
After the Megafires: Effects of Fire Severity on Reptile Species Richness and Occupancy in South-Eastern Australia




Abstract
1. Introduction
2. Materials and Methods
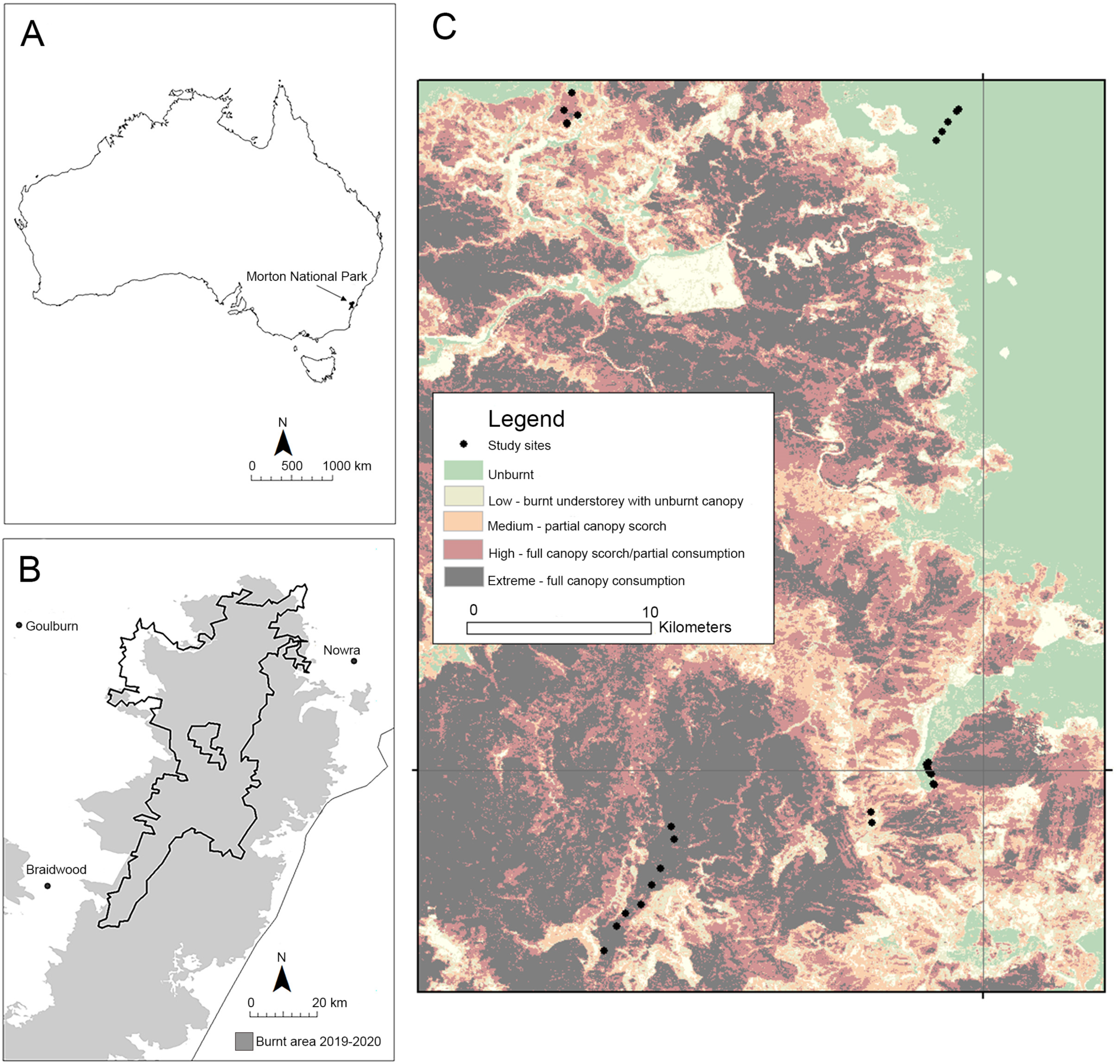
2.1. Study Area and Monitoring Plots
2.2. Reptile Surveys
2.3. Vegetation Surveys
2.4. Quantifying the Thermal Environment
2.5. Statistical Analyses
3. Results
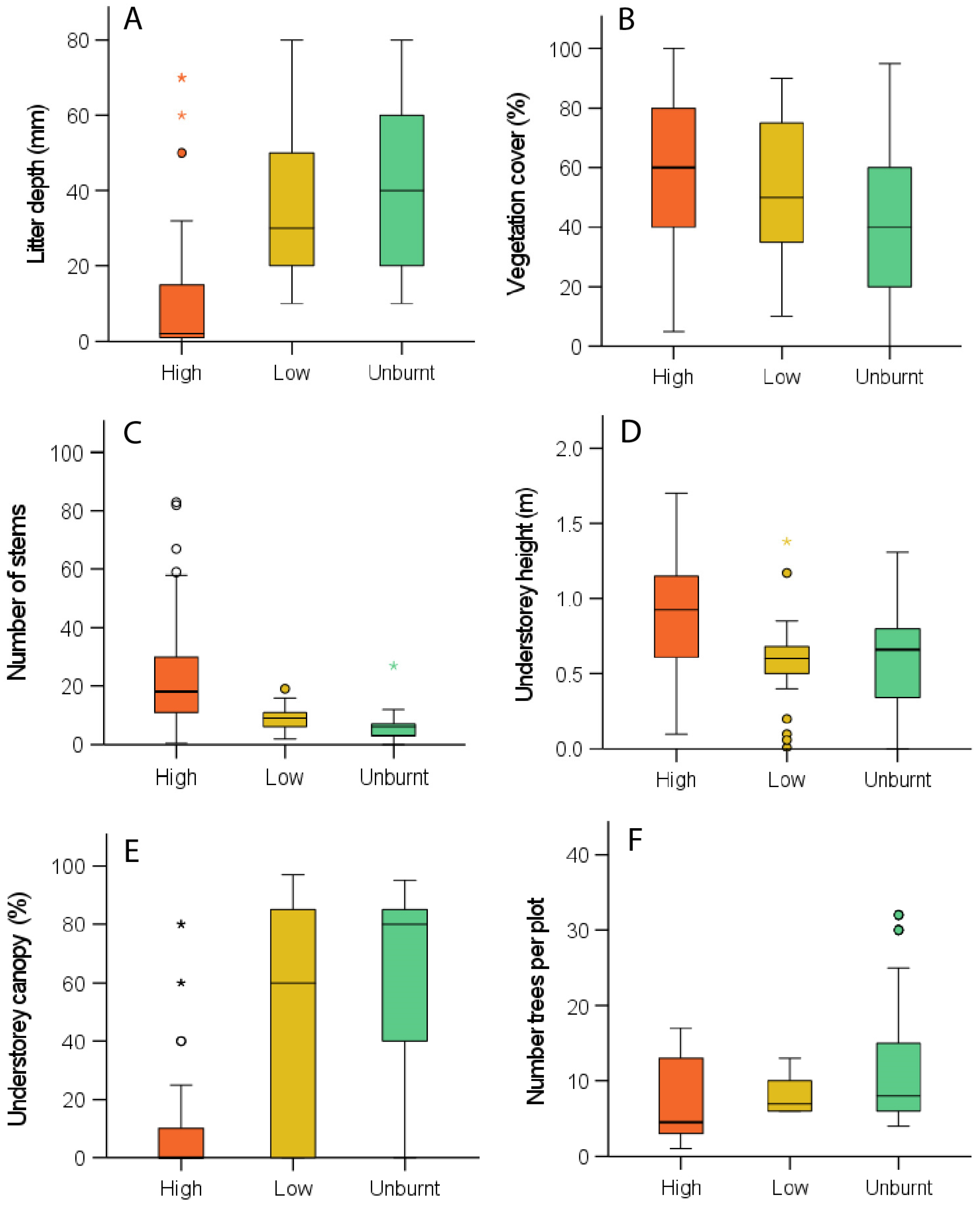
3.1. Vegetation Structure
3.2. Reptile Species Richness
3.3. Occupancy Estimates
3.4. Thermal Profiles of Reptile Retreat-Sites
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tran, B.N.; Tanase, M.A.; Bennett, L.T.; Aponte, C. High-severity wildfires in temperate Australian forests have increased in extent and aggregation in recent decades. PLoS ONE 2020, 15, e0242484. [Google Scholar] [CrossRef] [PubMed]
- Pausas, J.G.; Parr, C.L. Towards an understanding of the evolutionary role of fire in animals. Evol. Ecol. 2018, 32, 113–125. [Google Scholar] [CrossRef]
- Legge, S.; Woinarski, J.C.Z.; Scheele, B.C.; Garnett, S.T.; Lintermans, M.; Nimmo, D.G.; Whiterod, N.S.; Southwell, D.M.; Ehmke, G.; Buchan, A.; et al. Rapid assessment of the biodiversity impacts of the 2019–2020 Australian megafires to guide urgent management intervention and recovery and lessons for other regions. Divers. Distrib. 2021, 28, 571–591. [Google Scholar] [CrossRef]
- Santos, X.; Belliure, J.; Gonçalves, J.F.; Pausas, J.G. Resilience of reptiles to megafires. Ecol. Appl. 2022, 32, e2518. [Google Scholar] [CrossRef] [PubMed]
- Tomas, W.M.; Berlinck, C.N.; Chiaravalloti, R.M.; Faggioni, G.P.; Strüssmann, C.; Libonati, R.; Abrahão, C.R.; do Valle Alvarenga, G.; de Faria Bacellar, A.E.; de Queiroz Batista, F.R.; et al. Distance sampling surveys reveal 17 million vertebrates directly killed by the 2020’s wildfires in the Pantanal, Brazil. Sci. Rep. 2021, 11, 23547. [Google Scholar] [CrossRef]
