
Short-Term Effects of Prescribed Burn Seasonality on the Understory in a Pinus nigra Arnold subsp. salzmannii (Dunal) Franco Mediterranean Forest

 ,
,  ,
,  , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Experimental Design
Plots
2.3. Prescribed Burnings
2.4. Vegetation Indicators
2.4.1. Plant Coverage and α-Diversity
2.4.2. Life Form and Fire-Adapted Traits
2.5. Statistical Analyses
3. Results
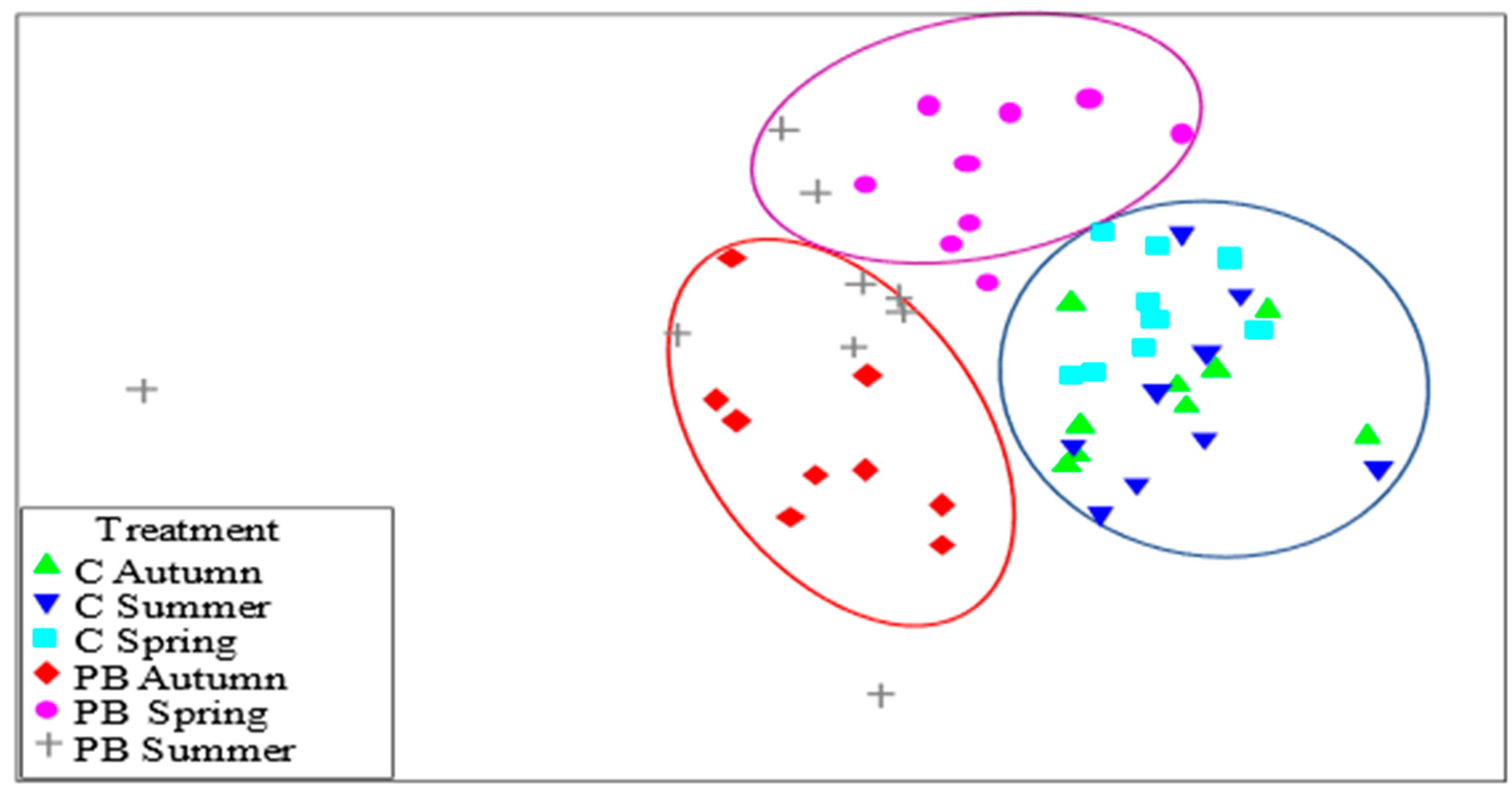
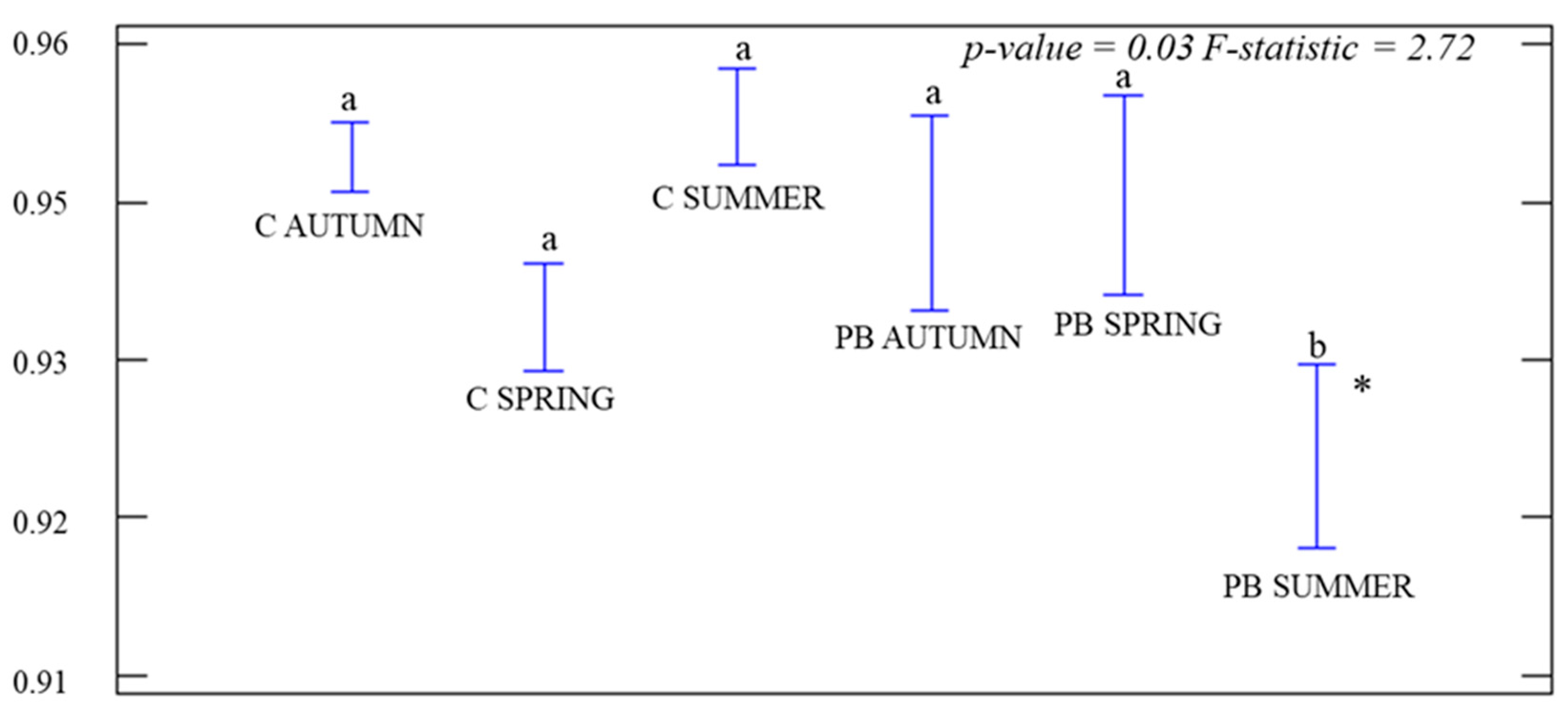
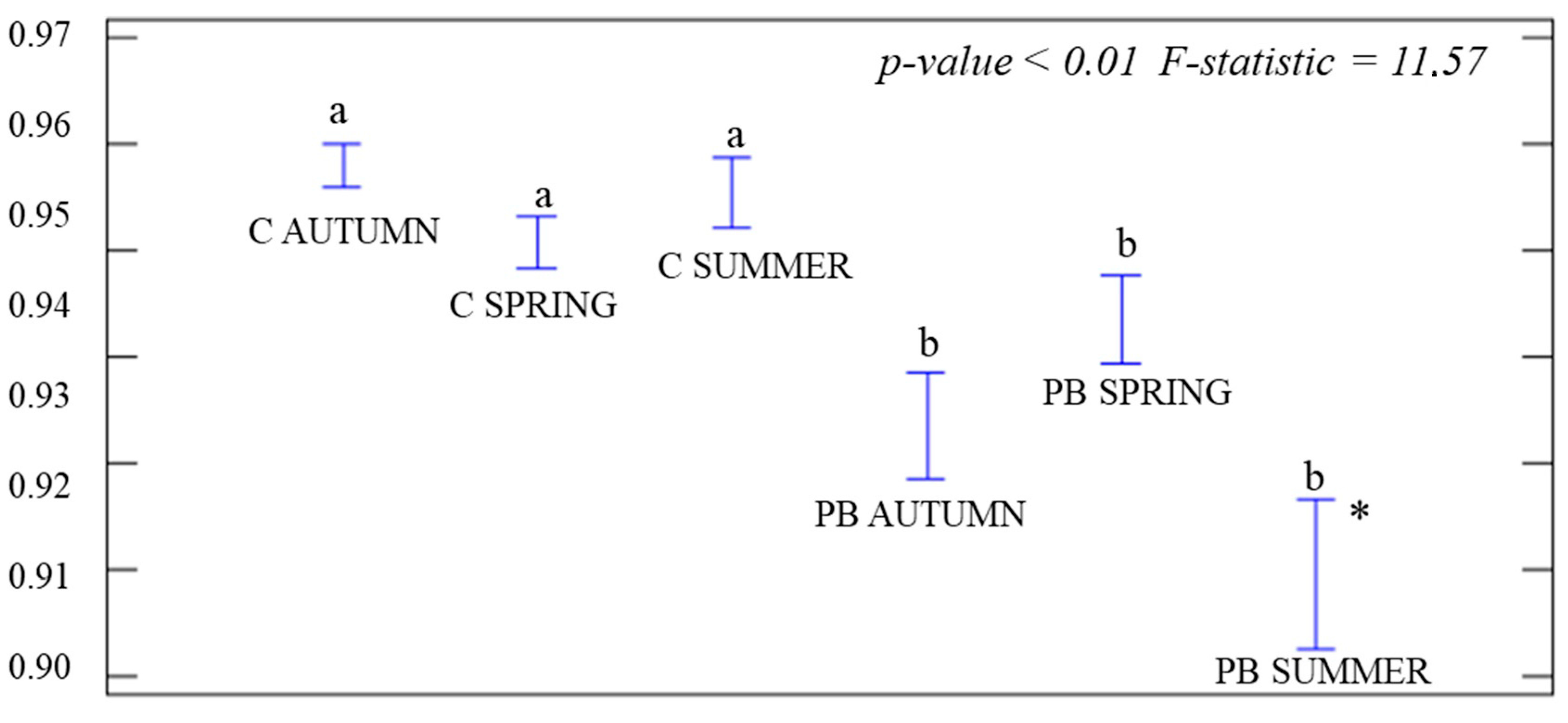
3.1. Analysis of Coverage Vegetation and α-Diversity
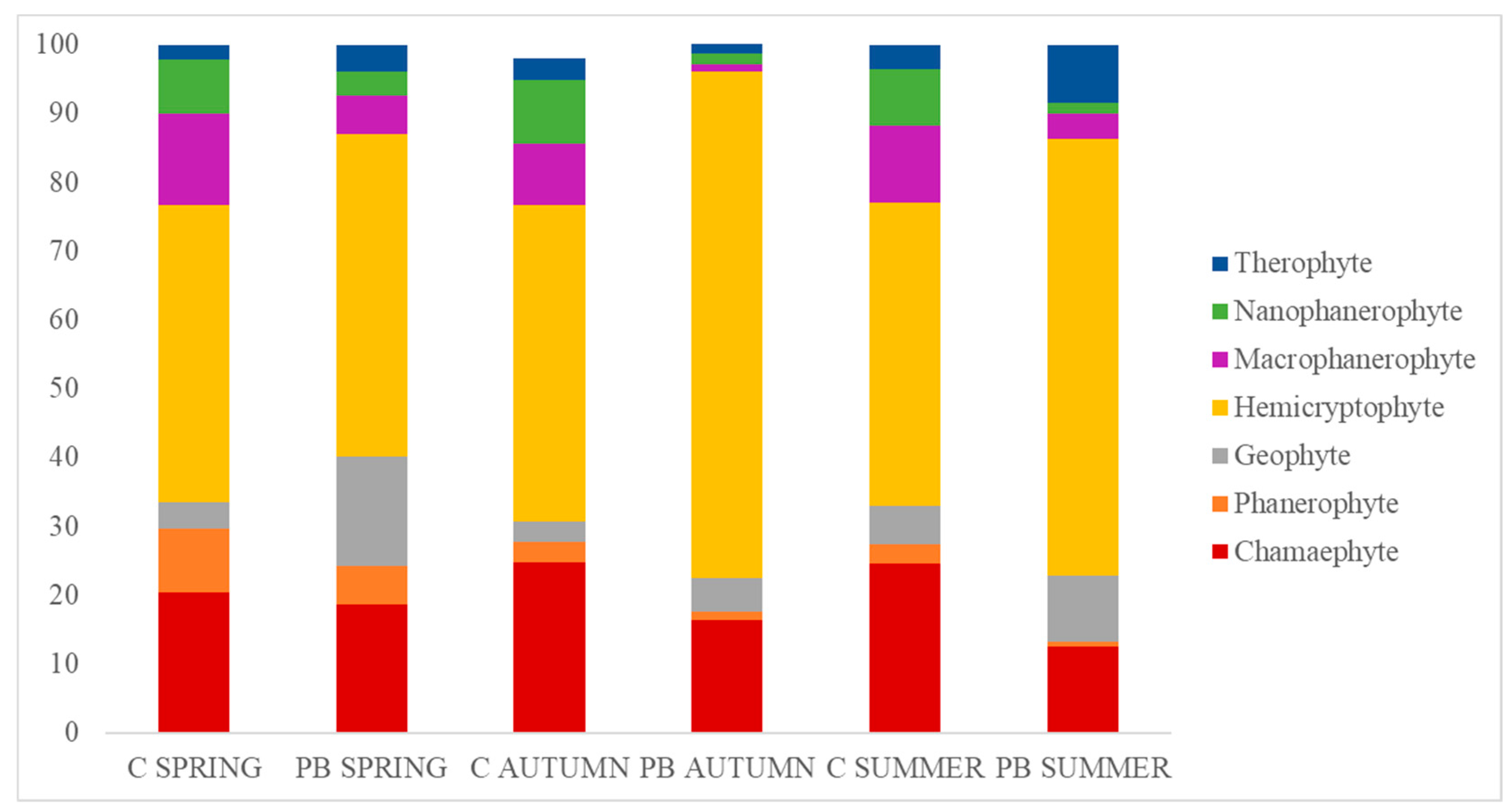
3.2. Analysis of Life Forms and Fire-Adapted Traits Classification
4. Discussion
4.1. Coverage Vegetation and α-Diversity
4.2. Raunkiaer and Fire-Adapted Traits Classification
4.3. Future Evaluations on the Effects of Preventive Treatments in Forest Management
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cardinale, B.J.; Duffy, J.E.; Gonzalez, A.; Hooper, D.U.; Perrings, C.; Venail, P.; Narwani, A.; Mace, G.M.; Tilman, D.; Wardle, D.A.; et al. Biodiversity loss and its impact on humanity. Nature 2012, 486, 59–67. [Google Scholar] [CrossRef]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; da Fonseca, G.A.B.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef]
- Tittensor, D.P.; Walpole, M.; Hill, S.L.L.; Boyce, D.G.; Britten, G.L.; Burgess, N.D.; Butchart, S.H.M.; Leadley, P.W.; Regan, E.C.; Alkemade, R.; et al. A mid-term analysis of progress toward international biodiversity targets. Science 2014, 346, 241–244. [Google Scholar] [CrossRef]
