Abstract
In fire-prone ecosystems, plant resilience to recurrent fires depends on certain fire-adaptive traits. However, how key functional and reproductive traits of graminoids respond to varying burning seasons is poorly understood. This meta-analysis, therefore, unpacks global perspectives on how resprouting, growth, reproductive, and productivity traits of graminoids (grasses and sedges) respond to different burning seasons. We recorded 569 observations from 80 experimental studies comparing graminoid plant trait responses in unburned vs. burned treatments over different seasons of burn. Weighted log response ratios and 95% confidence intervals (95%CI) were analyzed for each plant trait using random effects models and compared across burning seasons. Summer (0.35 (95%CI = 0.25 to 0.46)) and autumn burns (0.24 (95%CI = 0.16 to 0.31)) increased above-ground biomass m−2, while biomass plant−1 was increased only by spring burns (0.27 (95%CI = 0.22 to 0.32)). Bud production plant−1 and tiller−1 were reduced significantly by fire, especially spring, summer, and autumn burns. The shoot height (0.29 (95%CI = 0.17 to 0.41)), leaf length (0.15 (95%CI = 0.11 to 0.20)), and specific leaf area (0.08 (95%CI = 0.06 to 0.09)) increased only under summer burns, while flowering was enhanced by spring (0.19 (95%CI = 0.00 to 0.38)) and autumn burns [0.34 (95%CI = 0.02 to 0.66)]. However, seed production m−2 was reduced by spring and summer burns and the opposite was true for seed production plant−1. Overall, herbaceous plant trait responses to fire varied by the season of burn, disagreeing with the general principle that early spring burning is the best practice. We suggest that a decision on the season of burn should be informed by the objective of burning.
1. Introduction
Fire is an integral part of grasslands and woodlands and is key in preventing state shift to a bush encroached state and in increasing diversity and plant quality and productivity [,,,,,]. However, the effects of fire on vegetation are complex and vary both inter- and intra-specifically, depending on plant adaptive traits and the fire regime [,,]. Certain ecosystems co-evolved with fire; hence, their structure, functioning, and services are sustained by positive feedback between vegetation and fire [,,]. This feedback emanates from the fact that certain fire regimes select for certain plant adaptive traits [,,].
Fynbos shrublands of South Africa, Chaparral shrublands and prairies of America, and Cerrado grasslands of Brazil are typical ecosystems in which vegetation employs specialized strategies to adapt to recurrent fires []. Plants respond to fire mainly by seed dormancy release [], resprouting [], and fire-induced synchronous flowering [,]. For instance, summer fires increased the flowering of grasses in European [] and North American grasslands [], whereas spring fires promoted bunchgrass resprouting in African [] and Australian ecosystems []. Resprouting and recruitment from the seed, respectively, depend on bud and seed bank sizes [], and these post-fire recovery strategies promote plant population persistence [,]. The ability to recover clonally following fires depends largely on whether plant buds are located below or above ground []. Generally, plant buds that escape fire, more so below-ground plant buds, remain dormant and resprout when conditions are suitable following fires []. However, in other ecosystems, non-prescribed fires may be detrimental to fire-sensitive species that lack certain fire-adaptive traits [,]. A worst-case scenario is the extinction of these species, followed by the ingress of invaders, which exacerbates biodiversity loss [].
Graminoids, largely bunchgrasses, are amongst the most vulnerable growth forms to fires, due to the projection of their meristems to fire flame heights [,]. As a result, most bunch-type graminoids are obligate seeders, capitalizing largely on high flowering [] and en masse recruitment from the seed following fires [,]. Seeders allocate more resources to rapid growth to reach the reproductive stage prior to the next fire event, whereas clonal graminoids invest more to non-structural carbohydrate reserves and protective structures []. In some instances, light fires may reduce residual senesced fuel on grass tussocks, which increases light availability, thereby triggering the recruitment of new buds [].
Plant resilience to fire, however, is not exclusively dependent on plant phylogeny, but also on fire regime, including, amongst others, timing of burning []. The season of burn is crucial, as it affects the growth and reproductive cycles of graminoids, with consequential effects on plant persistence []. For example, burning when the meristematic tissues are elevated, and carbohydrate reserves are depleted during active growth hinders plant performance, survival, and persistence. Furthermore, burning during flowering has direct effects on plant fecundity, as seed production depends largely on the number of floral stalks []. These effects are modulated by seasonal variation in plant physiological and phenological changes and climatic conditions through influencing fuel load characteristics, which in turn influence the intensity of fire [].
A global rule of thumb is that burning in spring is the best practice []. However, this fire management prescription appears to be informed by community-scale responses without detailed understanding of its implications at a plant trait level. This limits our understanding of the mechanisms and adaptive traits underlying plant recovery and persistence following fires. Plant trait responses to fire may provide insights into plant performance, productivity, and reproductive capacity and help derive plant conservation strategies []. Furthermore, the future climate, characterized by recurrent droughts, is projected to shift burning seasons and frequencies, with fires likely to occur within short frequencies over extended seasons of burn. Thus, understanding plant responses to different burning seasons at a global scale form the basis for understanding resilience strategies of herbaceous plant communities to future climate-driven shifts in fire regime. A comprehensive assessment of plant trait responses to different burning seasons is crucial to unpack how plants recover and persist following fires of varying seasons of burn. This global meta-analysis was conducted to assess how growth, productivity, and reproductive traits of graminoids respond to varying seasons of burn.
