Abstract
The almost complete eradication of fire from grasslands in North America has led to non-linear hysteretic transitions to shrub- and woodlands that the reintroduction of low-intensity fire is unable to reverse. We explore the ability of the extreme ends of variation in fire behavior to help overcome hysteretic threshold behaviors in huisache (Vachellia farnesiana) encroached grasslands. We contrasted experimental fire treatments with unburned control areas to assess the ability of extreme fires burned during drought to alter the density and structure of huisache. We found that extreme fires reduced the density of huisache by over 30% compared to control plots, both through driving huisache mortality and reducing the number of new recruits following treatments. For instance, extreme fire drove 48% huisache mortality compared to 4% in control treatments. For surviving plants, the number of stems increased but the crown area did not significantly change. Prescribed fire, conducted under the right conditions, can drive high mortality in one of the most notorious encroaching species in the southern U.S. Great Plains. With the fire conditions observed in this study likely to increase under future climate projections, utilizing extreme fire as a management tool for huisache will help scale up management to meet the growing extent of woody encroachment into grasslands.
1. Introduction
Woody encroachment is a predominant threat to grassland systems across the globe, leading to herbaceous biomass loss and declines in grassland biodiversity [1,2,3]. These transitions are linked in many regions to fire suppression, which has removed frequent low-intensity fires that previously characterized many of the world’s grasslands, allowing woody seedlings to establish more readily [4,5]. Once established, woody vegetation promotes a positive feedback that further restricts fire by reducing herbaceous fuel loads and increasing fuel moisture [1,5,6]. Thus, woody encroachment into grasslands is often used as an example of ecosystem threshold dynamics, where abrupt transitions from a grassland to a shrub- or woodland represent non-linear hysteretic (the path into an alternative state is not the same as the path out; [7]) or irreversible transitions [1,8].
Management aimed at reintroducing low-intensity fires into woody-encroached areas has generally been unable to shift the system back towards its grassland state [5,8,9,10,11]. Resprouting woody species have below-ground meristematic tissues that are well insulated from fire; they also frequently have large carbon reserves that enable rapid new growth following low-intensity fire [12]. Studies focused on the reintroduction of low-intensity fire into woody-encroached grasslands have often found little to no impact of fire on the density of woody resprouting plants [13,14,15,16]. In some cases, low-intensity fire can actually increase the number of stems on individual plants and increase individual crown area [13,17,18].
Previous research has shown that the use of extreme fire (fires that exhibit rapid and erratic changes in fire behavior that include fire whirls and sudden blow-ups) can drive significant woody mortality [19] and may be able to overcome hysteresis in woody-encroached systems, driving a shift back towards a grassland state [20]. In the southern Great Plains, Twidwell et al. [21] found high levels of resprouting shrub mortality following extreme fire treatments when assessing the impacts of extreme fire on shrub communities. Critically, the responses of shrubs to extreme fire varied among the individual species within the study [21]. Since the presence and dominance of resprouting shrub species vary in different grasslands worldwide, assessing species-specific responses to extreme fire is important to accurately determine the potential for extreme fire to restore encroached grasslands at local scales.
In this study, we conduct a more in-depth assessment of the Twidwell et al. [21] extreme fire experiment, specifically evaluating huisache-specific responses. Huisache (Vachellia farnesiana, syn. Acacia farnesiana) is a particularly invasive woody species in the southern Great Plains, and its rapid encroachment and resprouting following disturbance makes it challenging to manage—particularly with low-intensity fires. Here, we assess the impacts of extreme fire on (1) huisache density relative to plant mortality and the number of new recruits and (2) huisache structure as a function of the number of plant stems and canopy cover. We predict that high-intensity fire will drive higher huisache mortality by killing root crowns while also reducing the huisache seed bank and, thus, the number of new recruits. Structurally, we predict that extreme fire will decrease the number of individual stems per plant and reduce the surviving crown area due to its potential to damage below-ground meristematic tissue.
2. Materials and Methods
2.1. Focal Species
Huisache (Vachellia farnesiana, syn. Acacia farnesiana) is a common woody encroacher that occurs or has invaded multiple continents, including the Americas [22,23], Australia [24,25], Asia [26], and Africa [27,28]. Huisache is a rapid encroacher; for instance, its canopy cover in an invaded area in Texas increased by 48% from 1964 to 1982 [29,30]. Its current range comprises the Gulf Coast from Texas to Florida, where it has encroached an estimated 2.7 million acres [31]. Huisache is concerning from both an economic and ecological perspective. It reduces forage production in large cattle-producing rangelands of the South Texas Plains [32], reduces native herbaceous species diversity, degrades grassland wildlife habitat, reduces plant available soil moisture, and contributes to fence degradation [33].