- Griffiths, A.D.; Christian, K.A. The effects of fire on the frillneck lizard (Chlamydosaurus kingii) in northern Australia. Aust. J. Ecol. 1996, 21, 386–398. [Google Scholar] [CrossRef]
- Ward, M.; Tulloch, A.I.T.; Radford, J.Q.; Williams, B.A.; Reside, A.E.; Macdonald, S.L.; Mayfield, H.J.; Maron, M.; Possingham, H.P.; Vine, S.J.; et al. Impact of 2019-2020 mega-fires on Australian fauna habitat. Nat. Ecol. Evol. 2020, 4, 1321–1326. [Google Scholar] [CrossRef] [PubMed]
- Nimmo, D.G.; Carthey, A.J.R.; Jolly, C.J.; Blumstein, D.T. Welcome to the Pyrocene: Animal survival in the age of megafire. Glob. Chang. Biol. 2021, 27, 5684–5693. [Google Scholar] [CrossRef]
- Jolly, C.J.; Dickman, C.R.; Doherty, T.S.; van Eeden, L.M.; Geary, W.L.; Legge, S.M.; Woinarski, J.C.Z.; Nimmo, D.G. Animal mortality during fire. Glob. Chang. Biol. 2022, 28, 2053–2065. [Google Scholar] [CrossRef]
- Nimmo, D.G.; Kelly, L.T.; Farnsworth, L.M.; Watson, S.J.; Bennett, A.F. Why do some species have geographically varying responses to fire history? Ecography 2014, 37, 805–813. [Google Scholar] [CrossRef]
- Driscoll, D.A.; Smith, A.L.; Blight, S.; Maindonald, J. Reptile responses to fire and the risk of post-disturbance sampling bias. Biodivers. Conserv. 2012, 21, 1607–1625. [Google Scholar] [CrossRef]
- Pringle, R.M.; Webb, J.K.; Shine, R. Canopy structure, microclimate, and habitat selection by a nocturnal snake. Hoplocephalus bungaroides. Ecology 2003, 84, 2668–2679. [Google Scholar] [CrossRef]
- Elzer, A.L.; Pike, D.A.; Webb, J.K.; Hammill, K.; Bradstock, R.A.; Shine, R. Forest-fire regimes affect thermoregulatory opportunities for terrestrial ectotherms. Austral Ecol. 2013, 38, 190–198. [Google Scholar] [CrossRef]