2. Materials and Methods
2.1. Data Collection
Using advanced search function, we searched peer-reviewed articles reporting effects of fire on functional traits of graminoids (grasses and sedges) from Web of Science, Scopus, Google Scholar, and BioOne Complete. The search was restricted only to studies published from 1985 to 2022. The following search terms and combinations were used: effect of “fire” or “burning” or “season of burn” on (1) grasses, (2) sedges, (3) graminoids, (4) functional traits, (5) tiller or bud production, (6) resprouting, (7) flowering or seed production or plant fecundity, (8) plant growth or height, and (9) biomass production.
For each study, we read the title to assess the relevance and thoroughly screened the abstracts to determine whether observations were conducted in burn vs. unburned scenario. Studies were scrutinized to meet the following criteria: (1) the study reports the effect of fire on graminoids (grasses and sedges), and (2) the following were provided: vegetation type (e.g., grassland, savanna, shrubland, etc.), season of burn, and sampling dates, and (3) the comparisons were conducted between burned and unburned scenario either in a form of Control-Impact (CIT), Before-After-Impact (BAIT) and or Before-After-Control-Impact (BACIT) experimental frameworks. The CIT framework refers to comparisons between burned and unburned treatments, whereas BAIT scenario refers to comparisons before and after burning []. The BACIT scenario refers to comparisons between burn treatment and control before and after fire application [].
The response variables assessed were shoot and root biomass (on g m−2 and g plant−1 basis), bud production [on per area (m2), plant and tiller basis], plant height (cm), leaf length (cm), specific leaf area (g cm−1) and seed production (on seeds m−2, plant−1, and stalk−1 basis). In scenarios where fire was applied in combination with other treatments, e.g., grazing, shrub control, etc., we selected a control which included fire only. Moreover, the historic background of the burned and unburned treatments was scrutinized and the studies in which previous management histories of the two treatments differed were rejected. In the case of BAIT and BACIT experimental frameworks, we selected mainly the studies in which covariates, e.g., rainfall, temperature, etc., were included in the statistical models and reported to have had no additive or confounding effect. For the studies in which repeated measures were conducted, each observation was considered as an independent study. In each study, we recorded the reference, longitude and latitudes, species name, plant functional type (grass or sedge) and season of burn. For studies that did not report coordinates, we searched them by name of the study area and country using coordinate converter (https://coordinates-converter.com; Accessed on 20 July 2023). In cases where the specific name of a study area was not reported, for example when reported as “the site was located between area A and B”, we recorded the coordinates of the area mentioned first (A).
The statistical measures recorded were mean (), standard deviation (SD) and the sample size (n) for both burned and unburned scenarios. The statistical measures were extracted from tables or through digitizing figures using WebPlot digitizer 4.5 (https://github.com/ankitrohatgi/WebPlotDigitizer; Accessed on 20 July 2020).
We obtained a total of 416 experimental studies, of which 95 lacked relevance; subsequently, they were disqualified. The remaining 321 studies were assessed for eligibility; 241 were disqualified for reasons of content and 80 studies reporting 569 observations qualified for this meta-analysis. Most studies (36%) were conducted in North America, whereas 20 and 23% of studies were conducted in South America and Africa, respectively (Figure 1). Of these studies, 50% were conducted in grasslands (Figure 1).
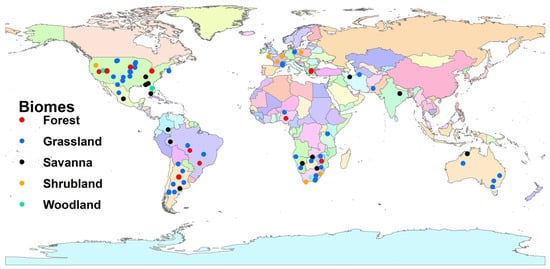
Figure 1.
Global distribution of studies reporting graminoid functional trait responses to fire across different biomes.
2.2. Meta-Analytical Procedures
Firstly, when the standard error of mean (SEM) was reported instead of SD, we converted it to SD using the following equation:
In cases where was reported with neither SD nor SEM, we computed the SD by dividing by 10. If 95% confidence interval (CI) was reported alongside with instead of SD or SEM, the following equation was employed to compute SD:
The random effect models (REMs) were used to compute weighted log response ratios (logRR) and their 95% confidence intervals. REM was selected for its high parsimony relative to fixed effects models, especially when between-study variance is expected to be high, due probably to differences in plant functional types, vegetation types, fire characteristics and area-specific climatic conditions []. The logRRs were computed according to the following equation and displayed in a forest plot together with their weightings:
where and are mean values of the response variables in burned and unburned treatments, respectively.
