpc [20,24,33]. Additionally, the open field test further corroborated the motor alterations induced by 6-OHDA injury, revealing significant motor impairments as evidenced by a reduced total distance traveled and lower average locomotor speed compared to baseline, as previously described [21].
The reduction in TH-immunoreactivity observed in the lesioned striatum—evidenced by a less reactive region indicative of dopaminergic terminal degeneration or decreased enzyme expression, as previously reported in both rodent and primate models [20,34,35,36]—together with the concomitant loss of TH-positive neurons in the ipsilateral SNpc, supports the evidence that 6-OHDA exerts a retrograde toxic effect on dopaminergic neurons following its uptake at striatal terminals [14,16]. It has been reported that a single intrastriatal injection of 6-OHDA can lead to the loss of approximately 67% of TH-positive neurons in the SNpc and 77% in the locus coeruleus [37], underscoring the robustness of this model in replicating key histopathological features of PD [38,39,40,41].
In line with these findings, our quantitative analysis revealed an approximate 40% reduction in the number of TH-immunoreactive neurons in the SNpc on the lesioned side, further validating the recognized effectiveness of the 6-OHDA model in inducing dopaminergic neurodegeneration in rodents [40,41,42]. Although the extent of neuronal loss in our study was somewhat lower than previously reported, it still stresses the pronounced vulnerability of the nigrostriatal pathway to 6-OHDA-induced toxicity [14,16]. Variability in lesion severity may be attributable to methodological factors such as the animal species or strain, neurotoxin concentration, stereotaxic coordinates, or post-lesion survival time, all of which are known to influence the extent of neurodegeneration. Additionally, aging itself may contribute to these differences, as it is associated with a natural decline in the number of dopaminergic neurons in the SNpc and altered TH-immunoreactivity in the striatum [43,44].
Moreover, our results show that TH-positive neurons on the lesioned side exhibited a 28% increase in soma size compared to the contralateral side, suggesting cellular hypertrophy as a potential compensatory or pathological response. This enlargement may reflect increased metabolic activity preceding apoptotic signaling, indicating that PLD +14 could be an early time point, with further neuronal loss potentially occurring at later stages. These findings underscore the need for longitudinal studies to clarify the temporal dynamics of neuronal compensation and degeneration following intrastriatal 6-OHDA lesions.
Our findings also revealed pronounced microglial activation in both the striatum and SN, as evidenced by the morphological transformation of Iba-1-positive cells in the vicinity of the 6-OHDA injection site. While non-lesioned regions exhibited resting microglia characterized by small somas and thin, ramified processes, the lesioned hemisphere showed reactive microglia with hypertrophic cell bodies and retracted, thickened processes. Similarly, astrocytic activation was indicated by increased GFAP-immunoreactivity, with reactive astrocytes in both the lesioned striatum and the ipsilateral SNpr displaying enlarged somas and shortened processes. Such results are in agreement with previous studies highlighting the central role of neuroinflammation in the progression of dopaminergic neurodegeneration in this model [45]. Following intrastriatal administration of 6-OHDA, early microglial activation is typically followed by reactive astrogliosis and the upregulation of pro-inflammatory mediators such as TNF-α, IL-1β, and iNOS [46,47]. This pro-inflammatory environment exacerbates oxidative damage and directly contributes to dopaminergic cell death [48,49], establishing a deleterious feedback loop in which glial activation and neuronal injury mutually reinforce one another, thereby accelerating neurodegeneration [18,19,50]. In this context, given the established link between oxidative stress and the etiology of PD, the in vivo assessment of oxidative stress, dopaminergic neurotransmission, and cell death pathways, particularly within the SN—the primary site of dopaminergic degeneration—has emerged as a critical step in advancing the development of new therapeutic approaches and the identification of reliable biomarkers in PD [51].
It is important to consider that GFAP upregulation, although frequently used as an indicator of neuroinflammation, is more accurately interpreted as a marker of reactive gliosis. Astrocytic reactivity may arise from diverse pathological stimuli, including mechanical damage, neuronal cell death, or pro-inflammatory signaling cascades [52]. Particularly, microglia-derived cytokines such as IL-1β and TNF-α are well-established drivers of astrocytic activation, thereby linking innate immune responses to the induction of gliosis [53,54]. This crosstalk highlights the bidirectional relationship between astrocytes and microglia, in which inflammatory mediators can amplify gliosis, while reactive astrocytes themselves release chemokines and neuroactive molecules that further modulate microglial activity. In the present study, our findings of GFAP upregulation in both striatum and SN should thus be interpreted within this broader framework of glial interplay. However, because morphological data were collected only at PLD +14, we cannot address the temporal sequence by which inflammation and gliosis interact. Future studies incorporating multiple time points would be essential to clarify whether astrocytic reactivity represents an early response to microglial activation or a secondary consequence of dopaminergic degeneration.
