
Mixed-Severity Wildfire as a Driver of Vegetation Change in an Arizona Madrean Sky Island System, USA



Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
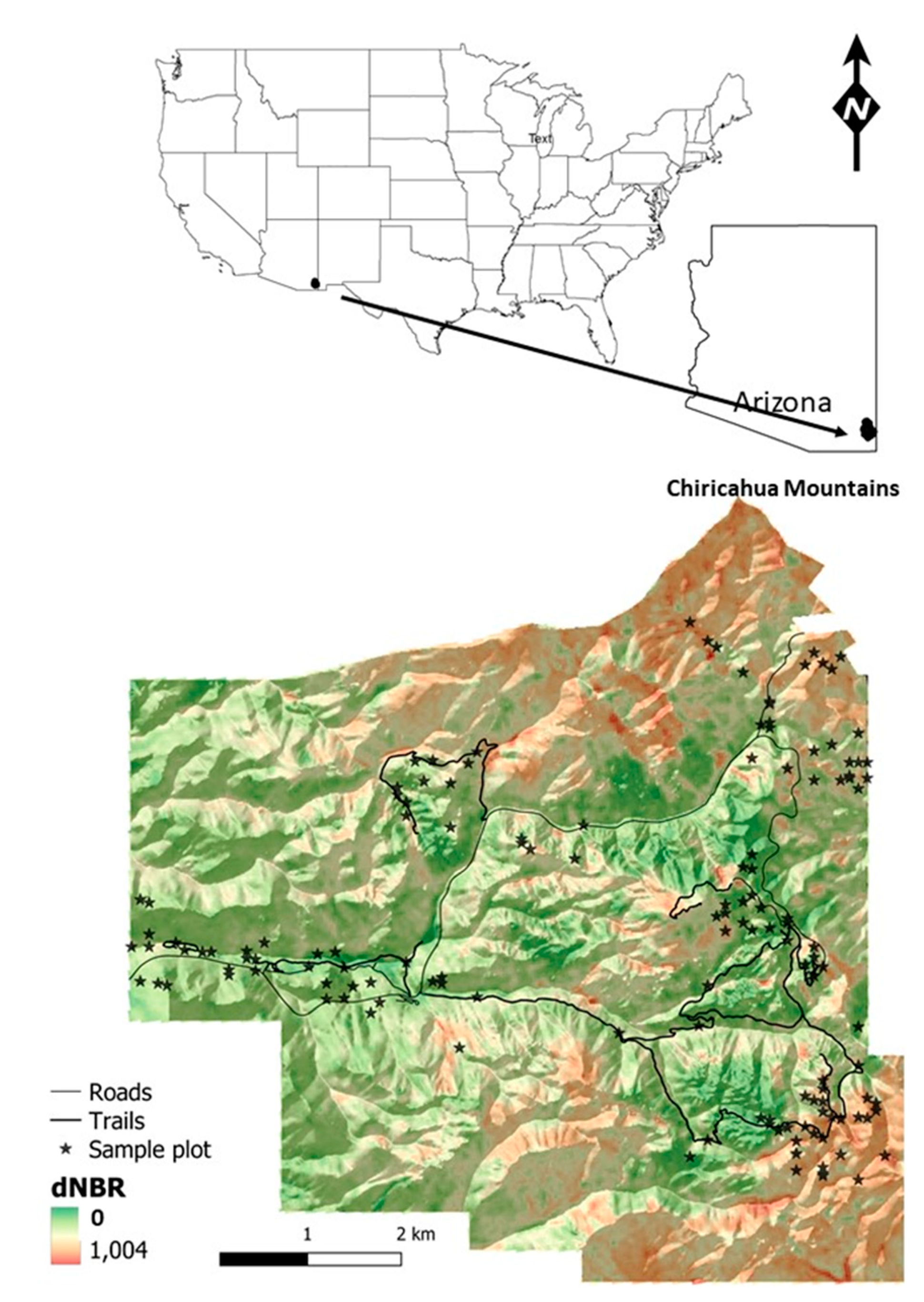
2.2. Field Sampling Methods
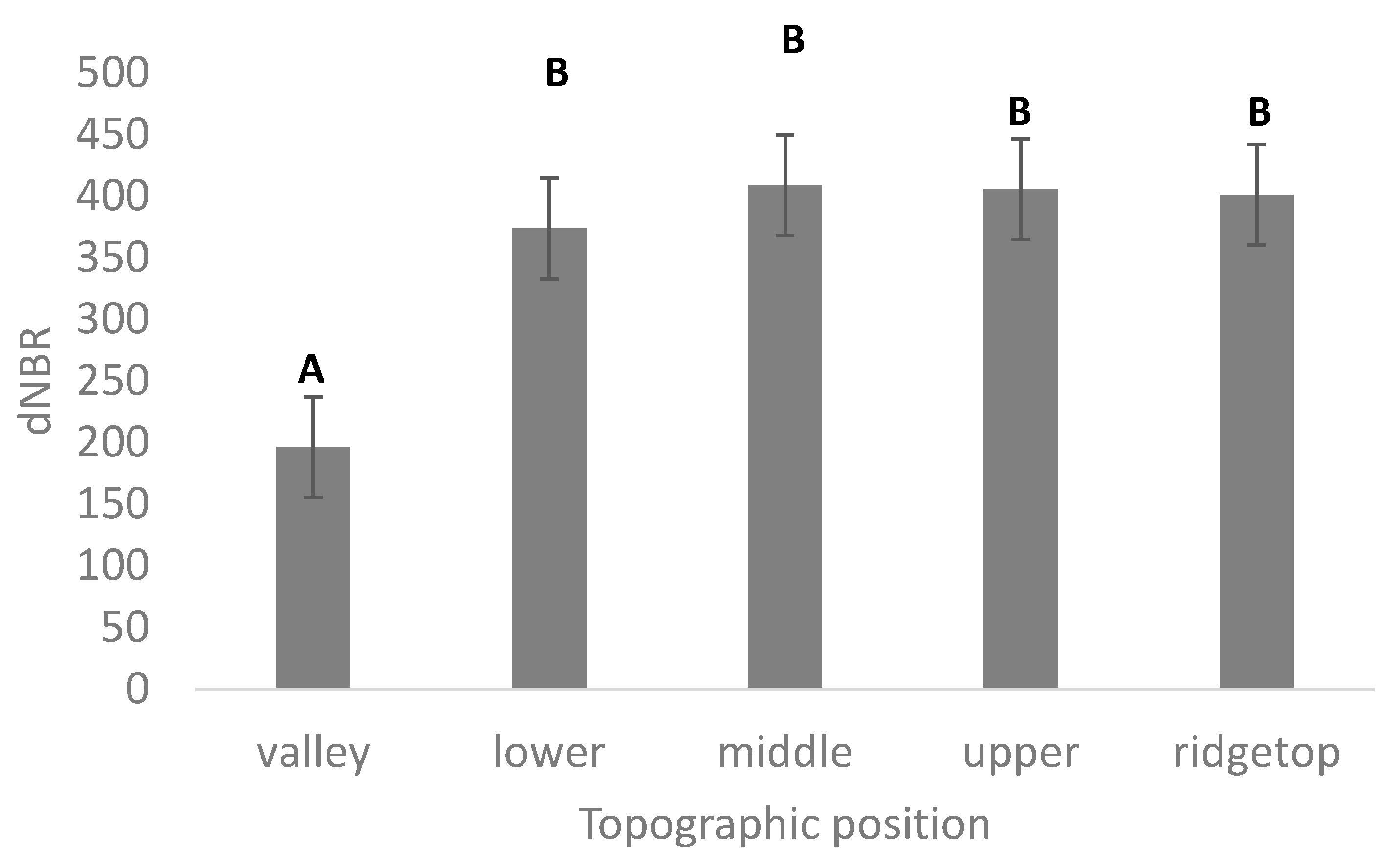
2.3. Fire Severity and Topography Data
2.4. Statistical Analyses
2.4.1. Community Compositional Shifts
2.4.2. Forest Stand Structural Changes
3. Results
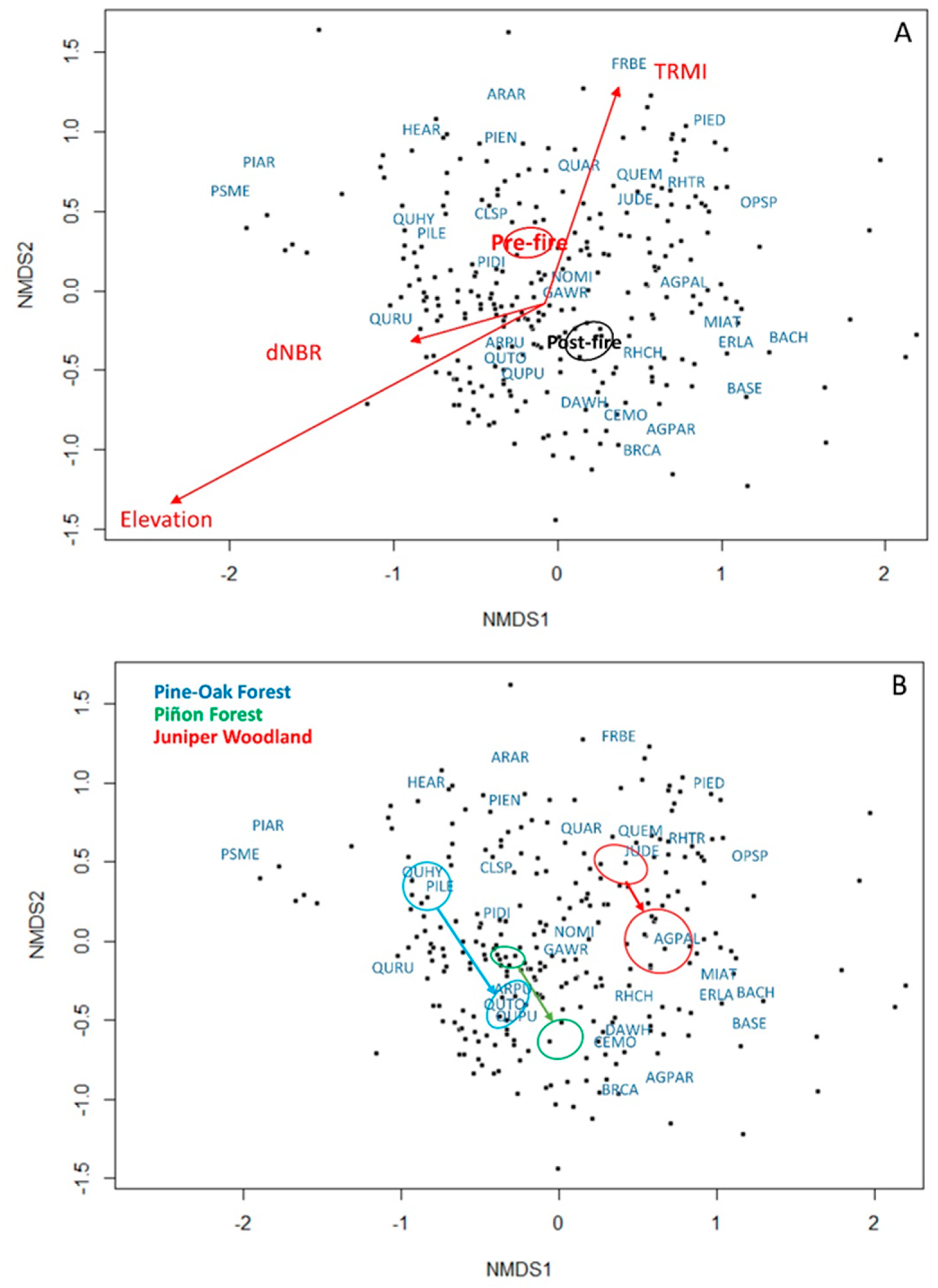
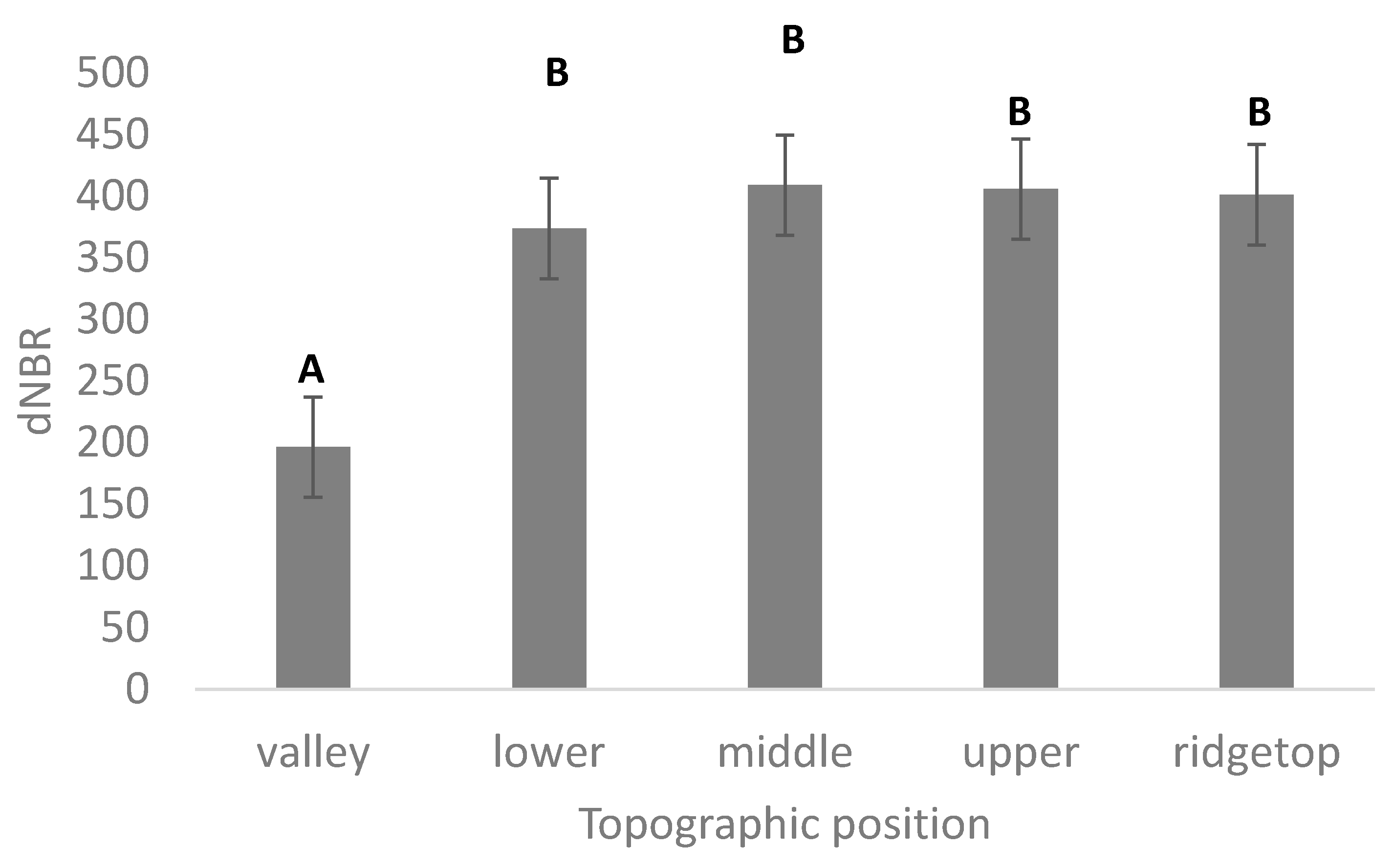
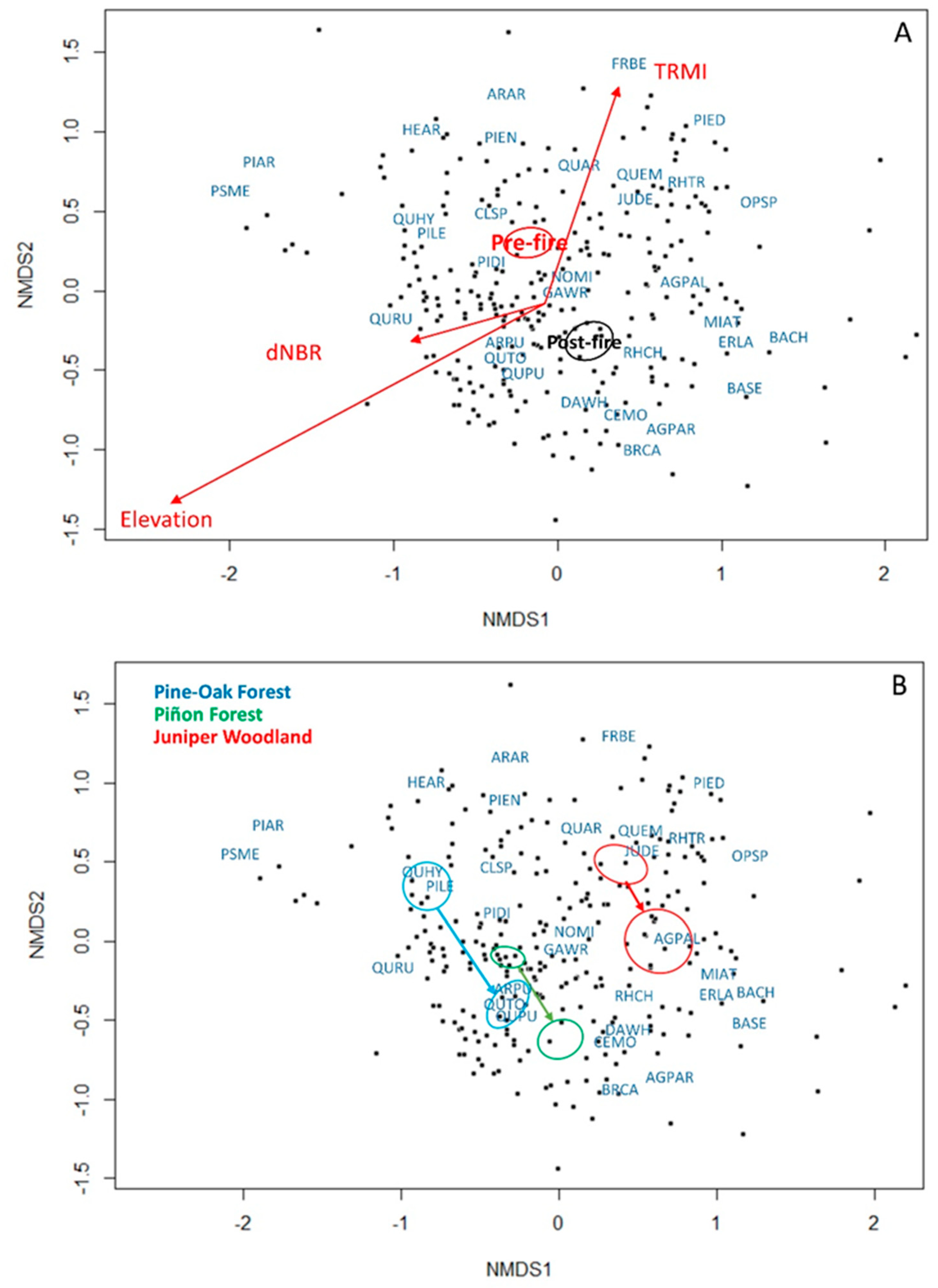
3.1. Vegetation Change, Fire Severity, and Topography
3.2. Vegetation Changes by Forest Cover Type
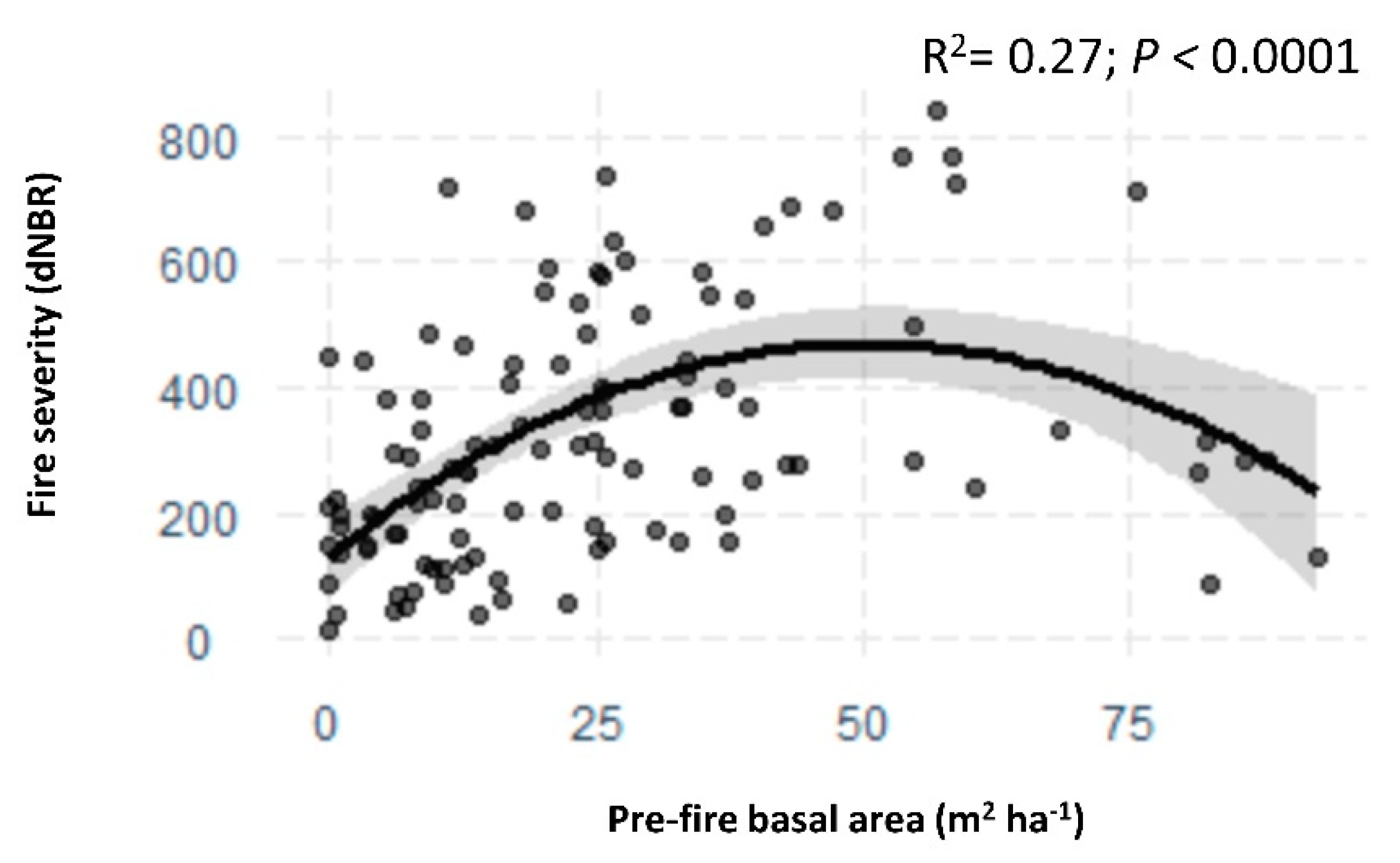
3.3. Fire Effects on Forest Stand Structure
4. Discussion
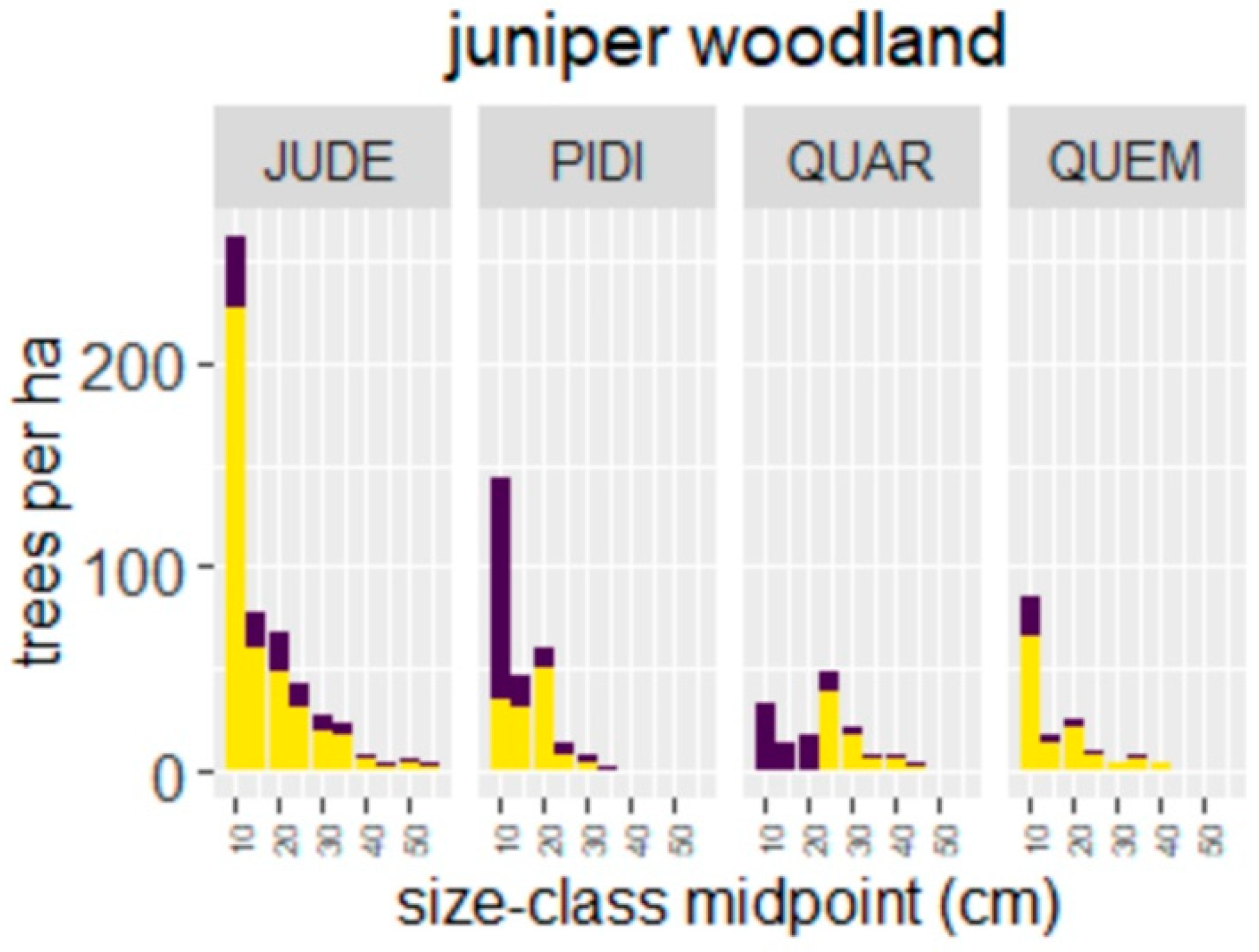
4.1. Wildfire Effects on Juniper Woodlands
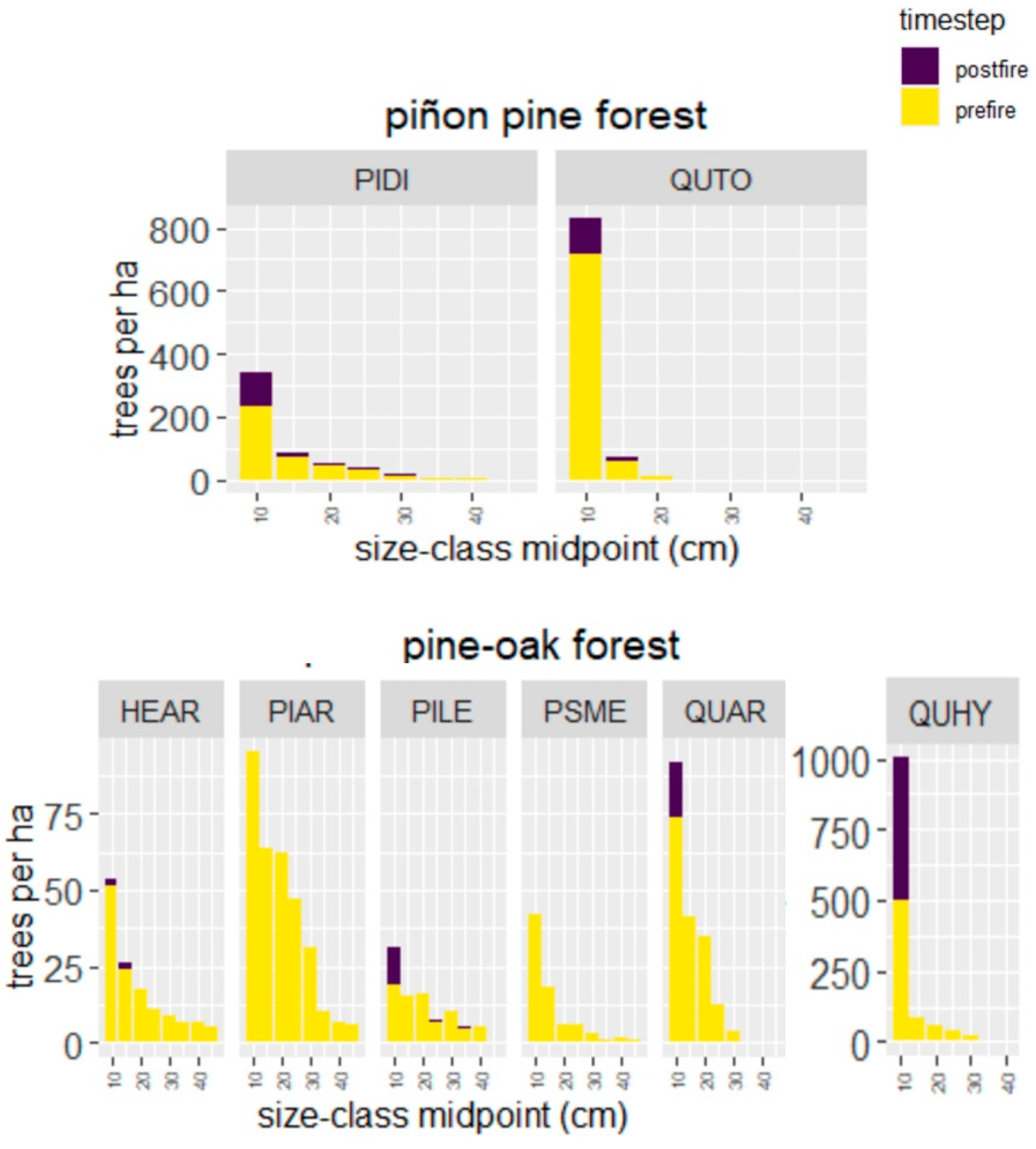
4.2. Wildfire Effects on Piñon Pine Forest
4.3. Wildfire Effects on Pine-Oak Forest
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | All Plots | Juniper Woodland | Pine-Oak Forest | Piñon Forest | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| t | df | p-Value | t | df | p-Value | t | df | p-Value | t | df | p-Value | |
| Agave palmeri | −2.45 | 30 | 0.014 | −1.79 | 16 | 0.075 | −2.2 | 13 | 0.03 | |||
| Arctostaphylos pungens | −8.37 | 90 | <0.001 | −3.39 | 29 | <0.001 | −4.06 | 25 | <0.001 | −10.1 | 36 | <0.001 |
| Dasylirion wheeleri | −3.22 | 27 | <0.001 | −1.99 | 12 | 0.048 | −2.73 | 14 | 0.007 | |||
| Garrya wrightii | −4.42 | 45 | <0.001 | −1.83 | 15 | 0.069 | −2.71 | 10 | 0.008 | −3.32 | 20 | 0.001 |
| Hesperocyparis arizonica | −3.68 | 29 | <0.001 | −1.91 | 6 | 0.058 | ||||||
| Juniperus deppeana | −1.92 | 78 | 0.054 | −1.96 | 46 | 0.051 | 0.22 | 11 | 0.823 | −1.13 | 21 | 0.264 |
| Mimosia aculeiticarpa | −8.45 | 18 | <0.001 | 0.93 | 16 | 0.352 | ||||||
| Nolina microcarpa | −3.02 | 8 | <0.001 | −8.45 | 34 | <0.001 | −2.59 | 18 | 0.011 | −1.14 | 32 | 0.19 |
| Pinus arizonica | −3.11 | 13 | 0.002 | −3.34 | 12 | 0.001 | ||||||
| Pinus discolor | −3.4 | 90 | <0.001 | −6.68 | 14 | <0.001 | −3.2 | 24 | 0.001 | −6.04 | 37 | <0.001 |
| Pinus engelmanii | −1.94 | 12 | 0.052 | −3.4 | 7 | 0.052 | −0.81 | 5 | 0.42 | |||
| Pinus edulis | −3.4 | 18 | <0.001 | −3.52 | 14 | <0.001 | ||||||
| Pinus leiophylla | −3.14 | 34 | 0.001 | −3.14 | 9 | 0.001 | −3.06 | 18 | 0.003 | |||
| Pseudotsuga menziesii | −1.5 | 13 | 0.133 | −1.38 | 11 | 0.168 | ||||||
| Quercus arizonica | −2.92 | 51 | 0.003 | −2.92 | 39 | 0.003 | −1.83 | 10 | 0.07 | |||
| Quercus emoryi | −1.92 | 36 | 0.055 | −1.92 | 33 | 0.055 | ||||||
| Quercus hypoleucoides | −6.11 | 70 | <0.001 | −6.11 | 10 | <0.001 | −7.62 | 40 | <0.001 | −3.35 | 20 | <0.001 |
| Quercus rugosa | −1.04 | 22 | 0.296 | −0.83 | 14 | 0.409 | ||||||
| Quercus toumeyi | −5.44 | 61 | <0.001 | −5.44 | 7 | <0.001 | −11.8 | 39 | <0.001 | |||
Appendix B
| PLANT SPECIES | Acronym | Adaptations & Reponses to Fire |
|---|---|---|
| TREES | ||
| Arziona Madrone (Arbutus arizonica) | ARAR | Variable trunk damage Can survive extensive bark and cambium damage Sprouting from the root crown and tree bole |
| Arizona Cypress (Hesperocyparis arizonica) | HEAR | Fire-sensitive trunk Serotinous cones |
| Alligator-bark Juniper (Juniperus deppeana) | JUDE | Large individuals can survive low-severity fire Sprouting from root crown, trunk, and branches |
| Arizona Pine (Pinus arizonica) | PIAR | Can survive low- and moderate-severity fire |
| Border Piñon (Pinus discolor) | PIDI | Thin bark, fire sensitive Seeds germinate from seeds cached by animals Nurse plants facilitate establishment in the open |
| Rocky Mountain Piñon (Pinus edulis) | PIED | Thin barked fire-sensitive trunk Seeds germinate from seeds cached by animals |
| Apache Pine (Pinus englemannii) | PIEN | Thicker bark and deeper roots, mature trees are fire resistant |
| Chihuahua Pine (Pinus leiophylla) | PILE | Thick bark, can survive low- and moderate-severity fire Modest sprouting, mainly in small stems Semi-serotinous cones |
| Douglas-fir (Pseudotsuga menziesii) | PSME | Can survive low-severity fire; moderate severity in large trees Susceptible to fire crowning due to low branches |
| Arizona White Oak (Quercus arizonica) | QUAR | Large individuals can survive low-severity fire Sprouting after top-kill |