- Lindenmayer, D.B.; Wood, J.T.; MacGregor, C.; Michael, D.R.; Cunningham, R.B.; Crane, M.; Montague-Drake, R.; Brown, D.; Muntz, R.; Driscoll, D.A. How predictable are reptile responses to wildfire? Oikos 2008, 117, 1086–1097. [Google Scholar] [CrossRef]
- Webb, J.K.; Shine, R. Differential effects of an intense wildfire on survival of sympatric snakes. J. Wildl. Manag. 2008, 72, 1394–1398. [Google Scholar] [CrossRef]
- Webb, J.K.; Jolly, C.J.; Hinds, M.; Adams, C.; Cuartas-Villa, S.; Lapwong, Y.; Letnic, M. Effects of the Australian 2019–2020 megafires on a population of endangered broad-headed snakes Hoplocephalus bungaroides. Austral Ecol. 2023, 48, 24–30. [Google Scholar] [CrossRef]
- Letnic, M.; Roberts, B.; Hodgson, M.; Ross, A.K.; Cuartas, S.; Lapwong, Y.; Price, O.; Sentinella, N.; Webb, J.K. Fire severity influences the post-fire habitat structure and abundance of a cool climate lizard. Austral Ecol. 2023, 48, 1440–1453. [Google Scholar] [CrossRef]
- Price, O.F.; Mikac, K.; Wilson, N.; Roberts, B.; Critescu, R.H.; Gallagher, R.; Mallee, J.; Donatiou, P.; Webb, J.; Keith, D.A.; et al. Short-term impacts of the 2019–20 fire season on biodiversity in eastern Australia. Austral Ecol. 2023, 48, 3–11. [Google Scholar] [CrossRef]
- Driscoll, D.A.; Lindenmayer, D.B.; Bennett, A.F.; Bode, M.; Bradstock, R.A.; Cary, G.J.; Clarke, M.F.; Dexter, N.; Fensham, R.; Friend, G.; et al. Fire management for biodiversity conservation: Key research questions and our capacity to answer them. Biol. Conserv. 2010, 143, 1928–1939. [Google Scholar] [CrossRef]
- Boer, M.M.; Resco de Dios, V.; Bradstock, R.A. Unprecedented burn area of Australian mega forest fires. Nat. Clim. Chang. 2020, 10, 171–172. [Google Scholar] [CrossRef]
