Changing Climate Suitability for Dominant Eucalyptus Species May Affect Future Fuel Loads and Flammability in Tasmania

Abstract
:1. Introduction
2. Method
2.1. Modelling Platform (The BCCVL)
2.2. Species Occurrence Data and Preparation
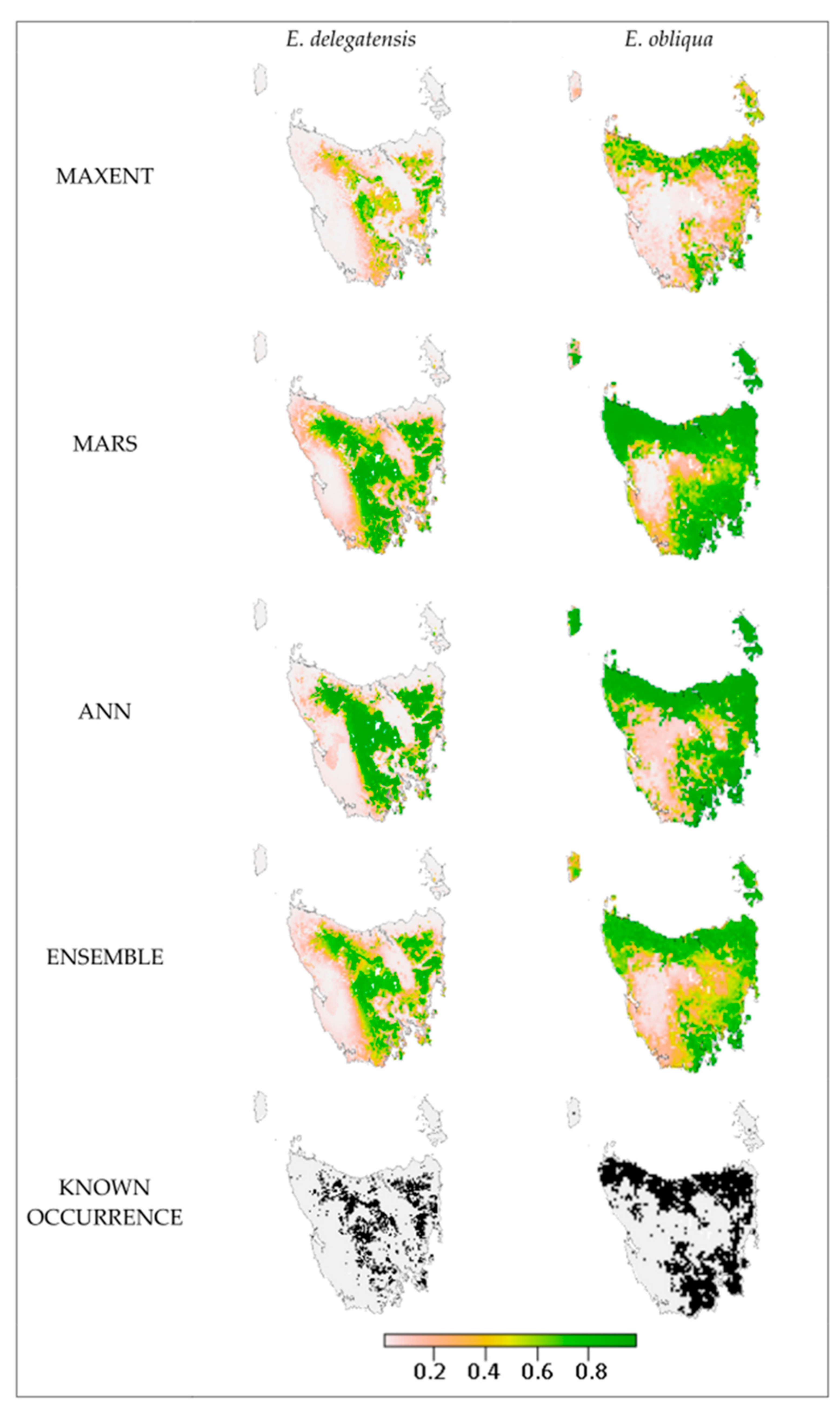
2.3. Species Distribution Models
2.4. Climate Variables and Soil Parameters
2.5. Climate Data—Current and Future
