

Advancements in Müller Glia Reprogramming: Pioneering Approaches for Retinal Neuron Regeneration


Abstract
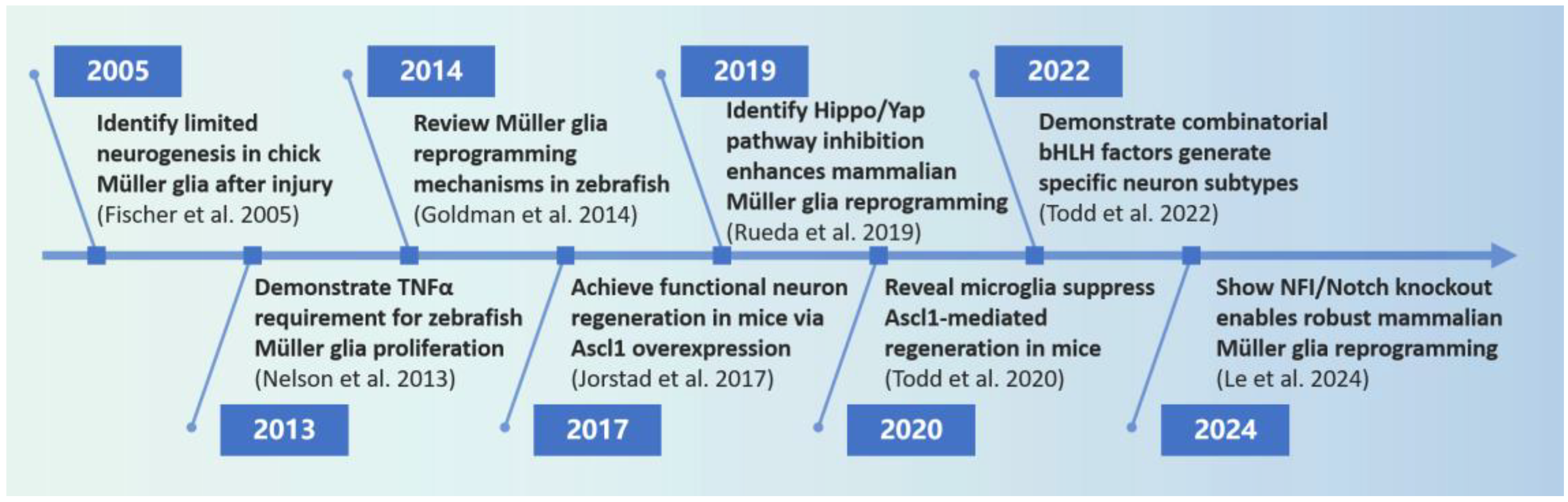
1. Introduction
2. Search Strategy
3. Müller Glia Responses to Injury
4. Müller Glia Reprogramming Mechanisms
4.1. Cytokines and Müller Glia Reprogramming
4.2. Key Signaling Pathways in Müller Glia Reprogramming
4.2.1. NFkB Signaling
4.2.2. Notch Signaling
4.2.3. Hippo/Yap Signaling
4.2.4. Wnt Signaling
4.2.5. Jak/Stat Signaling
4.2.6. MAPK Signaling
4.3. Transcriptional Regulation in Müller Glia Reprogramming
4.3.1. bHLH Transcription Factors
4.3.2. NFI Factors
4.4. Epigenetics in Müller Glia Reprogramming
4.4.1. Histone Modifications
4.4.2. DNA Methylation
4.4.3. MicroRNA-Mediated Gene Silencing

{kind=link}
{kind=link}
| Mechanism | Influence | Factors | Model | Reference |
|---|---|---|---|---|
| Cytokines | Induce MG proliferation | TNFα, IL-1β and IL-10 | Zebrafish | [14,28] |
| Promote MGPCs formation | HBEGF and FGF, MDK | Chick, Mouse | [30,32] | |
| Signaling pathway | Induce MG proliferation | Yap, Wnt | Mouse, Xenopus | [40,43] |
| Suppress MG proliferation | NFkB, Notch | Zebrafish, Mouse, Rat | [16,34,35,36,39] | |
| Promote MGPCs formation | Yap | Zebrafish | [41] | |
| prevent MGPCs formation | Hippo | Mouse | [17] | |
| Suppress differentiation of MGPCs into neurons | STAT | Mouse | [46] | |
| Transcriptional regulation | Enhance MG neurogenesis | Ascl1, Ascl1 + TSA, Atoh1/Ascl1 | Zebrafish, Mouse, Human MG culture | [3,46,53,55] |
| Influence regenerated neuron types | Atoh1/Ascl1, Ascl1/Pou4f2/Islet1/Atoh1 | Mouse | [18,46] | |
| Promote MGPCs proliferation | ID factors | Chick | [58] | |
| Suppress differentiation of MGPCs into neurons | ID factors, NFI factors | Chick, Zebrafish, Mouse | [19,34,36,58,60] | |
| Epigenetics | Suppress MG proliferation | Brd, miR-216a | Zebrafish | [65,80] |
| Promote MGPCs formation | miR-124/miR-9, miR-25/miR-124 | Mouse, Mouse MG culture | [75,76] | |
| Prevent MGPCs formation | Histone acetylation, Brd, DNA methylation, let 7, miR-216a | Zebrafish, Mouse, Rat | [61,65,69,70,77,80] | |
| Promote MGPCs proliferation | Dicer-dependent miRNA biogenesis pathway | Zebrafish | [78] | |
| Suppress MGPCs proliferation | Histone acetylation, miR-203 | Zebrafish | [61,78] | |
| Promote differentiation of MGPCs into neurons | DNA methylation, miR-18a | Zebrafish, Mouse | [70,81] | |
| Suppress Neuron differentiation | DNA methylation | Zebrafish | [72] | |
| Suppress MG neurogenesis | Histone acetylation, miR-25/miR-124 | Mouse | [3,76] |
4.5. Microenvironment in Müller Glia Reprogramming
5. Contribution of Müller Glia Reprogramming to Retinal Neuron Regeneration
6. Potential Implications of Müller Glia Reprogramming for Retinal Diseases
7. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bringmann, A.; Pannicke, T.; Grosche, J.; Francke, M.; Wiedemann, P.; Skatchkov, S.N.; Osborne, N.N.; Reichenbach, A. Müller Cells in the Healthy and Diseased Retina. Prog. Retin. Eye Res. 2006, 25, 397–424. [Google Scholar] [CrossRef] [PubMed]
- Vecino, E.; Rodriguez, F.D.; Ruzafa, N.; Pereiro, X.; Sharma, S.C. Glia-Neuron Interactions in the Mammalian Retina. Prog. Retin. Eye Res. 2016, 51, 1–40. [Google Scholar] [CrossRef]
- Jorstad, N.L.; Wilken, M.S.; Grimes, W.N.; Wohl, S.G.; VandenBosch, L.S.; Yoshimatsu, T.; Wong, R.O.; Rieke, F.; Reh, T.A. Stimulation of Functional Neuronal Regeneration from Müller Glia in Adult Mice. Nature 2017, 548, 103–107. [Google Scholar] [CrossRef] [PubMed]
- Goldman, D. Müller Glial Cell Reprogramming and Retina Regeneration. Nat. Rev. Neurosci. 2014, 15, 431–442. [Google Scholar] [CrossRef] [PubMed]
- Lahne, M.; Nagashima, M.; Hyde, D.R.; Hitchcock, P.F. Reprogramming Müller Glia to Regenerate Retinal Neurons. Annu. Rev. Vis. Sci. 2020, 6, 171–193. [Google Scholar] [CrossRef]
- Bringmann, A.; Iandiev, I.; Pannicke, T.; Wurm, A.; Hollborn, M.; Wiedemann, P.; Osborne, N.N.; Reichenbach, A. Cellular Signaling and Factors Involved in Müller Cell Gliosis: Neuroprotective and Detrimental Effects. Prog. Retin. Eye Res. 2009, 28, 423–451. [Google Scholar] [CrossRef]