- King, A.D.; Pitman, A.J.; Henley, B.J.; Ukkola, A.M.; Brown, J.R. The role of climate variability in Australian drought. Nat. Clim. Chang. 2020, 10, 177–179. [Google Scholar] [CrossRef]
- Collins, L.; Bradstock, R.A.; Clarke, H.; Clarke, M.F.; Nolan, R.H.; Penman, T.D. The 2019/2020 mega-fires exposed Australian ecosystems to an unprecedented extent of high-severity fire. Environ. Res. Lett. 2021, 16, 044029. [Google Scholar] [CrossRef]
- Keeley, J.E.; Fotheringham, C.J.; Baer-Keeley, M. Determinants of postfire recovery and succession in mediterranean-climate shrublands of California. Ecol. Appl. 2005, 15, 1515–1534. [Google Scholar] [CrossRef]
- Santos, X.; Badiane, A.; Matos, C. Contrasts in short- and long-term responses of Mediterranean reptile species to fire and habitat structure. Oecologia 2016, 180, 205–216. [Google Scholar] [CrossRef]
- Doherty, T.S.; Geary, W.L.; Jolly, C.J.; Macdonald, K.J.; Miritis, V.; Watchorn, D.J.; Cherry, M.J.; Conner, L.M.; González, T.M.; Legge, S.M.; et al. Fire as a driver and mediator of predator–prey interactions. Biol. Rev. Camb. Philos. Soc. 2022, 97, 1539–1558. [Google Scholar] [CrossRef]
- Black, D.E. The Vegetation of the Ettrema and Northern Budawangs Wilderness Areas, Morton National Park, New South Wales. Master’s Thesis, Department of Geology, University of Wollongong, Wollongong, Australia, 1995. [Google Scholar]
- State Government of NSW and NSW Department of Climate Change, Energy, the Environment and Water. Fire Extent and Severity Mapping (FESM). In The Sharing and Enabling Environmental Data Portal; 2020. Available online: https://datasets.seed.nsw.gov.au/dataset/33c2ee86-d2f7-4aaf-8c40-76b6d393a35c (accessed on 27 August 2024).