Astrocytes both respond to and regulate other immune cells: they release chemokines that recruit peripheral leukocytes, present signals that modulate microglial phenotypes, and can influence neutrophil and T-cell activity at sites of inflammation. Microglia-astrocyte loops (microglial cytokines → astrocyte activation → astrocyte secretome → further microglial or leukocyte recruitment) are central amplifiers of neuroinflammation in many CNS disorders [55].
Recent single-cell and spatial transcriptomic studies reveal wide heterogeneity among reactive astrocytes—multiple transcriptional states exist (more than the simple A1/A2 dichotomy), and their proportions and functions evolve over time after injury. Acute, transient reactivity may be protective; chronic, sustained reactivity is more likely to be maladaptive and to drive persistent neuroinflammation. This heterogeneity helps explain why blocking “gliosis” wholesale can be harmful, and why targeted modulation is the current focus [56,57].
Finally, our immunohistochemical analysis revealed increased caspase-3 expression in both the striatum and SNpc of lesioned animals, further supporting the involvement of apoptotic pathways in 6-OHDA-induced neurodegeneration. The presence of caspase-3-immunoreactive cells in these regions aligns with previous studies showing that oxidative stress and neuroinflammation converge to activate programmed cell death mechanisms, ultimately leading to neuronal loss [16,52].
In this sense, future studies incorporating anti-inflammatory agents, antioxidants, apoptotic pathway inhibitors, and glial response modulators are critical to attenuating neuroinflammation and safeguarding the integrity of the nigrostriatal system [12,58], with the ultimate goal of slowing disease progression or improving functional outcomes. In addition, increasing attention has been directed toward axonal degeneration as an early and pivotal event in PD pathology [59,60]. Therapeutic strategies that shift the focus from solely preserving neuronal cell bodies to also addressing neuroinflammation, as well as promoting axonal protection and regeneration, may enhance the effectiveness of neuroprotective interventions. Such approaches hold considerable promise in the translational pipeline for the development of novel treatments targeting the pathogenesis of PD.
Limitations and Future Perspectives
Despite providing relevant insights into the interplay between dopaminergic degeneration and neuroinflammation following focal 6-OHDA-induced striatal lesion, we acknowledge certain limitations in our experimental design. While Iba-1 and GFAP immunoreactivity effectively reveal microglial and astrocytic activation, additional markers of inflammatory activation such as MHC-II or CD68 [61], Western blot analysis of glial and apoptotic markers [62,63], and quantification of pro-inflammatory cytokines (e.g., TNF-α, IL-1β) [11] were not assessed, due to current resource constraints. These aspects represent avenues for future research and could further elucidate the molecular mechanisms underlying neuroinflammation and neuronal degeneration in this model.
Recent studies reinforce the importance of such molecular approaches. For instance, astrocytic JWA deletion has been shown to reduce glutamate transporters and exacerbate dopaminergic neurodegeneration [64], highlighting the functional contribution of astrocytes to excitotoxic processes. In addition, glial cells are now recognized as active regulators of neuronal ferroptosis, linking inflammatory responses to selective dopaminergic vulnerability [65]. Moreover, immunomodulatory strategies such as microneedle stimulation have been reported to attenuate PD-related pathology by modulating peripheral CD4+/CD8+ T cell dynamics and their brain infiltration [66], underscoring the translational relevance of integrating neuroimmune crosstalk in future models.
Other limitations include the use of relatively young adult rats (6–7 months old), which may limit the translational relevance of our findings given that PD typically manifests in older individuals. Future studies should consider the use of middle-aged or aged animals to better mimic age-related neurodegenerative processes. Additionally, only male rats were adopted to minimize potential confounding effects of hormonal fluctuations associated with the estrous cycle, which can influence behavioral and histological outcomes. Nevertheless, we acknowledge that including both sexes would strengthen translational relevance and represents a key direction for future investigations.
In addition, although functional assessments were performed at PLD +7 and +14, morphological data were only collected at the latter time point. The absence of structural data at earlier stages prevents a precise understanding of the temporal progression between neuroinflammation and cell death. Longitudinal analyses across multiple time points would be valuable to determine whether inflammatory processes precede, accompany, or follow dopaminergic degeneration.