The significant effect of the season of burn on plant traits was noted if the CI did not overlap with zero, whereas the significant differences between seasons of burn were noted if the CI of the two seasons did not overlap []. We generated 1000 bootstrap permutations to determine the intercept (model estimate) of the REM and used a Z-test to assess the significance of the REM intercept at α = 5%. The model fit was assessed using a restricted maximum likelihood (REML), log likelihood specifically. Due to assumed variability in study variances, the logRRs were weighted using the inverse of the variance as follows:
where SDb and SDub represent the standard deviation of burned and unburned treatments, respectively. The nb and nub are sample sizes of burned and unburned treatments, respectively. All the analysis was conducted using the package MAJOR in Jamovi 2.3, which is a graphical interface of R.
2.3. Detection of Study Bias
A general assumption is that studies reporting significant differences between treated vs. untreated scenarios are more likely to be published than those reporting marginal differences. For this scenario, we conducted meta-regression between sample sizes and response ratios using tau as an estimator, with a significant tau value at 95% confidence level indicating bias. We further assessed between-study variance using Q-test and a Fail-Safe N value and complemented them with funnel plots of sample sizes vs. logRRs for visualization of data asymmetry. After detecting bias, studentized residuals and Cook’s distance measure were used to identify the potential outlier studies and to check that no study overly influenced the logRR. The sensitivity analysis using trim and fill method was conducted to ascertain what the effect sizes would be if missing studies were included in the meta-analysis.
3. Results
3.1. Above- and Below-Ground Biomass
The overall plant responses to fire in terms of above-ground biomass per area (Z = 0.89, p = 0.375) and per plant (Z = −1.33, p = 0.182) was not significant (Figure 2a–c). However, above-ground biomass (AGB) varied by the season of burn, with AGB per area exhibiting a significant (p < 0.05) increase for summer and autumn burns compared to spring and winter burns (Figure 2a). In the latter two seasons of burn, AGB was negatively affected by fire, declining significantly by effect sizes of −0.04 (CI = −0.13; 0.05) and −0.17 (CI = −0.39; 0.04), respectively (Figure 2a). Similarly, AGB per plant varied by the season of burn, responding positively to spring burns, while it responded negatively to other seasons of burn, more so to summer burns (−1.95 (CI = −2.26; −1.65); Figure 2b). The overall response of root biomass (RB) to fire was not significant (Z = 1.66, p = 0.096), but RB varied by the season of burn (Figure 2c). The spring burns increased RB four-fold higher than autumn and summer burns, whereas winter burns reduced RB by an effect size of −0.06 (CI = 0.40; 0.29; Figure 2c).
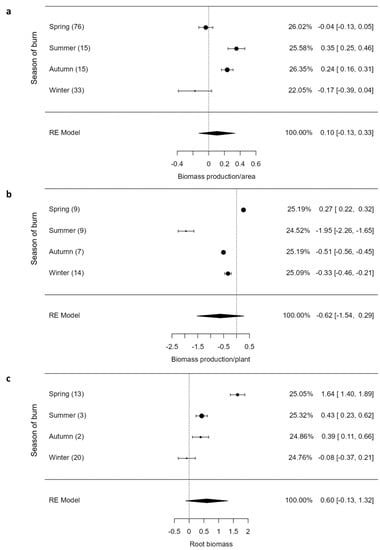
Figure 2.
Aboveground biomass per area (a) and plant (b) and root biomass (c) of graminoids across burning seasons. The values inside rounded parentheses are the number of studies (n) and those inside square parentheses denote upper and lower limits (±95% CIs). The values presented in percentages are weighting values for logRRs. The higher the contribution to the pooled effect size, the larger the weighting. RE model stands for random effects model.
3.2. Bud Production
The overall bud production per area showed no significant (Z = 0.613, p = 0.540) responses to fire. However, bud production per plant (Z = −2.35, p = 0.019) and per tiller (Z = −2.44, p = 0.015) showed significant responses to fire, with bud production responses varying by the season of burn (Figure 3b,c). Bud production per area increased under spring burns and decreased under summer and autumn burns (Figure 3a). For both per plant and tiller, bud production was negatively affected by fire, with summer burns causing more lethal effects [−1.02 (CI = −1.23; −0.88)] than autumn [−0.55 (CI = −0.68; −0.42)] and winter burns (0.11 (−0.24; 0.46); Figure 3c). Bud production per tiller was reduced significantly by fire, with more significant declines being evident under autumn burns than in spring burns (Figure 3c).
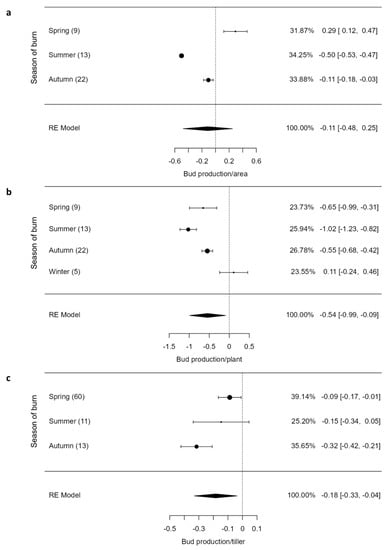
Figure 3.