- Nishino, R.; Nomura-Komoike, K.; Iida, T.; Fujieda, H. Cell Cycle-Dependent Activation of Proneural Transcription Factor Expression and Reactive Gliosis in Rat Müller Glia. Sci. Rep. 2023, 13, 22712. [Google Scholar] [CrossRef]
- Lenkowski, J.R.; Raymond, P.A. Müller Glia: Stem Cells for Generation and Regeneration of Retinal Neurons in Teleost Fish. Prog. Retin. Eye Res. 2014, 40, 94–123. [Google Scholar] [CrossRef]
- Turner, D.L.; Snyder, E.Y.; Cepko, C.L. Lineage-Independent Determination of Cell Type in the Embryonic Mouse Retina. Neuron 1990, 4, 833–845. [Google Scholar] [CrossRef]
- Jadhav, A.P.; Roesch, K.; Cepko, C.L. Development and Neurogenic Potential of Müller Glial Cells in the Vertebrate Retina. Prog. Retin. Eye Res. 2009, 28, 249–262. [Google Scholar] [CrossRef]
- Graf, T.; Enver, T. Forcing Cells to Change Lineages. Nature 2009, 462, 587–594. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Yang, Y.; Liu, J.; Qian, L. Direct Cell Reprogramming: Approaches, Mechanisms and Progress. Nat. Rev. Mol. Cell Biol. 2021, 22, 410–424. [Google Scholar] [CrossRef]
- Fischer, A.J. Neural Regeneration in the Chick Retina. Progress. Retin. Eye Res. 2005, 24, 161–182. [Google Scholar] [CrossRef]
- Nelson, C.M.; Ackerman, K.M.; O’Hayer, P.; Bailey, T.J.; Gorsuch, R.A.; Hyde, D.R. Tumor Necrosis Factor-Alpha Is Produced by Dying Retinal Neurons and Is Required for Muller Glia Proliferation during Zebrafish Retinal Regeneration. J. Neurosci. 2013, 33, 6524–6539. [Google Scholar] [CrossRef] [PubMed]
- Todd, L.; Finkbeiner, C.; Wong, C.K.; Hooper, M.J.; Reh, T.A. Microglia Suppress Ascl1-Induced Retinal Regeneration in Mice. Cell Rep. 2020, 33, 108507. [Google Scholar] [CrossRef]
- Le, N.; Vu, T.-D.; Palazzo, I.; Pulya, R.; Kim, Y.; Blackshaw, S.; Hoang, T. Robust Reprogramming of Glia into Neurons by Inhibition of Notch Signaling and Nuclear Factor I (NFI) Factors in Adult Mammalian Retina. Sci. Adv. 2024, 10, eadn2091. [Google Scholar] [CrossRef]
- Rueda, E.M.; Hall, B.M.; Hill, M.C.; Swinton, P.G.; Tong, X.; Martin, J.F.; Poché, R.A. The Hippo Pathway Blocks Mammalian Retinal Müller Glial Cell Reprogramming. Cell Rep. 2019, 27, 1637–1649.e6. [Google Scholar] [CrossRef] [PubMed]
- Todd, L.; Jenkins, W.; Finkbeiner, C.; Hooper, M.J.; Donaldson, P.C.; Pavlou, M.; Wohlschlegel, J.; Ingram, N.; Rieke, F.; Reh, T.A.; et al. Reprogramming Müller Glia to Regenerate Ganglion-like Cells in Adult Mouse Retina with Developmental Transcription Factors. Sci. Adv. 2022, 8, eabq7219. [Google Scholar] [CrossRef] [PubMed]
- Hoang, T.; Wang, J.; Boyd, P.; Wang, F.; Santiago, C.; Jiang, L.; Yoo, S.; Lahne, M.; Todd, L.J.; Jia, M.; et al. Gene Regulatory Networks Controlling Vertebrate Retinal Regeneration. Science 2020, 370, eabb8598. [Google Scholar] [CrossRef]
- Lyu, P.; Iribarne, M.; Serjanov, D.; Zhai, Y.; Hoang, T.; Campbell, L.J.; Boyd, P.; Palazzo, I.; Nagashima, M.; Silva, N.J.; et al. Common and Divergent Gene Regulatory Networks Control Injury-Induced and Developmental Neurogenesis in Zebrafish Retina. Nat. Commun. 2023, 14, 8477. [Google Scholar] [CrossRef]
- Martins, R.R.; Zamzam, M.; Tracey-White, D.; Moosajee, M.; Thummel, R.; Henriques, C.M.; MacDonald, R.B. Müller Glia Maintain Their Regenerative Potential despite Degeneration in the Aged Zebrafish Retina. Aging Cell 2022, 21, e13597. [Google Scholar] [CrossRef] [PubMed]
- Turkalj, B.; Quallich, D.; Bessert, D.A.; Kramer, A.C.; Cook, T.A.; Thummel, R. Development and Characterization of a Chronic Photoreceptor Degeneration Model in Adult Zebrafish That Does Not Trigger a Regenerative Response. Exp. Eye Res. 2021, 209, 108630. [Google Scholar] [CrossRef] [PubMed]
- Taylor, O.B.; El-Hodiri, H.M.; Palazzo, I.; Todd, L.; Fischer, A.J. Regulating the Formation of Müller Glia-Derived Progenitor Cells in the Retina. Glia 2025, 73, 4–24. [Google Scholar] [CrossRef] [PubMed]
