
Southernmost Eurasian Record of Reindeer (Rangifer) in MIS 8 at Galería (Atapuerca, Spain): Evidence of Progressive Southern Expansion of Glacial Fauna Across Climatic Cycles

 , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
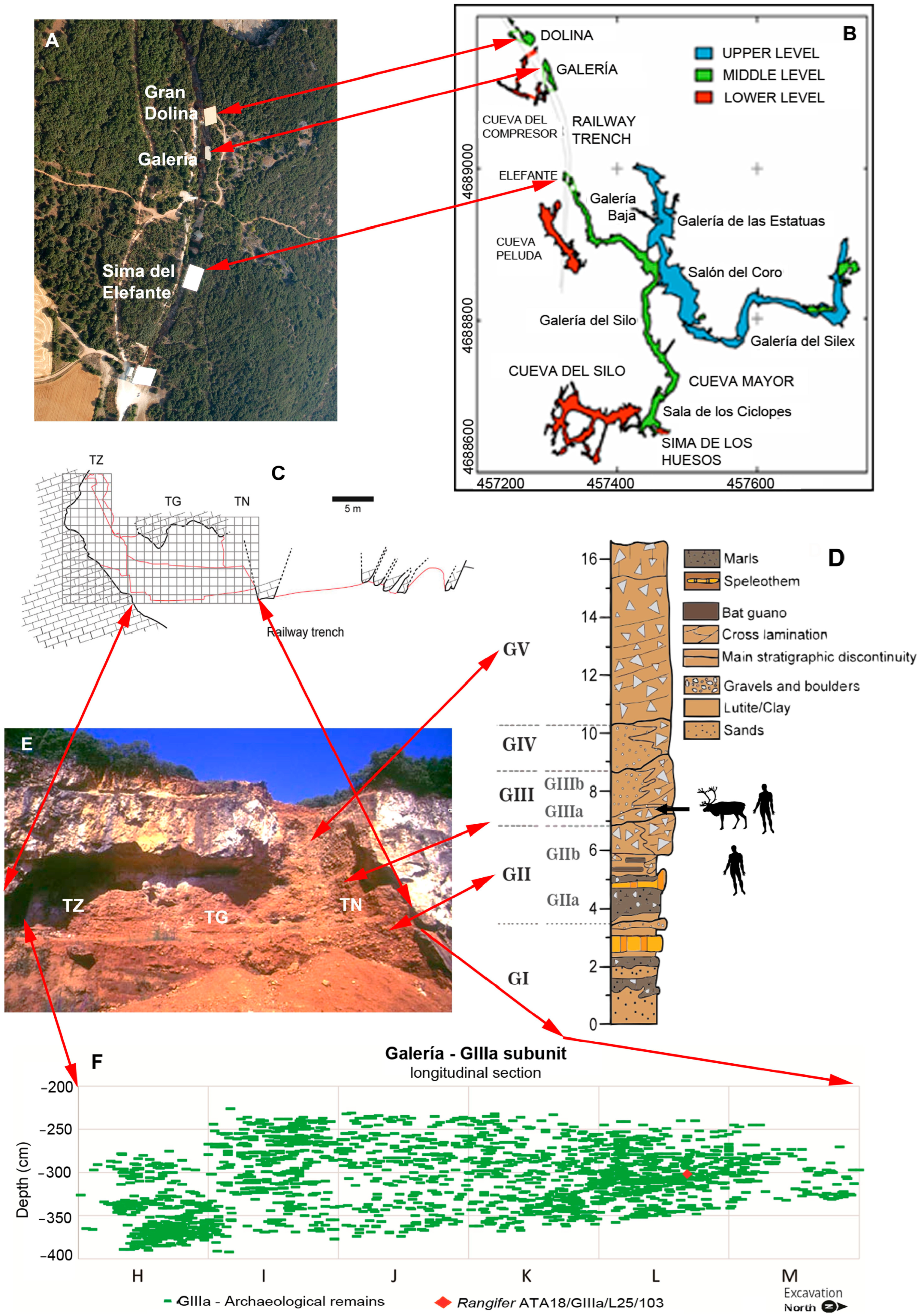
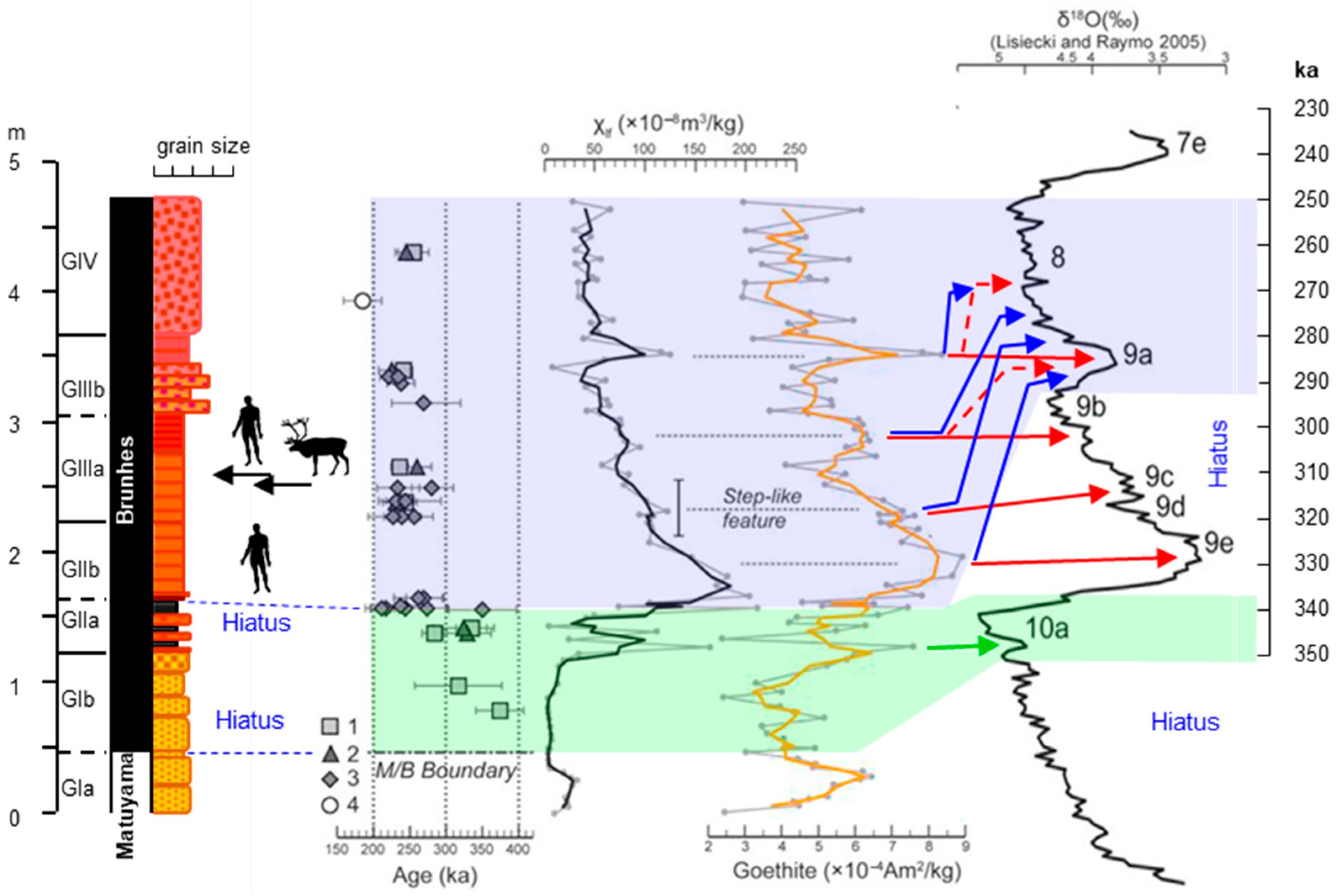
3. Stratigraphy and Dating of Galería—Exact Provenance of the Specimen
4. Results: Description and Comparison
5. Discussion
5.1. Age of the Rangifer Find from Galería
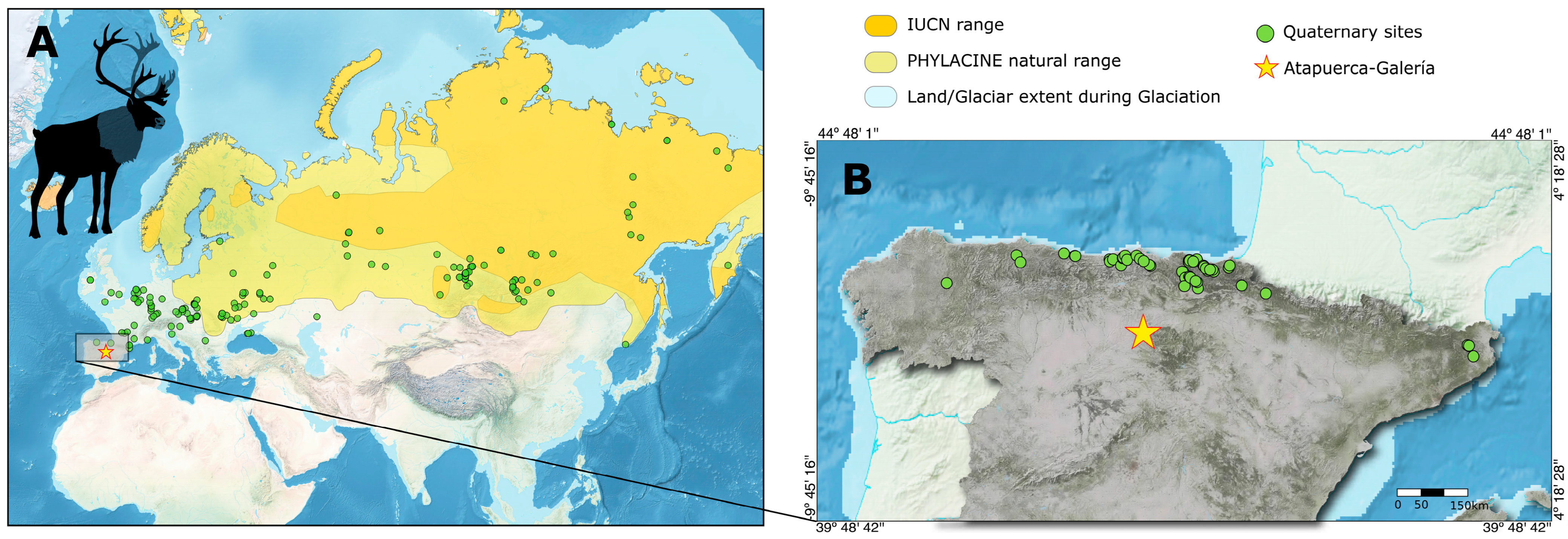
5.2. Rangifer from Galería in the Biogeographic Context of the Genus
5.3. Rangifer from Galería in the Context of the Cold-Adapted Fauna
5.4. The Expansion of Glacial Fauna into the Southern Refugia
5.5. Human Distribution During the Glacial Periods
6. Conclusions
- A single tooth demonstrates the presence of Rangifer in Galería unit GIII;
- This tooth, along with other fossils from the same stratigraphic level, dates to MIS 8;
- The accompanying fauna represents a typical “interglacial fauna” that inhabited the glacial refugium that was Spain back then;
- This finding constitutes the southernmost record of a reindeer in Eurasia during MIS 8;
- It also represents the oldest record of a cold-adapted large mammal in the Iberian Peninsula;
- During the subsequent glacial periods, cold-adapted mammals spread further into the Iberian glacial refugium;
- Humans inhabited the Sierra de Atapuerca during a period likely characterized by challenging local climatic conditions, as testified by the presence of reindeer.