Bud production per area (a), plant (b) and tiller (c) of graminoids across burning seasons. The values inside rounded parentheses is the number of studies (n) and those inside square parentheses denote upper and lower limits (±95% CIs). The values presented in percentages are weighting values of the logRRs. The higher the contribution to the pooled effect size, the larger the weighting. RE model stands for random effects model.
3.3. Height, Leaf Length, and Specific Leaf Area
The overall response of shoot height (SH), leaf length (LL), and specific leaf area (SLA) to fire was insignificant (p > 0.05), but these parameters varied by the season of burn (Figure 4a–c). The SH (0.29 (CI = 0.17; 0.41)) and LL (0.15 (CI = 0.11; 0.20)) were enhanced significantly only by summer burns (Figure 4a,b). Specifically, spring burns had remarkable negative effects on SH and LL, but LL was reduced more by autumn burns (Figure 4a,b). The SLA was marginally increased by an effect size of 0.08 (CI = 0.06; 0.09) by summer burns, whereas autumn and winter burns reduced it by −0.05 (CI = −0.07; 0.02) and −0.28 (CI = −1.19; 0.63), respectively (Figure 4c).
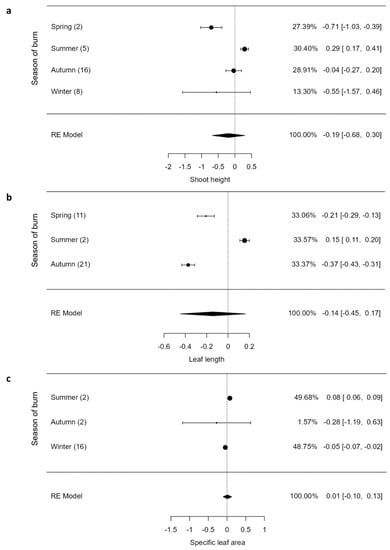
Figure 4.
Shoot height (a), leaf length (b), and specific leaf area (c) of graminoids across burning seasons. The values inside rounded parentheses is the number of studies (n) and those inside square parentheses denote upper and lower limits (±95% CIs). The values presented in percentages are weighting values of the logRRs. The higher the contribution to the pooled effect size, the larger the weighting. RE model stands for random effects model.
3.4. Reproductive Traits
The fire had a significant effect on flowering (Z = 2.85, p = 0.004), with spring and autumn burns significantly increasing flowering relative to winter burns, which reduced flowering by a magnitude of −0.17 (CI = −0.93; 0.60; Figure 5). The overall response of seed production per area (Z = −0.782, p = 0.434) and per plant (Z = 1.30, p = 0.194) to fire was not significant (Figure 6a,b). However, seed production on a per area, plant, and stalk basis varied remarkably across seasons of burn (Figure 6a–c). The seed production per area declined significantly in summer and spring burns and the opposite was true for autumn and winter burns (Figure 6a). Conversely, spring and summer burns increased seed production per plant compared to autumn and winter burns (Figure 6b).
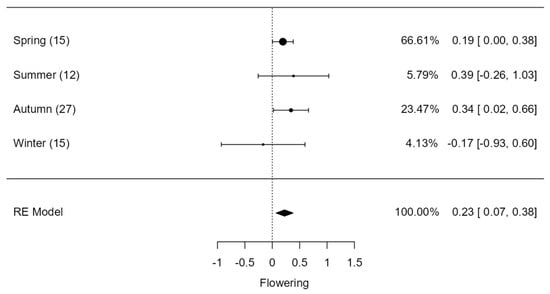
Figure 5.
Flowering of graminoids across burning seasons. The values inside rounded parentheses is the number of studies (n) and values inside square parentheses denote upper and lower limits (±95% CIs). The values presented in percentages are weighting values of the logRRs. The higher the contribution to the pooled effect size, the larger the weighting. RE model stands for random effects model.
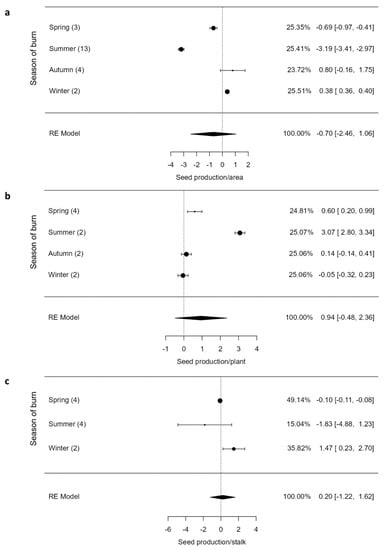
Figure 6.
Seed production per area (a), plant (b), and stalk (c) of graminoids across burning seasons. The values inside rounded parentheses is the number of studies (n) and those inside square parentheses denote upper and lower limits (±95% CIs). The values presented in percentages are weighting values of the logRRs. The higher the contribution to the pooled effect size, the larger the weighting. RE model stands for random effects model.