| Emory Oak (Quercus emoryi) | QUEM | Fire-sensitive trunk Sprouting after top-kill |
| Gambel Oak (Quercus gambelii) | QUGA | Sprouting after top-kill from root crown and roots Large individuals can survive low-severity fire |
| Silverleaf Oak (Quercus hypoleucoides) | QUHY | Fire-sensitive trunk, but Large individuals can survive low-severity fire Sprouting after top-kill |
| Southwestern Black Cherry (Prunus serotina var. virens) | PRSE | Fire-sensitive trunk Dormant seed banks stimulated by fire-created openings |
| White fir (Abies concolor) | ABCO | Fire-sensitive trunk |
| SHRUBS | Shrubs exhibit low shoot survival of fire | |
| Netleaf Oak (Quercus rugosa) | QURU | Sprouting after top-kill from root crown |
| Toumey Oak (Quercus toumeyi) | QUTO | Sprouting after top-kill from root crown |
| Pungent Oak (Quercus pungens) | QUPU | Sprouting after top-kill from root crown |
| Pointleaf Manzanita (Arctostaphylos pungens) | ARPU | Obligate seeder Dormant seed banks stimulated by fire Rooting of low branches observed |
| Mimosa catclaw (Mimosa aculeiticarpa var. biuncifera) | MIBI | Sprout from the root crown following top-kill by fire |
| Wright’s Silktassel (Garrya wrightii) | GAWR | Sprouting from root crown |
| Catclaw (Acacia greggii) | ACGR | Sprouting from root crown |
| Birchleaf Buckthorn (Frangula betulifolia) | FRBE | Sprouting from root crown |
| Sotol (Dasylirion wheeleri) | DAWH | moderately sensitive to fire; survivor species capable of sprouting from an aboveground caudex |
| Skunkbush and (Rhus trilobata) | RHTR | Sprouting Possible dormant seed bank |
| Littleleaf Sumac (Rhus microphylla) | RHMI | Sprouting Possible dormant seed bank |
| Mesquite (Prosopis glandulosa) | PRGL | Sprouting after top-kill from underground stem |
| Fendler’s Ceanothus (Ceanothus fendleri) | CEFE | Sprouting after top-kill from root crown Extensive seedling establishment after fire |
| Mountain-mahogany (Cercocarpus montanus) | CEMO | Sprouting after top-kill from root crown |
| Mountain yucca (Yucca madrensis) | YUMA | Sprouting after top-kill from roots |
Appendix C
| Vegetation Type | Species | Size-Class (cm) | t | df | p-Value |
|---|---|---|---|---|---|
| Piñon Forest | Pinus discolor | 10–15 | −3.93 | 76 | <0.001 |
| 15–20 | −5.33 | <0.001 | |||
| 20–25 | −5.22 | <0.001 | |||
| 25–30 | −4.01 | <0.001 | |||
| 30–35 | −3.22 | 0.002 | |||
| 35–40 | −2.29 | 0.024 | |||
| 40–45 | −1.71 | 0.089 | |||
| Quercus toumeyi | 40–45 | −11.9 | <0.001 | ||
| 15–20 | −2.64 | 0.010 | |||
| 20–25 | −0.97 | 0.330 | |||
| 25–30 | −0.98 | 0.329 | |||
| Juniper Forest | Pinus discolor | 10–15 | −3.44 | 116 | <0.001 |
| 15–20 | −3.07 | 0.002 | |||
| 20–25 | −2.68 | 0.008 | |||
| 25–30 | −2.19 | 0.029 | |||
| 30–35 | −2.00 | 0.047 | |||
| Juniperus deppeana | 10–15 | 2.13 | 0.035 | ||
| 15–20 | −2.28 | 0.024 | |||
| 20–25 | −0.46 | 0.640 | |||
| 25–30 | −1.25 | 0.211 | |||
| 30–35 | −0.92 | 0.210 | |||
| 35–40 | −0.87 | 0.385 | |||
| 40–45 | −1.76 | 0.080 | |||
| 45–50 | 0.15 | 0.880 | |||
| 50–55 | 0.06 | 0.952 | |||
| Quercus arizonica | 10–15 | 0.72 | 0.472 | ||
| 15–20 | −5.16 | <0.001 | |||
| 20–25 | −5.8 | <0.001 | |||
| 25–30 | −4.87 | <0.001 | |||
| 30–35 | −2.99 | 0.003 | |||
| 35–40 | −2.59 | 0.010 | |||
| 40–45 | −2.69 | 0.008 | |||
| Quercus emoryi | 10–15 | −1.29 | 0.199 | ||
| 15–20 | −2.47 | 0.014 | |||
| 20–25 | −3.1 | 0.002 | |||
| 25–30 | −3.04 | 0.003 | |||
| 30–35 | −2.23 | 0.027 | |||
| 35–40 | −2.65 | 0.009 | |||
| 40–45 | −1.5 | 0.134 | |||
| Pine-Oak Forest | Hesperocyparis arizonica | 10–15 | −1.79 | 78 | 0.077 |
| 15–20 | −2.4 | 0.018 | |||
| 20–25 | −2.36 | 0.020 | |||
| 25–30 | −1.9 | 0.059 | |||
| 30–35 | −2.22 | 0.029 | |||
| 35–40 | −1.84 | 0.068 | |||
| 40–45 | −1.5 | 0.135 | |||
| Pinus arizonica | 10–15 | −2.12 | 0.036 | ||
| 15–20 | −3.36 | 0.001 | |||
| 20–25 | −3.31 | 0.001 | |||
| 25–30 | −3.06 | 0.002 | |||
| 30–35 | −2.83 | 0.005 | |||
| 35–40 | −2.18 | 0.031 | |||
| 40–45 | −1.74 | 0.084 | |||
| 50–55 | −1.36 | 0.175 | |||
| Pinus engelmannii | 10–15 | 0.847 | 0.399 | ||
| 15–20 | −1.43 | 0.156 | |||
| 20–25 | −1.432 | 0.155 | |||
| 25–30 | −1.98 | 0.050 | |||
| 30–35 | −1.73 | 0.086 | |||
| Pinus leiophylla | 10–15 | −0.81 | 0.415 | ||