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| CENIEH | Centro Nacional de Investigación sobre la Evolución Humana |
| DAP | Antero-posterior diameter (maximum) |
| DAPb | DAP at the base of the crown |
| DTa | Transverse diameter of the anterior lobe |
| DTp | Transverse diameter of the posterior lobe |
| ESR | Electron spin resonance |
| ESR-US | ESR-uranium series |
| IPHES | Institut Català de Paleoecologia Humana i Evolució Social |
| IQW | Institut für Quartärpaläontologie Weimar, now Forschungsstation für Quartärpaläontologie of the Senckenberg Forschungsinstitut und Naturmuseum Frankfurt, Weimar |
| MIS | Marine isotope state |
| NBC | Naturalis Biodiversity Center, Leiden |
| NMM | Natuurhistorisch Museum, Maastricht |
| pIR-IR | Post infrared-infrared stimulated luminescence |
| TD | Trinchera Dolina, the site of Gran Dolina |
| TG | Trinchera Galería, the site or central part of Galería |
| TL | Thermo Luminisence |
| TT-OSL | Thermally transferred optically stimulated luminescence |
| TN | Trinchera Norte, a part of TG |
| TZ | Trinchera Zarpazos, Covacha de los Zarpazos, a part of Galería |
| ZPALUWr | Division of Palaeozoology, Department of Evolutionary Biology and Ecology, University of Wroclaw |
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon | Locality | Collection/Reference | Number | Side | DAP | DAPb | DTa | DTp |
|---|---|---|---|---|---|---|---|---|
| Rangifer | Atapuerca TG | IPHES | Ata18/GIIIa/L25/103 | 20.4 | 18.3 | 12.8 | 14.4 | |
| Maastricht-Belvedère | NMM | 2195 | d | 16.7 | 16.4 | 10.5 | 12.0 | |
| Recent Greenland | NBC | 20389 | s | 14.8 | 14.8 | 10.2 | 11.6 | |
| Recent zoo | NBC | 11665 | s | 14.3 | 13.0 | 9.1 | 10.2 | |
| NBC | NAM55 | s | 14.6 | 12.4 | 8.8 | 10.3 | ||
| NBC | 1 | s | 14.9 | 13.3 | 9.4 | 10.6 | ||
| Recent Lacunacha | IPHES | O-235 | s | 14.1 | 12.4 | 9.7 | 10.6 | |
| Procapreolus | Weze (Poland) | ZPALUWr [91] | MZ370 | d | 11.2 | 10.2 | 7.8 | 9.8 |
| Capreolus | Atapuerca TE7 | IPHES | Ata22/TE7c/G23/32 | d | 11.9 | 10.9 | 7.4 | 8.3 |
| Voigtstedt | IQW [92] | 1966/7411/Voi3595 | s | 11.9 | 10.9 | 8.4 | 8.7 | |
| Recent Lacunacha | IPHES | d | 11.0 | 9.7 | 7.1 | 8.2 | ||
| Alces carnutorum | Untermassfeld | [93] | IQW 1987/22044 (Mei21563) | d | 25.0 | 22.1 |
References
- Ehlers, J.; Gibbard, P.L. (Eds.) Quaternary Glaciations—Extent and Chronology. Part I: Europe; Elsevier Science: San Diego, CA, USA, 2004; Volume 2, ISBN 9780080540146. [Google Scholar]
- Kahlke, R.-D. Die Entstehungs-, Entwicklungs- Und Verbreitungsgeschichte Des Oberpleistozänen Mammuthus-Coelodonta-Faunenkomplexes in Eurasien (Grosssäuger). Abh. Der Senckenberg. Naturforschenden Ges. 1994, 546, 1–115, Figs. 1–23. [Google Scholar]
- Kahlke, R.-D. The Origin of Eurasian Mammoth Faunas (Mammuthus–Coelodonta Faunal Complex). Quat. Sci. Rev. 2014, 96, 32–49. [Google Scholar] [CrossRef]
- Álvarez-Lao, D.J.; García, N. Chronological Distribution of Pleistocene Cold-Adapted Large Mammal Faunas in the Iberian Peninsula. Quat. Int. 2010, 212, 120–128. [Google Scholar] [CrossRef]
- Guthrie, R.D. Frozen Fauna of the Mammoth Steppe: The Story of Blue Babe; The University of Chicago Press: Chicago, IL, USA; London, UK, 1990; pp. 1–323. [Google Scholar]
- Altuna, J. Arqueozoología de Los Niveles Paleolíticos de La Cueva de Abauntz (Arraiz, Navarra). Salduie Estud. Prehist. Arqueol. 2002, 1–26. [Google Scholar] [CrossRef]