4. Discussion
4.1. Biomass and Growth of Graminoids Depends on Burning Season
The effect of the season of burn on above-ground biomass (AGB) depended largely on the scale of measurement, with spring burns reducing biomass per area and increasing biomass per plant. This suggests that spring burns are detrimental at a stand level, probably due to fire eliminating fire-sensitive species, thereby reducing their contribution to stand-level biomass. Generally, spring burns are applied after the first rains [], a time coinciding with seedling recruitment and early plant establishment. Plant seedlings are more susceptible to disturbance at this stage and are less likely to survive fire []. Hence, burning in summer and autumn increased AGB per area because during these burning seasons, plants are well developed and the high moisture content in their phytomass insulates them against fire. Moreover, the nutrients from the ash produced during summer burn are readily available and accessible by plants because the root system is well-developed, and the soil moisture is adequate to solubilize these nutrients []. Thus, increased nutrient uptake from the ash and high soil moisture content in summer compensate the plants through early post-fire recovery []. Such a compensatory response could be justified by the increase in shoot height and leaf length observed under summer burns relative to spring burns (Figure 4a,b). The increase in SLA under summer burns, particularly, could be attributable to higher growth rates, as indicated by increased plant height relative to other seasons of burn (Figure 4a,c). Nonetheless, AGB per plant was increased by spring burns and reduced by summer and autumn burns, regardless of increased leaf length and stem growth under summer burns. This could be explained by the reduction in AGB per area under spring burns (Figure 2b), which probably reduced plant competition, in turn increasing resource availability to fire survivors []. The increase in AGB per plant for spring burns, however, appears to be caused neither by tillering nor by enhanced growth because bud production, plant growth and leaf length were reduced by spring burns (Figure 3 and Figure 4a). Thus, enhancement of AGB per plant by spring burns could be ascribed to other factors not considered in this study, e.g., leaf production.
The results further showed that root biomass (RB) was enhanced by all burning seasons except winter burns (Figure 2c). Plant dormancy, which increases fuel load together with low soil and fuel moisture facilitates intense winter fires that cause severe soil heating and combustion of plant crowns, leading to the death of plant roots []. The roots of graminoids are distributed shallower close to the soil surface; thus, they are more susceptible to intense fires [,]. This is more common where ground fires are prevalent as they burn below soil surface and cause severe soil heating []. Conversely, RB was enhanced by spring, summer, and autumn burns (Figure 2c), indicating that reduced AGB per plant in the latter two burning seasons is compensated for by below-ground biomass investment. Hence, one would expect higher post-fire plant recovery under these burning seasons relative to winter burns. High RB enhances water and nutrient uptake [], of which both enhance plant recovery following fires.
4.2. Burning during Active Growth Reduces Bud Production of Graminoids
In agreement with previous experimental studies, we found that fire reduces bud production, especially spring, summer, and autumn burns. During these seasons, plant growth is still active, and fire becomes lethal to plant buds because the meristematic tissues are elevated to fire flames. Moreover, carbohydrates are also depleted during active growth, as they are channeled to photosynthesis, and this limits bud regeneration after fires []. Generally, the reduction in the most active growth points such as intercalary meristems due to burning fosters plant reliance on axillary buds for photosynthetic regeneration []. Since buds are highly sensitive to heat, cool fire from senesced annual plants is adequate to inhibit bud emergence [], but perhaps not enough to remove apical dominance. Since the fuel load in spring, summer, and autumn is not as abundant as in winter, it is presumable that incomplete combustion due to high foliar moisture prevented the removal of apical dominance, thereby limiting the regeneration of axillary buds. This is explained by positive responses of bud production under winter burns relative to other burning seasons. The reduction in bud production by summer and autumn burns was also reported by Russell and Vermeire []. Reduction in bud production may exacerbate loss of plant vigor and population persistence, especially in the ecosystems dominated by perennials, which rely largely on clonal regeneration []. Our results suggest that herbaceous plant recovery depends largely on seed production rather than vegetative bud regeneration following spring, summer, and autumn fires.
4.3. Burning Enhances Flowering and Seed Production of Graminoids
Our results showed that burning facilitates flowering in graminoids, more so in spring, summer, and autumn burns (Figure 5). Nonetheless, when these different burning seasons were compared, there was no obvious difference on their effects, suggesting that fire stimulates flowering regardless of the burning season. This response is not surprising, as other studies, largely those conducted in grasslands, reported synchronized flowering in grasses after fires regardless of the timing of burn [,]. However, this effect appears not to translate to high seed production per area for spring and summer burns (Figure 6a), but rather increased seed production per plant in these seasons (Figure 6b). The increase in seed production per plant was expected given that the number of seeds correlates with the number of inflorescences produced []. Generally, there is a trade-off between seed production and resprouting []; hence, we recorded higher seed production per plant in spring and summer burns despite reduced bud production per plant. This is because plants, particularly seeders, invest more resources in fast growth and seed production instead of vegetative resprouting []. This meta-analysis suggests that, globally, seed production is the main persistence strategy employed by graminoids to recover from fires. Generally, this strategy maintains plant populations via two important ways, including forming large seed bank sizes that remain dormant if conditions are not suitable for recruitment []. Secondly, the seeds of these plants may be easily dispersed in safe sites for germination []. The effect of summer fires on flowering might depend on the time of fire ignition within the summer season, as the effect of fire before floral initiation may not be as severe as during floral initiation. It appears, however, that increase in flowering under summer burns is a common plant response triggered by chemical compounds of smoke and ash together with temperatures induced by fire [,]. Flowering is not only dependent on fire-induced cues, but also on soil nutrients and moisture [,,,], which are more abundant and easily accessible by plants in summer.