Moreover, the current investigation focused exclusively on the dorsal striatum and SNpc, without analyzing other relevant regions such as the ventral tegmental area (VTA), SN pars lateralis (SNpl), or the dorsal raphe nucleus (DRN), which may also contribute to the behavioral and neurochemical outcomes observed. Future studies should expand the anatomical scope and consider the involvement of serotonergic systems, especially given the potential links between PD, mood alterations, and DRN-mediated compensatory mechanisms.
In terms of behavioral assessment, our analyses were restricted to motor deficits. However, considering the well-documented interaction between motor and mood-related symptoms in PD [67], future experiments should incorporate tests for anxiety-like and motivational behaviors. Such approaches could better reflect the complexity of the disease and contribute to a more comprehensive preclinical model.
Lastly, although the 40% reduction in TH-positive neurons in the SNpc indicates a moderate lesion effect, this was accompanied by robust neuroinflammatory changes, including astrocytic and microglial activation, as well as apoptotic profiles in the SNpc. These findings highlight that even moderate dopaminergic degeneration is sufficient to trigger marked inflammatory and degenerative cascades, underscoring the translational relevance of targeting early neuroinflammatory processes in PD models. Previous tracing and lesion studies indicate that dopaminergic neuron vulnerability varies by projection target: for instance, VTA neurons are less susceptible to 6-OHDA than SNpc neurons [68], and post-lesion compensatory axonal sprouting from surviving SNpc neurons supports selective resilience [69,70]. Recent single-cell analyses further reveal molecularly distinct dopaminergic subpopulations with differential sensitivity to toxin-induced degeneration [71]. Together, these considerations suggest that the observed level of degeneration may reflect selective vulnerability and resilience among nigrostriatal subpopulations, rather than simple variability in lesion efficacy or toxin diffusion. Moreover, the observation that surviving neurons undergo hypertrophic changes provides a novel perspective on compensatory mechanisms that may influence disease progression and recovery. This integrative approach enhances the translational value of the 6-OHDA model and advances our ability to develop neuroprotective and disease-modifying strategies for PD.
5. Conclusions
Our findings support the utility of the intrastriatal 6-OHDA model for investigating the interplay between dopaminergic degeneration, neuroinflammation, and motor dysfunction in a rat model of PD, which reproduces key pathological features of PD, including selective neuronal loss, glial activation, and apoptotic signaling, accompanied by measurable motor impairments. In addition, our data raise the possibility of serotonergic involvement: dopaminergic denervation within the caudate–putamen may affect serotonergic projections, consistent with reports of functional cross-talk between the SN and DRN. This observation highlights the need to explore the serotonin/dopamine axis as a potential modulator of PD progression. From a translational perspective, these findings point to a therapeutic window in which interventions targeting neuroinflammation—such as selective inhibitors of glial activation or iNOS—may preserve neuronal integrity, modulate serotonergic dysregulation, and slow ongoing degeneration. Thus, the model provides a valuable platform not only for testing neuroprotective and anti-inflammatory strategies, but also for examining circuit-level interactions between dopaminergic and serotonergic systems. Future studies incorporating older animal cohorts, additional brain regions, and broader behavioral and molecular assessments may further enhance its translational relevance and contribute to the development of stage-specific therapeutic approaches for PD.
Author Contributions
Conceptualization, J.R.S. and M.A.M.F.; methodology, M.A.M.F., G.S.R. and J.R.S.; validation, G.S.R., L.B.J. and N.A.M.L.; formal analysis, M.A.M.F., N.A.M.L. and G.S.R.; investigation, M.A.M.F., G.S.R. and N.A.M.L.; resources, J.R.S., S.B. and R.R.L.; data curation, G.S.R., N.A.M.L. and L.B.J.; writing—original draft preparation, M.A.M.F., G.S.R., L.B.J., N.A.M.L., D.F. and H.W.M.S.; writing—review and editing, M.A.M.F., G.S.R., L.B.J., S.B., D.F. and H.W.M.S.; visualization, J.R.S., R.R.L. and D.F.; supervision, J.R.S.; project administration, J.R.S.; funding acquisition, J.R.S. All authors have read and agreed to the published version of the manuscript.
Funding
Research supported by grants from Coordination of Superior Level Staff Improvement—Brazil (CAPES) (Finance code 001) (88887.965130/2024-00), and National Council for Scientific and Technological Development—Brazil (CNPq) (141811/2023-4). MAMF was a recipient of a CAPES fellowship. GSR was a recipient of a CNPq fellowship.