- Fogerty, J.; Song, P.; Boyd, P.; Grabinski, S.E.; Hoang, T.; Reich, A.; Cianciolo, L.T.; Blackshaw, S.; Mumm, J.S.; Hyde, D.R.; et al. Notch Inhibition Promotes Regeneration and Immunosuppression Supports Cone Survival in a Zebrafish Model of Inherited Retinal Dystrophy. J. Neurosci. 2022, 42, 5144–5158. [Google Scholar] [CrossRef]
- Cheng, X.; Gao, H.; Tao, Z.; Yin, Z.; Cha, Z.; Huang, X.; Zhang, Y.; Zeng, Y.; He, J.; Ge, L.; et al. Repopulated Retinal Microglia Promote Müller Glia Reprogramming and Preserve Visual Function in Retinal Degenerative Mice. Theranostics 2023, 13, 1698–1715. [Google Scholar] [CrossRef]
- Fischer, A.J.; Zelinka, C.; Gallina, D.; Scott, M.A.; Todd, L. Reactive Microglia and Macrophage Facilitate the Formation of Müller Glia-Derived Retinal Progenitors. Glia 2014, 62, 1608–1628. [Google Scholar] [CrossRef]
- Conedera, F.M.; Pousa, A.M.Q.; Mercader, N.; Tschopp, M.; Enzmann, V. Retinal Microglia Signaling Affects Müller Cell Behavior in the Zebrafish Following Laser Injury Induction. Glia 2019, 67, 1150–1166. [Google Scholar] [CrossRef]
- Lu, C.; Hyde, D.R. Cytokines IL-1β and IL-10 Are Required for Müller Glia Proliferation Following Light Damage in the Adult Zebrafish Retina. Front. Cell Dev. Biol. 2024, 12, 1406330. [Google Scholar] [CrossRef]
- Yin, M.; Chen, Z.; Ouyang, Y.; Zhang, H.; Wan, Z.; Wang, H.; Wu, W.; Yin, X. Thrombin-Induced, TNFR-Dependent miR-181c Downregulation Promotes MLL1 and NF-κB Target Gene Expression in Human Microglia. J. Neuroinflamm. 2017, 14, 132. [Google Scholar] [CrossRef]
- El-Hodiri, H.M.; Bentley, J.R.; Reske, A.G.; Taylor, O.B.; Palazzo, I.; Campbell, W.A.; Halloy, N.R.; Fischer, A.J. Heparin-Binding Epidermal Growth Factor and Fibroblast Growth Factor 2 Rescue Müller Glia-Derived Progenitor Cell Formation in Microglia- and Macrophage-Ablated Chick Retinas. Development 2023, 150, dev202070. [Google Scholar] [CrossRef]
- Zhao, X.-F.; Wan, J.; Powell, C.; Ramachandran, R.; Myers, M.G.; Goldman, D. Leptin and IL-6 Family Cytokines Synergize to Stimulate Müller Glia Reprogramming and Retina Regeneration. Cell Rep. 2014, 9, 272–284. [Google Scholar] [CrossRef] [PubMed]
- Campbell, W.A.; Fritsch-Kelleher, A.; Palazzo, I.; Hoang, T.; Blackshaw, S.; Fischer, A.J. Midkine Is Neuroprotective and Influences Glial Reactivity and the Formation of Müller Glia-Derived Progenitor Cells in Chick and Mouse Retinas. Glia 2021, 69, 1515–1539. [Google Scholar] [CrossRef]
- Palazzo, I.; Kelly, L.; Koenig, L.; Fischer, A.J. Patterns of NFkB Activation Resulting from Damage, Reactive Microglia, Cytokines, and Growth Factors in the Mouse Retina. Exp. Neurol. 2023, 359, 114233. [Google Scholar] [CrossRef] [PubMed]
- Palazzo, I.; Todd, L.J.; Hoang, T.V.; Reh, T.A.; Blackshaw, S.; Fischer, A.J. NFkB-Signaling Promotes Glial Reactivity and Suppresses Müller Glia-Mediated Neuron Regeneration in the Mammalian Retina. Glia 2022, 70, 1380–1401. [Google Scholar] [CrossRef] [PubMed]
- Campbell, L.J.; Hobgood, J.S.; Jia, M.; Boyd, P.; Hipp, R.I.; Hyde, D.R. Notch3 and DeltaB Maintain Müller Glia Quiescence and Act as Negative Regulators of Regeneration in the Light-Damaged Zebrafish Retina. Glia 2021, 69, 546–566. [Google Scholar] [CrossRef] [PubMed]
- Sahu, A.; Devi, S.; Jui, J.; Goldman, D. Notch Signaling via Hey1 and Id2b Regulates Müller Glia’s Regenerative Response to Retinal Injury. Glia 2021, 69, 2882–2898. [Google Scholar] [CrossRef]
- Campbell, L.J.; Levendusky, J.L.; Steines, S.A.; Hyde, D.R. Retinal Regeneration Requires Dynamic Notch Signaling. Neural Regen. Res. 2022, 17, 1199–1209. [Google Scholar] [CrossRef]
- Elsaeidi, F.; Macpherson, P.; Mills, E.A.; Jui, J.; Flannery, J.G.; Goldman, D. Notch Suppression Collaborates with Ascl1 and Lin28 to Unleash a Regenerative Response in Fish Retina, But Not in Mice. J. Neurosci. 2018, 38, 2246–2261. [Google Scholar] [CrossRef]
- Del Debbio, C.B.; Mir, Q.; Parameswaran, S.; Mathews, S.; Xia, X.; Zheng, L.; Neville, A.J.; Ahmad, I. Notch Signaling Activates Stem Cell Properties of Müller Glia through Transcriptional Regulation and Skp2-Mediated Degradation of p27Kip1. PLoS ONE 2016, 11, e0152025. [Google Scholar] [CrossRef]