4.4. Limitations of the Study and Future Research Directions
This meta-analysis assessed plant trait responses to different seasons of burning. Generally, a fire regime includes various factors which could interact at large landscape scale with the season of burn, e.g., fire return intervals []. Thus, the assessment of functional trait responses to burning season alone may limit broad understanding of plant response dynamics under scenarios where a wide spectrum of fire regime factors interact. This knowledge is pivotal, given that future climate change is expected to cause changes not only in the season of burn, but also the frequency with which fires will occur []. This information is fundamental to creating awareness about climate-driven shifts in fire regime and to derive future coping mechanisms for these fire regime shifts for land users, particularly conservationists and livestock farmers. Thus, experimental research that incorporates burning season and fire return intervals could give a better perspective on recovery mechanisms and strategies utilized by plants to persist under recurrent fires. This could also consider the fact that plants with varying life histories and phylogenies respond interspecifically to varying fire regimes []. Thus, considering responses across plants of different photosynthetic pathways, growth habits, and lifespans may provide clear prospects on how ecosystems with varying plant functional traits will be shaped by recurrent fires of varying regimes.
5. Conclusions
The current meta-analysis indicated that biomass and bud production responses to fire depend on the season of burn. Biomass production varied considerably across seasons of burn, depending also on the scale of measurement, with spring burns reducing biomass per area, while it increased biomass per plant. Almost all fire seasons had negative effects on bud production, whereas they enhanced the flowering of graminoids, suggesting that sexual regeneration is a significant persistence strategy for recovery from fires. In a broader context, this meta-analysis revealed that global ecosystems dominated by graminoids respond differently to different burning seasons depending on the functional plant trait of concern. We, therefore, deduce that the general principle that spring is the best burning season is subjective and that the decision on burning season should be informed by the objective of burning. The results of this study are fundamental for understanding regeneration and persistence strategies of graminoids following fire for designing appropriate fire management prescriptions in respect to the season of burning.
Author Contributions
M.M., H.K.T. and B.G. conceived the research idea and designed the study. M.M. collected the data. M.M. conducted the analysis of the meta-analysis of data and wrote the manuscript. B.G. and H.K.T. reviewed and provided valuable and critical comments on the manuscript. M.M. used the software. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
The data used in this study will be made available on request from the corresponding author.
Acknowledgments
The authors would like to thank CB Kapiyana, T Nkemnkem and BV Maselana in the department of Livestock and Pasture at the University of Fort Hare for their comments which improved the quality of this manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Villarreal, M.L.; Norman, L.M.; Buckley, S.; Wallace, C.S.A.; Coe, M.A. Multi-index time series monitoring of drought and fire effects on desert grasslands. Remote Sens Environ. 2016, 183, 186–197. [Google Scholar]
- Moore, N.A.; Camac, J.S.; Morgan, J.W. Effects of drought and fire on resprouting capacity of 52 temperate Australian perennial native grasses. New Phytologist. 2018, 221, 1424–1433. [Google Scholar] [CrossRef] [PubMed]
- Neary, D.G.; Leonard, J.M. Effects of fire on grassland soils and water: A review. In Grasses and Grassland Aspects; Kindomihou, V.M., Ed.; IntechOpen: London, UK, 2020. [Google Scholar] [CrossRef]
- Stradic, L.S.; Roumet, C.; Durigan, G.; Cancian, L.; Fidelis, A. Variation in biomass allocation and root functional parameters in response to fire history in Brazilian savannas. J. Ecol. 2021, 109, 4143–4157. [Google Scholar] [CrossRef]
- Ansley, R.J.; Boutton, T.W.; Hollister, E.B. Can prescribed fires restore C4 grasslands invaded by a C3 woody species and a co-dominant C3 grass species? Ecosphere 2021, 12, e03885. [Google Scholar] [CrossRef]
- Da Sliva Goldas, C.; Podgaiski, L.R.; da Silva, C.V.C. Burning for grassland pollination: Recently burned patches promote plant flowering and insect pollinators. Austral Ecol. 2021, 47, 491–506. [Google Scholar]
- Simpson, K.J.; Ripley, B.S.; Christin, P.A.; Belcher, C.M.; Lehmann, C.E.R.; Thomas, G.H.; Osborne, C.P. Determinants of flammability in savanna grass species. J. Ecol. 2016, 104, 138–148. [Google Scholar] [CrossRef]