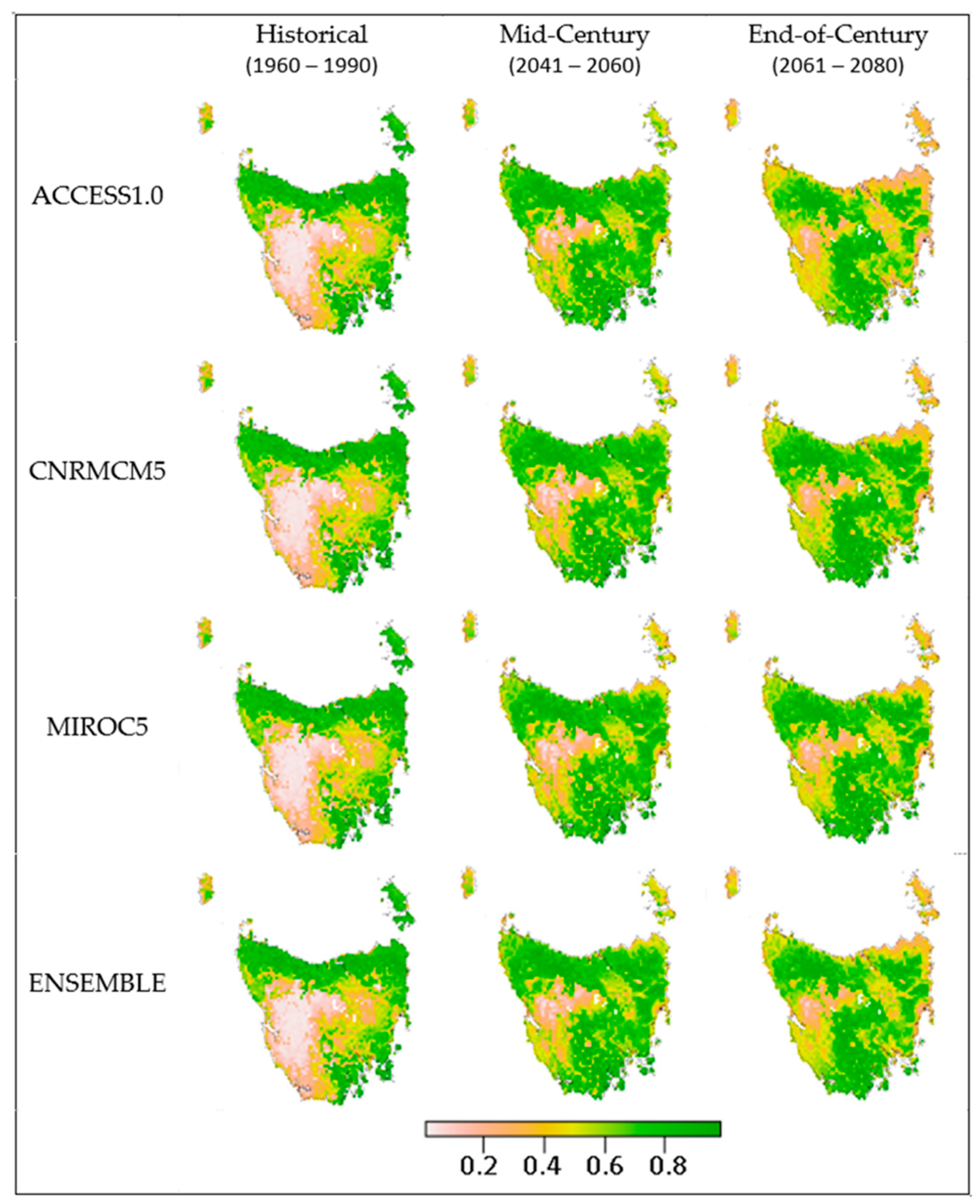
2.6. Global Climate Models
2.7. Emissions Scenarios
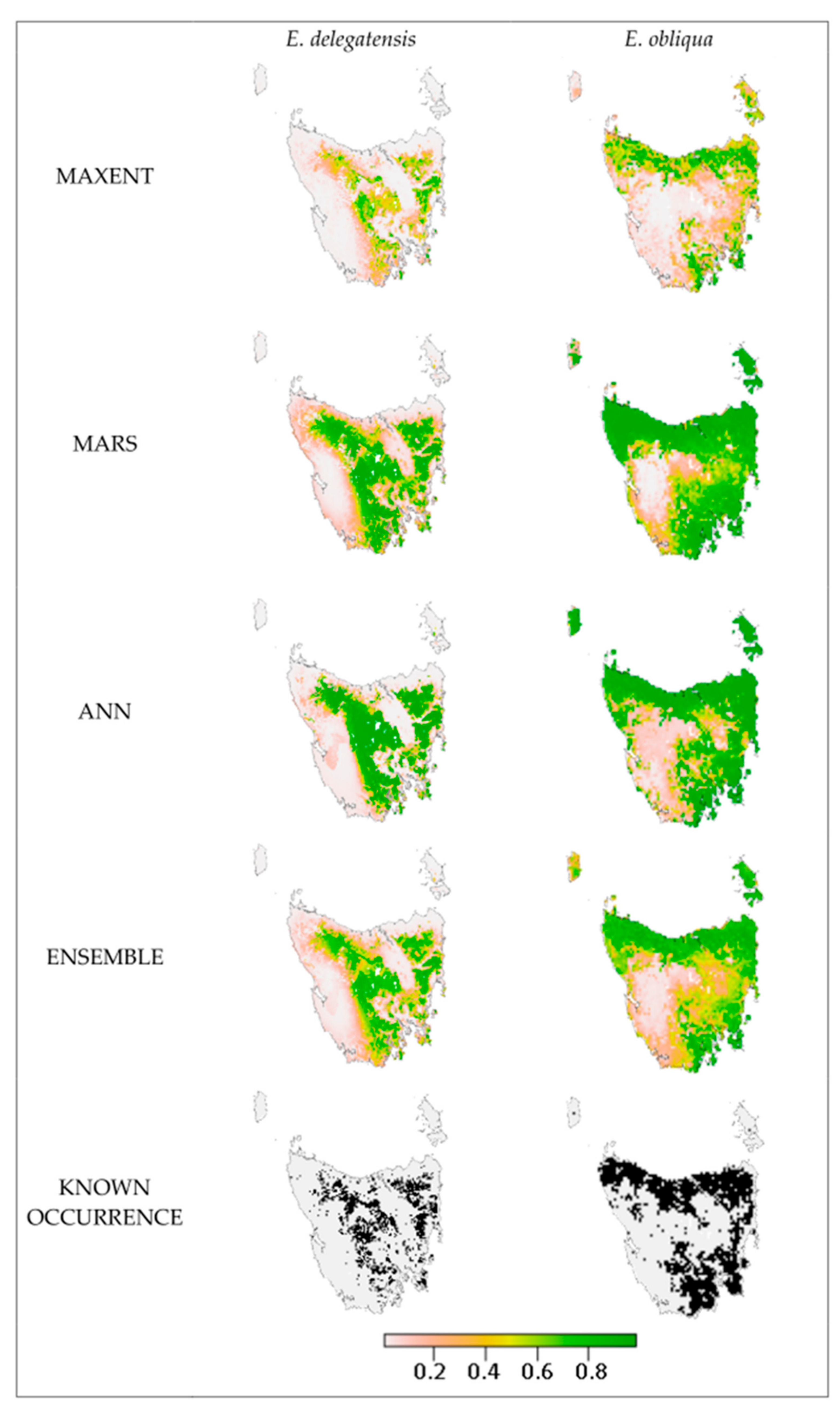
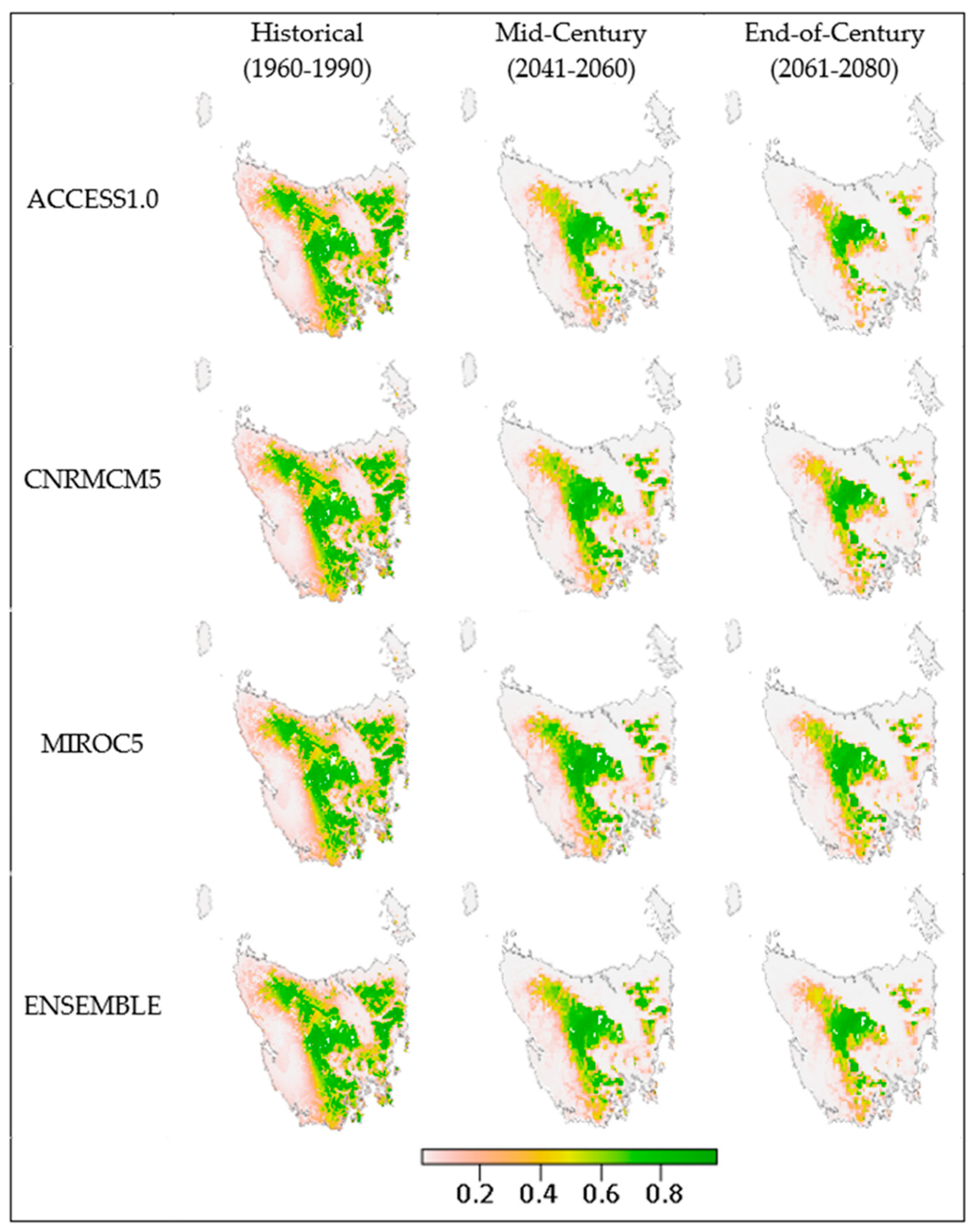
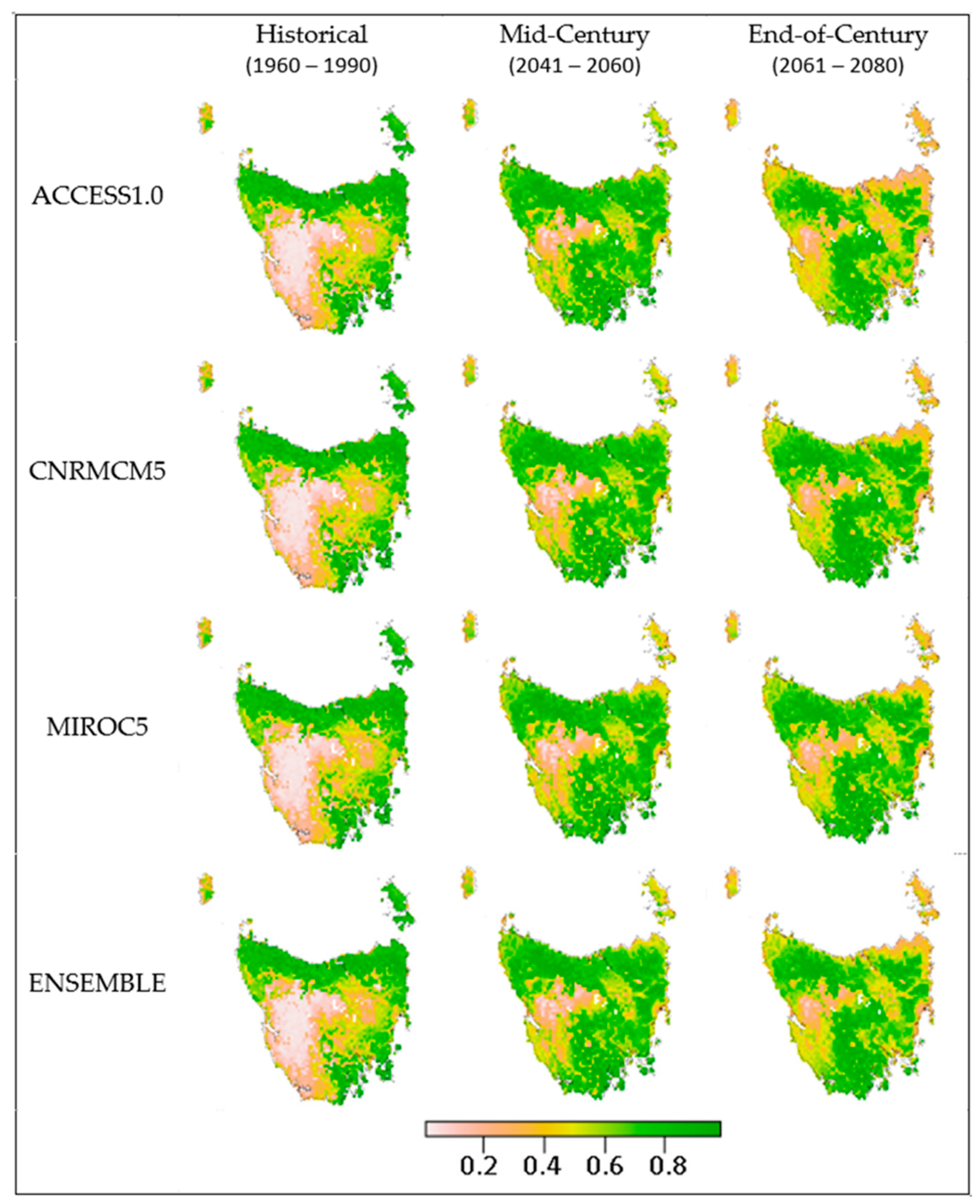
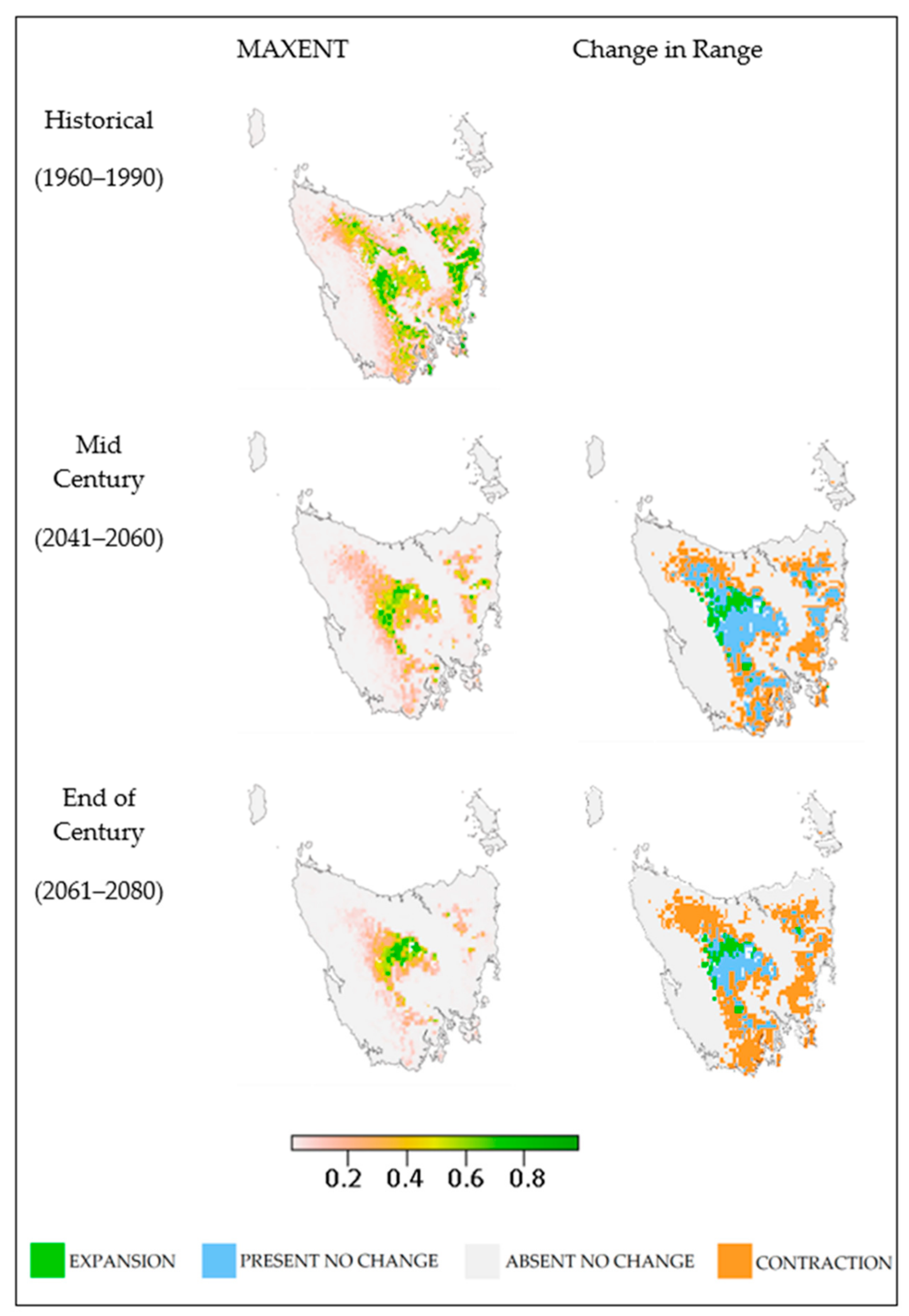
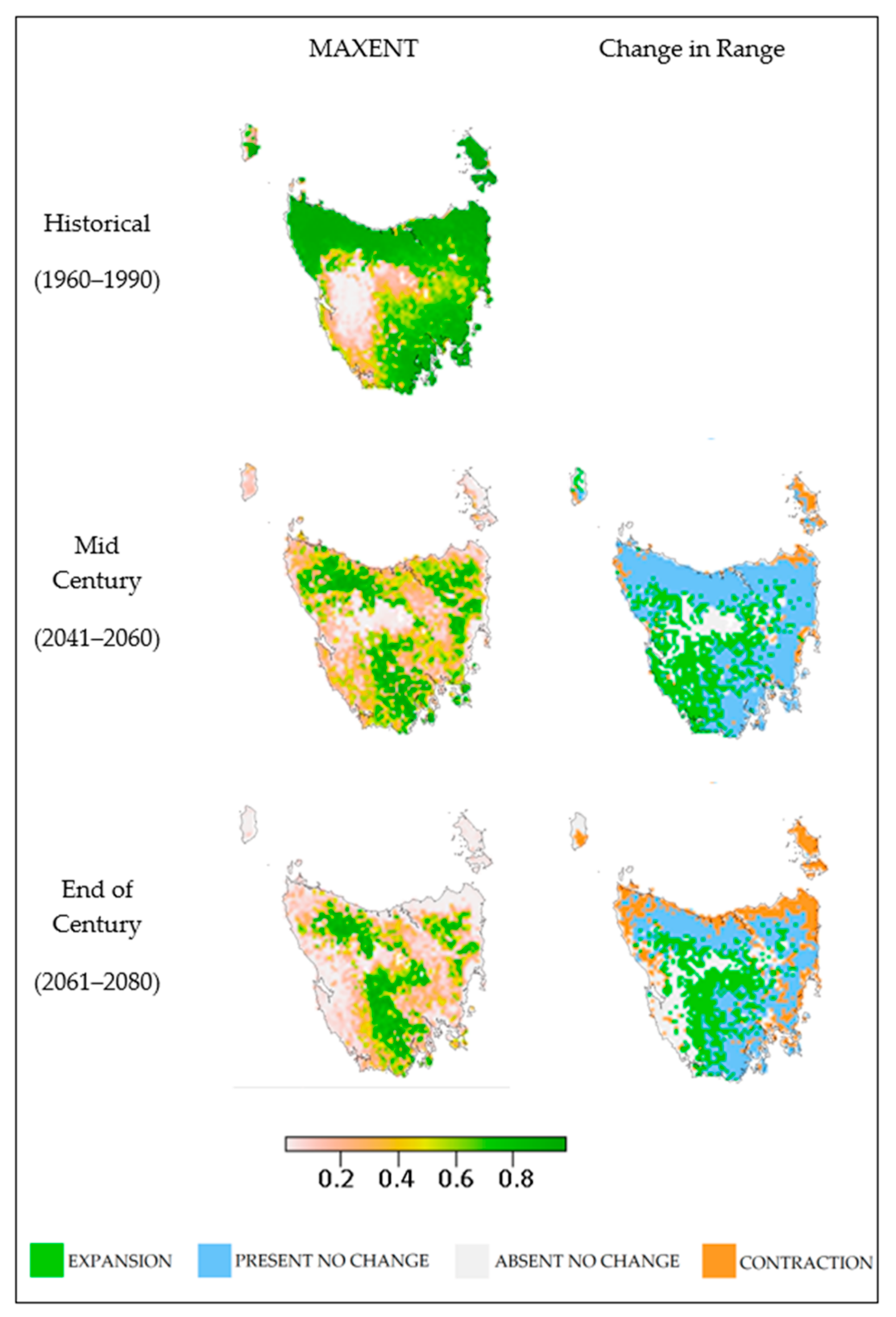
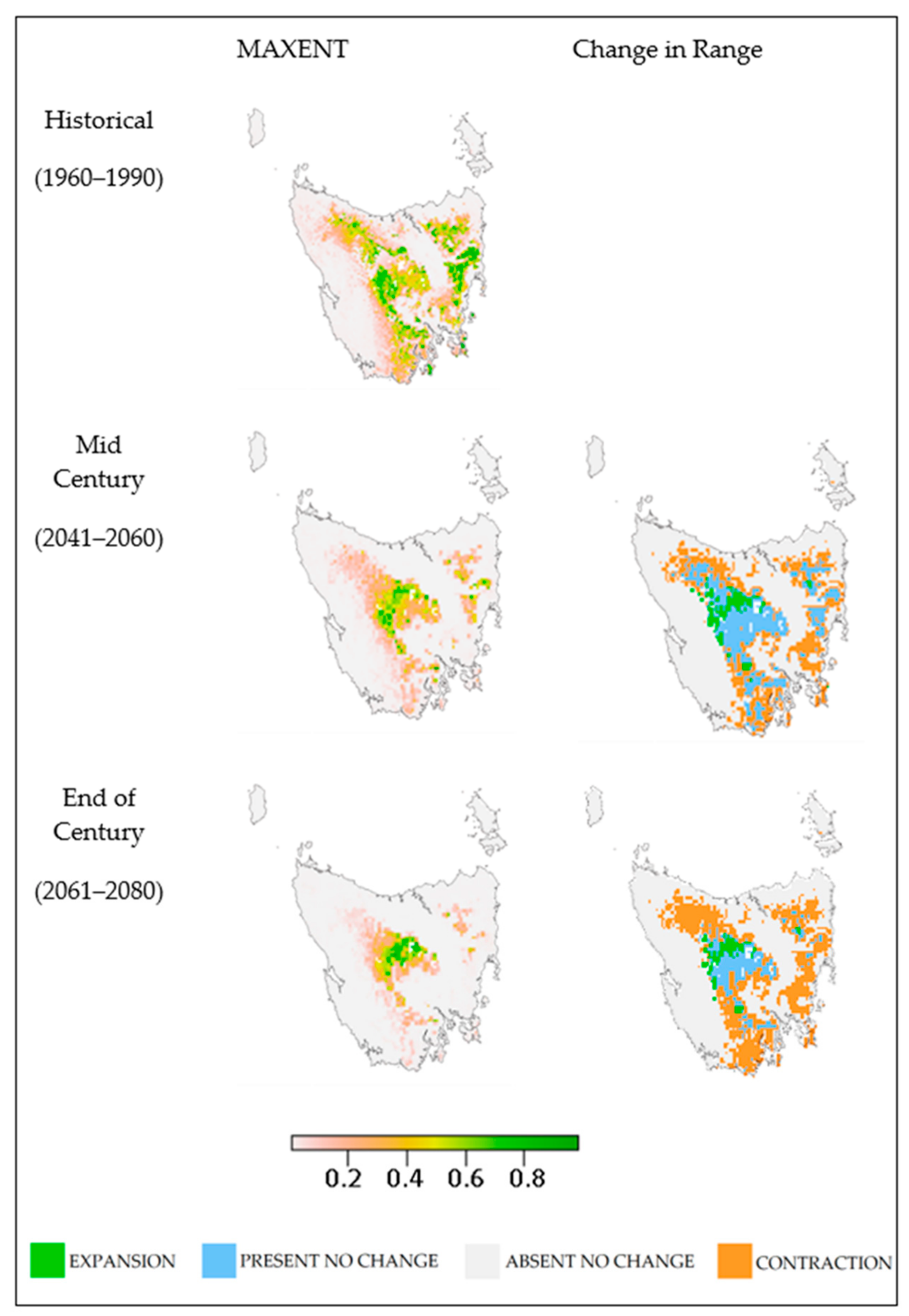
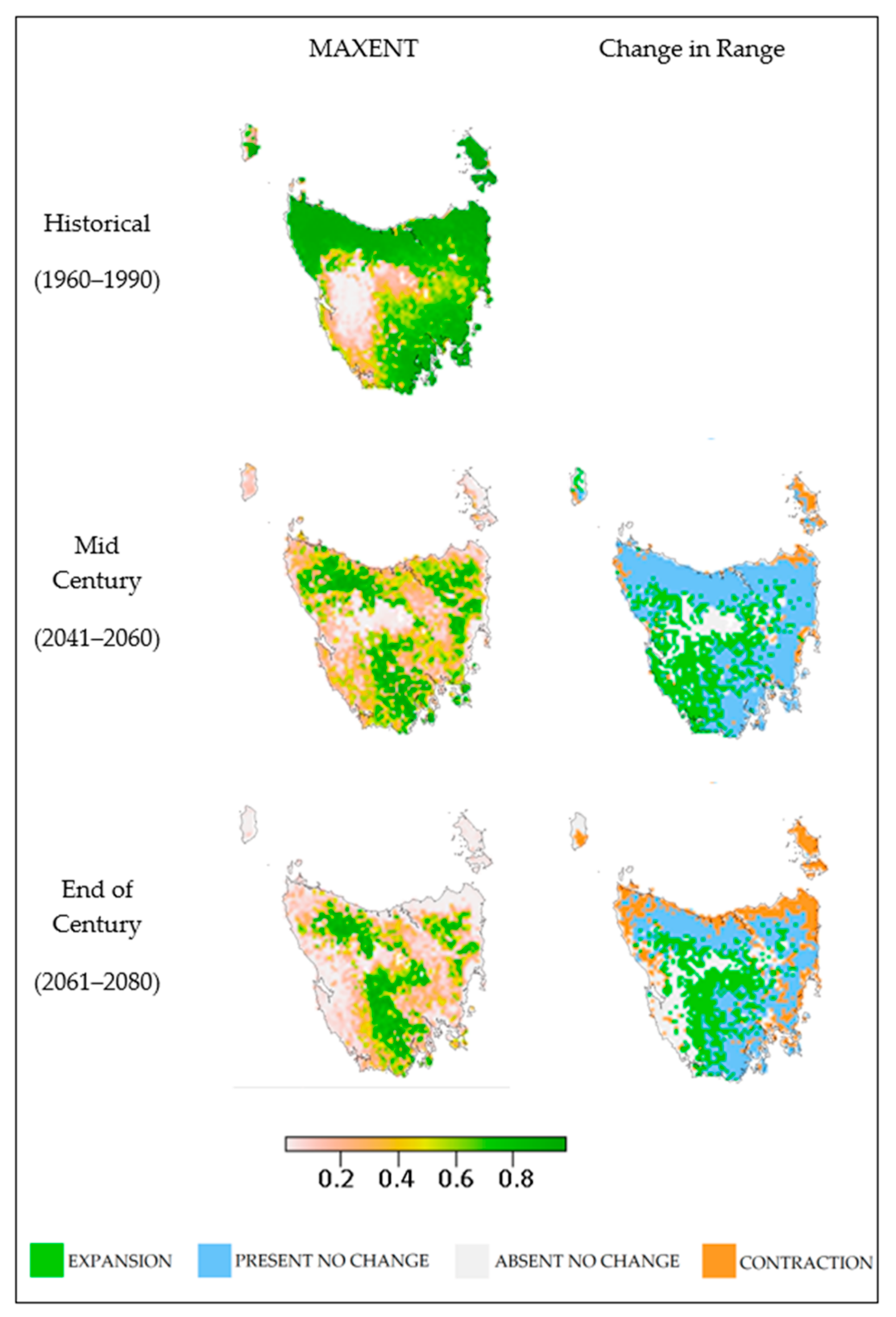
3. Results
4. Discussion
4.1. Outcomes