- Poorter, L.; van der Sande, M.T.; Thompson, J.; Arets, E.J.M.M.; Alarcón, A.; Álvarez-Sánchez, J.; Ascarrunz, N.; Balvanera, P.; Barajas-Guzmán, G.; Boit, A.; et al. Diversity enhances carbon storage in tropical forests. Glob. Ecol. Biogeogr. 2015, 24, 1314–1328. [Google Scholar] [CrossRef]
- Cassell, B.A.; Scheller, R.M.; Lucash, M.S.; Hurteau, M.D.; Loudermilk, E.L. Widespread severe wildfires under climate change lead to increased forest homogeneity in dry mixed-conifer forests. Ecosphere 2019, 10, e02934. [Google Scholar] [CrossRef]
- Roces-Díaz, J.V.; Santín, C.; Martínez-Vilalta, J.; Doerr, S.H. A global synthesis of fire effects on ecosystem services of forests and woodlands. Front. Ecol. Environ. 2022, 20, 170–178. [Google Scholar] [CrossRef]
- Anav, A.; Mariotti, A. Sensitivity of natural vegetation to climate change in the Euro-Mediterranean area. Clim. Res. 2011, 46, 277–292. [Google Scholar] [CrossRef]
- Cantón, Y.; Solé-Benet, A.; de Vente, J.; Boix-Fayos, C.; Calvo-Cases, A.; Asensio, C.; Puigdefábregas, J. A review of runoff generation and soil erosion across scales in semiarid south-eastern Spain. J. Arid. Environ. 2011, 75, 1254–1261. [Google Scholar] [CrossRef]
- Barredo, J.I.; Caudullo, G.; Dosio, A. Mediterranean habitat loss under future climate conditions: Assessing impacts on the Natura 2000 protected area network. Appl. Geogr. 2016, 75, 83–92. [Google Scholar] [CrossRef]