- Hamon, A.; García-García, D.; Ail, D.; Bitard, J.; Chesneau, A.; Dalkara, D.; Locker, M.; Roger, J.E.; Perron, M. Linking YAP to Müller Glia Quiescence Exit in the Degenerative Retina. Cell Rep. 2019, 27, 1712–1725.e6. [Google Scholar] [CrossRef]
- Lourenço, R.; Brandão, A.S.; Borbinha, J.; Gorgulho, R.; Jacinto, A. Yap Regulates Müller Glia Reprogramming in Damaged Zebrafish Retinas. Front. Cell Dev. Biol. 2021, 9, 667796. [Google Scholar] [CrossRef] [PubMed]
- Osakada, F.; Ooto, S.; Akagi, T.; Mandai, M.; Akaike, A.; Takahashi, M. Wnt Signaling Promotes Regeneration in the Retina of Adult Mammals. J. Neurosci. 2007, 27, 4210–4219. [Google Scholar] [CrossRef]
- Yao, K.; Qiu, S.; Tian, L.; Snider, W.D.; Flannery, J.G.; Schaffer, D.V.; Chen, B. Wnt Regulates Proliferation and Neurogenic Potential of Müller Glial Cells via a Lin28/Let-7 miRNA-Dependent Pathway in Adult Mammalian Retinas. Cell Rep. 2016, 17, 165–178. [Google Scholar] [CrossRef] [PubMed]
- Sanges, D.; Simonte, G.; Di Vicino, U.; Romo, N.; Pinilla, I.; Nicolás, M.; Cosma, M.P. Reprogramming Müller Glia via in Vivo Cell Fusion Regenerates Murine Photoreceptors. J. Clin. Investig. 2016, 126, 3104–3116. [Google Scholar] [CrossRef]
- Todd, L.; Squires, N.; Suarez, L.; Fischer, A.J. Jak/Stat Signaling Regulates the Proliferation and Neurogenic Potential of Müller Glia-Derived Progenitor Cells in the Avian Retina. Sci. Rep. 2016, 6, 35703. [Google Scholar] [CrossRef]
- Jorstad, N.L.; Wilken, M.S.; Todd, L.; Finkbeiner, C.; Nakamura, P.; Radulovich, N.; Hooper, M.J.; Chitsazan, A.; Wilkerson, B.A.; Rieke, F.; et al. STAT Signaling Modifies Ascl1 Chromatin Binding and Limits Neural Regeneration from Muller Glia in Adult Mouse Retina. Cell Rep. 2020, 30, 2195–2208.e5. [Google Scholar] [CrossRef]
- Fischer, A.J.; Scott, M.A.; Ritchey, E.R.; Sherwood, P. Mitogen-Activated Protein Kinase-Signaling Regulates the Ability of Müller Glia to Proliferate and Protect Retinal Neurons against Excitotoxicity. Glia 2009, 57, 1538–1552. [Google Scholar] [CrossRef] [PubMed]
- Ornitz, D.M.; Itoh, N. The Fibroblast Growth Factor Signaling Pathway. Wiley Interdiscip. Rev. Dev. Biol. 2015, 4, 215–266. [Google Scholar] [CrossRef]
- Kelly, L.E.; El-Hodiri, H.M.; Crider, A.; Fischer, A.J. Protein Phosphatases Regulate the Formation of Müller Glia-Derived Progenitor Cells in the Chick Retina. Mol. Cell. Neurosci. 2024, 129, 103932. [Google Scholar] [CrossRef]
- Wan, J.; Zhao, X.-F.; Vojtek, A.; Goldman, D. Retinal Injury, Growth Factors, and Cytokines Converge on β-Catenin and pStat3 Signaling to Stimulate Retina Regeneration. Cell Rep. 2014, 9, 285–297. [Google Scholar] [CrossRef]
- Gallina, D.; Zelinka, C.; Fischer, A.J. Glucocorticoid Receptors in the Retina, Müller Glia and the Formation of Müller Glia-Derived Progenitors. Development 2014, 141, 3340–3351. [Google Scholar] [CrossRef] [PubMed]
- Todd, L. Inducing Neural Regeneration from Glia Using Proneural bHLH Transcription Factors. Adv. Exp. Med. Biol. 2023, 1415, 577–582. [Google Scholar] [CrossRef]
- Fausett, B.V.; Gumerson, J.D.; Goldman, D. The Proneural Basic Helix-Loop-Helix Gene Ascl1a Is Required for Retina Regeneration. J. Neurosci. 2008, 28, 1109–1117. [Google Scholar] [CrossRef]
- Karl, M.O.; Reh, T.A. Regenerative Medicine for Retinal Diseases: Activating Endogenous Repair Mechanisms. Trends Mol. Med. 2010, 16, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Todd, L.; Hooper, M.J.; Haugan, A.K.; Finkbeiner, C.; Jorstad, N.; Radulovich, N.; Wong, C.K.; Donaldson, P.C.; Jenkins, W.; Chen, Q.; et al. Efficient Stimulation of Retinal Regeneration from Müller Glia in Adult Mice Using Combinations of Proneural bHLH Transcription Factors. Cell Rep. 2021, 37, 109857. [Google Scholar] [CrossRef]
- Wohlschlegel, J.; Finkbeiner, C.; Hoffer, D.; Kierney, F.; Prieve, A.; Murry, A.D.; Haugan, A.K.; Ortuño-Lizarán, I.; Rieke, F.; Golden, S.A.; et al. ASCL1 Induces Neurogenesis in Human Müller Glia. Stem Cell Rep. 2023, 18, 2400–2417. [Google Scholar] [CrossRef] [PubMed]