Low-intensity fire has negligible impacts on huisache density and undesired impacts on plant structure that can potentially decrease herbaceous productivity in encroached areas. Huisache density depends on both fire-driven mortality and the occurrence of new recruits following fire. While low-intensity fire can topkill huisache, mortality is rare; huisache often rapidly resprouts from surviving root crowns [22,32,34,35,36]. Seeds also typically survive low-intensity fire, and seed dormancy can be broken by fire scarification [25,37]; thus, recruitment is likely uninhibited and potentially stimulated by low-intensity fire.
Along with resprouter density, changes in shrub structure can reduce herbaceous production and promote positive feedbacks associated with woody encroachment [32]. Like other resprouters, low-intensity fire has also been shown to greatly increase the number of huisache stems per plant by breaking apical dominance [32,35,36], which increases plant stem density and decreases subsequent fire management potential. Low-intensity fire has been shown to reduce canopy cover one year post-fire [34]; however, canopies are rapidly replaced [32]. In contrast, the species-specific summary of huisache’s response to fire reported by Twidwell et al. [21] highlighted high levels of mortality from extreme fire treatments, but did not conduct an in-depth assessment of morphological and reproductive outcomes of extreme fire specific to huisache.
2.2. Study Area and Design
Experiments using extreme fire application were conducted in the uplands of the Western Gulf Coastal Plain ecoregion at the Rob and Bessie Welder Wildlife Refuge, located 11 km North of Sinton, Texas (Figure 1). This area was once dominated by coastal tallgrass prairie species, but due to a lack of fire, it is now co-dominated by the thorn scrub shrub species huisache and mesquite (Prosopis glandulosa) [38]. The mean annual precipitation in the area is 900 mm [21]. Historically, wildfires typically occurred during the growing season; however, recent increases in wildfire numbers and total hectares burned are associated with a more consistent number of wildfires throughout the year [39].
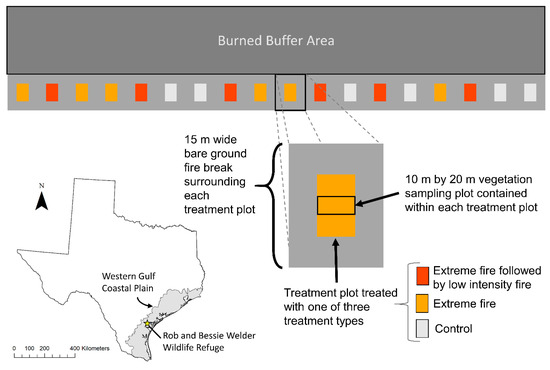
Figure 1.
The experimental design used to assess the impacts of extreme fire and extreme fire followed by low-intensity fire on huisache (Vachellia farnesiana) density and structure in encroached grasslands in the Rob and Bessie Welder Wildlife Refuge, Texas, U.S.
Eighteen 30 m × 20 m experimental plots were distributed across a pasture with a high density of resprouting shrubs, no downwind fire hazards, and sufficient space to create fire breaks wide enough to contain extreme fire (Figure 1). Each plot was randomly assigned to one of three treatments: (1) extreme fire treatment (n = 6), (2) extreme fire treatment followed by low-intensity fire (n = 6), and (3) unburned control sites (n = 6; Figure 1). Maximum woody plant canopy cover was 75%, while surface herbaceous cover ranged from 60 to 100%. Fire treatments were conducted using special exemptions during government-imposed burn restrictions when extremely low fuel moisture was able to facilitate extreme fire behavior. Each plot was burned independently with a flank–head fire.
2.3. Treatment Application
Extreme fire treatments were applied to 12 plots in June 2008, when total precipitation was 96% below the historical monthly mean, based on nearby weather station estimates from Sinton, TX. The second low-intensity fire treatment was applied in July 2009 to 6 plots, 1 year after the initial extreme fire treatments. See Twidwell et al. [21] for more detailed drought information during experiments. Fireline intensities (I) during treatments were calculated using visual estimations of flame length (FL) along a 10 m transect within each subplot, following the methods of Rothermel and Deeming [40] using the equation from Byram [41] and Alexander [42]:
Temperatures were estimated using ceramic tile pyrometers with 25 temperature-indicating lacquers that melted at a range of 79 to 1038 °C (OMEGALAQ® Liquid Temperature Lacquers; DwyerOmega, 102 Indiana Hwy. 212, Michigan City, IN, USA). Extreme fire behavior (fire whirls and sudden blow-ups) was recorded during the first application of fires across multiple treatment plots. Fireline intensity in the extreme fire treatments in 2008 reached up to 8595 kW m−1, while fire temperatures reached at least 1038 °C. Low-intensity fire treatments conducted in 2009 had maximum fire line intensities of 58 kW m−1 and a peak recorded temperature of 760 °C.