4.2. Potential Ecological Impacts
4.3. Modelled and Realised Distributions
4.4. Species Variation
4.5. Model Evaluation
4.6. Implications for Fire Management
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Mutlu, M.; Popescu, S.C.; Stripling, C.; Spencer, T. Mapping surface fuel models using lidar and multispectral data fusion for fire behavior. Remote Sens. Environ. 2008, 112, 274–285. [Google Scholar] [CrossRef]
- Clarke, H.; Lucas, C.; Smith, P. Changes in Australian fire weather between 1973 and 2010. Int. J. Clim. 2012, 33, 931–944. [Google Scholar] [CrossRef]
- Bradstock, R.A.; Penman, T.D.; Boer, M.M.; Price, O.; Clarke, H. Divergent responses of fire to recent warming and drying across south-eastern Australia. Glob. Chang. Biol. 2014, 20, 1412–1428. [Google Scholar] [CrossRef] [PubMed]
- Press, T. Tasmanian Wilderness World Heritage Area Bushfire and Climate Change Research Project; Tasmanian Government: Hobart, Australia, 2016.
- Australasian Fire and Emergency Service Authorities Council. AFAC Independent Operational Review: A Review of the Management of the Tasmanian Fires of December 2018–March 2019. Available online: http://www.fire.tas.gov.au/userfiles/AFAC/AFAC_Review.pdf (accessed on 1 March 2020).