- Torres-Machorro, A.L. Homodimeric and Heterodimeric Interactions among Vertebrate Basic Helix-Loop-Helix Transcription Factors. Int. J. Mol. Sci. 2021, 22, 12855. [Google Scholar] [CrossRef]
- Taylor, O.B.; Patel, S.P.; Hawthorn, E.C.; El-Hodiri, H.M.; Fischer, A.J. ID Factors Regulate the Ability of Müller Glia to Become Proliferating Neurogenic Progenitor-like Cells. Glia 2024, 72, 1236–1258. [Google Scholar] [CrossRef]
- Wilczynska, K.M.; Singh, S.K.; Adams, B.; Bryan, L.; Rao, R.R.; Valerie, K.; Wright, S.; Griswold-Prenner, I.; Kordula, T. Nuclear Factor I Isoforms Regulate Gene Expression during the Differentiation of Human Neural Progenitors to Astrocytes. Stem Cells 2009, 27, 1173–1181. [Google Scholar] [CrossRef]
- El-Hodiri, H.M.; Campbell, W.A.; Kelly, L.E.; Hawthorn, E.C.; Schwartz, M.; Jalligampala, A.; McCall, M.A.; Meyer, K.; Fischer, A.J. Nuclear Factor I in Neurons, Glia and during the Formation of Müller Glia-Derived Progenitor Cells in Avian, Porcine and Primate Retinas. J. Comp. Neurol. 2022, 530, 1213–1230. [Google Scholar] [CrossRef]
- Mitra, S.; Sharma, P.; Kaur, S.; Khursheed, M.A.; Gupta, S.; Ahuja, R.; Kurup, A.J.; Chaudhary, M.; Ramachandran, R. Histone Deacetylase-Mediated Müller Glia Reprogramming through Her4.1-Lin28a Axis Is Essential for Retina Regeneration in Zebrafish. iScience 2018, 7, 68–84. [Google Scholar] [CrossRef] [PubMed]
- Strahl, B.D.; Allis, C.D. The Language of Covalent Histone Modifications. Nature 2000, 403, 41–45. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zang, C.; Cui, K.; Schones, D.E.; Barski, A.; Peng, W.; Zhao, K. Genome-Wide Mapping of HATs and HDACs Reveals Distinct Functions in Active and Inactive Genes. Cell 2009, 138, 1019–1031. [Google Scholar] [CrossRef]
- Sanhueza Salas, L.F.; García-Venzor, A.; Beltramone, N.; Capurro, C.; Toiber, D.; Silberman, D.M. Metabolic Imbalance Effect on Retinal Müller Glial Cells Reprogramming Capacity: Involvement of Histone Deacetylase SIRT6. Front. Genet. 2021, 12, 769723. [Google Scholar] [CrossRef]
- Lee, J.; Lee, B.-K.; Gross, J.M. Brd Activity Regulates Müller Glia-Dependent Retinal Regeneration in Zebrafish. Glia 2023, 71, 2866–2883. [Google Scholar] [CrossRef] [PubMed]
- Ueno, K.; Iwagawa, T.; Ochiai, G.; Koso, H.; Nakauchi, H.; Nagasaki, M.; Suzuki, Y.; Watanabe, S. Analysis of Müller Glia Specific Genes and Their Histone Modification Using Hes1-Promoter Driven EGFP Expressing Mouse. Sci. Rep. 2017, 7, 3578. [Google Scholar] [CrossRef]
- Sawicka, A.; Seiser, C. Histone H3 Phosphorylation—A Versatile Chromatin Modification for Different Occasions. Biochimie 2012, 94, 2193–2201. [Google Scholar] [CrossRef]
- Coppedè, F. Mitochondrial DNA Methylation and Mitochondria-Related Epigenetics in Neurodegeneration. Neural Regen. Res. 2024, 19, 405–406. [Google Scholar] [CrossRef]
- Reyes-Aguirre, L.I.; Lamas, M. Oct4 Methylation-Mediated Silencing As an Epigenetic Barrier Preventing Müller Glia Dedifferentiation in a Murine Model of Retinal Injury. Front. Neurosci. 2016, 10, 523. [Google Scholar] [CrossRef]
- Powell, C.; Grant, A.R.; Cornblath, E.; Goldman, D. Analysis of DNA Methylation Reveals a Partial Reprogramming of the Müller Glia Genome during Retina Regeneration. Proc. Natl. Acad. Sci. USA 2013, 110, 19814–19819. [Google Scholar] [CrossRef]
- Dvoriantchikova, G.; Seemungal, R.J.; Ivanov, D. Development and Epigenetic Plasticity of Murine Müller Glia. Biochim. Biophys. Acta Mol. Cell Res. 2019, 1866, 1584–1594. [Google Scholar] [CrossRef] [PubMed]
- Seritrakul, P.; Gross, J.M. Tet-Mediated DNA Hydroxymethylation Regulates Retinal Neurogenesis by Modulating Cell-Extrinsic Signaling Pathways. PLoS Genet. 2017, 13, e1006987. [Google Scholar] [CrossRef]
- Konar, G.J.; Ferguson, C.; Flickinger, Z.; Kent, M.R.; Patton, J.G. miRNAs and Müller Glia Reprogramming During Retina Regeneration. Front. Cell Dev. Biol. 2020, 8, 632632. [Google Scholar] [CrossRef] [PubMed]