- Álvarez Lao, D.J. Revisión Paleontológica de Los Macromamíferos Indicadores de Clima Frío En El Pleistoceno de La Península Ibérica. Ph.D. Thesis, Universidad de Oviedo, Oviedo, Spain, 2007. [Google Scholar]
- Álvarez-Lao, D.J.; García, N. Geographical Distribution of Pleistocene Cold-Adapted Large Mammal Faunas in the Iberian Peninsula. Quat. Int. 2011, 233, 159–170. [Google Scholar] [CrossRef]
- García, N.; Arsuaga, J.L. Last Glaciation Cold-Adapted Faunas in the Iberian Peninnsula. Deinsea 2003, 9, 159–170. [Google Scholar]
- Ortega, A.I. El Yacimiento de Galería (Sierra de Atapuerca). Crónica de Una Intervención. In Ocupaciones Humanas y Paleoecología del Yacimiento de Galería; Díez, J.-C., Rosas, A., Carbonell, E., Eds.; Junta de Castilla y León: Valladolid, Spain, 1999; pp. 13–17. [Google Scholar]
- Gómez-Olivencia, A.; Arceredillo, D.; Álvarez-Lao, D.J.; Garate, D.; San Pedro, Z.; Castaños, P.; Rios-Garaizar, J. New Evidence for the Presence of Reindeer (Rangifer Tarandus) on the Iberian Peninsula in the Pleistocene: An Archaeopalaeontological and Chronological Reassessment: Reindeer (Rangifer tarandus) on the Iberian Peninsula in the Pleistocene. Boreas 2014, 43, 286–308. [Google Scholar] [CrossRef]
- Álvarez-Lao, D.; García, N. A New Site from the Spanish Middle Pleistocene with Cold-Resistant Faunal Elements: La Parte (Asturias, Spain). Quat. Int. 2006, 142–143, 107–118. [Google Scholar] [CrossRef]
- Rodríguez, J.; Willmes, C.; Mateos, A. Shivering in the Pleistocene. Human Adaptations to Cold Exposure in Western Europe from MIS 14 to MIS 11. J. Hum. Evol. 2021, 153, 102966. [Google Scholar] [CrossRef]
- Gunn, A. Rangifer Tarandus. The IUCN Red List of Threatened Species. 2016. Available online: https://www.iucnredlist.org/species/29742/22167140 (accessed on 28 October 2024).
- Faurby, S.; Davis, M.; Pedersen, R.Ø.; Schowanek, S.D.; Antonelli, A.; Svenning, J.-C. PHYLACINE 1.2: The Phylogenetic Atlas of Mammal Macroecology. Ecology 2018, 99, 2626. [Google Scholar] [CrossRef]
- Žliobaitė, I.; Fortelius, M.; Bernor, R.L.; van den Hoek Ostende, L.W.; Janis, C.M.; Lintulaakso, K.; Säilä, L.K.; Werdelin, L.; Casanovas-Vilar, I.; Croft, D.A.; et al. The NOW Database of Fossil Mammals. In Vertebrate Paleobiology and Paleoanthropology; Springer International Publishing: Cham, Switzerland, 2023; pp. 33–42. [Google Scholar] [CrossRef]
- Ollé, A.; Mosquera, M.; Rodríguez-Álvarez, X.P.; García-Medrano, P.; Barsky, D.; de Lombera-Hermida, A.; Carbonell, E. The Acheulean from Atapuerca: Three Steps Forward, One Step Back. Quat. Int. 2016, 411, 316–328. [Google Scholar] [CrossRef]
- Bermúdez de Castro, J.M.; Carbonell, E.; Cáceres, I.; Díez, J.C.; Fernández-Jalvo, Y.; Mosquera, M.; Ollé, A.; Rodríguez, J.; Rodríguez, X.P.; Rosas, A.; et al. The TD6 (Aurora Stratum) Hominid Site. Final Remarks and New Questions. J. Hum. Evol. 1999, 37, 695–700. [Google Scholar] [CrossRef]
- Bermúdez de Castro, J.M.; Arsuaga, J.L.; Carbonell, E.; Rosas, A.; Martínez, I.; Mosquera, M. A Hominid from the Lower Pleistocene of Atapuerca, Spain: Possible Ancestor to Neandertals and Modern Humans. Science 1997, 276, 1392–1395. [Google Scholar] [CrossRef] [PubMed]
- Carbonell, E.; Bermudez de Castro, J.M.; Parés, J.; Pérez-González, A.; Cuenca-Bescós, G.; Ollé, A.; Mosquera, M.; Huguet, R.; Made, J.; Rosas, A.; et al. The First Hominin of Europe. Nature 2008, 452, 465–469. [Google Scholar] [CrossRef] [PubMed]