| 15–20 | −2.17 | 0.033 | |||
| 20–25 | −2.69 | 0.008 | |||
| 25–30 | −2 | 0.048 | |||
| 30–35 | −1.69 | 0.093 | |||
| 35–40 | −2.05 | 0.043 | |||
| 45–50 | −1.68 | 0.096 | |||
| 50–55 | −1 | 0.320 | |||
| Quercus arizonica | 10–15 | −1.45 | 0.148 | ||
| 15–20 | −1.91 | 0.059 | |||
| 20–25 | −2.78 | 0.006 | |||
| 25–30 | −2.13 | 0.036 | |||
| 30–35 | −1.31 | 0.193 | |||
| Quercus hypoleucoides | 10–15 | −8.7 | <0.001 | ||
| 15–20 | −3.01 | 0.003 | |||
| 20–25 | −2.94 | 0.004 | |||
| 25–30 | −3.23 | 0.001 | |||
| 30–35 | −2.11 | 0.037 |
Appendix D
| Model | Estimate | Standard Error | z-Value | P |
|---|---|---|---|---|
| (Intercept) | 2.51 | 0.47 | 5.40 | <0.001 |
| dNBR | −0.01 | 0.00 | −5.79 | <0.001 |
References
- Bond, W.J.; Keeley, J.E. Fire as a global ‘herbivore’: The ecology and evolution of flammable ecosystems. Trends Ecol. Evol. 2005, 20, 387–394. [Google Scholar] [CrossRef]
- He, T.; Lamont, B.B.; Pausas, J.G. Fire as a key driver of Earth’s biodiversity. Biol. Rev. 2019, 94, 1983–2010. [Google Scholar] [CrossRef] [PubMed]
- Pausas, J.G.; Ribeiro, E. The global fire–productivity relationship. Glob. Ecol. Biogeogr. 2013, 22, 728–736. [Google Scholar] [CrossRef]
- McLaughlin, B.C.; Blakey, R.; Weitz, A.P.; Feng, X.; Brown, B.J.; Ackerly, D.D.; Dawson, T.E.; Thompson, S.E. Weather underground: Subsurface hydrologic processes mediate tree vulnerability to extreme climatic drought. Glob. Chang. Biol. 2020, 26, 3091–3107. [Google Scholar] [CrossRef] [PubMed]
- Kolden, C.A.; Bleeker, T.; Smith, A.; Poulos, H.M.; Camp, A.E. Fire Effects on Historical Wildfire Refugia in Contemporary Wildfires. Forests 2017, 8, 400. [Google Scholar] [CrossRef] [Green Version]
- Enright, N.; Keith, D.A.; Clarke, M.F.; Miller, B.P. Fire regimes in Australian sclerophyllous shrubby ecosystems: Heathlands, heathy woodlands and mallee woodlands. In Flammable Australia: Fire Regimes and Biodiversity in a Changing World; CSIRO Publishing: Melbourne, Australia, 2012; pp. 215–235. [Google Scholar]
- Gonzalez, M.E.; Veblen, T.T.; Sibold, J.S. Influence of fire severity on stand development of Araucaria araucana–Nothofagus pumilio stands in the Andean cordillera of south-central Chile. Austral Ecol. 2010, 35, 597–615. [Google Scholar] [CrossRef]
- Fulé, P.Z.; Crouse, J.E.; Heinlein, T.A.; Moore, M.M.; Covington, W.W.; Verkamp, G. Mixed-severity fire regime in a high-elevation forest of Grand Canyon, Arizona, USA. Landsc. Ecol. 2003, 18, 465–486. [Google Scholar] [CrossRef]
- Cohn, J.S.; Di Stefano, J.; Christie, F.; Cheers, G.; York, A. How do heterogeneity in vegetation types and post-fire age-classes contribute to plant diversity at the landscape scale? For. Ecol. Manag. 2015, 346, 22–30. [Google Scholar] [CrossRef]
- Parr, C.L.; Andersen, A.N. Patch mosaic burning for biodiversity conservation: A critique of the pyrodiversity paradigm. Conserv. Biol. 2006, 20, 1610–1619. [Google Scholar] [CrossRef]
- Clarke, P.J.; Lawes, M.J.; Murphy, B.P.; Russell-Smith, J.; Nano, C.E.M.; Bradstock, R.; Enright, N.J.; Fontaine, J.B.; Gosper, C.R.; Radford, I.; et al. A synthesis of postfire recovery traits of woody plants in Australian ecosystems. Sci. Total Environ. 2015, 534, 31–42. [Google Scholar] [CrossRef]
- He, T.; Lamont, B.B. Baptism by fire: The pivotal role of ancient conflagrations in evolution of the Earth’s flora. Natl. Sci. Rev. 2018, 5, 237–254. [Google Scholar] [CrossRef] [Green Version]
- Lamont, B.B.; Enright, N.J.; He, T. Fitness and evolution of resprouters in relation to fire. Plant Ecol. 2011, 212, 1945–1957. [Google Scholar] [CrossRef]
- Krawchuk, M.A.; Haire, S.L.; Coop, J.; Parisien, M.A.; Whitman, E.; Chong, G.; Miller, C. Topographic and fire weather controls of fire refugia in forested ecosystems of northwestern North America. Ecosphere 2016, 7, e01632. [Google Scholar] [CrossRef]
- Archibald, S.; Lehmann, C.E.; Belcher, C.; Bond, W.J.; Bradstock, R.A.; Daniau, A.; Dexter, K.; Forrestel, E.; Greve, M.; He, T. Biological and geophysical feedbacks with fire in the Earth system. Environ. Res. Lett. 2018, 13, 033003. [Google Scholar] [CrossRef] [Green Version]
- Poulos, H.M.; Taylor, A.H.; Beaty, R.M. Environmental controls on dominance and diversity of woody plant species in a Madrean, Sky Island ecosystem, Arizona, USA. Plant Ecol. 2007, 193, 15–30. [Google Scholar] [CrossRef]