- Sharples, J.J.; Cary, G.J.; Fox-Hughes, P.; Mooney, S.; Evans, J.P.; Fletcher, M.-S.; Fromm, M.; Grierson, P.F.; McRae, R.; Baker, P. Natural hazards in Australia: Extreme bushfire. Clim. Chang. 2016, 139, 85–99. [Google Scholar] [CrossRef]
- Fox-Hughes, P.; Harris, R.; Lee, G.; Grose, M.; Bindoff, N. Future fire danger climatology for Tasmania, Australia, using a dynamically downscaled regional climate model. Int. J. Wildland Fire 2014, 23, 309–321. [Google Scholar] [CrossRef]
- Clarke, H.; Pitman, A.; Kala, J.; Carouge, C.; Haverd, V.; Evans, J.P. An investigation of future fuel load and fire weather in Australia. Clim. Chang. 2016, 139, 591–605. [Google Scholar] [CrossRef] [Green Version]
- Pearson, R.G.; Dawson, T.P. Predicting the impacts of climate change on the distribution of species: Are bioclimate envelope models useful? Glob. Ecol. Biogeogr. 2003, 12, 361–371. [Google Scholar] [CrossRef] [Green Version]
- Root, T.L.; Price, J.T.; Hall, K.R.; Schneider, S.H.; Rosenzweigk, C.; Pounds, J.A. Fingerprints of global warming on wild animals and plants. Nat. Cell Biol. 2003, 421, 57–60. [Google Scholar] [CrossRef]
- Hughes, L. Biological consequences of global warming: Is the signal already apparent? Trends Ecol. Evol. 2000, 15, 56–61. [Google Scholar] [CrossRef]
- Harris, R.M.B.; Remenyi, T.A.; Williamson, G.J.; Bindoff, N.L.; Bowman, D.M.J.S. Climate-vegetation-fire interactions and feedbacks: Trivial detail or major barrier to projecting the future of the Earth system? Wiley Interdiscip. Rev. Clim. Chang. 2016, 7, 910–931. [Google Scholar] [CrossRef]
- Cornelissen, J.H.; Grootemaat, S.; Verheijen, L.M.; Cornwell, W.K.; Van Bodegom, P.M.; Van Der Wal, R.; Aerts, R. Are litter decomposition and fire linked through plant species traits? New Phytol. 2017, 216, 653–669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Porfirio, L.L.; Harris, R.M.B.; Lefroy, E.C.; Hugh, S.; Gould, S.F.; Lee, G.; Bindoff, N.L.; Mackey, B. Improving the Use of Species Distribution Models in Conservation Planning and Management under Climate Change. PLoS ONE 2014, 9, e113749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, R.M.B.; Porfirio, L.L.; Hugh, S.; Lee, G.; Bindoff, N.L.; Mackey, B.; Beeton, N. To Be Or Not to Be? Variable selection can change the projected fate of a threatened species under future climate. Ecol. Manag. Restor. 2013, 14, 230–234. [Google Scholar] [CrossRef]
- Matthews, S.; Sullivan, A.L.; Watson, P.; Williams, R.J. Climate change, fuel and fire behaviour in a eucalypt forest. Glob. Chang. Biol. 2012, 18, 3212–3223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, J.K. Surface Fuel Loadings and Predicted Fire Behavior for Vegetation Types in the Northern Rocky Mountains; US Department of Agriculture, Forest Service, Intermountain Research Station: Corvallis, OR, USA, 1986. Available online: https://books.google.com/books?hl=en&lr=&id=sysbvZ_vTyMC&oi=fnd&pg=PP3&dq=Surface+Fuel+Loadings+and+Predicted+Fire+Behaviour+for+Vegetation+Types+in+the+Northern+Rocky+Mountains&ots=rJhHCLRFKD&sig=pyipuLhPRXI68H4eRAwtEzBG2ss (accessed on 10 December 2020).
- Arroyo, L.A.; Pascual, C.; Manzanera, J.A. Fire models and methods to map fuel types: The role of remote sensing. For. Ecol. Manag. 2008, 256, 1239–1252. [Google Scholar] [CrossRef] [Green Version]
- Cassandra, V.A.; Van Altena, C.; Van Logtestijn, R.; Cornwell, W.; Cornelissen, J.H. Species composition and fire: Non-additive mixture effects on ground fuel flammability. Front. Plant Sci. 2012, 3, 63. [Google Scholar] [CrossRef] [Green Version]
- Gill, A.M.; Zylstra, P. Flammability of Australian forests. Aust. For. 2005, 68, 87–93. [Google Scholar] [CrossRef] [Green Version]
- Potts, B.M.; Williams, K.J. The natural distribution of Eucalyptus species in Tasmania. Tasforests 1996, 8, 39–165. [Google Scholar]
- Williamson, G.J.; Prior, L.D.; Grose, M.R.; Harris, R.M.B.; Bowman, D.M.J.S. Projecting canopy cover change in Tasmanian eucalypt forests using dynamically downscaled regional climate models. Reg. Environ. Chang. 2014, 14, 1373–1386. [Google Scholar] [CrossRef]
- Butt, N.; Pollock, L.J.; McAlpine, C.A. Eucalypts face increasing climate stress. Ecol. Evol. 2013, 3, 5011–5022. [Google Scholar] [CrossRef]
- Hughes, L.; Cawsey, E.M.; Westoby, M. Climatic Range Sizes of Eucalyptus Species in Relation to Future Climate Change. Glob. Ecol. Biogeogr. Lett. 1996, 5. [Google Scholar] [CrossRef]
- Garzón, M.B.; De Dios, R.S.; Sainz, H. Effects of climate change on the distribution of Iberian tree species. Appl. Veg. Sci. 2008, 11, 169–178. [Google Scholar] [CrossRef]
- Booth, T.; Broadhurst, L.; Pinkard, E.; Prober, S.; Dillon, S.; Bush, D.; Pinyopusarerk, K.; Doran, J.; Ivkovich, M.; Young, A. Native forests and climate change: Lessons from eucalypts. For. Ecol. Manag. 2015, 347, 18–29. [Google Scholar] [CrossRef]
- Hallgren, W.; Beaumont, L.; Bowness, A.; Chambers, L.; Graham, E.; Holewa, H.; Laffan, S.; Mackey, B.; Nix, H.; Price, J.; et al. The Biodiversity and Climate Change Virtual Laboratory: Where ecology meets big data. Environ. Model. Softw. 2016, 76, 182–186. [Google Scholar] [CrossRef] [Green Version]
- Lucas, J.; Harris, R. Delegatensis Occurrence. 2020. Available online: https://biocache.ala.org.au/occurrences/3febe465-fde2-4e81-8aea-9628380b5e9b (accessed on 28 November 2020).
- Lucas, J.; Harrris, R. Obliqua Occurrence. 2020. Available online: https://doi.ala.org.au/doi/7d232471-18ad-4aec-ba39-f78b9e74749c (accessed on 28 November 2020).
- Gueta, T.; Carmel, Y. Quantifying the value of user-level data cleaning for big data: A case study using mammal distribution models. Ecol. Inform. 2016, 34, 139–145. [Google Scholar] [CrossRef]
- Feeley, K.J.; Silman, M. Modelling the responses of Andean and Amazonian plant species to climate change: The effects of georeferencing errors and the importance of data filtering. J. Biogeogr. 2010, 37, 733–740. [Google Scholar] [CrossRef]
- Lucas, J.; Harris, R.; Mendeley Data. Eucalyptus Obliqua and Eucalyptus Delegatensis Occurances Tasmania (ALA Cleaned). 2020. Available online: https://data.mendeley.com/datasets/bmx6zsjxy8/1 (accessed on 28 November 2020).