- Gibson, R.; Danaher, T.; Hehir, W.; Collins, L. A remote sensing approach to mapping fire severity in south-eastern Australia using sentinel 2 and random forest. Remote Sens. Environ. 2020, 240, 111702. [Google Scholar] [CrossRef]
- Michael, D.R.; Florance, D.; Crane, M.; Blanchard, W.; Lindenmayer, D.B. Barking up the right tree: Comparative use of arboreal and terrestrial artificial refuges to survey reptiles in temperate eucalypt woodlands. Wildl. Res. 2018, 45, 185–192. [Google Scholar] [CrossRef]
- Foster, C.N.; Barton, P.S.; Sato, C.F.; Wood, J.T.; MacGregor, C.I.; Lindenmayer, D.B. Herbivory and fire interact to affect forest understory habitat, but not its use by small vertebrates. Anim. Conserv. 2016, 19, 15–25. [Google Scholar] [CrossRef]
- Wilson, S.; Swan, G. A Complete Guide to Reptiles of Australia; Reed New Holland Publishers: Sydney, Australia, 2021. [Google Scholar]
- Doherty, T.S.; Davis, R.A.; van Etten, E.J.B.; Collier, N.; Krawiec, J. Response of a shrubland mammal and reptile community to a history of landscape-scale wildfire. Int. J. Wildland Fire 2015, 24, 534–543. [Google Scholar] [CrossRef]
- Kruskal, W.H.; Wallis, W.A. Use of ranks in one-criterion variance analysis. J. Am. Stat. Assoc. 1952, 47, 583–621. [Google Scholar] [CrossRef]
- Spearman, C. The proof and measurement of association between two things. Am. J. Psychol. 1904, 15, 72–101. [Google Scholar] [CrossRef]
- Bakus, J.G. Types of data, standardizations and transformations, introduction to biometrics, experimental design. In Quantitative Analysis of Marine Biological Communities; John Wiley & Sons: Hoboken, NJ, USA, 2007; pp. 62–122. [Google Scholar]
- Dunn, O.J. Multiple comparisons using rank sums. Technometrics 1964, 6, 241–252. [Google Scholar] [CrossRef]
- MacKenzie, D.I.; Nichols, J.D.; Royle, J.A.; Pollock, K.H.; Bailey, L.; Hines, J.E. Occupancy Estimation and Modeling: Inferring Patterns and Dynamics of Species Occurrence; Elsevier Science & Technology: San Diego, CA, USA, 2017. [Google Scholar]
- MacKenzie, D.I.; Nichols, J.D.; Lachman, G.B.; Droege, S.; Royle, J.A.; Langtimm, C.A. Estimating site occupancy rates when detection probabilities are less than one. Ecology 2002, 83, 2248–2255. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R. Multimodel Inference: Understanding AIC and BIC in Model Selection. Sociol. Methods Res. 2004, 33, 261–304. [Google Scholar] [CrossRef]
- Goulet, C.T.; Thompson, M.B.; Chapple, D.G. Repeatability and correlation of physiological traits: Do ectotherms have a “thermal type”? Ecol. Evol. 2017, 7, 710–719. [Google Scholar] [CrossRef]
- Blanca, M.J.; Arnau, J.; García-Castro, F.J.; Alarcón, R.; Bono, R. Repeated measures ANOVA and adjusted F-tests when sphericity is violated: Which procedure is best? Front. Psychol. 2023, 14, 1192453. [Google Scholar] [CrossRef] [PubMed]
- Barker, J.W.; Price, O.F.; Jenkins, M.E. High severity fire promotes a more flammable eucalypt forest structure. Austral Ecol. 2022, 47, 519–529. [Google Scholar] [CrossRef]
- Denham, A.J.; Vincent, B.E.; Clarke, P.J.; Auld, T.D. Responses of tree species to a severe fire indicate major structural change to Eucalyptus–Callitris forests. Plant Ecol. 2016, 217, 617–629. [Google Scholar] [CrossRef]
- Bassett, M.; Leonard, S.W.J.; Chia, E.K.; Clarke, M.F.; Bennett, A.F. Interacting effects of fire severity, time since fire and topography on vegetation structure after wildfire. For. Ecol. Manag. 2017, 396, 26–34. [Google Scholar] [CrossRef]
- Gordon, C.E.; Price, O.F.; Tasker, E.M.; Denham, A.J. Acacia shrubs respond positively to high severity wildfire: Implications for conservation and fuel hazard management. Sci. Total Environ. 2017, 575, 858–868. [Google Scholar] [CrossRef]
- Lindsay, M.N.; Lewis, D.B.; Halstead, N.; Gainsbury, A.M. Fire severity effects on the herpetofaunal diversity of the Florida scrub, a biodiversity hotspot. Biodivers. Conserv. 2023, 32, 1857–1878. [Google Scholar] [CrossRef]