- Poulos, H.M.; Camp, A.E. Topographic influences on vegetation mosaics and tree diversity in the Chihuahuan Desert Borderlands. Ecology 2010, 91, 1140–1151. [Google Scholar] [CrossRef]
- Coblentz, D.D.; Riitters, K.H. Topographic controls on the regional-scale biodiversity of the south-western USA. J. Biogeogr. 2004, 31, 1125–1138. [Google Scholar] [CrossRef]
- Van Devender, T.R.; Avila-Villegas, S.; Emerson, M.; Turner, D.; Flesch, A.D.; Deyo, N.S. Biodiversity in the Madrean Archipelago of Sonora, Mexico. In Proceedings of the Merging Science and Management in a Rapidly Changing World: Biodiversity and Management of the Madrean Archipelago III and 7th Conference on Research and Resource Management in the Southwestern Deserts, Tucson, AZ, USA, 1–5 May 2012. [Google Scholar]
- Felger, R.S.; Wilson, M.F. Northern Sierra Madre Occidental and its Apachian outliers: A neglected center of biodiversity. In Biodiversity and Management of the Madrean Archipelago: The Sky Islands of Southwestern United States and Northwestern Mexico; 1995; pp. 36–51. [Google Scholar]
- O’Connor, C.D.; Falk, D.A.; Lynch, A.M.; Swetnam, T.W. Fire severity, size, and climate associations diverge from historical precedent along an ecological gradient in the Pinaleño Mountains, Arizfona, USA. For. Ecol. Manag. 2014, 329, 264–278. [Google Scholar] [CrossRef] [Green Version]
- Yocom, L.L.; Fulé, P.Z.; Falk, D.A.; García-Domínguez, C.; Cornejo-Oviedo, E.; Brown, P.M.; Villanueva-Díaz, J.; Cerano, J.; Montaño, C.C. Fine-scale factors influence fire regimes in mixed-conifer forests on three high mountains in Mexico. Int. J. Wildland Fire 2014, 23, 959–968. [Google Scholar] [CrossRef]
- Margolis, E.Q.; Swetnam, T.W.; Allen, C.D. Historical stand-replacing fire in upper montane forests of the Madrean Sky Islands and Mogollon Plateau, southwestern USA. Fire Ecol. 2011, 7, 88–107. [Google Scholar] [CrossRef]
- Margolis, E.Q. Fire regime shift linked to increased forest density in a piñon–juniper savanna landscape. Int. J. Wildland Fire 2014, 23, 234–245. [Google Scholar] [CrossRef]
- Morino, K.A.; Baisan, C.H.; Swetnam, T.W. Historical Fire Regimes in the Chiricahua Mountains, Arizona: An Examination of Fire along an Elevation Gradient and in Mixed-Confier Forest; Laboratory of Tree-Ring Research, The University of Arizona: Tucson, AZ, USA, 2000. [Google Scholar]
- Taylor, A.H.; Poulos, H.M.; Kluber, J.; Issacs, R.; Pawlikowski, N.; Barton, A.M. Controls on spatial patterns of wildfire severity and early post-fire vegetation development in an Arizona Sky Island, USA. Landsc. Ecol. 2021, 36, 2637–2656. [Google Scholar] [CrossRef]
- Schoennagel, T.; Balch, J.K.; Brenkert-Smith, H.; Dennison, P.E.; Harvey, B.J.; Krawchuk, M.A.; Mietkiewicz, N.; Morgan, P.; Moritz, M.A.; Rasker, R.; et al. Adapt to more wildfire in western North American forests as climate changes. Proc. Natl. Acad. Sci. USA 2017, 114, 4582–4590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villarreal, M.L.; Iniguez, J.M.; Flesch, A.D.; Sanderlin, J.S.; Cortés Montaño, C.; Conrad, C.R.; Haire, S.L. Contemporary Fire Regimes Provide a Critical Perspective on Restoration Needs in the Mexico-United States Borderlands. Air Soil Water Res. 2020, 13, 1178622120969191. [Google Scholar] [CrossRef]
- Villarreal, M.L.; Haire, S.L.; Iniguez, J.M.; Montaño, C.C.; Poitras, T.B. Distant neighbors: Recent wildfire patterns of the Madrean Sky Islands of southwestern United States and northwestern Mexico. Fire Ecol. 2019, 15, 2. [Google Scholar] [CrossRef]
- Haire, S.L.; McGarigal, K. Effects of landscape patterns of fire severity on regenerating ponderosa pine forests (Pinus ponderosa) in New Mexico and Arizona, USA. Landsc. Ecol. 2010, 25, 1055–1069. [Google Scholar] [CrossRef]
- Haire, S.L.; McGarigal, K. Changes in fire severity across gradients of climate, fire size, and topography: A landscape ecological perspective. Fire Ecol. 2009, 5, 86–103. [Google Scholar] [CrossRef]
- Haffey, C.M. Patterns and Predictors of Crown Fire Induced Type Conversion in Dry Conifer Forests; Northern Arizona University: Flagstaff, AZ, USA, 2014. [Google Scholar]