Institutional Review Board Statement
The animal study protocol was approved by the Institutional Review Board (UFS Ceua 2315030919) and conducted in accordance with the NIH Guide for the Care and Use of Laboratory Animals (NIH Publications No. 80–23, revised 1985).
Informed Consent Statement
Not applicable.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors on request.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Abbreviations
The following abbreviations are used in this manuscript:
| 6-OHDA | 6-hydroxydopamine |
| DRN | Dorsal raphe nucleus |
| GFAP | Glial fibrillary acid protein |
| Iba-1 | Ionized calcium-binding adapter molecule 1 |
| IL-1β | Interleukin-1 beta |
| iNOS | Inducible nitric oxide synthase |
| MFB | Medial forebrain bundle |
| OD | Optical density |
| PD | Parkinson’s disease |
| PLD | Post-lesion days |
| ROS | Reactive oxygen species |
| SN | Substantia nigra |
| SNpc | Substantia nigra pars compacta |
| SNpl | Substantia nigra pars lateralis |
| SNpr | Substantia nigra pars reticulata |
| TH | Tyrosine hydroxylase |
| TNF-α | Tumor necrosis factor alpha |
| VTA | Ventral tegmental area |
References
- Bloem, B.R.; Okun, M.S.; Klein, C. Parkinson’s disease. Lancet 2021, 397, 2284–2303. [Google Scholar] [CrossRef]
- Hou, Y.; Dan, X.; Babbar, M.; Wei, Y.; Hasselbalch, S.G.; Croteau, D.L.; Bohr, V.A. Ageing as a risk factor for neurodegenerative disease. Nat. Rev. Neurol. 2019, 15, 565–581. [Google Scholar] [CrossRef]
- Pringsheim, T.; Jette, N.; Frolkis, A.; Steeves, T.D.L. The prevalence of Parkinson’s disease: A systematic review and meta-analysis. Mov. Disord. 2014, 29, 1583–1590. [Google Scholar] [CrossRef] [PubMed]
- Dauer, W.; Przedborski, S. Parkinson’s disease: Mechanisms and models. Neuron 2003, 39, 889–909. [Google Scholar] [CrossRef] [PubMed]
- Poewe, W.; Seppi, K.; Tanner, C.; Halliday, G.M.; Brundin, P.; Volkman, J.; Schrag, A.E.; Lang, A.E. Parkinson disease. Nat. Rev. Dis. Primers 2017, 3, 17013. [Google Scholar] [CrossRef] [PubMed]
- Kalia, L.V.; Lang, A.E. Parkinson’s disease. Lancet 2015, 386, 896–912. [Google Scholar] [CrossRef]
- Aarsland, D.; Batzu, L.; Halliday, G.M.; Geurtsen, G.J.; Ballard, C.; Chaudhuri, K.R.; Weintraub, D. Parkinson disease-associated cognitive impairment. Nat. Rev. Dis. Primers 2021, 7, 47. [Google Scholar] [CrossRef]
- Hussein, A.; Guevara, C.A.; Del Valle, P.; Gupta, S.; Benson, D.L.; Huntley, G.W. Non-motor symptoms of Parkinson’s disease: The neurobiology of early psychiatric and cognitive dysfunction. Neuroscientist 2023, 29, 97–116. [Google Scholar] [CrossRef]
- Wang, T.; Shi, C.; Luo, H.; Zheng, H.; Fan, L.; Tang, M.; Su, Y.; Yang, J.; Mao, C.; Xu, Y. Neuroinflammation in Parkinson’s Disease: Triggers, Mechanisms, and Immunotherapies. Neuroscientist 2022, 28, 364–381. [Google Scholar] [CrossRef]
- Tansey, M.G.; McCoy, M.K.; Frank-Cannon, T.C. Neuroinflammatory mechanisms in Parkinson’s disease: Potential environmental triggers, pathways, and targets for early therapeutic intervention. Exp. Neurol. 2007, 208, 1–25. [Google Scholar] [CrossRef]
- Hirsch, E.C.; Hunot, S. Neuroinflammation in Parkinson’s disease: A target for neuroprotection? Lancet Neurol. 2009, 8, 382–397. [Google Scholar] [CrossRef]
- Tansey, M.G.; Goldberg, M.S. Neuroinflammation in Parkinson’s disease: Its role in neuronal death and implications for therapeutic intervention. Neurobiol. Dis. 2010, 37, 510–518. [Google Scholar] [CrossRef] [PubMed]