- Rajaram, K.; Harding, R.L.; Bailey, T.; Patton, J.G.; Hyde, D.R. Dynamic miRNA Expression Patterns during Retinal Regeneration in Zebrafish: Reduced Dicer or miRNA Expression Suppresses Proliferation of Müller Glia-Derived Neuronal Progenitor Cells. Dev. Dyn. 2014, 243, 1591–1605. [Google Scholar] [CrossRef]
- Wohl, S.G.; Reh, T.A. miR-124-9-9* Potentiates Ascl1-Induced Reprogramming of Cultured Müller Glia. Glia 2016, 64, 743–762. [Google Scholar] [CrossRef]
- Wohl, S.G.; Hooper, M.J.; Reh, T.A. MicroRNAs miR-25, Let-7 and miR-124 Regulate the Neurogenic Potential of Müller Glia in Mice. Development 2019, 146, dev179556. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Tao, Z.; Xue, L.; Zeng, Y.; Wang, Y.; Xu, H.; Yin, Z.Q. Lin28b Stimulates the Reprogramming of Rat Müller Glia to Retinal Progenitors. Exp. Cell Res. 2017, 352, 164–174. [Google Scholar] [CrossRef]
- Rajaram, K.; Harding, R.L.; Hyde, D.R.; Patton, J.G. miR-203 Regulates Progenitor Cell Proliferation during Adult Zebrafish Retina Regeneration. Dev. Biol. 2014, 392, 393–403. [Google Scholar] [CrossRef]
- Thummel, R.; Kassen, S.C.; Enright, J.M.; Nelson, C.M.; Montgomery, J.E.; Hyde, D.R. Characterization of Müller Glia and Neuronal Progenitors during Adult Zebrafish Retinal Regeneration. Exp. Eye Res. 2008, 87, 433–444. [Google Scholar] [CrossRef]
- Kara, N.; Kent, M.R.; Didiano, D.; Rajaram, K.; Zhao, A.; Summerbell, E.R.; Patton, J.G. The miR-216a-Dot1l Regulatory Axis Is Necessary and Sufficient for Müller Glia Reprogramming during Retina Regeneration. Cell Rep. 2019, 28, 2037–2047.e4. [Google Scholar] [CrossRef]
- Magner, E.; Sandoval-Sanchez, P.; Kramer, A.C.; Thummel, R.; Hitchcock, P.F.; Taylor, S.M. Disruption of miR-18a Alters Proliferation, Photoreceptor Replacement Kinetics, Inflammatory Signaling, and Microglia/Macrophage Numbers During Retinal Regeneration in Zebrafish. Mol. Neurobiol. 2022, 59, 2910–2931. [Google Scholar] [CrossRef]
- Si, T.-E.; Li, Z.; Zhang, J.; Su, S.; Liu, Y.; Chen, S.; Peng, G.-H.; Cao, J.; Zang, W. Epigenetic Mechanisms of Müller Glial Reprogramming Mediating Retinal Regeneration. Front. Cell Dev. Biol. 2023, 11, 1157893. [Google Scholar] [CrossRef] [PubMed]
- Ghorbani, S.; Yong, V.W. The Extracellular Matrix as Modifier of Neuroinflammation and Remyelination in Multiple Sclerosis. Brain 2021, 144, 1958–1973. [Google Scholar] [CrossRef]
- Melrose, J.; Hayes, A.J.; Bix, G. The CNS/PNS Extracellular Matrix Provides Instructive Guidance Cues to Neural Cells and Neuroregulatory Proteins in Neural Development and Repair. Int. J. Mol. Sci. 2021, 22, 5583. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Liu, M.; Xie, D.; Liu, X.; Yan, H. Role of the Extracellular Matrix and YAP/TAZ in Cell Reprogramming. Differ. Res. Biol. Divers. 2021, 122, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Campbell, W.A., IV; Deshmukh, A.; Blum, S.; Todd, L.; Mendonca, N.; Weist, J.; Zent, J.; Hoang, T.V.; Blackshaw, S.; Leight, J.; et al. Matrix-Metalloproteinase Expression and Gelatinase Activity in the Avian Retina and Their Influence on Müller Glia Proliferation. Exp. Neurol. 2019, 320, 112984. [Google Scholar] [CrossRef]
- Silva, N.J.; Nagashima, M.; Li, J.; Kakuk-Atkins, L.; Ashrafzadeh, M.; Hyde, D.R.; Hitchcock, P.F. Inflammation and Matrix Metalloproteinase 9 (Mmp-9) Regulate Photoreceptor Regeneration in Adult Zebrafish. Glia 2020, 68, 1445–1465. [Google Scholar] [CrossRef]
- Zhang, Z.; Hou, H.; Yu, S.; Zhou, C.; Zhang, X.; Li, N.; Zhang, S.; Song, K.; Lu, Y.; Liu, D.; et al. Inflammation-Induced Mammalian Target of Rapamycin Signaling Is Essential for Retina Regeneration. Glia 2020, 68, 111–127. [Google Scholar] [CrossRef]
- Vay, S.U.; Flitsch, L.J.; Rabenstein, M.; Rogall, R.; Blaschke, S.; Kleinhaus, J.; Reinert, N.; Bach, A.; Fink, G.R.; Schroeter, M.; et al. The Plasticity of Primary Microglia and Their Multifaceted Effects on Endogenous Neural Stem Cells in Vitro and in Vivo. J. Neuroinflamm. 2018, 15, 226. [Google Scholar] [CrossRef]