- Coop, J.D.; Parks, S.A.; McClernan, S.R.; Holsinger, L.M. Influences of prior wildfires on vegetation response to subsequent fire in a reburned southwestern landscape. Ecol. Appl. 2016, 26, 346–354. [Google Scholar] [CrossRef]
- Barton, A.M.; Poulos, H.M. Pine vs. oaks revisited: Conversion of Madrean pine-oak forest to oak shrubland after high-severity wildfire in the Sky Islands of Arizona. For. Ecol. Manag. 2018, 414, 28–40. [Google Scholar] [CrossRef]
- Barton, A.M. Pines versus oaks: Effects of fire on the composition of Madrean forests in Arizona. For. Ecol. Manag. 1999, 120, 143–156. [Google Scholar] [CrossRef]
- Coop, J.D.; Parks, S.A.; Stevens-Rumann, C.S.; Crausbay, S.D.; Higuera, P.E.; Hurteau, M.D.; Tepley, A.; Whitman, E.; Assal, T.; Collins, B.M. Wildfire-driven forest conversion in western North American landscapes. BioScience 2020, 70, 659–673. [Google Scholar] [CrossRef]
- Guiterman, C.H.; Margolis, E.Q.; Allen, C.D.; Falk, D.A.; Swetnam, T.W. Long-term persistence and fire resilience of oak shrubfields in dry conifer forests of northern New Mexico. Ecosystems 2018, 21, 943–959. [Google Scholar] [CrossRef]
- Stevens-Rumann, C.S.; Morgan, P. Tree regeneration following wildfires in the western US: A review. Fire Ecol. 2019, 15, 15. [Google Scholar] [CrossRef] [Green Version]
- Johnston, B.C. Woodland classification: The pinyon-juniper formation. Gen. Tech. Rep. INT 1989, 160. [Google Scholar]
- Wood, M.K.; Scanlon, R.; Cram, D.S. Occurrence of sprouts and seedlings of Pinyon Pines, Alligator Junipers, and Gray Oaks following harvest of fuelwood and prescribed burning. Southwest. Nat. 2012, 57, 51–57. [Google Scholar] [CrossRef]
- Márquez Linares, M.A. Distribución, Germinación y Estructura de Poblaciones de Arctostaphylos pungens HBK, y su Relación con el Fuego en Durango, México; Universidad Autónoma de Nuevo León: San Nicolás de los Garza, Mexico, 2004. [Google Scholar]
- Arechederra-Romero, L. Southwest Fire Science Consortium Field Trip to the Chiricahua National Monument: Discussion of the Impacts of the 2011 Horseshoe 2 Fire. Environ. Geol. 2012. [Google Scholar]
- Drewes, H.; Williams, F. Mineral Resources of the Chiricahua Wilderness Area; US Geological Survey Bulletin: Cochise County, AZ, USA, 1973.
- Adams, D.K.; Comrie, A.C. The North American Monsoon. Bull. Am. Meteorol. Soc. 1997, 78, 2197–2213. [Google Scholar] [CrossRef] [Green Version]
- Minor, J.; Falk, D.A.; Barron-Gafford, G.A. Fire Severity and Regeneration Strategy Influence Shrub Patch Size and Structure Following Disturbance. Forests 2017, 8, 221. [Google Scholar] [CrossRef] [Green Version]
- Barton, A.M. Factors controlling plant distributions: Drought, competition, and fire in montane pines in Arizona. Ecol. Monogr. 1993, 63, 367–397. [Google Scholar] [CrossRef]
- Barton, A.M.; Teeri, J.A. The ecology of elevational positions in plants: Drought resistance in five montane pine species in southeastern Arizona. Am. J. Bot. 1993, 80, 15–25. [Google Scholar] [CrossRef]
- USDA. USDA Plants Database; USDA, 2021. Available online: https://plants.usda.gov/ (accessed on 20 July 2020).
- Swetnam, T.W.; Baisan, C.H. Fire Histories of Montane Forests in the Madrean Borderlands; United States Department of Agriculture Forest Service: Washington, DC, USA, 1996.
- Barton, A.M.; Swetnam, T.W.; Baisan, C.H. Arizona pine (Pinus arizonica) stand dynamics: Local and regional factors in a fire-prone madrean gallery forest of Southeast Arizona, USA. Landsc. Ecol. 2001, 16, 351–369. [Google Scholar] [CrossRef]
- Swetnam, T.W.; Baisan, C.H.; Brown, P.M.; Caprio, A.C. Fire History of Rhyolite Canyon, Chiricahua National Monument; Cooperative National Park Resources Studies Unit, School of Renewable Natural Resources, University of Arizona: Tucson, AZ, USA, 1989. [Google Scholar]
- Eidenshink, J.; Schwind, B.; Brewer, K.; Zhu, Z.-L.; Quayle, B.; Howard, S. A project for monitoring trends in burn severity. Fire Ecol. 2007, 3, 3–21. [Google Scholar] [CrossRef]
- Miller, J.D.; Thode, A.E. Quantifying burn severity in a heterogeneous landscape with a relative version of the delta Normalized Burn Ratio (dNBR). Remote Sens. Environ. 2007, 109, 66–80. [Google Scholar] [CrossRef]
- QGIS. QGIS Geographic Information System; QGIS, 2020. Available online: qgis.osgeo.org (accessed on 20 July 2020).