- Peñuelas, J.; Sardans, J. Global Change and Forest Disturbances in the Mediterranean Basin: Breakthroughs, Knowledge Gaps, and Recommendations. Forests 2021, 12, 603. [Google Scholar] [CrossRef]
- Halofsky, J.E.; Peterson, D.L.; Harvey, B.J. Changing wildfire, changing forests: The effects of climate change on fire regimes and vegetation in the Pacific Northwest, USA. Fire Ecol. 2020, 16, 4. [Google Scholar] [CrossRef]
- Chick, M.P.; Nitschke, C.R.; Cohn, J.S.; Penman, T.D.; York, A. Factors influencing above-ground and soil seed bank vegetation diversity at different scales in a quasi-Mediterranean ecosystem. J. Veg. Sci. 2018, 29, 684–694. [Google Scholar] [CrossRef]
- Barbéro, M.; Loisel, R.; Quézel, P.; Richardson, D.; Romane, F. Pines of the Mediterranean basin. In Ecology and Biogeography of Pinus; Cambridge University Press: Cambridge, UK, 1998; p. 8. [Google Scholar]
- Change, M.E.C.; Cramer, W.; Guiot, J.; Marini, K.; Azzopardi, B.; Balzan, M.V.; Cherif, S.; Doblas-Miranda, E.; dos Santos, M.; Dobrinski, P. Climate and Environmental Change in the Mediterranean Basin—Current Situation and Risks for the Future; First Mediterranean Assessment Report; MedECC: Lisbon, Portugal, 2020. [Google Scholar]
- EFFIS. Forest Fires in Europe, Middle East and North Africa 2021. 2023. Available online: https://effis.jrc.ec.europa.eu/ (accessed on 12 July 2023).