- Thiele, S.L.; Warre, R.; Nash, J.E. Development of a unilaterally-lesioned 6-OHDA mouse model of Parkinson’s disease. J. Vis. Exp. 2012, 60, 3234. [Google Scholar] [CrossRef]
- Jagmag, S.A.; Tripathi, N.; Shukla, S.D.; Maiti, S.; Khurana, S. Evaluation of models of Parkinson’s Disease. Front. Neurosci. 2016, 9, 503. [Google Scholar] [CrossRef] [PubMed]
- Ungerstedt, U. 6-hydroxy-dopamine induced degeneration of central monoamine neurons. Eur. J. Pharmacol. 1968, 5, 107–110. [Google Scholar] [CrossRef]
- Simola, N.; Morelli, M.; Carta, A.R. The 6-Hydroxydopamine model of Parkinson’s disease. Neurotox. Res. 2007, 11, 151–167. [Google Scholar] [CrossRef]
- Vivekanantham, S.; Shah, S.; Dewji, R.; Dewji, A.; Khatri, C.; Ologunde, R. Neuroinflammation in Parkinson’s disease: Role in neurodegeneration and tissue repair. Int. J. Neurosci. 2015, 125, 717–725. [Google Scholar] [CrossRef]
- Bayo-Olugbami, A.; Nafiu, A.B.; Amin, A.; Ogundele, O.M.; Lee, C.C.; Owoyele, B.V. Vitamin D attenuated 6-OHDA-induced behavioural deficits, dopamine dysmetabolism, oxidative stress, and neuro-inflammation in mice. Nutr. Neurosci. 2022, 25, 823–834. [Google Scholar] [CrossRef]
- Lugrin, J.; Rosenblatt-Velin, N.; Parapanov, R.; Liaudet, L. The role of oxidative stress during inflammatory processes. Biol. Chem. 2014, 395, 203–230. [Google Scholar] [CrossRef]
- Barata-Antunes, S.; Teixeira, F.G.; Mendes-Pinheiro, B.; Domingues, A.V.; Vilaça-Faria, H.; Marote, A.; Silva, D.; Sousa, R.A.; Salgado, A.J. Impact of aging on the 6-OHDA-induced rat model of Parkinson’s disease. Int. J. Mol. Sci. 2020, 21, 3459. [Google Scholar] [CrossRef]
- Su, R.J.; Zhen, J.L.; Wang, W.; Zhang, J.L.; Zheng, Y.; Wang, X.M. Time-course behavioral features are correlated with Parkinson’s disease-associated pathology in a 6-hydroxydopamine hemiparkinsonian rat model. Mol. Med. Rep. 2018, 17, 3356–3363. [Google Scholar] [CrossRef] [PubMed]
- Olsson, M.; Nikkhah, G.; Bentlage, C.; Björklund, A. Forelimb akinesia in the rat Parkinson model: Differential effects of dopamine agonists and nigral transplants as assessed by a new stepping test. J. Neurosci. 1995, 5, 3863–3875. [Google Scholar] [CrossRef] [PubMed]
- Iancu, R.; Mohapel, P.; Brundin, P.; Paul, G. Behavioral characterization of a unilateral 6-OHDA-lesion model of Parkinson’s disease in mice. Behav. Brain Res. 2005, 162, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Messaoudi, I.; Nsibi, A.; Saoud, H.; Saidi, N.E.; Mani, S. Unilateral 6-hydroxydopamine-lesioned rat as relevant model to study the pain related to Parkinson’s disease. Neurol. Neurobiol. 2020, 3, 1–5. [Google Scholar] [CrossRef]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates, 7th ed.; Academic Press: Cambridge, MA, USA, 2013. [Google Scholar]
- Li, J.; Sun, Y.; Chen, J. Transcriptome sequencing in a 6-hydroxydopamine rat model of Parkinson’s disease. Genes Genet. Syst. 2019, 94, 61–69. [Google Scholar] [CrossRef]
- Lu, J.; Liu, X.; Tian, Y.; Li, H.; Ren, Z.; Liang, S.; Zhang, G.; Zhao, C.; Li, X.; Wang, T.; et al. Moxibustion Exerts a Neuroprotective Effect through Antiferroptosis in Parkinson’s Disease. Evid. Based Complement. Alternat Med. 2019, 2019, 2735492. [Google Scholar] [CrossRef]
- Schallert, T.; Fleming, S.M.; Leasure, J.L.; Tillerson, J.L.; Bland, S.T. CNS plasticity and assessment of forelimb sensorimotor outcome in unilateral rat models of stroke, cortical ablation, parkinsonism and spinal cord injury. Neuropharmacology 2000, 39, 777–787. [Google Scholar] [CrossRef]