- Parker, A.J. The topographic relative moisture index: An approach to soil-moisture assessment in mountain terrain. Phys. Geogr. 1982, 3, 160–168. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’hara, R.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Package ‘vegan’. Community Ecol. Package Version 2013, 2, 1–295. [Google Scholar]
- Husson, F.; Josse, J.; Le, S.; Mazet, J.; Husson, M.F. Package ‘FactoMineR’. R Package 2016, 96, 698. [Google Scholar]
- De Caceres, M.; Jansen, F.; De Caceres, M.M. Package ‘indicspecies’. Indicators 2016, 8, 1. [Google Scholar]
- Jackman, S.; Tahk, A.; Zeileis, A.; Maimone, C.; Fearon, J.; Meers, Z.; Jackman, M.S.; Imports, M. Package ‘pscl’; Political Science Computational Laboratory: Stanford, CA, USA, 2015; Volume 18. [Google Scholar]
- Huffman, D.W.; Floyd, M.L.; Hanna, D.P.; Crouse, J.E.; Fulé, P.Z.; Meador, A.J.S.; Springer, J.D. Fire regimes and structural changes in oak-pine forests of the Mogollon Highlands ecoregion: Implications for ecological restoration. For. Ecol. Manag. 2020, 465, 118087. [Google Scholar] [CrossRef]
- Savage, M.; Mast, J.N. How resilient are southwestern ponderosa pine forests after crown fires? Can. J. For. Res. 2005, 35, 967–977. [Google Scholar] [CrossRef] [Green Version]
- Strom, B.A.; Fulé, P.Z. Pre-wildfire fuel treatments affect long-term ponderosa pine forest dynamics. Int. J. Wildland Fire 2007, 16, 128–138. [Google Scholar] [CrossRef]
- Savage, M.; Mast, J.N.; Feddema, J.J. Double whammy: High-severity fire and drought in ponderosa pine forests of the Southwest. Can. J. For. Res. 2013, 43, 570–583. [Google Scholar] [CrossRef]
- Shive, K.L.; Sieg, C.H.; Fulé, P.Z. Pre-wildfire management treatments interact with fire severity to have lasting effects on post-wildfire vegetation response. For. Ecol. Manag. 2013, 297, 75–83. [Google Scholar] [CrossRef]
- Roccaforte, J.P.; Fuleé, P.Z.; Chancellor, W.W.; Laughlin, D.C. Woody debris and tree regeneration dynamics following severe wildfires in Arizona ponderosa pine forests. Can. J. For. Res. 2012, 42, 593–604. [Google Scholar] [CrossRef]
- Stevens-Rumann, C.S.; Kemp, K.B.; Higuera, P.E.; Harvey, B.J.; Rother, M.T.; Donato, D.C.; Morgan, P.; Veblen, T.T. Evidence for declining forest resilience to wildfires under climate change. Ecol. Lett. 2018, 21, 243–252. [Google Scholar] [CrossRef] [PubMed]
- Collins, B.M.; Roller, G.B. Early forest dynamics in stand-replacing fire patches in the northern Sierra Nevada, California, USA. Landsc. Ecol. 2013, 28, 1801–1813. [Google Scholar] [CrossRef]
- Young, D.; Latimer, A. Post-fire forest regeneration in a changing climate. Joint Fire Sci. Program Rep. 2017. [Google Scholar]
- Barton, A.M. Intense wildfire in southeastern Arizona: Transformation of a Madrean oak–pine forest to oak woodland. For. Ecol. Manag. 2002, 165, 205–212. [Google Scholar] [CrossRef]
- Iniguez, J.M.; Swetnam, T.W.; Baisan, C.H. Spatially and temporally variable fire regime on RIncon Peak, Arizona, USA. Fire Ecol. Spec. Issue 2009, 5, 3–21. [Google Scholar] [CrossRef]
- Fulé, P.Z.; Covington, W. Spatial patterns of Mexican pine-oak forests under different recent fire regimes. Plant Ecol. 1998, 134, 197–209. [Google Scholar] [CrossRef]
- Fulé, P. Effects of an intense wildfire in a Mexican oak-pine forest. For. Sci. 2000, 46, 52–61. [Google Scholar]
- Romme, W.H.; Allen, C.D.; Bailey, J.D.; Baker, W.L.; Bestelmeyer, B.T.; Brown, P.M.; Eisenhart, K.S.; Floyd, M.L.; Huffman, D.W.; Jacobs, B.F. Historical and modern disturbance regimes, stand structures, and landscape dynamics in piñon–juniper vegetation of the western United States. Rangel. Ecol. Manag. 2009, 62, 203–222. [Google Scholar] [CrossRef] [Green Version]
- Floyd, M.L.; Romme, W.H.; Hanna, D.P.; Hanna, D.D. Historical and Modern Fire Regimes in Piñon-Juniper Woodlands, Dinosaur National Monument, United States. Rangel. Ecol. Manag. 2017, 70, 348–355. [Google Scholar] [CrossRef]
- Floyd, M.L.; Clifford, M.; Cobb, N.S.; Hanna, D.; Delph, R.; Ford, P.; Turner, D. Relationship of stand characteristics to drought-induced mortality in three Southwestern piñon-juniper woodlands. Ecol. Appl. 2009, 19, 1223–1230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Floyd, M.L.; Hanna, D.D.; Romme, W.H. Historical and recent fire regimes in piñon–juniper woodlands on Mesa Verde, Colorado, USA. For. Ecol. Manag. 2004, 198, 269–289. [Google Scholar] [CrossRef]
- Poulos, H.; Gatewood, R.; Camp, A. Fire regimes of the piñon-juniper woodlands of Big Bend National Park and the Davis Mountains, west Texas, USA. Can. J. For. Res. 2009, 39, 1236–1246. [Google Scholar] [CrossRef]
- Poulos, H.M.; Reemts, C.M.; Wogan, K.A.; Karges, J.P.; Gatewood, R.G. Multiple wildfires with minimal consequences: Low-severity wildfire effects on West Texas piñon-juniper woodlands. For. Ecol. Manag. 2020, 473, 118293. [Google Scholar] [CrossRef]
- Baisan, C.H.; Morino, K.A. Fire History in Chiricahua National Monument; National Park Service: Washington, DC, USA, 2000. [Google Scholar]
- Falk, D.A.; Watts, A.C.; Thode, A.E. Scaling ecological resilience. Front. Ecol. Evol. 2019, 7, 275. [Google Scholar] [CrossRef] [Green Version]
- Lanner, R.M. Made for Each Other: A symbiosis of Birds and Pines; Oxford University Press on Demand: Oxford, UK, 1996. [Google Scholar]
- Vander Wall, S.B. Dispersal of singleleaf pinon pine (Pinus monophylla) by seed-caching rodents. J. Mammal. 1997, 78, 181–191. [Google Scholar] [CrossRef]
- Kane, V.R.; Cansler, C.A.; Povak, N.A.; Kane, J.T.; McGaughey, R.J.; Lutz, J.A.; Churchill, D.J.; North, M.P. Mixed severity fire effects within the Rim fire: Relative importance of local climate, fire weather, topography, and forest structure. For. Ecol. Manag. 2015, 358, 62–79. [Google Scholar] [CrossRef] [Green Version]
- Kane, J.M.; Dugi, F.L.; Kolb, T.E. Establishment and growth of pinon pine regeneration vary by nurse type along a soil substrate age gradient in northern Arizona. J. Arid Environ. 2015, 115, 113–119. [Google Scholar] [CrossRef]
- Keeley, J.E.; Fotheringham, C. Role of fire in regeneration from seed. Seeds Ecol. Regen. Plant Communities 2000, 2, 311–330. [Google Scholar]
- Pausas, J.G.; Bradstock, R.A.; Keith, D.A.; Keeley, J.E. Plant functional traits in relation to fire in crown-fire ecosystems. Ecology 2004, 85, 1085–1100. [Google Scholar] [CrossRef] [Green Version]
- Pausas, J.G.; Keeley, J.E. Evolutionary ecology of resprouting and seeding in fire-prone ecosystems. New Phytol. 2014, 204, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Barton, A.M.; Poulos, H.M. Response of Arizona cypress (Hesperocyparis arizonica) to the Horseshoe Two Megafire in a south-eastern Arizona Sky Island mountain range. Int. J. Wildland Fire 2019, 28, 62–69. [Google Scholar] [CrossRef]