- Yu, C.; Roubeix, C.; Sennlaub, F.; Saban, D.R. Microglia versus Monocytes: Distinct Roles in Degenerative Diseases of the Retina. Trends Neurosci. 2020, 43, 433–449. [Google Scholar] [CrossRef]
- Gascón, S.; Murenu, E.; Masserdotti, G.; Ortega, F.; Russo, G.L.; Petrik, D.; Deshpande, A.; Heinrich, C.; Karow, M.; Robertson, S.P.; et al. Identification and Successful Negotiation of a Metabolic Checkpoint in Direct Neuronal Reprogramming. Cell Stem Cell 2016, 18, 396–409. [Google Scholar] [CrossRef] [PubMed]
- Tsogtbaatar, E.; Landin, C.; Minter-Dykhouse, K.; Folmes, C.D.L. Energy Metabolism Regulates Stem Cell Pluripotency. Front. Cell Dev. Biol. 2020, 8, 87. [Google Scholar] [CrossRef]
- Polyzos, A.A.; Lee, D.Y.; Datta, R.; Hauser, M.; Budworth, H.; Holt, A.; Mihalik, S.; Goldschmidt, P.; Frankel, K.; Trego, K.; et al. Metabolic Reprogramming in Astrocytes Distinguishes Region-Specific Neuronal Susceptibility in Huntington Mice. Cell Metab. 2019, 29, 1258–1273.e11. [Google Scholar] [CrossRef]
- Fridman, E.S.; Ginini, L.; Gil, Z. The Role of Extracellular Vesicles in Metabolic Reprogramming of the Tumor Microenvironment. Cells 2022, 11, 1433. [Google Scholar] [CrossRef] [PubMed]
- Yin, Z.; Kang, J.; Cheng, X.; Gao, H.; Huo, S.; Xu, H. Investigating Müller Glia Reprogramming in Mice: A Retrospective of the Last Decade, and a Look to the Future. Neural Regen. Res. 2025, 20, 946–959. [Google Scholar] [CrossRef] [PubMed]
- Yao, K.; Qiu, S.; Wang, Y.V.; Park, S.J.H.; Mohns, E.J.; Mehta, B.; Liu, X.; Chang, B.; Zenisek, D.; Crair, M.C.; et al. Restoration of Vision after de Novo Genesis of Rod Photoreceptors in Mammalian Retinas. Nature 2018, 560, 484–488. [Google Scholar] [CrossRef]
- Xiao, D.; Jin, K.; Qiu, S.; Lei, Q.; Huang, W.; Chen, H.; Su, J.; Xu, Q.; Xu, Z.; Gou, B.; et al. In Vivo Regeneration of Ganglion Cells for Vision Restoration in Mammalian Retinas. Front. Cell Dev. Biol. 2021, 9, 755544. [Google Scholar] [CrossRef]
- Bernardos, R.L.; Barthel, L.K.; Meyers, J.R.; Raymond, P.A. Late-Stage Neuronal Progenitors in the Retina Are Radial Müller Glia That Function as Retinal Stem Cells. J. Neurosci. 2007, 27, 7028–7040. [Google Scholar] [CrossRef]
- Jayaram, H.; Jones, M.F.; Eastlake, K.; Cottrill, P.B.; Becker, S.; Wiseman, J.; Khaw, P.T.; Limb, G.A. Transplantation of Photoreceptors Derived from Human Muller Glia Restore Rod Function in the P23H Rat. Stem Cells Transl. Med. 2014, 3, 323–333. [Google Scholar] [CrossRef]
- Eastlake, K.; Wang, W.; Jayaram, H.; Murray-Dunning, C.; Carr, A.J.F.; Ramsden, C.M.; Vugler, A.; Gore, K.; Clemo, N.; Stewart, M.; et al. Phenotypic and Functional Characterization of Müller Glia Isolated from Induced Pluripotent Stem Cell-Derived Retinal Organoids: Improvement of Retinal Ganglion Cell Function upon Transplantation. Stem Cells Transl. Med. 2019, 8, 775–784. [Google Scholar] [CrossRef]
- Eastlake, K.; Lamb, W.D.B.; Luis, J.; Khaw, P.T.; Jayaram, H.; Limb, G.A. Prospects for the Application of Müller Glia and Their Derivatives in Retinal Regenerative Therapies. Prog. Retin. Eye Res. 2021, 85, 100970. [Google Scholar] [CrossRef] [PubMed]
- Johnson, T.V.; Bull, N.D.; Martin, K.R. Neurotrophic Factor Delivery as a Protective Treatment for Glaucoma. Exp. Eye Res. 2011, 93, 196–203. [Google Scholar] [CrossRef]
- Mead, B.; Amaral, J.; Tomarev, S. Mesenchymal Stem Cell-Derived Small Extracellular Vesicles Promote Neuroprotection in Rodent Models of Glaucoma. Investig. Ophthalmol. Vis. Sci. 2018, 59, 702–714. [Google Scholar] [CrossRef]
- Clark, B.S.; Stein-O’Brien, G.L.; Shiau, F.; Cannon, G.H.; Davis-Marcisak, E.; Sherman, T.; Santiago, C.P.; Hoang, T.V.; Rajaii, F.; James-Esposito, R.E.; et al. Single-Cell RNA-Seq Analysis of Retinal Development Identifies NFI Factors as Regulating Mitotic Exit and Late-Born Cell Specification. Neuron 2019, 102, 1111–1126.e5. [Google Scholar] [CrossRef]
- Collin, J.; Queen, R.; Zerti, D.; Dorgau, B.; Hussain, R.; Coxhead, J.; Cockell, S.; Lako, M. Deconstructing Retinal Organoids: Single Cell RNA-Seq Reveals the Cellular Components of Human Pluripotent Stem Cell-Derived Retina. Stem Cells 2019, 37, 593–598. [Google Scholar] [CrossRef] [PubMed]