- Vadilonga, T.; Úbeda, X.; Germann, P.F.; Lorca, M. Effects of prescribed burnings on soil hydrological parameters. Hydrol. Process. 2008, 22, 4249–4256. [Google Scholar] [CrossRef]
- Pausas, J.G.; Keeley, J.E. Wildfires and global change. Front. Ecol. Environ. 2021, 19, 387–395. [Google Scholar] [CrossRef]
- Clark, D.A.; Brown, S.; Kicklighter, D.W.; Chambers, J.Q.; Thomlinson, J.R.; Ni, J. Measuring Net Primary Production in Forests: Concepts and Field Methods. Ecol. Appl. 2001, 11, 356–370. [Google Scholar] [CrossRef]
- Vázquez-Veloso, A.; Dejene, T.; Oria-de-Rueda, J.A.; Guijarro, M.; Hernando, C.; Espinosa, J.; Madrigal, J.; Martín-Pinto, P. Prescribed burning in spring or autumn did not affect the soil fungal community in Mediterranean Pinus nigra natural forests. For. Ecol. Manag. 2022, 512, 120161. [Google Scholar] [CrossRef]
- Reinking, D.L. Fire regimes and avian responses in the central tallgrass prairie. Stud. Avian Biol. 2005, 30, 116. [Google Scholar]
- García-Llamas, P.; Suárez-Seoane, S.; Fernández-Manso, A.; Quintano, C.; Calvo, L. Evaluation of fire severity in fire prone-ecosystems of Spain under two different environmental conditions. J. Environ. Manag. 2020, 271, 110706. [Google Scholar] [CrossRef]
- Bowd, E.J.; Egidi, E.; Lindenmayer, D.B.; Wardle, D.A.; Kardol, P.; Cary, G.J.; Foster, C. Direct and indirect effects of fire on microbial communities in a pyrodiverse dry-sclerophyll forest. J. Ecol. 2022, 110, 1687–1703. [Google Scholar] [CrossRef]
- Quevedo, L.; Rodrigo, A.; Espelta, J.M. Post-fire resprouting ability of 15 non-dominant shrub and tree species in Mediterranean areas of NE Spain. Ann. For. Sci. 2007, 64, 883–890. [Google Scholar] [CrossRef]
- Moreira, F.; Ascoli, D.; Safford, H.; Adams, M.A.; Moreno, J.M.; Pereira, J.M.C.; Catry, F.X.; Armesto, J.; Bond, W.; González, M.E.; et al. Wildfire management in Mediterranean-type regions: Paradigm change needed. Environ. Res. Lett. 2020, 15, 011001. [Google Scholar] [CrossRef]
- Lucas-Borja, M.E.; Heydari, M.; Miralles, I.; Zema, D.A.; Manso, R. Effects of Skidding Operations after Tree Harvesting and Soil Scarification by Felled Trees on Initial Seedling Emergence of Spanish Black Pine (Pinus nigra Arn. ssp. salzmannii). Forests 2020, 11, 767. [Google Scholar] [CrossRef]
- Valor, T.; Battipaglia, G.; Piqué, M.; Altieri, S.; González-Olabarria, J.R.; Casals, P. The effect of prescribed burning on the drought resilience of Pinus nigra ssp. salzmannii Dunal (Franco) and P. sylvestris L. Ann. For. Sci. 2020, 77, 13. [Google Scholar] [CrossRef]
- del Cerro Barja, A.; Borja, M.L.; García, E.M.; Serrano, F.L.; Abellán, M.A.; Morote, F.G.; López, R.N. Influence of stand density and soil treatment on the Spanish Black Pine (Pinus nigra Arn. ssp. Salzmannii) regeneration in Spain. For. Syst. 2009, 18, 167–180. [Google Scholar]
- Martín-Alcón, S.; Coll, L. Unraveling the relative importance of factors driving post-fire regeneration trajectories in non-serotinous Pinus nigra forests. For. Ecol. Manag. 2016, 361, 13–22. [Google Scholar] [CrossRef]
- Keane, R.E.; Agee, J.K.; Fulé, P.; Keeley, J.E.; Key, C.; Kitchen, S.G.; Miller, R.; Schulte, L.A.; Keane, R.E.; Agee, J.K.; et al. Ecological effects of large fires on US landscapes: Benefit or catastrophe? Int. J. Wildland Fire 2008, 17, 696–712. [Google Scholar] [CrossRef]
- Pausas, J.G.; Carbó, E.; Neus Caturla, R.; Gil, J.M.; Vallejo, R. Post-fire regeneration patterns in the eastern Iberian Peninsula. Acta Oecol. 1999, 20, 499–508. [Google Scholar] [CrossRef]
- Raunkiaer, C. The Life Forms of Plants and Statistical Plant Geography; Being the Collected Papers of C. Raunkiaer; Clarendon Press: Oxford, UK, 1934. [Google Scholar]
- Teixeira, A.M.C.; Curran, T.J.; Jameson, P.E.; Meurk, C.D.; Norton, D.A. Post-Fire Resprouting in New Zealand Woody Vegetation: Implications for Restoration. Forests 2020, 11, 269. [Google Scholar] [CrossRef]
- Martinez, J.L.; Lucas-Borja, M.E.; Plaza-Alvarez, P.A.; Denisi, P.; Moreno, M.A.; Hernández, D.; González-Romero, J.; Zema, D.A. Comparison of Satellite and Drone-Based Images at Two Spatial Scales to Evaluate Vegetation Regeneration after Post-Fire Treatments in a Mediterranean Forest. Appl. Sci. 2021, 11, 5423. [Google Scholar] [CrossRef]