2.4. Vegetation Measurements
Huisache shrubs exceeding 100 cm in height were identified in each experimental plot in 2006 before extreme fire application. Each shrub was measured and tagged with a metal flag identifying each unique individual to track individual response to fire. Shrub survival was tracked annually until 2011. Shrub measurements were repeated in 2011—3 years after the initial extreme fire treatments to ensure sufficient time to assess mortality. Counts of shrubs per plot were used to calculate huisache density before and after fire treatments. Metal tags were used to identify shrubs to determine the impacts of extreme fire on huisache mortality, as well as to identify new recruits that occurred following treatments. Shrub mortality was assumed if a shrub had been fully top killed and showed no evidence of resprouting 3 years post-extreme fire treatment. New recruits were identified as individuals that were greater than 100 cm in height and were recorded in 2011 but were not recorded in 2006. The number of stems per plant, along with crown area, were measured before and after treatments for each individual shrub to determine the impact of fire treatments on shrub structure.
2.5. Analysis
Shrub density and the number of new recruits were assessed per plot using Type II ANOVA and post hoc Tukey Honest Significant Difference test [43] to determine differences among treatments. Shrub mortality along with shrub structural changes in crown area and number of stems were assessed per plant using a generalized linear mixed effect model and linear mixed effect models, respectively, with plot number included as a random effect [44]. All statistical analyses were conducted in R version 4.0.4 [45].
3. Results
3.1. Huisache Density
There was a negative percent change in huisache density on average in both extreme fire treatments, while untreated control plots increased in huisache density (Table 1; Figure 2). Huisache density increased by an average of 21% ± 6.2 SE in control plots between 2006 and 2011, shifting from a mean of 580 plants per hectare to 717 plants per hectare. In contrast, extreme fire and extreme fire treatments followed by low-intensity fire had an average decrease in huisache density of 1% ± 7.5 SE and 8% ± 8.2 SE, respectively, relative to what occurred 5 years prior. There was no significant difference in the change in huisache density between plots treated with extreme fire and plots treated with extreme fire followed by low-intensity fire (Table 1).

Table 1.
The modeled response in the percent change in huisache (Vachellia farnesiana) density, huisache plant mortality, and huisache new recruits among plots treated with extreme fire treatments and control plots.
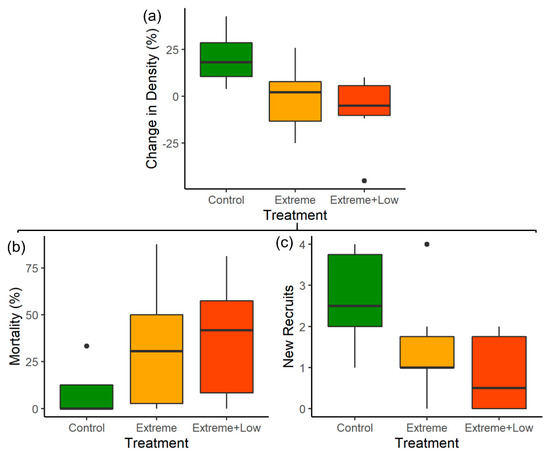
Figure 2.
A comparison of the (a) percent change in huisache (Vachellia farnesiana) density, (b) huisache percent mortality, and (c) number of new huisache recruits across control plots (unburned), plots treated with extreme fire, and plots treated with extreme fire followed by low-intensity fire.
Huisache mortality was significantly higher in both extreme fire treatments compared to control plots (Table 1; Figure 2), with 48% huisache mortality (32 of 67) compared to 4% in control treatments (2 of 55). Though there was high variability among plots, there was an average mortality of 9% ± 5.9 SE in control plots, compared to 33% ± 14.4 SE and 37% ± 13.4 SE in plots treated with extreme fire and those treated with extreme fire followed by low-intensity fire, respectively. There were no significant differences in percent mortality between extreme fire treatment types (Table 1).
Similarly, extreme fire treatments typically had fewer new recruits than control plots, though extreme fire treatments followed by low-intensity fire were the most effective at reducing new recruits (Table 1; Figure 2). There was an average of 3 ± 0.5 SE new recruits per plot in the control treatment, compared to 2 ± 0.5 SE new recruits in plots treated with extreme fire and 1 ± 0.5 SE new recruits in plots treated with extreme fire followed by low-intensity fire.
3.2. Structure
While there was a high level of variability among plants, plants that survived extreme fire treatments had significantly greater stem density (Table 2; Figure 3). The maximum percent increase in the number of stems ranged from 200% in control plots to 900% in plots treated with extreme fire followed by low-intensity fire. The change in stem number per plant was over 150% greater in plots treated with extreme fire compared to control plots (Table 2).
Table 2.
The modeled response in the percent change in the number of stems per huisache (Vachellia farnesiana) plant and huisache plant crown area among plots treated with extreme fire treatments and control plots.