- Olden, J.D.; Lawler, J.J.; Poff, N.L. Machine Learning Methods without Tears: A Primer for Ecologists. Q. Rev. Biol. 2008, 83, 171–193. [Google Scholar] [CrossRef] [Green Version]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of MaxEnt for ecologists. Divers. Distrib. 2010, 17, 43–57. [Google Scholar] [CrossRef]
- Friedman, J.H. Multivariate Adaptive Regression Splines. Ann. Stat. 1991, 19, 1–67. [Google Scholar] [CrossRef]
- Breiner, F.T.; Guisan, A.; Bergamini, A.; Nobis, M.P. Overcoming limitations of modelling rare species by using ensembles of small models. Methods Ecol. Evol. 2015, 6, 1210–1218. [Google Scholar] [CrossRef]
- Booth, T.H.; Nix, H.A.; Busby, J.R.; Hutchinson, M.F. bioclim: The first species distribution modelling package, its early applications and relevance to most currentMaxEntstudies. Divers. Distrib. 2014, 20, 1–9. [Google Scholar] [CrossRef]
- Beaumont, L.J.; Hughes, L.; Poulsen, M. Predicting species distributions: Use of climatic parameters in BIOCLIM and its impact on predictions of species’ current and future distributions. Ecol. Model. 2005, 186, 251–270. [Google Scholar] [CrossRef]
- Vanderwal, J.J. All Future Climate Layers for Australia—5 km Resolution. 2012. Available online: https://researchdata.edu.au/future-climate-layers-5km-resolution/10856 (accessed on 28 November 2020).
- National Soil Grids Australia. 2012. Available online: https://www.asris.csiro.au/themes/NationalGrids.html (accessed on 28 November 2020).
- Austin, M.P.; Van Niel, K.P. Impact of landscape predictors on climate change modelling of species distributions: A case study with Eucalyptus fastigata in southern New South Wales, Australia. J. Biogeogr. 2010, 38, 9–19. [Google Scholar] [CrossRef]
- Harris, R.M.B.; Grose, M.R.; Lee, G.; Bindoff, N.L.; Porfirio, L.L.; Fox-Hughes, P. Climate projections for ecologists. Wiley Interdiscip. Rev. Clim. Chang. 2014, 5, 621–637. [Google Scholar] [CrossRef]
- Meteorology, C.a.B.o. Climate Change in Australia. Information for Australia’s Natural Resource Management Regions: Technical Report; CSIRO and Bureau of Meteorology: Canberra, Australia, 2015. [Google Scholar]
- England, M.H.; McGregor, S.; Spence, P.; Meehl, G.A.; Timmermann, A.; Cai, W.; Gupta, A.S.; McPhaden, M.J.; Purich, A.; Santoso, A. Recent intensification of wind-driven circulation in the Pacific and the ongoing warming hiatus. Nat. Clim. Chang. 2014, 4, 222–227. [Google Scholar] [CrossRef]
- Riahi, K.; Rao, S.; Krey, V.; Cho, C.; Chirkov, V.; Fischer, G.; Kindermann, G.; Nakicenovic, N.; Rafaj, P. RCP 8.5—A scenario of comparatively high greenhouse gas emissions. Clim. Chang. 2011, 109, 33–57. [Google Scholar] [CrossRef] [Green Version]
- Peters, G.P.; Andrew, R.M.; Boden, T.; Canadell, J.G.; Ciais, P.; Le Quéré, C.; Marland, G.; Raupach, M.R.; Wilson, C. The challenge to keep global warming below 2 °C. Nat. Clim. Chang. 2012, 3, 4–6. [Google Scholar] [CrossRef]
- BCCVL. SDM—Interpretation of Model Outputs. Available online: https://support.bccvl.org.au/support/solutions/articles/6000127046-sdm-interpretation-of-model-outputs (accessed on 29 June 2020).
- Payne, J.L.; Finnegan, S. The effect of geographic range on extinction risk during background and mass extinction. Proc. Natl. Acad. Sci. USA 2007, 104, 10506–10511. [Google Scholar] [CrossRef] [Green Version]
- Parmesan, C. Range and Abundance Changes. In Biodiversity and Climate Change: Transforming the Biosphere; Lovejoy, T.E., Lee, H., Eds.; Yale University Press: London, UK, 2019. [Google Scholar]
- Parmesan, C.; Yohe, G. A globally coherent fingerprint of climate change impacts across natural systems. Nat. Cell Biol. 2003, 421, 37–42. [Google Scholar] [CrossRef]
- Travis, J.M.; Delgado, M.; Bocedi, G.; Baguette, M.; Bartoń, K.; Bonte, D.; Boulangeat, I.; Hodgson, J.A.; Kubisch, A.; Penteriani, V.; et al. Dispersal and species’ responses to climate change. Oikos 2013, 122, 1532–1540. [Google Scholar] [CrossRef] [Green Version]
- Angert, A.L.; Crozier, L.G.; Rissler, L.J.; Gilman, S.E.; Tewksbury, J.J.; Chunco, A.J. Do species’ traits predict recent shifts at expanding range edges? Ecol. Lett. 2011, 14, 677–689. [Google Scholar] [CrossRef] [PubMed]
- Saura, S.; Bodin, Ö.; Fortin, M.-J. EDITOR’S CHOICE: Stepping stones are crucial for species’ long-distance dispersal and range expansion through habitat networks. J. Appl. Ecol. 2014, 51, 171–182. [Google Scholar] [CrossRef]
- Scheller, R.; Mladenoff, D. Simulated effects of climate change, fragmentation, and inter-specific competition on tree species migration in northern Wisconsin, USA. Clim. Res. 2008, 36, 191–202. [Google Scholar] [CrossRef] [Green Version]
- Von Oheimb, P.V.; Albrecht, C.; Riedel, F.; Bössneck, U.; Zhang, H.; Wilke, T. Testing the role of the Himalaya Mountains as a dispersal barrier in freshwater gastropods (Gyraulus spp.). Biol. J. Linn. Soc. 2013, 109, 526–534. [Google Scholar] [CrossRef] [Green Version]
- Green, D.G.; Sadedin, S. Interactions matter—complexity in landscapes and ecosystems. Ecol. Complex. 2005, 2, 117–130. [Google Scholar] [CrossRef]
- Metzger, J.P.; Martensen, A.C.; Dixo, M.; Bernacci, L.C.; Ribeiro, M.C.; Teixeira, A.M.G.; Pardini, R. Time-lag in biological responses to landscape changes in a highly dynamic Atlantic forest region. Biol. Conserv. 2009, 142, 1166–1177. [Google Scholar] [CrossRef]
- Stoneman, G.L. Ecology and physiology of establishment of eucalypt seedlings from seed: A review. Aust. For. 1994, 57, 11–29. [Google Scholar] [CrossRef]
- Special Climate Statement 73: Extreme Heat and Fire Weather in December 2019 and January 2020. Available online: http://www.bom.gov.au/climate/current/statements/scs73.pdf (accessed on 29 June 2020).