- Carrillo-García, C.; Girola-Iglesias, L.; Guijarro, M.; Hernando, C.; Madrigal, J.; Mateo, R.G. Ecological niche models applied to post-megafire vegetation restoration in the context of climate change. Sci. Total Environ. 2023, 855, 158858. [Google Scholar] [CrossRef]
- Pauli, H.; Gottfried, M.; Dullinger, S.; Abdaladze, O.; Akhalkatsi, M.; Alonso, J.L.B.; Coldea, G.; Dick, J.; Erschbamer, B.; Calzado, R.F.; et al. Recent Plant Diversity Changes on Europe’s Mountain Summits. Science 2012, 336, 353–355. [Google Scholar] [CrossRef]
- Pausas, J.G.; Keeley, J.E. Epicormic Resprouting in Fire-Prone Ecosystems. Trends Plant Sci. 2017, 22, 1008–1015. [Google Scholar] [CrossRef]
- Rabin, S.S.; Gérard, F.N.; Arneth, A. The influence of thinning and prescribed burning on future forest fires in fire-prone regions of Europe. Environ. Res. Lett. 2022, 17, 055010. [Google Scholar] [CrossRef]
- Ewing, A.L.; Engle, D.M. Effects of Late Summer Fire on Tallgrass Prairie Microclimate and Community Composition. Am. Midl. Nat. 1988, 120, 212–223. [Google Scholar] [CrossRef]
- Moghli, A.; Santana, V.M.; Baeza, M.J.; Pastor, E.; Soliveres, S. Fire Recurrence and Time Since Last Fire Interact to Determine the Supply of Multiple Ecosystem Services by Mediterranean Forests. Ecosystems 2022, 25, 1358–1370. [Google Scholar] [CrossRef]
- Keeley, J.E.; Pausas, J.G.; Keeley, J.E.; Pausas, J.G. Distinguishing disturbance from perturbations in fire-prone ecosystems. Int. J. Wildland Fire 2019, 28, 282–287. [Google Scholar] [CrossRef]
- Stevens, J.T.; Haffey, C.M.; Coop, J.D.; Fornwalt, P.J.; Yocom, L.; Allen, C.D.; Bradley, A.; Burney, O.T.; Carril, D.; Chambers, M.E.; et al. Tamm Review: Postfire landscape management in frequent-fire conifer forests of the southwestern United States. For. Ecol. Manag. 2021, 502, 119678. [Google Scholar] [CrossRef]
- Thomsen, A.M.; Ooi, M.K.J. Shifting season of fire and its interaction with fire severity: Impacts on reproductive effort in resprouting plants. Ecol. Evol. 2022, 12, e8717. [Google Scholar] [CrossRef] [PubMed]
- Kottek, M.; Grieser, J.; Beck, C.; Rudolf, B.; Rubel, F. World Map of the Köppen-Geiger climate classification updated. Meteorol. Z. 2006, 15, 259–263. [Google Scholar] [CrossRef]
- AEMET. Weather Data from Beteta Station (Cuenca-Spain) 2000–2022; State Meteorological Agency of Spanish Government (AEMET): Madrid, Spain, 2022; Available online: http://www.aemet.es/ (accessed on 3 May 2022).
- IGN. Mapa de Suelos de España: Escala 1:1.000.000; Instituto Geográfico Nacional: Madrid, Spain, 2006. [Google Scholar]
- Working Group WRB IUSS. Base Referencial Mundial del Recurso Suelo 2014, Actualización 2015. Informes Sobre Recursos Mundiales de Suelos 106, FAO, Roma. 2016; ISBN 978-92-5-308369-5. Available online: https://www.fao.org/home/en/ (accessed on 13 April 2022).
- Espinosa, J.; Martin-Benito, D.; Rodríguez de Rivera, Ó.; Hernando, C.; Guijarro, M.; Madrigal, J. Tree Growth Response to Low-Intensity Prescribed Burning in Pinus nigra Stands: Effects of Burn Season and Fire Severity. Appl. Sci. 2021, 11, 7462. [Google Scholar] [CrossRef]
- Blaser, S.; Prati, D.; Senn-Irlet, B.; Fischer, M. Effects of forest management on the diversity of deadwood-inhabiting fungi in Central European forests. For. Ecol. Manag. 2013, 304, 42–48. [Google Scholar] [CrossRef]
- Rossetti, I.; Cogoni, D.; Calderisi, G.; Fenu, G. Short-Term Effects and Vegetation Response after a Megafire in a Mediterranean Area. Land 2022, 11, 2328. [Google Scholar] [CrossRef]
- Alizoti, P.G.; Kilimis, K.; Gallios, P. Temporal and spatial variation of flowering among Pinus nigra Arn. clones under changing climatic conditions. For. Ecol. Manag. 2010, 259, 786–797. [Google Scholar] [CrossRef]
- Vega, J.A.; Landsberg, J.; Bará, S.; Paysen, T.; Fontúrbel, M.T.; Alonso, M. Efectos del fuego prescrito bajo arbolado de P. Pinaster en suelos forestales de Galicia y Andalucía. Cuad. Soc. Esp. Cienc. For. 2000, 9, 123–136. [Google Scholar]
- Rothermel, R.C. A Mathematical Model for Predicting Fire Spread in Wildland Fuels; Research Paper INT-116 1972; US Department of Agriculture, Intermountain Forest and Range Experiment Station: Ogden, UT, USA, 1972; 40p.