- Boix, J.; Padel, T.; Paul, G. A partial lesion model of Parkinson’s disease in mice--characterization of a 6-OHDA-induced medial forebrain bundle lesion. Behav. Brain Res. 2015, 284, 196–206. [Google Scholar] [CrossRef]
- Carvalho, M.M.; Campos, F.L.; Coimbra, B.; Pego, J.M.; Rodrigues, C.; Lima, R.; Rodrigues, A.J.; Sousa, N.; Salgado, A.J. Behavioral characterization of the 6-hydroxidopamine model of Parkinson’s disease and pharmacological rescuing of non-motor deficits. Mol. Neurodegener. 2013, 8, 14. [Google Scholar] [CrossRef]
- Freire, M.A.M.; Guimaraes, J.S.; Santos, J.R.; Simplicio, H.; Gomes-Leal, W. Morphometric analysis of NADPH diaphorase reactive neurons in a rat model of focal excitotoxic striatal injury. Neuropathology 2016, 36, 527–534. [Google Scholar] [CrossRef]
- Abercrombie, M. Estimation of nuclear population from microtome sections. Anat. Rec. 1946, 94, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Li, X.; Zhang, L.; Zhang, Y.; Qi, X.; Wang, S.; Qin, C. Longitudinal assessment of motor function following the unilateral intrastriatal 6-hydroxydopamine lesion model in mice. Front. Behav. Neurosci. 2022, 16, 982218. [Google Scholar] [CrossRef] [PubMed]
- Glajch, K.E.; Fleming, S.M.; Surmeier, D.J.; Osten, P. Sensorimotor assessment of the unilateral 6-hydroxydopamine mouse model of Parkinson’s disease. Behav. Brain Res. 2012, 230, 309–316. [Google Scholar] [CrossRef] [PubMed]
- Santana, M.B.; Halje, P.; Simplício, H.; Richter, U.; Freire, M.A.M.; Petersson, P.; Nicolelis, M. Spinal cord stimulation alleviates motor deficits in a primate model of Parkinson Disease. Neuron 2014, 84, 716–722. [Google Scholar] [CrossRef]
- Stott, S.R.W.; Barker, R.A. Time course of dopamine neuron loss and glial response in the 6-OHDA striatal mouse model of Parkinson’s disease. Eur. J. Neurosci. 2014, 39, 1042–1056. [Google Scholar] [CrossRef]
- Bustelli, I.B.; Oliveira, L.M.; Netto, N.F.C.; Stilhano, R.S.; Caetano, A.L. Behavioral effects of 6-hydroxydopamine-induced damage to nigro-striatal pathway and locus coeruleus as a rodent model of Parkinson’s disease. Behav. Brain Res. 2024, 462, 114873. [Google Scholar] [CrossRef]
- Bagga, V.; Dunnett, S.B.; Fricker, R.A. The 6-OHDA mouse model of Parkinson’s disease—Terminal striatal lesions provide a superior measure of neuronal loss and replacement than median forebrain bundle lesions. Behav. Brain Res. 2015, 288, 107–117. [Google Scholar] [CrossRef]
- Porras, A.R.B.; Ramirez, L.B.; Turner, L.F. Striatal vs. Nigral 6-OHDA lesion models: A closer look at Parkinson’s disease pathology. Braz. J. Anim. Environ. Res. 2025, 8, e80427. [Google Scholar] [CrossRef]
- Grealish, S.; Mattsson, B.; Draxler, P.; Björklund, A. Characterisation of behavioural and neurodegenerative changes induced by intrastatial 6-OHDA lesion in mice. Eur. J. Neurosci. 2010, 31, 2266–2278. [Google Scholar] [CrossRef]
- Francardo, V.; Recchia, A.; Popovic, N.; Andersson, D.; Nissbrandt, H.; Cenci, M.A. Impact of the lesion procedure on the profiles of motor impairment and molecular responsiveness to L-DOPA in the 6-hydroxydopamine mouse model of Parkinson’s disease. Neurobiol. Dis. 2011, 42, 327–340. [Google Scholar] [CrossRef]