- Kaib, M.; Baisan, C.H.; Grissino-Mayer, H.D.; Swetnam, T.W. Fire History of the Gallery Pine-Oak Forests and Adjacent Grasslands of the Chiricahua Mountains of Arizona; General Technical Report RM; United States Department of Agriculture Forest Service: Tucson, AZ, USA, 1996; pp. 253–264.
- Davis, K.T.; Dobrowski, S.Z.; Higuera, P.E.; Holden, Z.A.; Veblen, T.T.; Rother, M.T.; Parks, S.A.; Sala, A.; Maneta, M.P. Wildfires and climate change push low-elevation forests across a critical climate threshold for tree regeneration. Proc. Natl. Acad. Sci. USA 2019, 116, 6193–6198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Owen, S.M.; Sieg, C.H.; Sánchez Meador, A.J.; Fulé, P.Z.; Iniguez, J.M.; Baggett, L.S.; Fornwalt, P.J.; Battaglia, M.A. Spatial patterns of ponderosa pine regeneration in high-severity burn patches. For. Ecol. Manag. 2017, 405, 134–149. [Google Scholar] [CrossRef]
- Peeler, J.L.; Smithwick, E.A. Seed source pattern and terrain have scale-dependent effects on post-fire tree recovery. Landsc. Ecol. 2020, 35, 1945–1959. [Google Scholar] [CrossRef]
- Harris, L.; Taylor, A.H. Topography, fuels, and fire exclusion drive fire severity of the Rim Fire in an old-growth mixed-conifer forest, Yosemite National Park, USA. Ecosystems 2015, 18, 1192–1208. [Google Scholar] [CrossRef]
- Harris, L.B.; Taylor, A.H. Rain-shadow forest margins resilient to low-severity fire and climate change but not high-severity fire. Ecosphere 2020, 11, e03258. [Google Scholar] [CrossRef]
- Bonnet, V.H.; Schoettle, A.W.; Shepperd, W.D. Postfire environmental conditions influence the spatial pattern of regeneration for Pinus ponderosa. Can. J. For. Res. 2005, 35, 37–47. [Google Scholar] [CrossRef] [Green Version]
- Downing, W.M.; Krawchuk, M.A.; Coop, J.D.; Meigs, G.W.; Haire, S.L.; Walker, R.B.; Whitman, E.; Chong, G.; Miller, C.; Tortorelli, C. How do plant communities differ between fire refugia and fire-generated early-seral vegetation? J. Veg. Sci. 2020, 31, 26–39. [Google Scholar] [CrossRef]
- Meddens, A.J.; Kolden, C.A.; Lutz, J.A.; Smith, A.M.; Cansler, C.A.; Abatzoglou, J.T.; Meigs, G.W.; Downing, W.M.; Krawchuk, M.A. Fire refugia: What are they, and why do they matter for global change? BioScience 2018, 68, 944–954. [Google Scholar] [CrossRef]
- Krawchuk, M.A.; Meigs, G.W.; Cartwright, J.M.; Coop, J.D.; Davis, R.; Holz, A.; Kolden, C.; Meddens, A.J. Disturbance refugia within mosaics of forest fire, drought, and insect outbreaks. Front. Ecol. Environ. 2020, 18, 235–244. [Google Scholar] [CrossRef]
- Lauvaux, C.A.; Skinner, C.N.; Taylor, A.H. High severity fire and mixed conifer forest-chaparral dynamics in the southern Cascade Range, USA. For. Ecol. Manag. 2016, 363, 74–85. [Google Scholar] [CrossRef]
- Cook, B.I.; Ault, T.R.; Smerdon, J.E. Unprecedented 21st century drought risk in the American Southwest and Central Plains. Sci. Adv. 2015, 1, e1400082. [Google Scholar] [CrossRef] [Green Version]
- Ault, T.R.; Mankin, J.S.; Cook, B.I.; Smerdon, J.E. Relative impacts of mitigation, temperature, and precipitation on 21st-century megadrought risk in the American Southwest. Sci. Adv. 2016, 2, e1600873. [Google Scholar] [CrossRef] [Green Version]
- Adams, M.A. Mega-fires, tipping points and ecosystem services: Managing forests and woodlands in an uncertain future. For. Ecol. Manag. 2013, 294, 250–261. [Google Scholar] [CrossRef]
- Abatzoglou, J.T.; Kolden, C.A. Relationships between climate and macroscale area burned in the western United States. Int. J. Wildland Fire 2013, 22, 1003–1020. [Google Scholar] [CrossRef]
- Singleton, M.P.; Thode, A.E.; Meador, A.J.S.; Iniguez, J.M. Increasing trends in high-severity fire in the southwestern USA from 1984 to 2015. For. Ecol. Manag. 2019, 433, 709–719. [Google Scholar] [CrossRef]
- Kemp, K.B.; Higuera, P.E.; Morgan, P.; Abatzoglou, J.T. Climate will increasingly determine post-fire tree regeneration success in low-elevation forests, Northern Rockies, USA. Ecosphere 2019, 10, e02568. [Google Scholar] [CrossRef]
- Liang, S.; Hurteau, M.D.; Westerling, A.L. Response of Sierra Nevada forests to projected climate–wildfire interactions. Glob. Chang. Biol. 2017, 23, 2016–2030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Connor, C.D.; Falk, D.A.; Garfin, G.M. Projected climate-fire interactions drive forest to shrubland transition on an Arizona sky island front. Environ. Sci. 2020, 8. [Google Scholar]
- Yanahan, A.D.; Moore, W. Impacts of 21st-century climate change on montane habitat in the Madrean Sky Island Archipelago. Divers. Distrib. 2019, 25, 1625–1638. [Google Scholar] [CrossRef] [Green Version]
- Gratani, L. Plant Phenotypic Plasticity in Response to Environmental Factors. Adv. Bot. 2014, 2014, 17. [Google Scholar] [CrossRef] [Green Version]
- Hess, V.A.; Fule, P.Z. Is a Mexican pine species better adapted to the warming climate of the southwestern USA? Front. For. Glob. Chang. 2020, 3, 60. [Google Scholar] [CrossRef]
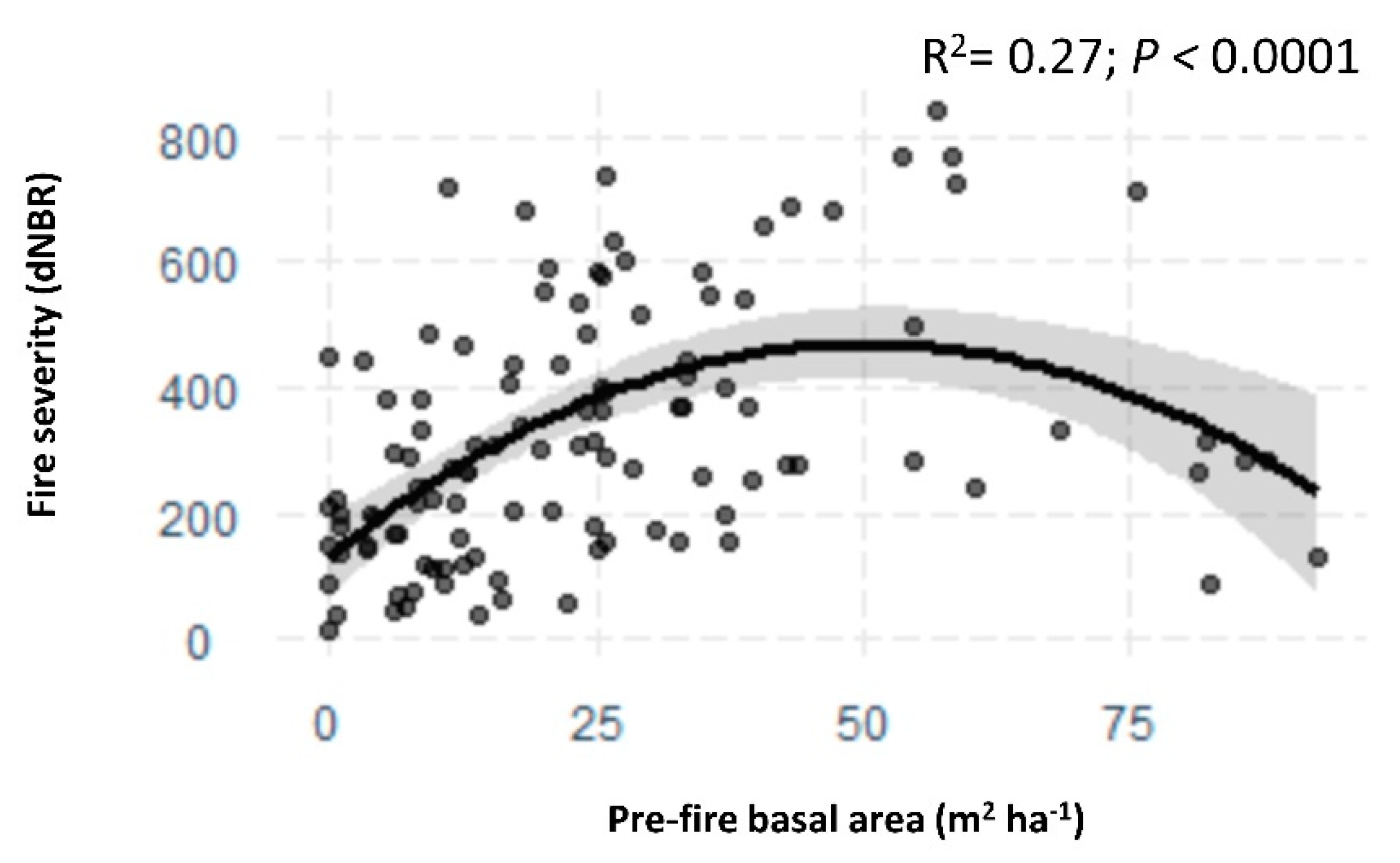
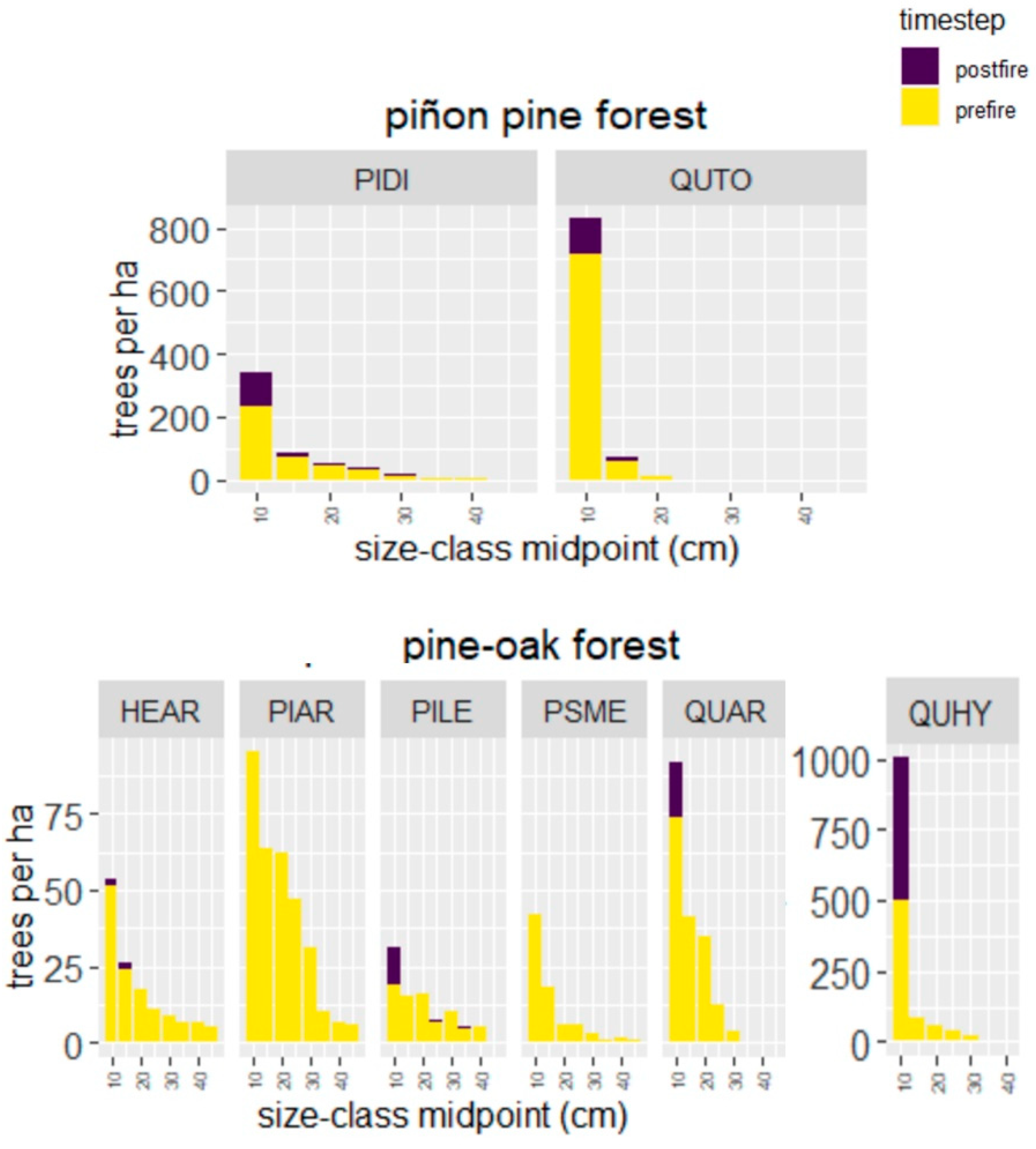
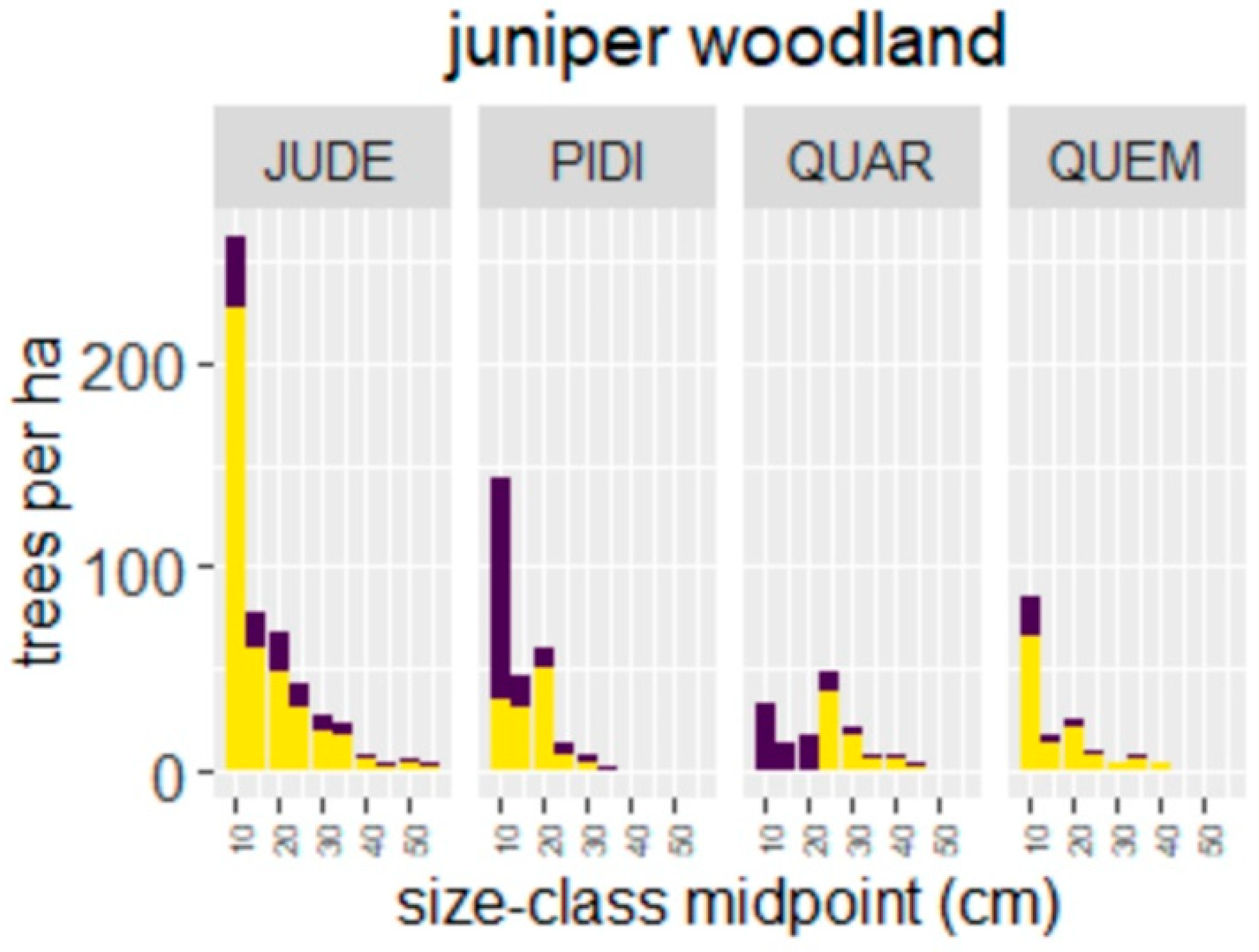
| Species | n | year | Juvenile Plant Density (#/ha) | Basal Area (m2/ha) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Hesperocyparis arizonica | 26 | pre | 44.3 | ± | 27.1 | ** | 2.6 | ± | 2.2 | ** |
| post | 5.8 | ± | 4.8 | 0.2 | ± | 0.4 | ||||
| Juniperus deppeana | 78 | pre | 20.3 | ± | 77.8 | ns | 4.5 | ± | 8.2 | ** |
| post | 36.0 | ± | 103.0 | 0.4 | ± | 1.1 | ||||
| Pinus arizonica | 13 | pre | 20.8 | ± | 11.6 | ns | 3.0 | ± | 1.6 | ns |
| post | 0 | ± | 0 | 0 | ± | 0 | ||||
| Pinus discolor | 90 | pre | 165.0 | ± | 276.0 | ** | 2.4 | ± | 3.9 | ** |
| post | 23.7 | ± | 82.0 | 0.2 | ± | 1.6 | ||||
| Pinus edulis | 16 | pre | 23.2 | ± | 100.0 | ** | 0.8 | ± | 2.9 | ** |
| post | 0.0 | ± | 0.0 | 0.0 | ± | 0.0 | ||||
| Pinus leiophylla | 34 | pre | 12.1 | ± | 43.6 | * | 1.6 | ± | 6.0 | ** |
| post | 4.4 | ± | 23.3 | 0.0 | ± | 0.1 | ||||
| Platanus wrightii | 5 | pre | 4.8 | ± | 7.1 | ns | 0.2 | ± | 2.1 | ns |
| post | 2.9 | ± | 9.5 | 0.0 | ± | 0.0 | ||||
| Pseudotsuga menziesii | 13 | pre | 10.4 | ± | 2.2 | ns | 0.6 | ± | 2.2 | * |
| post | 0.0 | ± | 0.0 | 0.0 | ± | 0.0 | ||||
| Pinus engelmannii | 12 | pre | 3.1 | ± | 23.0 | ns | 2.2 | ± | 8.7 | * |
| post | 4.4 | ± | 34.3 | 0.0 | ± | 0.2 | ||||
| Quercus arizonica | 51 | pre | 29.9 | ± | 119.6 | ns | 4.4 | ± | 8.0 | ** |
| post | 22.0 | ± | 74.6 | 0.0 | ± | 0.3 | ||||
| Quercus emoryi | 36 | pre | 107.9 | ± | 110.4 | ** | 1.4 | ± | 4.4 | ns |
| post | 9.2 | ± | 72.8 | 0.0 | ± | 0.1 | ||||
| Quercus hypoleucoides | 70 | pre | 176.0 | ± | 279.8 | ** | 1.8 | ± | 6.8 | ** |
| post | 4.0 | ± | 25.5 | 0.0 | ± | 0.0 | ||||
| TOTAL | pre | 585.6 | ± | 619.9 | ** | 29.0 | ± | 32.4 | ** | |
| post | 108.1 | ± | 200.5 | 1.4 | ± | 5.1 | ||||
| Species | Acronym | Lifeform | NMDS1 | NMDS2 | r2 | P-Value | Significance |
|---|---|---|---|---|---|---|---|
| Agave palmeri | AGPAL | shrub | 0.967 | 0.256 | 0.064 | 0.001 | *** |