- Albini, F.A. Estimating Wildfire Behavior and Effects; General Technical Reports INT-GTR-30; U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station: Ogden, UT, USA, 1976; Volume 30, 92p.
- Canfield, R.H. Application of the Line Interception Method in Sampling Range Vegetation. J. For. 1941, 39, 388–394. [Google Scholar]
- Pokswinski, S.; Gallagher, M.R.; Skowronski, N.S.; Loudermilk, E.L.; Hawley, C.; Wallace, D.; Everland, A.; Wallace, J.; Hiers, J.K. A simplified and affordable approach to forest monitoring using single terrestrial laser scans and transect sampling. MethodsX 2021, 8, 101484. [Google Scholar] [CrossRef]
- Magurran, A.E. Ecological Diversity and Its Measurement; Springer: Dordrecht, The Netherlands, 1988. [Google Scholar]
- Simpson, E.H. Measurement of Diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Shannon, C.E. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef]
- Paula, S.; Arianoutsou, M.; Kazanis, D.; Tavsanoglu, Ç.; Lloret, F.; Buhk, C.; Ojeda, F.; Luna, B.; Moreno, J.M.; Rodrigo, A.; et al. Fire-related traits for plant species of the Mediterranean Basin. Ecology 2009, 90, 1420. [Google Scholar] [CrossRef]
- Fares, S.; Bajocco, S.; Salvati, L.; Camarretta, N.; Dupuy, J.-L.; Xanthopoulos, G.; Guijarro, M.; Madrigal, J.; Hernando, C.; Corona, P. Characterizing potential wildland fire fuel in live vegetation in the Mediterranean region. Ann. For. Sci. 2017, 74, 1. [Google Scholar] [CrossRef]
- Tavşanoğlu, Ç.; Pausas, J.G. A functional trait database for Mediterranean Basin plants. Sci. Data 2018, 5, 180135. [Google Scholar] [CrossRef] [PubMed]
- Gachet, S.; Véla, E.; Tatoni, T. BASECO: A floristic and ecological database of Mediterranean French flora. Biodivers. Conserv. 2005, 14, 1023–1034. [Google Scholar] [CrossRef]
- Santana, V.M.; Alday, J.G.; Baeza, M.J. Effects of fire regime shift in Mediterranean Basin ecosystems: Changes in soil seed bank composition among functional types. Plant Ecol. 2014, 215, 555–566. [Google Scholar] [CrossRef]
- Manseau, M.; Huot, J.; Crete, M. Effects of Summer Grazing by Caribou on Composition and Productivity of Vegetation: Community and Landscape Level. J. Ecol. 1996, 84, 503–513. [Google Scholar] [CrossRef]
- Elliott, K.J.; Hendrick, R.L.; Major, A.E.; Vose, J.M.; Swank, W.T. Vegetation dynamics after a prescribed fire in the southern Appalachians. For. Ecol. Manag. 1999, 114, 199–213. [Google Scholar] [CrossRef]
- Grant, C.D.; Loneragan, W.A. The effects of burning on the understorey composition of rehabilitated bauxite mines in Western Australia: Community changes and vegetation succession. For. Ecol. Manag. 2001, 145, 255–279. [Google Scholar] [CrossRef]
- Chu, C.-J.; Maestre, F.T.; Xiao, S.; Weiner, J.; Wang, Y.-S.; Duan, Z.-H.; Wang, G. Balance between facilitation and resource competition determines biomass–density relationships in plant populations. Ecol. Lett. 2008, 11, 1189–1197. [Google Scholar] [CrossRef]
- Heydari, M.; Faramarzi, M.; Pothier, D. Post-fire recovery of herbaceous species composition and diversity, and soil quality indicators one year after wildfire in a semi-arid oak woodland. Ecol. Eng. 2016, 94, 688–697. [Google Scholar] [CrossRef]
- Hashemi, A.; Aghbash, F.G.; Zarafshar, M.; Bazot, S. 80-years livestock transit impact on permanent path soil in Zagros oak forest, Iran. Appl. Soil Ecol. 2019, 138, 189–194. [Google Scholar] [CrossRef]
- Valdez, C.G.; Guzmán, M.A.; Valdés, A.; Forougbakhch, R.; Alvarado, M.A.; Rocha, A.; Valdez, C.G.; Guzmán, M.A.; Valdés, A.; Forougbakhch, R.; et al. Estructura y diversidad de la vegetación en un matorral espinoso prístino de Tamaulipas, México. Rev. Biol. Trop. 2018, 66, 1674–1682. [Google Scholar] [CrossRef]
- Mattingly, W.B.; Orrock, J.L.; Collins, C.D.; Brudvig, L.A.; Damschen, E.I.; Veldman, J.W.; Walker, J.L. Historical agriculture alters the effects of fire on understory plant beta diversity. Oecologia 2015, 177, 507–518. [Google Scholar] [CrossRef]
- Mahood, A.L.; Balch, J.K. Repeated fires reduce plant diversity in low-elevation Wyoming big sagebrush ecosystems (1984–2014). Ecosphere 2019, 10, e02591. [Google Scholar] [CrossRef]
- Moya, D.; Sagra, J.; Lucas-Borja, M.E.; Plaza-Álvarez, P.A.; González-Romero, J.; De Las Heras, J.; Ferrandis, P. Post-Fire Recovery of Vegetation and Diversity Patterns in Semiarid Pinus halepensis Mill. Habitats after Salvage Logging. Forests 2020, 11, 1345. [Google Scholar] [CrossRef]
- Novak, L.; Scholl, J.P.; Kiefer, G.; Iler, A.M. Prescribed burning has limited effects on the population dynamics of rare plants. Conserv. Sci. Pract. 2022, 4, e12792. [Google Scholar] [CrossRef]