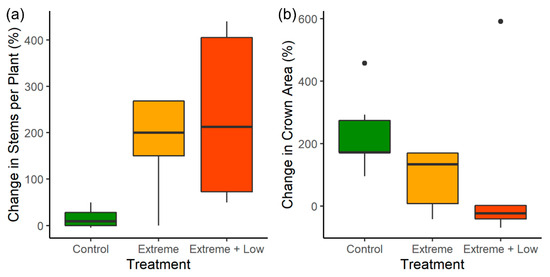
Figure 3.
A comparison of the average (a) percent change in the number of stems per huisache (Vachellia farnesiana) plant and (b) percent change in huisache plant crown area per plot for control plots (unburned), plots treated with extreme fire (Extreme), and plots treated with extreme fire followed by low-intensity fire (Extreme + Low).
Three years post-fire, extreme treatments had not resulted in any significant change in huisache plant crown area across plots in surviving plants (Table 2; Figure 3). Like huisache stem number, there was a high level of variability in the percentage change in crown area among plants. In control plots, the percentage change in crown area ranged from a 97% decrease to a 1610% increase. In comparison, plots treated with extreme fire ranged from an 85% decrease to a 650% increase in crown area and plots treated with extreme fire followed by low-intensity fire ranged from a 92% decrease to a 1200% increase in crown area.
4. Discussion
Unlike the negligible effects recorded in studies assessing the impacts of low-intensity fire treatments on huisache [34,35,36], extreme fire during drought reduced huisache density by killing over a third of huisache plants on average and reducing huisache recruitment. This finding highlights the need for management to move beyond the burned-unburned perspective of fire, particularly when dealing with hysteretic threshold dynamics. Surface fire intensity is associated with woody plant mortality thresholds [46], and modest fluctuations in both wind speed and relative humidity during a prescribed burn affect surface fire intensity and heat transfer, thereby affecting mortality and resprouting responses [47]. High-intensity fire can overcome thresholds in grasslands that have shifted to a woody-encroached state [20]. Moreover, extreme fire, even during drought, has been found to either increase or have no influence on grassland species richness, cover, and biomass [20,38,48]. Based on our results, it is possible that with continued application of extreme fires during drought, thresholds associated with huisache invasion could be overcome.
Extreme fire was able to control huisache density by increasing adult mortality and reducing recruitment. While some mortality occurred in control plots, possibly from intense drought conditions within the study period [49], we found significantly higher mortality in plots treated with extreme fire. High levels of soil heating caused by fire can drive shrub mortality [50]. Twidwell et al. [21] found that extreme fires were able to drive high levels of mortality in resprouting shrub communities. We find this is consistent with the species-specific response seen in huisache. Recruitment was also lower in plots treated with extreme fire. While huisache seeds are generally resistant to low-intensity fire and are typically able to survive in the soil seed bank [25], higher soil heating from fire has been correlated with a decreased shrub soil seed bank [50], suggesting that higher levels of soil heating in extreme fire treatments may have increased seed mortality. In addition, huisache seeds are produced in large numbers during the growing season [51], which may have been mitigated by the growing season fire treatments applied during this study. Seed pods attached to plants are likely more vulnerable to fire, though this remains understudied [25].
Extreme fire treatments did not largely alter the structural outcomes in huisache observed among low-intensity treatments in other studies. Low-intensity fire has been found to greatly increase the number of huisache stems per plant [32,35,36]. While we found high variability in the change in stem number per plant within treatments, plants that experienced extreme fire and survived generally saw a large increase in the number of stems. A higher density of stems can reinforce feedbacks that encourage shrub encroachment by decreasing herbaceous surface fuels and increasing moisture below crowns that helps prevent damage from fire [17]. However, we found that low-intensity fires were able to carry through our plots treated with extreme fire one year post-fire and were associated, on average, with slightly lower levels of new recruits, higher levels of mortality, and lower huisache density. When assessed across shrub communities, crown area has been shown to decrease following extreme fire [21]. However, we found that extreme fire did not largely impact the crown area of huisache. Combined treatments of fire and grazing have been shown to better reduce canopy area in other resprouting shrub species [11]. Investigations into the impacts of combined grazing and extreme fire treatments are needed.