- Hartong, D.T.; Berson, E.L.; Dryja, T.P. Retinitis Pigmentosa. Lancet 2006, 368, 1795–1809. [Google Scholar] [CrossRef] [PubMed]
- Antonetti, D.A.; Silva, P.S.; Stitt, A.W. Current Understanding of the Molecular and Cellular Pathology of Diabetic Retinopathy. Nat. Rev. Endocrinol. 2021, 17, 195–206. [Google Scholar] [CrossRef]
- Coughlin, B.A.; Feenstra, D.J.; Mohr, S. Müller Cells and Diabetic Retinopathy. Vis. Res. 2017, 139, 93–100. [Google Scholar] [CrossRef]
- Curcio, C.A.; Medeiros, N.E.; Millican, C.L. Photoreceptor Loss in Age-Related Macular Degeneration. Investig. Ophthalmol. Vis. Sci. 1996, 37, 1236–1249. [Google Scholar]
- Zhao, N.; Hao, X.-N.; Huang, J.-M.; Song, Z.-M.; Tao, Y. Crosstalk between Microglia and Müller Glia in the Age-Related Macular Degeneration: Role and Therapeutic Value of Neuroinflammation. Aging Dis. 2024, 15, 1132–1154. [Google Scholar] [CrossRef]
- Almasieh, M.; Wilson, A.M.; Morquette, B.; Cueva Vargas, J.L.; Di Polo, A. The Molecular Basis of Retinal Ganglion Cell Death in Glaucoma. Prog. Retin. Eye Res. 2012, 31, 152–181. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Su, J.; Hu, X.; Zhou, C.; Li, H.; Chen, Z.; Xiao, Q.; Wang, B.; Wu, W.; Sun, Y.; et al. Glia-to-Neuron Conversion by CRISPR-CasRx Alleviates Symptoms of Neurological Disease in Mice. Cell 2020, 181, 590–603.e16. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; Zhu, J.; Duan, Y.; Li, G.; Cai, H.; Zheng, L.; Qian, H.; Zhang, C.; Jin, Z.; Fu, X.-D.; et al. Visual Function Restoration in Genetically Blind Mice via Endogenous Cellular Reprogramming. bioRxiv 2020. [Google Scholar] [CrossRef]
- Xie, Y.; Zhou, J.; Chen, B. Critical Examination of Ptbp1-Mediated Glia-to-Neuron Conversion in the Mouse Retina. Cell Rep. 2022, 39, 110960. [Google Scholar] [CrossRef]
- Xie, Y.; Zhou, J.; Wang, L.-L.; Zhang, C.-L.; Chen, B. New AAV Tools Fail to Detect Neurod1-Mediated Neuronal Conversion of Müller Glia and Astrocytes in Vivo. EBioMedicine 2023, 90, 104531. [Google Scholar] [CrossRef]
- Hoang, T.; Kim, D.W.; Appel, H.; Pannullo, N.A.; Leavey, P.; Ozawa, M.; Zheng, S.; Yu, M.; Peachey, N.S.; Blackshaw, S. Genetic Loss of Function of Ptbp1 Does Not Induce Glia-to-Neuron Conversion in Retina. Cell Rep. 2022, 39, 110849. [Google Scholar] [CrossRef]
- Chen, G. In Vivo Confusion over in Vivo Conversion. Mol. Ther. 2021, 29, 3097–3098. [Google Scholar] [CrossRef]
- Blackshaw, S.; Sanes, J.R. Turning Lead into Gold: Reprogramming Retinal Cells to Cure Blindness. J. Clin. Investig. 2021, 131, e146134. [Google Scholar] [CrossRef]
- Grabinski, S.E.; Parsana, D.; Perkins, B.D. Comparative Analysis of Transcriptional Changes in Zebrafish Cep290 and Bbs2 Mutants by RNA-Seq Reveals Upregulation of Inflammatory and Stress-Related Pathways. Front. Mol. Neurosci. 2023, 16, 1148840. [Google Scholar] [CrossRef]
- Parain, K.; Chesneau, A.; Locker, M.; Borday, C.; Perron, M. Regeneration from Three Cellular Sources and Ectopic Mini-Retina Formation upon Neurotoxic Retinal Degeneration in Xenopus. Glia 2024, 72, 759–776. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, Y.; Qin, S.; Wu, H. Advancements in Müller Glia Reprogramming: Pioneering Approaches for Retinal Neuron Regeneration. Neuroglia 2025, 6, 20. https://doi.org/10.3390/neuroglia6020020
Zhou Y, Qin S, Wu H. Advancements in Müller Glia Reprogramming: Pioneering Approaches for Retinal Neuron Regeneration. Neuroglia. 2025; 6(2):20. https://doi.org/10.3390/neuroglia6020020
Chicago/Turabian StyleZhou, Yuyan, Song Qin, and Haixiang Wu. 2025. "Advancements in Müller Glia Reprogramming: Pioneering Approaches for Retinal Neuron Regeneration" Neuroglia 6, no. 2: 20. https://doi.org/10.3390/neuroglia6020020
APA StyleZhou, Y., Qin, S., & Wu, H. (2025). Advancements in Müller Glia Reprogramming: Pioneering Approaches for Retinal Neuron Regeneration. Neuroglia, 6(2), 20. https://doi.org/10.3390/neuroglia6020020