- Vidal-Cordero, J.M.; Angulo, E.; Molina, F.P.; Boulay, R.; Cerdá, X. Long-term recovery of Mediterranean ant and bee communities after fire in southern Spain. Sci. Total Environ. 2023, 887, 164132. [Google Scholar] [CrossRef] [PubMed]
- Valkó, O.; Deák, B. Increasing the potential of prescribed burning for the biodiversity conservation of European grasslands. Curr. Opin. Environ. Sci. Health 2021, 22, 100268. [Google Scholar] [CrossRef]
- Hanes, T.L. Succession after Fire in the Chaparral of Southern California. Ecol. Monogr. 1971, 41, 27–52. [Google Scholar] [CrossRef]
- DeJong, T.M. A Comparison of Three Diversity Indices Based on Their Components of Richness and Evenness. Oikos 1975, 26, 222–227. [Google Scholar] [CrossRef]
- James, V.; Frankie, L.; Kumar, R.; Karan, N.M. Species-area and species-individual relationships for tropical trees. J. Ecol. 1996, 84, 549–562. [Google Scholar]
- Fuerst-Bjeliš, B. Mediterranean Identities: Environment, Society, Culture; IntechOpen: London, UK, 2017. [Google Scholar]
- Myers, J.A.; Chase, J.M.; Crandall, R.M.; Jiménez, I. Disturbance alters beta-diversity but not the relative importance of community assembly mechanisms. J. Ecol. 2015, 103, 1291–1299. [Google Scholar] [CrossRef]
- Strand, E.K.; Satterberg, K.L.; Hudak, A.T.; Byrne, J.; Khalyani, A.H.; Smith, A.M.S. Does burn severity affect plant community diversity and composition in mixed conifer forests of the United States Intermountain West one decade post fire? Fire Ecol. 2019, 15, 25. [Google Scholar] [CrossRef]
- Moradizadeh, H.; Heydari, M.; Omidipour, R.; Mezbani, A.; Prévosto, B. Ecological effects of fire severity and time since fire on the diversity partitioning, composition and niche apportionment models of post-fire understory vegetation in semi-arid oak forests of Western Iran. Ecol. Eng. 2020, 143, 105694. [Google Scholar] [CrossRef]
- Martínez-Sánchez, J.J.; Ferrandis, P.; Trabaud, L.; Galindo, R.; Franco, J.A.; Herranz, J.M. Comparative root system structure of post-fire Pinus halepensis Mill. and Cistus monspeliensis L. saplings. Plant Ecol. 2003, 168, 309–320. [Google Scholar] [CrossRef]
- Dymov, A.A.; Startsev, V.V.; Milanovsky, E.Y.; Valdes-Korovkin, I.A.; Farkhodov, Y.R.; Yudina, A.V.; Donnerhack, O.; Guggenberger, G. Soils and soil organic matter transformations during the two years after a low-intensity surface fire (Subpolar Ural, Russia). Geoderma 2021, 404, 115278. [Google Scholar] [CrossRef]
- Condit, R.; Ashton, P.S.; Baker, P.; Bunyavejchewin, S.; Gunatilleke, S.; Gunatilleke, N.; Hubbell, S.P.; Foster, R.B.; Itoh, A.; LaFrankie, J.V.; et al. Spatial Patterns in the Distribution of Tropical Tree Species. Science 2000, 288, 1414–1418. [Google Scholar] [CrossRef] [PubMed]
- Pausas, J.G.; Verdú, M. Fire Reduces Morphospace Occupation in Plant Communities. Ecology 2008, 89, 2181–2186. [Google Scholar] [CrossRef]
- Bond, W.J.; Keeley, J.E. Fire as a global ‘herbivore’: The ecology and evolution of flammable ecosystems. Trends Ecol. Evol. 2005, 20, 387–394. [Google Scholar] [CrossRef]
- Tangney, R.; Paroissien, R.; Le Breton, T.D.; Thomsen, A.; Doyle, C.A.T.; Ondik, M.; Miller, R.G.; Miller, B.P.; Ooi, M.K.J. Success of post-fire plant recovery strategies varies with shifting fire seasonality. Commun. Earth Environ. 2022, 3, 126. [Google Scholar] [CrossRef]
- Pausas, J.G.; Bond, W.J. Alternative Biome States in Terrestrial Ecosystems. Trends Plant Sci. 2020, 25, 250–263. [Google Scholar] [CrossRef]
- Ruprecht, E.; Fenesi, A.; Fodor, E.I.; Kuhn, T. Prescribed burning as an alternative management in grasslands of temperate Europe: The impact on seeds. Basic Appl. Ecol. 2013, 14, 642–650. [Google Scholar] [CrossRef]
- Pausas, J.G.; Ribeiro, E. Fire and plant diversity at the global scale. Glob. Ecol. Biogeogr. 2017, 26, 889–897. [Google Scholar] [CrossRef]
- Huerta, S.; Fernández-García, V.; Calvo, L.; Marcos, E. Soil Resistance to Burn Severity in Different Forest Ecosystems in the Framework of a Wildfire. Forests 2020, 11, 773. [Google Scholar] [CrossRef]
- Stewart, J.A.E.; van Mantgem, P.J.; Young, D.J.N.; Shive, K.L.; Preisler, H.K.; Das, A.J.; Stephenson, N.L.; Keeley, J.E.; Safford, H.D.; Wright, M.C.; et al. Effects of postfire climate and seed availability on postfire conifer regeneration. Ecol. Appl. 2021, 31, e02280. [Google Scholar] [CrossRef] [PubMed]
- Węgrzyn, M.H.; Fałowska, P.; Alzayany, K.; Waszkiewicz, K.; Dziurowicz, P.; Wietrzyk-Pełka, P. Seasonal Changes in the Photosynthetic Activity of Terrestrial Lichens and Mosses in the Lichen Scots Pine Forest Habitat. Diversity 2021, 13, 642. [Google Scholar] [CrossRef]