Enhancing the range of conditions under which we apply fire to landscapes displaying hysteretic dynamics from woody encroachment will help improve our ability to reverse unwanted ecological transitions. Current limitations in environmental conditions, such as moisture and wind speed during the application of prescribed fire, preclude fire practitioners or grassland restorationists from targeting the conditions observed in this study, limiting fire as a management tool in the face of novel change [52,53]. This represents a substantial change in the occurrence of fire on the landscape; that is, the occurrence of high-intensity fires during severe drought may play a more important role than previously thought in regulating some resprouting shrubs and trees, like huisache. Fire is more economical and a less risky grassland management tool than alternative methods like mechanical removal or herbicide application [54,55], particularly over the large scales and rates at which woody encroachment is occuring in grassland areas [56]. While other approaches, such as fuel loading, can be used to promote extreme fire conditions [20], the conditions observed in this study are likely to occur more frequently in the future based on future climate projections [57]. With increasing wildfires occurring under drought conditions [39,48] and the high mortality of huisache observed, grassland restoration objectives may be more likely to be achieved by following the occurrence of wildfires during drought with a prescribed burning regime to further limit recruitment. This, coupled with targeted management of surviving individuals, may provide a more opportunistic, cost-effective approach in some cases.
We demonstrate the ability of fires that burn outside of typical prescribed burn conditions to help control a serial invader that occurs across many of the world’s continents. Future research should continue to investigate the ability of extreme fire to control other encroaching woody species and assess whether there is regional variation in huisache response to extreme fire. Extreme fire treatments increased native forb species richness and had no effect on non-native herbaceous plants in our study system [38]; however, assessments of the effects of extreme prescribed burns on soil and other ecosystem properties are warranted, particularly across different ecosystem types. Fire in grassland fuels typically generates low amounts of soil heating below the soil surface [58]; however, the substantial woody canopy can create patches of high heat input to the soil due to the longer residence time of woody debris that has fallen to the soil surface and continues to smolder [50]. Conducting extreme fire during drought may mitigate some soil heating effects as dry soil has slower heat transfer than wet soil [59]. Studies could investigate and contrast the ecological impacts of extreme fire behaviors generated through means other than drought, such as fuel loading, to determine the role of drought in shaping the ecological outcomes observed in our assessment.
Author Contributions
Conceptualization, V.M.D., D.T., W.E.R. and U.P.K.; methodology, D.T. and V.M.D.; formal analysis, V.M.D.; investigation, D.T.; writing—original draft preparation, V.M.D., A.V.S. and J.W.; writing—review and editing, V.M.D., C.L.W., D.A.M., A.V.S., J.W., D.T. and W.E.R.; funding acquisition, W.E.R. and U.P.K. All authors have read and agreed to the published version of the manuscript.
Funding
Funding was provided by the University of Florida Institute of Food and Agricultural Sciences, University of Nebraska-Lincoln, USDA Natural Resource Conservation Service Conservation Innovation Grant 68-3A75-5-180, USDA-CSREES National Needs Fellowship 2009-38420-05631, Rob and Bessie Welder Wildlife Foundation WWF #711, Tom Slick Foundation, and Texas A&M AgriLife Research.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Ratajczak, Z.; Nippert, J.B.; Ocheltree, T.W. Abrupt Transition of Mesic Grassland to Shrubland: Evidence for Thresholds, Alternative Attractors, and Regime Shifts. Ecology 2014, 95, 2633–2645. [Google Scholar] [CrossRef]