- Branchi, I.; D’Andrea, I.; Armida, M.; Cassano, T.; Pezzola, A.; Potenza, R.L.; Morgese, M.G.; Popoli, P.; Alleva, E. Nonmotor symptoms in Parkinson’s disease: Investigating early-phase onset of behavioral dysfunction in the 6-hydroxydopamine-lesioned rat model. J. Neurosci. Res. 2010, 88, 2051–2062. [Google Scholar] [CrossRef]
- Collier, T.J.; Kanaan, N.M.; Kordower, J.H. Ageing as a primary risk factor for Parkinson’s disease: Evidence from studies of non-human primates. Nat. Rev. Neurosci. 2007, 8, 499–504. [Google Scholar] [CrossRef]
- Norrara, B.; Arrais, A.C.; Costa, I.M.; Santos, J.R.; Engelberth, R.C.G.J.; Cavalcante, J.S.; Guzen, F.; Cavalcanti, J.R.; Freire, M.A.M. Pattern of tyrosine hydroxylase expression during aging of mesolimbic pathway of the rat. J. Chem. Neuroanat. 2018, 92, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, V.S.L.; Valente-Amaral, A.; Monteiro, R.F.M.; Meira, C.L.S.; de Meira, N.S.; da Silva, M.N.; Pinheiro, J.J.V.; Bastos, G.N.T.; Felicio, J.S.; Yamada, E.S. Aqueous extract of Swietenia macrophylla leaf exerts an anti-inflammatory effect in a murine model of Parkinson’s disease induced by 6-OHDA. Front. Neurosci. 2024, 18, 1351718. [Google Scholar] [CrossRef] [PubMed]
- Koprich, J.B.; Reske-Nielsen, C.; Mithal, P.; Isacson, O. Neuroinflammation mediated by IL-1β increases susceptibility of dopamine neurons to degeneration in an animal model of Parkinson’s disease. J. Neuroinflamm. 2008, 5, 8. [Google Scholar] [CrossRef] [PubMed]
- Villar-Cheda, B.; Valenzuela, R.; Rodriguez-Perez, A.I.; Guerra, M.J.; Labandeira-Garcia, J.L. Aging-related changes in the nigral angiotensin system enhances proinflammatory and pro-oxidative markers and 6-OHDA-induced dopaminergic degeneration. Neurobiol. Aging 2012, 33, 204.e1–204.e11. [Google Scholar] [CrossRef]
- McGeer, P.L.; McGeer, E.G. Inflammation and neurodegeneration in Parkinson’s disease. Parkinsonism Relat. Disord. 2004, 10 (Suppl. S1), S3–S7. [Google Scholar] [CrossRef]
- Peterson, L.J.; Flood, P.M. Oxidative stress and microglial cells in Parkinson’s disease. Mediators Inflamm. 2012, 2012, 401264. [Google Scholar] [CrossRef]
- Block, M.L.; Zecca, L.; Hong, J.S. Microglia-mediated neurotoxicity: Uncovering the molecular mechanisms. Nat. Rev. Neurosci. 2007, 8, 57–69. [Google Scholar] [CrossRef]
- Trist, B.G.; Hare, D.J.; Double, K.L. Oxidative stress in the aging substantia nigra and the etiology of Parkinson’s disease. Aging Cell 2019, 18, e13031. [Google Scholar] [CrossRef]
- Zamanian, J.L.; Xu, L.; Foo, L.C.; Nouri, N.; Zhou, L.; Giffard, R.G.; Barres, B.A. Genomic analysis of reactive astrogliosis. J. Neurosci. 2012, 32, 6391–6410. [Google Scholar] [CrossRef] [PubMed]
- Liddelow, S.A.; Barres, B.A. Reactive astrocytes: Production, function, and therapeutic potential. Immunity 2017, 46, 957–967. [Google Scholar] [CrossRef] [PubMed]
- Freire, M.A.M.; Rocha, G.S.; Bittencourt, L.O.; Falcao, D.; Lima, R.R.; Cavalcanti, J.R. Cellular and molecular pathophysiology of traumatic brain injury: What have we learned so far? Biology 2023, 12, 1139. [Google Scholar] [CrossRef] [PubMed]
- Matejuk, A.; Ransohoff, R.M. Crosstalk Between Astrocytes and Microglia: An Overview. Front. Immunol. 2020, 11, 1416. [Google Scholar] [CrossRef]
- Patani, R.; Hardingham, G.E.; Liddelow, S.A. Functional roles of reactive astrocytes in neuroinflammation and neurodegeneration. Nat. Rev. Neurol. 2023, 19, 395–409. [Google Scholar] [CrossRef]