| Arbutus arizonica | ARAR | tree | −0.273 | 0.962 | 0.055 | 0.001 | *** |
| Arctostaphylos pungens | ARPU | shrub | −0.972 | −0.236 | 0.141 | 0.001 | *** |
| Cercocarpus montanus | CEMO | shrub | 0.488 | −0.873 | 0.006 | 0.434 | |
| Daslylironwrightii | DAWH | shrub | 0.652 | −0.759 | 0.034 | 0.004 | ** |
| Frangula betulafolia | FRBE | shrub | 0.225 | 0.974 | 0.020 | 0.061 | |
| Garrya wrightii | GAWR | shrub | 0.014 | 1.000 | 0.007 | 0.352 | |
| Hesperocyparis ariconica | HEAR | tree | −0.522 | 0.853 | 0.141 | 0.001 | *** |
| Juniperus deppeana | JUDE | tree | 0.433 | 0.901 | 0.342 | 0.001 | *** |
| Mimosa aculeaticarpa | MIAT | shrub | 0.998 | −0.059 | 0.134 | 0.001 | *** |
| Nolina microcarpa | NOMI | shrub | 0.042 | 0.999 | 0.023 | 0.061 | |
| Opuntia spinosior | OPSP | shrub | 0.803 | 0.596 | 0.125 | 0.001 | *** |
| Pinus arizonica | PIAR | tree | −0.890 | 0.455 | 0.175 | 0.001 | *** |
| Pinus discolor | PIDI | tree | −0.789 | 0.615 | 0.095 | 0.001 | *** |
| Pinus edulis | PIED | tree | 0.511 | 0.859 | 0.087 | 0.001 | *** |
| Pinus engelmannii | PIEN | tree | −0.425 | 0.905 | 0.054 | 0.001 | *** |
| Pinus leiophylla | PILE | tree | −0.787 | 0.617 | 0.087 | 0.001 | *** |
| Platanus wrightii | PLWR | tree | −0.155 | 0.988 | 0.103 | 0.001 | *** |
| Prosopis glandulosa | PRGL | shrub | 0.980 | 0.201 | 0.064 | 0.001 | *** |
| Pseudotsuga menzeisii | PSME | tree | −0.927 | 0.376 | 0.050 | 0.002 | ** |
| Quercus arizonica | QUAR | shrub/tree | 0.049 | 0.999 | 0.178 | 0.001 | *** |
| Quercus emoryi | QUEM | shrub/tree | 0.468 | 0.884 | 0.088 | 0.001 | *** |
| Quercus hypoleucoides | QUHY | shrub/tree | −0.818 | 0.576 | 0.348 | 0.001 | *** |
| Quercus rugosa | QURU | shrub | −0.998 | 0.070 | 0.091 | 0.001 | *** |
| Quercus toumeyi | QUTO | shrub | −0.795 | −0.606 | 0.065 | 0.001 | *** |
| Rhus choriophylla | RHCH | shrub | 0.935 | −0.354 | 0.008 | 0.306 | |
| Rhrus trilobata | RHTR | shrub | 0.539 | 0.843 | 0.054 | 0.002 | ** |
| Environmental Variables | |||||||
| TRMI | 0.30212 | 0.95327 | 0.1599 | 0.001 | *** | ||
| dNBR | −0.9483 | −0.31738 | 0.0491 | 0.002 | ** | ||
| Elevation | −0.8584 | −0.51298 | 0.4257 | 0.001 | *** | ||
| severity | 0.0257 | 0.031 | * | ||||
| timestep | 0.1454 | 0.001 | *** | ||||
| timestep * vegetation type | 0.4992 | 0.001 | *** | ||||
| Species | Acronym | Pine-Oak Forest | Piñon Forest | Juniper Woodland | Indicator Species P-Value |
|---|---|---|---|---|---|
| Agave palmeri | AGPAL | 1 | 0.537 | 0.004 | 0.012 |
| Arbutus arizonica | ARAR | 0.012 | 1 | 0.913 | 0.036 |
| Arctostaphylos pungens | ARPU | 0.845 | 0.001 | 1 | 0.003 |
| Dasylirion wheeleri | DAWH | 1 | 0.01 | 0.229 | 0.030 |
| Frangula betulifolia | FRBE | 0.225 | 0.974 | 0.020 | 0.061 |
| Garrya wrightii | GAWR | 0.872 | 0.01 | 0.888 | 0.030 |
| Hesperocyparis arizonica | HEAR | 0.221 | 0.802 | 0.433 | 0.527 |
| Juniperus deppeana | JUDE | 0.99 | 0.996 | 0.001 | 0.003 |
| Mimosa aculeaticarpa | MIAT | 0.994 | 0.994 | 0.001 | 0.003 |
| Nolina microcarpa | NOMI | 1 | 0.001 | 0.652 | 0.003 |
| Opuntia spinosior | OPSP | 0.998 | 1 | 0.001 | 0.003 |
| Pinus arizonica | PIAR | 0.001 | 0.982 | 1 | 0.003 |
| Pinus discolor | PIDI | 0.286 | 0.003 | 1 | 0.009 |
| Pinus edulis | PIED | 0.974 | 1 | 0.001 | 0.003 |
| Pinus engelmannii | PIEN | 0.001 | 1 | 0.956 | 0.003 |
| Pinus leiophylla | PILE | 0.002 | 0.958 | 0.937 | 0.006 |
| Platanus wrightii | PLWR | 0.398 | 1 | 0.186 | 0.461 |
| Prosopis glandulosa | PRGL | 1 | 1 | 0.013 | 0.038 |
| Pseudotsuga menziesii | PSME | 0.004 | 0.98 | 0.914 | 0.012 |
| Quercus arizonica | QUAR | 0.016 | 1 | 0.205 | 0.047 |
| Quercus emoryi | QUEM | 1 | 1 | 0.001 | 0.003 |
| Quercus hypoleucoides | QUHY | 0.001 | 0.996 | 1 | 0.003 |
| Quercus rugosa | QURU | 0.179 | 0.853 | 0.502 | 0.447 |
| Quercus toumeyi | QUTO | 1 | 0.001 | 1 | 0.003 |
| Rhus choriophylla | RHCH | 0.953 | 0.102 | 0.512 | 0.276 |
| Rhus trilobata | RHTR | 0.555 | 1 | 0.031 | 0.090 |
| Species | All Transects | Juniper Woodland | Pine-Oak Forest | Piñon Forest | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Pre-Fire | Post-Fire | Pre-Fire | Post-Fire | PRE-FIRE | Post-Fire | Pre-Fire | Post-Fire | |||||
| Agave palmeri | 26.7 | 13.9 | ↓ | 60.7 | 25.8 | ↓ | 3.3 | 2.0 | 34.6 | 8.2 | ||
| Arctostaphylos pungens | 177.4 | 59.4 | ↓ | 95.1 | 32.5 | ↓ | 187.9 | 74.0 | ↓ | 291.5 | 67.7 | ↓ |
| Dasylirion wheeleri | 39.5 | 11.1 | ↓ | 43.9 | 16.3 | ↓ | 1.4 | 2.0 | 72.2 | 12.3 | ↓ | |
| Garrya wrightii | 38.1 | 10.4 | ↓ | 27.5 | 10.8 | ↓ | 32.3 | 2.0 | ↓ | 60.2 | 18.5 | ↓ |
| Hesperocyparis arizonica | 99.4 | 6.9 | ↓ | 94.5 | 9.5 | ↓ | 142.3 | 4.0 | 62.8 | 6.2 | ||
| Juniperus deppeana | 113.7 | 76.5.5 | ↓ | 210.2 | 134.2 | ↓ | 27.5 | 30.0 | 56.3 | 36.9 | ||
| Mimosa aculeaticarpa | 10.1 | 13.9 | 22.4 | 27.1 | 1.3 | 4.0 | 1.3 | 4.1 | ||||
| Nolina microcarpa | 180.9 | 30.1 | ↓ | 170.9 | 25.8 | ↓ | 103.5 | 22.0 | ↓ | 275.7 | 45.1 | ↓ |
| Pinus arizonica | 95.9 | 0 | ↓ | 0.0 | 0.0 | 326.9 | 0.0 | ↓ | 4.1 | 0.0 | ||
| Pinus discolor | 253.3 | 44.0 | ↓ | 221.7 | 5.4 | ↓ | 155.1 | 44.0 | ↓ | 402.0 | 102.6 | ↓ |
| Pinus edulis | 50.3 | 1.7 | ↓ | 111.7 | 1.4 | 9.0 | 0.0 | 0.0 | 4.1 | |||
| Pinus engelmannii | 24.1 | 6.3 | 38.9 | 5.4 | ↓ | 26.0 | 14.0 | ↓ | 0.0 | 0.0 | ||
| Pinus leiophylla | 45.2 | 9.8 | ↓ | 38.2 | 9.5 | ↓ | 81.4 | 16.0 | ↓ | 18.7 | 4.1 | |
| Pseudotsuga menziesii | 25.5 | 0.0 | 4.5 | 0.0 | 80.4 | 0.0 | 1.3 | 0.0 | ||||
| Quercus arizonica | 134.2 | 60.2 | ↓ | 192.3 | 92.2 | ↓ | 171.3 | 48.0 | 8.3 | 24.6 | ||
| Quercus emoryi | 55.2 | 23.1 | 125.2 | 48.8 | ↓ | 1.3 | 2.0 | 4.7 | 6.2 | |||
| Quercus hypoleucoides | 269.9 | 50.4 | ↓ | 51.6 | 24.4 | ↓ | 716.3 | 106.0 | ↓ | 142.3 | 32.8 | ↓ |
| Quercus rugosa | 28.6 | 16.8 | 23.3 | 5.4 | 52.4 | 38.0 | 12.3 | 12.3 | ||||
| Quercus toumeyi | 244.5 | 58.5 | ↓ | 14.1 | 31.2 | ↓ | 56.7 | 48.0 | 785.8 | 110.8 | ↓ | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Poulos, H.M.; Freiburger, M.R.; Barton, A.M.; Taylor, A.H. Mixed-Severity Wildfire as a Driver of Vegetation Change in an Arizona Madrean Sky Island System, USA. Fire 2021, 4, 78. https://doi.org/10.3390/fire4040078
Poulos HM, Freiburger MR, Barton AM, Taylor AH. Mixed-Severity Wildfire as a Driver of Vegetation Change in an Arizona Madrean Sky Island System, USA. Fire. 2021; 4(4):78. https://doi.org/10.3390/fire4040078
Chicago/Turabian StylePoulos, Helen M., Michael R. Freiburger, Andrew M. Barton, and Alan H. Taylor. 2021. "Mixed-Severity Wildfire as a Driver of Vegetation Change in an Arizona Madrean Sky Island System, USA" Fire 4, no. 4: 78. https://doi.org/10.3390/fire4040078
APA StylePoulos, H. M., Freiburger, M. R., Barton, A. M., & Taylor, A. H. (2021). Mixed-Severity Wildfire as a Driver of Vegetation Change in an Arizona Madrean Sky Island System, USA. Fire, 4(4), 78. https://doi.org/10.3390/fire4040078