- Stevens, N.; Lehmann, C.E.R.; Murphy, B.P.; Durigan, G. Savanna Woody Encroachment Is Widespread across Three Continents. Glob. Change Biol. 2017, 23, 235–244. [Google Scholar] [CrossRef]
- Twidwell, D.; Rogers, W.E.; Fuhlendorf, S.D.; Wonkka, C.L.; Engle, D.M.; Weir, J.R.; Kreuter, U.P.; Taylor, C.A. The Rising Great Plains Fire Campaign: Citizens’ Response to Woody Plant Encroachment. Front. Ecol. Environ. 2013, 11, e64–e71. [Google Scholar] [CrossRef]
- Ratajczak, Z.; Nippert, J.B.; Briggs, J.M.; Blair, J.M. Fire Dynamics Distinguish Grasslands, Shrublands and Woodlands as Alternative Attractors in the Central Great Plains of North America. J. Ecol. 2014, 102, 1374–1385. [Google Scholar] [CrossRef]
- D’Odorico, P.; Okin, G.S.; Bestelmeyer, B.T. A Synthetic Review of Feedbacks and Drivers of Shrub Encroachment in Arid Grasslands. Ecohydrology 2012, 5, 520–530. [Google Scholar] [CrossRef]
- Ratajczak, Z.; Nippert, J.B.; Hartman, J.C.; Ocheltree, T.W. Positive Feedbacks Amplify Rates of Woody Encroachment in Mesic Tallgrass Prairie. Ecosphere 2011, 2, art121. [Google Scholar] [CrossRef]
- Angeler, D.G.; Allen, C.R. Quantifying Resilience. J. Appl. Ecol. 2016, 53, 617–624. [Google Scholar] [CrossRef]
- Collins, S.L.; Nippert, J.B.; Blair, J.M.; Briggs, J.M.; Blackmore, P.; Ratajczak, Z. Fire Frequency, State Change and Hysteresis in Tallgrass Prairie. Ecol. Lett. 2021, 24, 636–647. [Google Scholar] [CrossRef]
- Ansley, R.J.; Wiedemann, H.T. Reversing the Woodland Steady State: Vegetation Responses During Restoration of Juniperus-Dominated Grasslands with Chaining and Fire. In Western North American Juniperus Communities: A Dynamic Vegetation Type; Van Auken, O.W., Ed.; Ecological Studies; Springer: New York, NY, USA, 2008; pp. 272–290. ISBN 978-0-387-34003-6. [Google Scholar]
- Briggs, J.M.; Knapp, A.K.; Brock, B.L. Expansion of Woody Plants in Tallgrass Prairie: A Fifteen-Year Study of Fire and Fire-Grazing Interactions. Am. Midl. Nat. 2002, 147, 287–294. [Google Scholar] [CrossRef]
- O’Connor, R.C.; Taylor, J.H.; Nippert, J.B. Browsing and Fire Decreases Dominance of a Resprouting Shrub in Woody Encroached Grassland. Ecology 2020, 101, e02935. [Google Scholar] [CrossRef]
- Clarke, P.J.; Lawes, M.J.; Midgley, J.J.; Lamont, B.B.; Ojeda, F.; Burrows, G.E.; Enright, N.J.; Knox, K.J.E. Resprouting as a Key Functional Trait: How Buds, Protection and Resources Drive Persistence after Fire. New Phytol. 2013, 197, 19–35. [Google Scholar] [CrossRef]
- Briggs, J.M.; Knapp, A.K.; Blair, J.M.; Heisler, J.L.; Hoch, G.A.; Lett, M.S.; McCarron, J.K. An Ecosystem in Transition: Causes and Consequences of the Conversion of Mesic Grassland to Shrubland. BioScience 2005, 55, 243–254. [Google Scholar] [CrossRef]
- Canadell, J.; Lloret, F.; López-Soria, L. Resprouting Vigour of Two Mediterranean Shrub Species after Experimental Fire Treatments. Vegetatio 1991, 95, 119–126. [Google Scholar] [CrossRef]
- Drewa, P.B. Effects of Fire Season and Intensity on Prosopis glandulosa Torr. var. glandulosa. Int. J. Wildland Fire 2003, 12, 147–157. [Google Scholar] [CrossRef]
- Miller, J.E.D.; Damschen, E.I.; Ratajczak, Z.; Özdoğan, M. Holding the Line: Three Decades of Prescribed Fires Halt but Do Not Reverse Woody Encroachment in Grasslands. Landsc. Ecol. 2017, 32, 2297–2310. [Google Scholar] [CrossRef]
- Eldridge, D.J.; Bowker, M.A.; Maestre, F.T.; Roger, E.; Reynolds, J.F.; Whitford, W.G. Impacts of Shrub Encroachment on Ecosystem Structure and Functioning: Towards a Global Synthesis. Ecol. Lett. 2011, 14, 709–722. [Google Scholar] [CrossRef]
- Enslin, B.W.; Potgieter, A.L.F.; Biggs, H.C.; Biggs, R. Long Term Effects of Fire Frequency and Season on the Woody Vegetation Dynamics of the Sclerocarya birrea/Acacia nigrescens Savanna of the Kruger National Park. Koedoe 2000, 43, 27–37. [Google Scholar] [CrossRef]
- Scholtz, R.; Donovan, V.M.; Strydom, T.; Wonkka, C.; Kreuter, U.P.; Rogers, W.E.; Taylor, C.; Smit, I.P.; Govender, N.; Trollope, W.; et al. High-Intensity Fire Experiments to Manage Shrub Encroachment: Lessons Learned in South Africa and the United States. Afr. J. Range Forage Sci. 2022, 39, 148–159. [Google Scholar] [CrossRef]