- Legge, S.; Murphy, S.; Heathcote, J.; Flaxman, E.; Augusteyn, J.; Crossman, M. The short-term effects of an extensive and high-intensity fire on vertebrates in the tropical savannas of the central Kimberley, northern Australia. Wildl. Res. 2008, 35, 33–43. [Google Scholar] [CrossRef]
- Taylor, J.E.; Ellis, M.V.; Williams, N.; Kloecker, U. Responses of birds and reptiles in Warrumbungle National Park after the extensive 2013 wildfire. Proc. Linn. Soc. N. S. W. 2020, 142, S155–S208. [Google Scholar]
- Evans, M.J.; MacGregor, C.; Lindenmayer, D. A misleading tail: A long-term study of reptile responses to multiple disturbances undermined by a change in surveying techniques. PLoS ONE 2024, 19, e0305518. [Google Scholar] [CrossRef] [PubMed]
- Spence-Bailey, L.M.; Nimmo, D.G.; Kelly, L.T.; Bennett, A.F.; Clarke, M.F. Maximising trapping efficiency in reptile surveys: The role of seasonality, weather conditions and moon phase on capture success. Wildl. Res. 2010, 37, 104–115. [Google Scholar] [CrossRef]
- Letnic, M.; Dickman, C.R.; Tischler, M.K.; Tamayo, B.; Beh, C.L. The responses of small mammals and lizards to post-fire succession and rainfall in arid Australia. J. Arid Environ. 2004, 59, 85–114. [Google Scholar] [CrossRef]
- Hodgson, M.; Ross, A.; Lapwong, Y.; Cuartas, S.; Roberts, B.; Price, O.; Webb, J.K.; Sentinella, N.; Lee, J.; Laffan, S.; et al. A cryptic elapid snake persists in the wake of catastrophic wildfires. Oryx 2024, in press.
- Taylor, J.E.; Fox, B.J. Disturbance effects from fire and mining produce different lizard communities in eastern Australian forests. Austral Ecol. 2001, 26, 193–204. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Number and Percentage of Plots in Which Each Species Was Detected | |||
|---|---|---|---|
| Reptile Species | Unburnt (n = 10) | Low Severity (n = 5) | High Severity (n = 13) |
| Amalosia lesueurii | 0 | 1 (20%) | 0 |
| Acritoscincus platynotus | 0 | 1 (20%) | 3 (23.1%) |
| Ctenotus taeniolatus | 0 | 0 | 2 (15.4%) |
| Lampropholis delicata | 5 (50%) | 1 (20%) | 6 (46.2%) |
| Lampropholis guichenoti | 1 (10%) | 0 | 2 (15.4%) |
| Saproscincus mustelina | 0 | 3 (60%) | 0 |
| Drysdalia rhodogaster | 1 (10%) | 0 | 1 (7.7%) |
| Detection | Occupancy | AICc | Delta AICc | AICc Weight | Model Likelihood | N |
|---|---|---|---|---|---|---|
| Time | Constant | 99.10 | 0.000 | 0.48 | 1.00 | 6 |
| Constant | Constant | 99.81 | 0.709 | 0.34 | 0.70 | 2 |
| Fire severity | Time | 102.89 | 3.797 | 0.07 | 0.15 | 4 |
| Constant | Fire severity | 103.03 | 3.925 | 0.07 | 0.14 | 4 |
| Time | Fire severity | 104.34 | 5.234 | 0.03 | 0.07 | 8 |
| Fire severity | Fire severity | 106.76 | 7.660 | 0.01 | 0.02 | 6 |
| Fire severity × time | Constant | 154.91 | 55.809 | 0.00 | 0.00 | 16 |
| Fire severity × time | Fire severity | 179.82 | 80.714 | 0.00 | 0.00 | 18 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Archer, M.L.; Letnic, M.; Murray, B.R.; Webb, J.K. After the Megafires: Effects of Fire Severity on Reptile Species Richness and Occupancy in South-Eastern Australia. Fire 2024, 7, 349. https://doi.org/10.3390/fire7100349
Archer ML, Letnic M, Murray BR, Webb JK. After the Megafires: Effects of Fire Severity on Reptile Species Richness and Occupancy in South-Eastern Australia. Fire. 2024; 7(10):349. https://doi.org/10.3390/fire7100349
Chicago/Turabian StyleArcher, Maddison L., Mike Letnic, Brad R. Murray, and Jonathan K. Webb. 2024. "After the Megafires: Effects of Fire Severity on Reptile Species Richness and Occupancy in South-Eastern Australia" Fire 7, no. 10: 349. https://doi.org/10.3390/fire7100349
APA StyleArcher, M. L., Letnic, M., Murray, B. R., & Webb, J. K. (2024). After the Megafires: Effects of Fire Severity on Reptile Species Richness and Occupancy in South-Eastern Australia. Fire, 7(10), 349. https://doi.org/10.3390/fire7100349