- Cieri, M.B.; Ramos, A.J. Astrocytes, reactive astrogliosis, and glial scar formation in traumatic brain injury. Neural Regen. Res. 2025, 20, 973–989. [Google Scholar] [CrossRef]
- Subramaniam, S.R.; Federoff, H.J. Targeting microglial activation states as a therapeutic avenue in Parkinson’s disease. Front. Aging Neurosci. 2017, 9, 176. [Google Scholar] [CrossRef]
- Wong, Y.C.; Luk, K.; Purtell, K.; Nanni, S.B.; Stoessi, A.J.; Trudeau, L.E.; Yue, Z.; Krainc, D.; Oertel, W.; Obeso, J.A.; et al. Neuronal vulnerability in Parkinson disease: Should the focus be on axons and synaptic terminals? Mov. Disord. 2019, 34, 1406–1422. [Google Scholar] [CrossRef]
- Rocha, G.S.; Freire, M.A.M.; Falcao, D.; Outeiro, T.F.; Lima, R.R.; Santos, J.R. Neurodegeneration in Parkinson’s disease: Are we looking at the right spot? Mol. Brain 2025, 18, 68. [Google Scholar] [CrossRef]
- Graeber, M.B.; Streit, W.J. Microglia: Biology and pathology. Acta Neuropathol. 2020, 119, 89–105. [Google Scholar] [CrossRef]
- Le, T.M.; Takarada-Iemata, M.; Ta, H.M.; Roboon, J.; Ishii, H.; Tamatani, T.; Hattori, T.; Hori, O. Ndrg2 deficiency ameliorates neurodegeneration in experimental autoimmune encephalomyelitis. J. Neurochem. 2018, 145, 139–153. [Google Scholar] [CrossRef]
- Guo, M.; Lu, B.; Gan, J.; Wang, S.; Jiang, X.; Li, H. Apoptosis detection: A purpose-dependent approach selection. Cell Cycle 2021, 20, 1033–1040. [Google Scholar] [CrossRef]
- Wang, R.; Zhao, X.; Xu, J.; Wen, Y.; Li, A.; Lu, M.; Zhou, J. Astrocytic JWA deletion exacerbates dopaminergic neurodegeneration by decreasing glutamate transporters in mice. Cell Death Dis. 2018, 9, 352. [Google Scholar] [CrossRef]
- Li, M.; Chen, M.; Li, H.; Gao, D.; Zhao, L.; Zhu, M. Glial cells improve Parkinson’s disease by modulating neuronal function and regulating neuronal ferroptosis. Front. Cell Dev. Biol. 2025, 12, 1510897. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Choi, Y.; Kim, J.S.; Lee, H.; Ju, I.G.; Yoo, N.Y.; La, S.; Jeong, D.H.; Na, C.; Park, H.-J.; et al. Stimulation of microneedles alleviates pathology of Parkinson’s disease in mice by regulating the CD4+/CD8+ cells from the periphery to the brain. Front. Immunol. 2024, 15, 1454102. [Google Scholar] [CrossRef] [PubMed]
- Freire, M.A.M.; Santos, J.R. Parkinson’s disease: General features, effects of levodopa treatment and future directions. Front. Neuroanat. 2010, 4, 146. [Google Scholar] [CrossRef] [PubMed]
- Grant, R.J.; Clarke, P.B.S. Susceptibility of ascending dopamine projections to 6-hydroxydopamine in rats: Effect of hypothermia. Neuroscience 2002, 115, 1281–1294. [Google Scholar] [CrossRef]
- Tanguay, W.; Ducrot, C.; Giguère, N.; Bourque, M.J.; Trudeau, L.E. Neonatal 6-OHDA lesion of the SNc induces striatal compensatory sprouting from surviving SNc dopaminergic neurons without VTA contribution. Eur. J. Neurosci. 2021, 54, 6618–6632. [Google Scholar] [CrossRef]
- Paß, T.; Ricke, K.M.; Hofmann, P.H.; Chowdhury, R.S.; Nie, Y.; Chinnery, P.; Endepols, H.; Neumaier, B.; Carvalho, A.; Rigoux, L.; et al. Preserved striatal innervation maintains motor function despite severe loss of nigral dopaminergic neurons. Brain 2024, 147, 3189–3203. [Google Scholar] [CrossRef]
- Salmani, B.Y.; Lahti, L.; Gillberg, L.; Jacobsen, J.K.; Mantas, I.; Svenningsson, P.; Perlmann, T. Transcriptomic atlas of midbrain dopamine neurons uncovers differential vulnerability in a Parkinsonism lesion model. Elife 2024, 12, RP89482. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).