- Bielski, C.H.; Scholtz, R.; Donovan, V.M.; Allen, C.R.; Twidwell, D. Overcoming an “Irreversible” Threshold: A 15-Year Fire Experiment. J. Environ. Manag. 2021, 291, 112550. [Google Scholar] [CrossRef]
- Twidwell, D.; Rogers, W.E.; Wonkka, C.L.; Taylor, C.A., Jr.; Kreuter, U.P. Extreme Prescribed Fire during Drought Reduces Survival and Density of Woody Resprouters. J. Appl. Ecol. 2016, 53, 1585–1596. [Google Scholar] [CrossRef]
- Fulbright, T.E.; Dacy, E.C.; Drawe, D.L. Does Browsing Reduce Shrub Survival and Vigor Following Summer Fires? Acta Oecologica 2011, 37, 10–15. [Google Scholar] [CrossRef]
- Watson, P.A.; Alexander, H.D.; Moczygemba, J.D. Coastal Prairie Recovery in Response to Shrub Removal Method and Degree of Shrub Encroachment. Rangel. Ecol. Manag. 2019, 72, 275–282. [Google Scholar] [CrossRef]
- Burrows, W.H.; Carter, J.O.; Scanlan, J.C.; Anderson, E.R. Management of Savannas for Livestock Production in North-East Australia: Contrasts Across the Tree-Grass Continuum. J. Biogeogr. 1990, 17, 503–512. [Google Scholar] [CrossRef]
- Erkovan, H.I.; Clarke, P.J.; Whalley, R.D.B.; Erkovan, H.I.; Clarke, P.J.; Whalley, R.D.B. Seed Bank Dynamics of Acacia farnesiana (L.) Willd. and Its Encroachment Potential in Sub-Humid Grasslands of Eastern Australia. Rangel. J. 2013, 35, 427–433. [Google Scholar] [CrossRef]
- Sekar, K.C. Invasive Alien Plants of Indian Himalayan Region—Diversity and Implication. Am. J. Plant Sci. 2012, 3, 177–184. [Google Scholar] [CrossRef]
- Arévalo, J.R.; Afonso, L.; Naranjo, A.; Salas, M. Invasion of the Gran Canaria Ravines Ecosystems (Canary Islands) by the Exotic Species Acacia farnesiana. Plant Ecol. 2009, 206, 185. [Google Scholar] [CrossRef]
- Cheek, M.D.; Boon, R.G.C. Vachellia farnesiana (L.) Wight & Arn., a Potentially Invasive Tree in KwaZulu-Natal, South Africa. S. Afr. J. Bot. 2019, 124, 387–390. [Google Scholar] [CrossRef]
- Smith, H.N.; Rechenthin, C.A. Grassland Restoration Part 1: The Texas Brush Problem; U.S. Department of Agriculture Soil Conservation Service: Temple, TX, USA, 1964. [Google Scholar]
- U.S. Department of Agriculture Soil Conservation Service (USDA). Fact Sheet: Range Management—Brush Management; U.S. Department of Agriculture Soil Conservation Service (USDA): Temple, TX, USA, 1985. [Google Scholar]
- Clayton, M.K.; Lyons, R.K.; McGinty, J.A. Huisache Ecology and Management; Texas A&M Agrilife Extension Bulletin: College Station, TX, USA, 2014. [Google Scholar]
- Scifres, C.J.; Mutz, J.L.; Drawe, D.L. Ecology and Management of Huisache on the Texas Coastal Prairie; Texas FARMER Collection; Texas A&M Agrilife Extension Bulletin: College Station, TX, USA, 1982. [Google Scholar]
- Allred, B.W. Distribution and Control of Several Woody Plants in Texas and Oklahoma. J. Range Manag. 1948, 1, 17–27. [Google Scholar] [CrossRef]
- Box, T.W.; White, R.S. Fall and Winter Burning of South Texas Brush Ranges. J. Range Manag. 1969, 22, 373–376. [Google Scholar] [CrossRef]
- Dacy, E.C.; Fulbright, T.E. Survival of Sprouting Shrubs Following Summer Fire: Effects of Morphological and Spatial Characteristics. Rangel. Ecol. Manag. 2009, 62, 179–185. [Google Scholar] [CrossRef]
- Rasmussen, G.; Scifres, C.; Drawe, D. Huisache Growth, Browse Quality, and Use Following Burning Acacia farnesiana, Texas, Nutritional Status. Rangel. Ecol. Manag./J. Range Manag. Arch. 1983, 36, 337–342. [Google Scholar]
- Scifres, C.J. Salient Aspects of Huisache Seed Germination. Southwest. Nat. 1974, 18, 383–391. [Google Scholar] [CrossRef]
- Twidwell, D.; Rogers, W.E.; McMahon, E.A.; Thomas, B.R.; Kreuter, U.P.; Blankenship, T.L. Prescribed Extreme Fire Effects on Richness and Invasion in Coastal Prairie. Invasive Plant Sci. Manag. 2012, 5, 330–340. [Google Scholar] [CrossRef]
- Donovan, V.M.; Wonkka, C.L.; Twidwell, D. Surging Wildfire Activity in a Grassland Biome. Geophys. Res. Lett. 2017, 44, 5986–5993. [Google Scholar] [CrossRef]