- Pausas, J.G.; Paula, S. Grasses and fire: The importance of hiding buds: A response to Moore et al. (2019) ‘Effects of drought and fire on resprouting capacity of 52 temperate Australian perennial native grasses’. New Phytol. 2020, 226, 957–959. [Google Scholar]
- Te Beest, M.; Kleinjan, A.; Tuijnman, V.; Findlay, N.; Mvelase, T.; le Roux, E.; Tedder, M.; Gordijn, P.; Janse van Rensburg, S. Grass functional trait responses to experimental warming and fire in Afromontane grasslands. Afr. J. Range Forage Sci. 2021, 38, 88–101. [Google Scholar] [CrossRef]
- Zylstra, P. Linking fire behavior and its ecological effects to plant traits using FRaME in R. Methods Ecol. Evol. 2020, 12, 1365–1378. [Google Scholar] [CrossRef]
- Bombo, A.B.; Appezzato-da-Glória, B.; Fidelis, A. Fire exclusion changes belowground bud bank and bud-bearing organ composition jeopardizing open savanna resilience. Oecologia 2022, 199, 153–164. [Google Scholar]
- Keleey, J.E.; Pausas, J.G.; Rundell, P.W.; Bond, W.J.; Bradstock, R.A. Fire as an evolutionary pressure shaping plant traits. Trends Plant Sci. 2011, 16, 406–410. [Google Scholar]
- Lamont, B.B. Historical links between climate and fire on species dispersion and trait evolution. Plant Ecol. 2022, 223, 711–732. [Google Scholar] [CrossRef]
- Fidelis, A.; Rodrigues, C.A.; Dairrel, M.; Blanco, C.C.; Pillar, V.D.; Pfadenhauer, J. What matters for vegetation regeneration in Brazilian subtropical grasslands: Seeders or resprouters? Flora 2021, 279, 151817. [Google Scholar]
- Burger, N.; Bond, W.J. Flammability traits of Cape shrubland species with different post-fire recruitment strategies. S. Afr. J. Bot. 2015, 101, 40–48. [Google Scholar]
- Fontenele, H.G.V.; Cruz-Lima, L.F.S.; Pachecho-Filho, J.L. Burning grasses, poor seeds: Post-fire reproduction of early flowering Neotropical savanna grasses produces low-quality seeds. Plant Ecol. 2020, 221, 1265–1274. [Google Scholar] [CrossRef]
- Wagenius, S.; Beck, J.; Kiefer, G. Fire synchronizes flowering and boosts reproduction in a widespread but declining prairie species. Proc. Natl. Acad. Sci. USA 2020, 117, 3000–3005. [Google Scholar] [PubMed]
- Vidaller, C.; Dutoit, T.; Ramone, H.; Bischoff, A. Fire increases the reproduction of the dominant grass Brachypodium retusum and Mediterranean steppe diversity in a combined burning and grazing experiment. Appl. Veg. Sci. 2019, 22, 127–137. [Google Scholar]
- Simpson, K.J.; Olofsson, J.K.; Ripley, B.S.; Osborne, C.P. Frequent fires prime plant developmental responses to burning. Proc. R. Soc. B 2019, 286, 20191315. [Google Scholar] [CrossRef]
- Gallagher, R.V.; Allen, S.; Mackenzie, B.D.E.; Yates, C.J.; Gosper, C.R.; Keith, D.A.; Merow, C.; White, M.D.; Wenk, E.; Maitner, B.S.; et al. High fire frequency and the impact of the 2019–2020 megafires on Australian plant diversity. Divers. Distrib. 2021, 27, 1166–1179. [Google Scholar] [CrossRef]
- Hiers, Q.A.; Treadwell, M.L.; Dickinson, M.B.; Kavanagh, K.L.; Lodge, A.G.; Starns, H.D.; Tolleson, D.R.; Twidwell, D.; Wonkka, C.L.; Rogers, W.E. Grass bud responses to fire in a semiarid savanna system. Ecol. Evol. 2021, 11, 6620–6633. [Google Scholar]
- Smith-Ramírez, C.; Castillo-Mandujano, J.; Beserra, P.; Sandoval, L.; Allende, R.; Fuentes, R. Recovery of Chilean Mediterranean vegetation after different frequencies of fires. For. Ecol. Manag. 2021, 485, 118922. [Google Scholar] [CrossRef]
- Bestelmeyer, B.T.; Burkett, L.M.; Lister, L. Effects of Managed Fire on a Swale Grassland in the Chihuahuan Desert. Rangelands 2021, 43, 181–184. [Google Scholar]
- Emery, S.M.; Gross, K.L. Effects of timing of prescribed fire on the demography of an invasive plant, Spotted Knapweed Centaurea maculosa. J. Appl. Ecol. 2005, 42, 60–69. [Google Scholar]
- Dagleish, H.J.; Ott, J.P.; Setshogo, M.P.; Hartnet, D.C. Inter-specific variation in bud banks and flowering effort among semi-arid African savanna grasses. S. Afr. J. Bot. 2012, 83, 127–133. [Google Scholar]
- Laterra, P.; Vignolio, O.R.; Linares, M.P.; Giaquinta, A.; Maceira, N. Cumulative effects of fire on a tussock pampa grassland. J. Veg. Sci. 2003, 14, 43–54. [Google Scholar] [CrossRef]
- Tangney, R.; Paroissien, R.; Le Breton, T.D.; Thomsen, A.; Doyle, C.A.T.; Ondik, M.; Miller, R.G.; Miller, B.P.; Ooi, M.K.J. Success of post-fire plant recovery strategies varies with shifting fire seasonality. Commun. Earth Environ. 2022, 3, 126. [Google Scholar] [CrossRef]
- Knapp, E.E.; Estes, B.L.; Skinner, C.N. Ecological Effects of Prescribed Fire Season: A Literature Review and Synthesis for Managers; JFSP Synthesis Reports; United States Department of Agriculture Forest Service, Pacific Southwest Research Station: Albany, CA, USA, 2009; Volume 4, Available online: http://digitalcommons.unl.edu/jfspsynthesis/4 (accessed on 5 July 2023).