- Pereira-Silva, E.F.L.; Casals, P.; Sodek, L.; Delitti, W.B.C.; Vallejo, V.R. Post-fire nitrogen uptake and allocation by two resprouting herbaceous species with contrasting belowground traits. Environ. Exp. Bot. 2019, 159, 157–167. [Google Scholar] [CrossRef]
- Paroissien, R.; Ooi, M.K.J. Effects of fire season on the reproductive success of the post-fire flowerer Doryanthes excelsa. Environ. Exp. Bot. 2021, 192, 104634. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Season | Temperature Ambient (°C) | Moisture Ambient (%) | Wind Speed (km/h) | 2 cm Depth Soil Surface (°C) | Mineral Soil Surface (°C) | Organic Layer (°C) | 30 cm Above Soil Surface (°C) | With Temperatures Above 100 °C (mint) |
|---|---|---|---|---|---|---|---|---|
| Spring | 20 | 41 | 3.1 | 3.1 ± 1.3 | 28.1 ± 5.2 | 204.8 ± 11.5 | 361.7 ± 65.3 | 1 ± 0.1 |
| Autumn | 16 | 47 | 3.2 | 18 ± 0.8 | 37.1 ± 5.4 | 389.7 ± 32.1 | 702.3 ± 59.6 | 5 ± 2.1 |
| Summer | 24 | 37 | 2.3 | 26 ± 1.5 | 68.1 ± 7.1 | 591.8 ± 21.7 | 635.9 ± 42.1 | 15 ± 5.2 |
| Groups | R-Statistic | Significance Level (%) | Possible Permutations | Number ≥ Observed |
|---|---|---|---|---|
| C VS. PB AUTUMN | 0.577 | 0.1 | 24,310 | 0 |
| C VS. PB SUMMER | 0.519 | 0.1 | 24,310 | 0 |
| C VS. PB SPRING | 0.520 | 0.1 | 24,310 | 0 |
| Species | Average | Contribution % | Cum. % |
|---|---|---|---|
| C AUTUMN: Average similarity = 49.55 | |||
| Bromus erectus | 20.5 | 14.23 | 14.23 |
| Genista scorpius | 13.06 | 9.26 | 23.49 |
| Pilosella castellana | 7.41 | 5.61 | 29.1 |
| C SUMMER: Average similarity = 47.56 | |||
| Bromus erectus | 21.08 | 14.9 | 14.9 |
| Genista scorpius | 12.56 | 9.03 | 23.92 |
| Thymus bracteatus | 8.09 | 5.5 | 29.42 |
| C SPRING: Average similarity = 50.97 | |||
| Bromus erectus | 26.43 | 20.11 | 20.11 |
| Genista scorpius | 12.97 | 9.15 | 29.26 |
| Quercus fajinea | 10.18 | 7.93 | 37.19 |
| PB AUTUMN: Average similarity = 43.43 | |||
| Bromus erectus | 17.57 | 22.57 | 22.57 |
| Ononis spinosa | 9.53 | 13.55 | 36.11 |
| Geum sylvaticum | 7.83 | 8.56 | 44.67 |
| PB SPRING: Average similarity = 44.91 | |||
| Bromus erectus | 15.77 | 22.28 | 22.28 |
| Thalictrum tuberosum | 8.81 | 8.72 | 31 |
| Geum sylvaticum | 7.31 | 8.47 | 39.47 |
| PB SUMMER: Average similarity = 37.37 | |||
| Bromus erectus | 18.3 | 28.6 | 28.6 |
| Ononis spinosa | 8.28 | 14.98 | 43.58 |
| Pilosella castellana | 7.19 | 6.81 | 50.39 |
| Species | Average 1 | Average 2 | Contribution % | Cum. % |
|---|---|---|---|---|
| C AUTUMN (1) and PB AUTUMN (2): Average dissimilarity = 63.93 | ||||
| Genista scorpius | 13.06 | 2.73 | 4.67 | 4.67 |
| Ranunculus bulbosus | 0.5 | 8.81 | 3.97 | 8.63 |
| Bromus erectus | 20.5 | 17.57 | 3.93 | 12.57 |
| C SPRING (1) and PB SPRING (2): Average dissimilarity = 59.19 | ||||
| Bromus erectus | 26.43 | 15.77 | 5.44 | 5.44 |
| Rosa canina | 9.71 | 3.97 | 4.24 | 9.68 |
| Pinus nigra | 10.39 | 1.94 | 4.18 | 13.87 |
| C SUMMER (1) and PB SUMMER (2): Average dissimilarity = 68.45 | ||||
| Bromus erectus | 21.08 | 18.3 | 4.46 | 4.46 |
| Genista scorpius | 12.56 | 3.25 | 4.24 | 8.7 |
| Thymus bracteatus | 8.09 | 0 | 3.78 | 12.47 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fajardo-Cantos, Á.; Moya, D.; Lucas-Borja, M.E.; Plaza-Álvarez, P.A.; Peña-Molina, E.; González-Romero, J.; de las Heras-Ibañez, J. Short-Term Effects of Prescribed Burn Seasonality on the Understory in a Pinus nigra Arnold subsp. salzmannii (Dunal) Franco Mediterranean Forest. Fire 2023, 6, 283. https://doi.org/10.3390/fire6080283
Fajardo-Cantos Á, Moya D, Lucas-Borja ME, Plaza-Álvarez PA, Peña-Molina E, González-Romero J, de las Heras-Ibañez J. Short-Term Effects of Prescribed Burn Seasonality on the Understory in a Pinus nigra Arnold subsp. salzmannii (Dunal) Franco Mediterranean Forest. Fire. 2023; 6(8):283. https://doi.org/10.3390/fire6080283
Chicago/Turabian StyleFajardo-Cantos, Álvaro, Daniel Moya, Manuel Esteban Lucas-Borja, Pedro Antonio Plaza-Álvarez, Esther Peña-Molina, Javier González-Romero, and Jorge de las Heras-Ibañez. 2023. "Short-Term Effects of Prescribed Burn Seasonality on the Understory in a Pinus nigra Arnold subsp. salzmannii (Dunal) Franco Mediterranean Forest" Fire 6, no. 8: 283. https://doi.org/10.3390/fire6080283
APA StyleFajardo-Cantos, Á., Moya, D., Lucas-Borja, M. E., Plaza-Álvarez, P. A., Peña-Molina, E., González-Romero, J., & de las Heras-Ibañez, J. (2023). Short-Term Effects of Prescribed Burn Seasonality on the Understory in a Pinus nigra Arnold subsp. salzmannii (Dunal) Franco Mediterranean Forest. Fire, 6(8), 283. https://doi.org/10.3390/fire6080283