- Rothermel, R.C.; Deeming, J.E. Measuring and Interpreting Fire Behavior for Correlation with Fire Effects; US Department of Agriculture, Forest Service: Washington, DC, USA, 1980; p. 9. [Google Scholar]
- Byram, G.M. Combustion of Forest Fuels. In Forest Fire: Control and Use; Davis, K.P., Ed.; McGraw-Hill: New York, NY, USA, 1959; pp. 61–89. [Google Scholar]
- Alexander, M.E. Calculating and Interpreting Forest Fire Intensities. Can. J. Bot. 1982, 60, 349–357. [Google Scholar] [CrossRef]
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous Inference in General Parametric Models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using Lme4. J. Stat. Softw. 2014, 67, 1–48. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Twidwell, D.; Fuhlendorf, S.D.; Taylor, C.A.; Rogers, W.E. Refining Thresholds in Coupled Fire–Vegetation Models to Improve Management of Encroaching Woody Plants in Grasslands. J. Appl. Ecol. 2013, 50, 603–613. [Google Scholar] [CrossRef]
- Starns, H.D.; Wonkka, C.L.; Dickinson, M.B.; Lodge, A.G.; Treadwell, M.L.; Kavanagh, K.L.; Tolleson, D.R.; Twidwell, D.; Rogers, W.E. Prosopis glandulosa Persistence Is Facilitated by Differential Protection of Buds during Low- and High-Energy Fires. J. Environ. Manag. 2022, 303, 114141. [Google Scholar] [CrossRef]
- Donovan, V.M.; Twidwell, D.; Uden, D.R.; Tadesse, T.; Wardlow, B.D.; Bielski, C.H.; Jones, M.O.; Allred, B.W.; Naugle, D.E.; Allen, C.R. Resilience to Large, “Catastrophic” Wildfires in North America’s Grassland Biome. Earth’s Future 2020, 8, e2020EF001487. [Google Scholar] [CrossRef]
- Johnson, D.M.; Domec, J.-C.; Berry, Z.C.; Schwantes, A.M.; McCulloh, K.A.; Woodruff, D.R.; Polley, H.W.; Wortemann, R.; Swenson, J.J.; Mackay, D.S.; et al. Co-Occurring Woody Species Have Diverse Hydraulic Strategies and Mortality Rates during an Extreme Drought. Plant Cell Environ. 2018, 41, 576–588. [Google Scholar] [CrossRef]
- Odion, D.C.; Davis, F.W. Fire, Soil Heating, and the Formation of Vegetation Patterns in Chaparral. Ecol. Monogr. 2000, 70, 149–169. [Google Scholar] [CrossRef]
- Francis, J.K. Wildland Shrubs of the United States and Its Territories: Thamnic Descriptions, Volume 1; General Technical Reports IITF-GTR-26; US Department of Agriculture, Forest Service, International Institute of Tropical Forestry: San Juan, Puerto Rico; US Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2004; 830p. [Google Scholar]
- Weir, J.R. Conducting Prescribed Fires: A Comprehensive Manual; Texas A&M University Press: College Station, TX, USA, 2009. [Google Scholar]
- Twidwell, D.; Bielski, C.H.; Scholtz, R.; Fuhlendorf, S.D. Advancing Fire Ecology in 21st Century Rangelands. Rangel. Ecol. Manag. 2021, 78, 201–212. [Google Scholar] [CrossRef]
- Teague, R.; Ansley, J.; Kreuter, U.; McGrann, J.; Pinchak, B. Fire vs. Herbicide? Rangelands 2001, 23, 9–14. [Google Scholar] [CrossRef]
- Van Liew, D.; Richard Conner, J.; Kreuter, U.P.; Teague, R. An Economic Comparison of Prescribed Extreme Fire and Alternative Methods for Managing Invasive Brush Species in Texas: A Modeling Approach. Open Agric. J. 2012, 6, 17–26. [Google Scholar] [CrossRef]
- Fogarty, D.T.; Roberts, C.P.; Uden, D.R.; Donovan, V.M.; Allen, C.R.; Naugle, D.E.; Jones, M.O.; Allred, B.W.; Twidwell, D. Woody Plant Encroachment and the Sustainability of Priority Conservation Areas. Sustainability 2020, 12, 8321. [Google Scholar] [CrossRef]
- Kloesel, K.; Bartush, B.; Banner, J.; Brown, D.; Lemery, J.; Lin, X.; Loeffler, C.; McManus, G.; Mullens, E.; Nielsen-Gammon, J.; et al. Southern Great Plains. In Impacts, Risks, and Adaptation in the United States: Fourth National Climate Assessment, Volume II; U.S. Global Change Research Program: Washington, DC, USA, 2018. [Google Scholar]
- McGranahan, D.A.; Wonkka, C.L.; Dangi, S.; Spiess, J.W.; Geaumont, B. Mineral Nitrogen and Microbial Responses to Soil Heating in Burned Grassland. Geoderma 2022, 424, 116023. [Google Scholar] [CrossRef]
- Campbell, G.S.; Jungbauer, J., Jr.; Bristow, K.L.; Hungerford, R.D. Soil Temperature and Water Content beneath a Surface Fire. Soil Sci. 1995, 159, 363–374. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).