- Gonzalez, T.M.; Gonzalez-Trujillo, J.D.; Munoz, A.; Armenteras, D. Effects of fre history on animal communities: A systematic review. Ecol. Process. 2022, 11, 11. [Google Scholar] [CrossRef]
- Christie, A.P.; Amano, T.; Martin, P.A.; Shackelford, G.E.; Simmons, B.I.; Sutherland, W.J. Simple study designs in ecology produce inaccurate estimates of biodiversity responses. J. Appl. Ecol. 2019, 56, 2742–2754. [Google Scholar] [CrossRef]
- Mndela, M.; Tjelele, J.T.; Madakadze, I.C.; Mangwane, M.; Samuels, I.M.; Muller, F.; Pule, H.T. A global meta-analysis of woody plant responses to elevated CO2: Implications on biomass, growth, leaf N content, photosynthesis, and water relations. Ecol. Process. 2022, 11, 52. [Google Scholar] [CrossRef]
- Snyman, H.A. Short-term influence of fire on seedling establishment in a semi-arid grassland of South Africa. S. Afr. J. Bot. 2004, 70, 215–226. [Google Scholar]
- Ithurrat, L.S.; Busso, C.A.; Torres, Y.A.; Giorgeti, H.D.; Rodriguez, G.D.; Ambrosino, M.L. Adequate management of post-fire defoliation would not affect the metabolic activity of axillary buds in grasses. Ann. Bot. Fennici. 2019, 56, 267–276. [Google Scholar]
- Brown, J.K.; Smith, J.K. (Eds.) Wildland Fire in Ecosystems: Effects of Fire on Flora; Gen. Tech. Rep. RMRS-GTR-42-vol. 2; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Ogden, UT, USA, 2000; 257p. [Google Scholar]
- Russell, M.L.; Vermeire, L.T.; Ganguli, A.C.; Hendrickson, J.R. Fire Return Interval and Season of Fire Alter Bud Banks. Range Ecol. Manag. 2019, 72, 542–550. [Google Scholar]
- Simpson, K.J.; Jardine, E.C.; Archibald, S.; Forrestel, E.J.; Lehmann, C.E.R.; Thomas, G.H.; Osborne, C.P. Resprouting grasses are associated with less frequent fire than seeders. New Phytol. 2021, 230, 832–844. [Google Scholar]
- Richardson, L.K.; Wagenius, S. Fire influences reproductive outcomes by modifying flowering phenology and mate-availability. New Phytol. 2022, 233, 2083–2093. [Google Scholar] [CrossRef]
- Liu, F.; Chen, J.M.; Wang, Q.F. Trade-offs between sexual and asexual reproduction in a monoecious species Sagittaria pygmaea (Alismataceae): The effect of different nutrient levels. Plant Syst. Evol. 2009, 277, 61–65. [Google Scholar]
- Day, N.J. When do grasses resprout after fire? New Phytol. 2021, 230, 406–407. [Google Scholar] [PubMed]
- Baruzzi, C.; Medina-Irizarry, N.; Armstrong, M.N.; Crandall, R.M. Fire season and canopy cover interact to affect reproduction of a pyrogenic bunchgrass, Aristida beyrichiana. Plant Ecol. 2022, 223, 263–272. [Google Scholar]
- Bowen, B.J.; Pate, J.S. Effect of season of burn on shoot recovery and post-fire flowering performance in the resprouter Stirlingia latifolia R. Br. (Proteaceae). Austral Ecol. 2004, 29, 145–155. [Google Scholar]
- Kross, C.S.; Rohli, R.V.; Moon, J.A.; Fournier, A.M.V.; Woodrey, M.S.; Nyman, J.A. Preferred atmospheric circulations associated with favorable prescribed burns in the Gulf of Mexico coast, USA. Fire Ecol. 2023, 19, 7. [Google Scholar] [CrossRef]
- Cao, W.; Rohli, R.V.; Han, F.; Vega, A.J.; Bushra, N.; Nyman, J.A. Atmospheric circulation regimes for prescribed burns along the U.S. Gulf of Mexico coast. Appl. Geogr. 2021, 136, 102587. [Google Scholar] [CrossRef]
- Smith, M.D.; van Wilgen, B.W.; Burns, C.E.; Govender, N.; Potgieter, A.L.F.; Andelman, S.; Biggs, H.C.; Botha, J.; Trollope, W.S.W. Long-term effects of fire frequency and season on herbaceous vegetation in savannas of the Kruger National Park, South Africa. J. Plant Ecol. 2013, 6, 71–83. [Google Scholar] [CrossRef]
- Enright, N.J.; Fontaine, J.B.; Lamont, B.B.; Miller, B.P.; Westcott, V.C. Resistance and resilience to changing climate and fire regime depend on plant functional traits. J. Ecol. 2014, 102, 1572–1581. [Google Scholar]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).