- Hoffmann, A.A.; Rymer, P.; Byrne, M.; Ruthrof, K.X.; Whinam, J.; McGeoch, M.; Bergstrom, D.M.; Guerin, G.R.; Sparrow, B.; Joseph, L.; et al. Impacts of recent climate change on terrestrial flora and fauna: Some emerging Australian examples. Austral Ecol. 2019, 44, 3–27. [Google Scholar] [CrossRef] [Green Version]
- Bowman, D.M.J.S.; Murphy, B.P.; Neyland, D.L.J.; Williamson, G.J.; Prior, L.D. Abrupt fire regime change may cause landscape-wide loss of mature obligate seeder forests. Glob. Chang. Biol. 2014, 20, 1008–1015. [Google Scholar] [CrossRef]
- Harris, R.M.B.; Remenyi, T.; Fox-Hughes, P.; Love, P.; Bindoff, N.L. Exploring the Future of Fuel Loads in Tasmania, Australia: Shifts in Vegetation in Response to Changing Fire Weather, Productivity, and Fire Frequency. Forests 2018, 9, 210. [Google Scholar] [CrossRef] [Green Version]
- Harris, R.M.B.; Beaumont, L.J.; Vance, T.R.; Tozer, C.R.; Remenyi, T.A.; Perkins-Kirkpatrick, S.; Mitchell, P.J.; Nicotra, A.B.; McGregor, S.; Andrew, N.R.; et al. Biological responses to the press and pulse of climate trends and extreme events. Nat. Clim. Chang. 2018, 8, 579–587. [Google Scholar] [CrossRef]
- Parmesan, C.; Root, T.L.; Willig, M.R. Impacts of extreme weather and climate on terrestrial biota. Bull. Am. Meteorol. Soc. 2000, 81, 443–450. [Google Scholar] [CrossRef] [Green Version]
- Seneviratne, S.I.; Nicholls, N.; Easterling, D.; Goodess, C.M.; Kanae, S.; Kossin, J.; Luo, Y.; Marengo, J.; McInnes, K.; Rahimi, M.; et al. Changes in climate extremes and their impacts on the natural physical environment. In Managing the Risks of Extreme Events and Disasters to Advance Climate Change Adaptation; A Special Report of Working Groups I and II of the Intergov-Ernmental Panel on Climate Change (IPCC); Field, C.B., Barros, T.F.V., Stocker, D., Qin, D.J., Dokken, K.L., Ebi, M.D., Mastrandrea, K.J., Mach, G.-K., Plattner, S.K., et al., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2012; pp. 109–230. [Google Scholar]
- Franklin, J. Moving beyond static species distribution models in support of conservation biogeography. Divers. Distrib. 2010, 16, 321–330. [Google Scholar] [CrossRef]
- Harris, D.J. Generating realistic assemblages with a joint species distribution model. Methods Ecol. Evol. 2015, 6, 465–473. [Google Scholar] [CrossRef]
- Radosavljevic, A.; Anderson, R.P. Making better Maxent models of species distributions: Complexity, overfitting and evaluation. J. Biogeogr. 2014, 41, 629–643. [Google Scholar] [CrossRef]
- Turesson, G. The Species and the Variety as Ecological Units. Hereditas 2010, 3, 100–113. [Google Scholar] [CrossRef]
- Turrill, W.B. The Ecotype Concept. A Consideration with Appreciation and Criticism, Especially Oe Recent Trends. New Phytol. 1946, 45, 34–43. [Google Scholar] [CrossRef]
- Anderson, C.; Ladiges, P. A Comparison of Three Populations of Eucalyptus obliqua L’hérit. Growing on Acid and Calcareous Soils in Southern Victoria. Aust. J. Bot. 1978, 26, 93–109. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Algorithm | Description | Strengths | Weaknesses |
|---|---|---|---|
| Artificial Neural Network (ANN) | A collection of models consisting of interconnected nodes arranged in three layers. Layer one is an input layer with a node for each environmental variable (EV), layer two is a hidden layer consisting of different weighted combinations of the EVs, and the third layer is an output layer representing a prediction of presence. The model learns by calculating the difference between the output layer and known occurrences and back propagating to achieve more accurate outcomes [33] | Powerful predictions; Handles large amounts of data; Can model non-linear relationships | Does not deal well with missing or outlying data; Inefficient handling of mixed data types; Long processing times |
| Maxent | Maximum entropy modelling finds the most uniform distribution that falls within the limits of the environments observed at known occurrence points. These limits are implemented as constraints of six types, being linear, quadratic, product, threshold, hinge and categorical [34] | Does not require absence data; Compatible with continuous and categorical variables; Considers interactions between variables; Inbuilt mechanisms to avoid overfitting | Provides suitability rather than probability of presence |
| Multivariate Adaptive Regression Splines (MARS) | A model that finds and partitions at appropriate points in environmental data and builds a linear regression model for each partitioned section which is then knotted together. The model has built-in pruning mechanisms to avoid overfitting, and makes no assumptions about response-predictor variable relationships (as linear regression models do) [35]. | Handles many variables; Detects inter-variable interactions; Complex but fast; Handles outliers well | Overfitting; Difficult interpretation; Handles missing data poorly |
| Climate Model | Projection for 2100 in Relation to the CMIP5 Archive Mean |
|---|---|
| ACCESS1.0 | A hot, dry future. Warming exceeds 2.5 °C across most of Australia, and >3.5 °C in central Australia. Drying is projected over most areas, with decreased annual precipitation of 15%. This model shows high skill in modelling historical climate. Maximum consensus for many regions across Australia. |
| MIROC5 | Moderate warming and slight changes in annual precipitation, with declines in north-east Queensland and south-west Australia; Relatively low warming, wetter model, with a 1.5–3 °C temperature increase and −5% to 5% change in annual precipitation. |
| CNRM-CM5 | Hot/wet end of range in Southern Australia. A temperature increase of 1.5–3 °C and −15% to −5% change in annual precipitation. It also has a good representation of extreme El Niño in CMIP5 evaluations [44]. |
| 2050 | 2070 | |||
|---|---|---|---|---|
| km2 | % | km2 | % | |
| Contraction | 18,763 | 56 ± 7.3 | 24,591 | 67 ± 22.7 |
| No Change | 14,500 | 44 ± 7.3 | 8783 | 26 ± 7.8 |
| Expansion | 2097 | 6 ± 1.4 | 1658 | 5 ± 1.8 |
| 2050 | 2070 | |||
|---|---|---|---|---|
| km2 | % | km2 | % | |
| Contraction | 59,264 | 40 ± 13.9 | 81,098 | 55 ± 16.8 |
| No Change | 85,342 | 60 ± 12.1 | 63,474 | 45 ± 16.8 |
| Expansion | 26,085 | 18 ± 5.1 | 24,398 | 17 ± 6.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lucas, J.; Harris, R.M.B. Changing Climate Suitability for Dominant Eucalyptus Species May Affect Future Fuel Loads and Flammability in Tasmania. Fire 2021, 4, 1. https://doi.org/10.3390/fire4010001
Lucas J, Harris RMB. Changing Climate Suitability for Dominant Eucalyptus Species May Affect Future Fuel Loads and Flammability in Tasmania. Fire. 2021; 4(1):1. https://doi.org/10.3390/fire4010001
Chicago/Turabian StyleLucas, Jessica, and Rebecca M. B. Harris. 2021. "Changing Climate Suitability for Dominant Eucalyptus Species May Affect Future Fuel Loads and Flammability in Tasmania" Fire 4, no. 1: 1. https://doi.org/10.3390/fire4010001
APA StyleLucas, J., & Harris, R. M. B. (2021). Changing Climate Suitability for Dominant Eucalyptus Species May Affect Future Fuel Loads and Flammability in Tasmania. Fire, 4(1), 1. https://doi.org/10.3390/fire4010001