- Huguet, R.; Rodríguez-Álvarez, X.P.; Martinón-Torres, M.; Vallverdú, J.; López-García, J.; Lozano, M.; Terradillos-Bernal, M.; Expósito, I.; Ollé, A.; Santos, E.; et al. The Earliest Human Face of Western Europe. Nature 2025, 640, 707–713. [Google Scholar] [CrossRef]
- Arsuaga, J.L.; Martínez, I.; Gracia, A.; Carretero, J.M.; Carbonell, E. Three New Human Skulls from the Sima de Los Huesos Middle Pleistocene Site in Sierra de Atapuerca, Spain. Nature 1993, 362, 534–537. [Google Scholar] [CrossRef]
- Cáceres, I.; Huguet, R.; Rosell, J.; Esteban, M.; Saladié, P.S.; Diéz, J.C.; Ollé, A.; Vallverú, J.; García-Medrano, P.; Carbonell, E. El Yacimiento de Galería (Sierra de Atapuerca, Burgos, España): Un Enclave Para La Obtención de Recursos Cárnicos En El Pleistoceno Medio. Zona Arqueol. 2010, 13, 186–196. [Google Scholar]
- García-Medrano, P.; Cáceres, I.; Ollé, A.; Carbonell, E. The Occupational Pattern of the Galería Site (Atapuerca, Spain): A Technological Perspective. Quat. Int. 2017, 433, 363–378. [Google Scholar] [CrossRef]
- Rodríguez, J.; Burjachs, F.; Cuenca-Bescós, G.; García, N.; Van der Made, J.; Pérez González, A.; Blain, H.-A.; Expósito, I.; López-García, J.M.; García Antón, M.; et al. One Million Years of Cultural Evolution in a Stable Environment at Atapuerca (Burgos, Spain). Quat. Sci. Rev. 2011, 30, 1396–1412. [Google Scholar] [CrossRef]
- García, N.; Arsuaga, J.-L. The Carnivore Remains from the Hominid-Bearing Trinchera-Galería, Sierra de Atapuerca, Middle Pleistocene Site (Spain). Geobios 1998, 31, 659–674. [Google Scholar] [CrossRef]
- Van der Made, J. Artiodactylia Del Yacimiento Mesopleistoceno de Galería (Sierra de Atapuerca). In Atapuerca: Ocupaciones Humanas y Paleoecología del Yacimiento de Galería; Díez, J.-C., Rosas, A., Carbonell, E., Eds.; Junta de Castilla y León: Valladolid, Spain, 1999; pp. 143–167. [Google Scholar]
- Van der Made, J. First Description of the Large Mammals from the Locality of Penal, and Updated Faunal Lists for the Atapuerca Ungulates—Equus altidens, Bison and Human Dispersal into Western Europe. Quat. Int. 2013, 295, 36–47. [Google Scholar] [CrossRef]
- Sánchez, B. Perissodactylia Del Yacimiento Del Pleistoceno Medio de Galería:(Sierra de Atapuerca). In Atapuerca: Ocupaciones Humanas y Paleoecología del Yacimiento de Galería; Díez, J.-C., Rosas, A., Carbonell, E., Eds.; Junta de Castilla y León: Valladolid, Spain, 1999; pp. 169–174. [Google Scholar]
- Van der Made, J. The Dwarfed “Giant Deer” Megaloceros matritensis n.sp. from the Middle Pleistocene of Madrid—A Descendant of M. savini and Contemporary to M. giganteus. Quat. Int. 2019, 520, 110–139. [Google Scholar] [CrossRef]
- Ollé, A.; Huguet, R. Secuencia Arqueoestratigráfica Del Yacimiento de Galería, Atapuerca. In Atapuerca: Ocupaciones Humanas y Paleoecología del Yacimiento de Galería; Díez, J.-C., Rosas, A., Carbonell, E., Eds.; Junta de Castilla y León: Valladolid, Spain, 1999; pp. 55–62. [Google Scholar]
- Pérez-González, A.; Parés, J.M.; Carbonell, E.; Aleixandre, T.; Ortega, A.I.; Benito, A.; Martin Merino, M.Á. Géologie de la Sierra de Atapuerca et stratigraphie des remplissages karstiques de Galería et Dolina (Burgos, Espagne). Anthropologie 2001, 105, 27–43. [Google Scholar] [CrossRef]
- Pérez González, A.; Parés, J.M.; Gallardo, J.; Aleixandre, T.; Ortega, A.I.; Pinilla, A. Geología y Estratigrafía Del Relleno de Galería de La Sierra de Atapuerca (Burgos). In Ocupaciones Humanas y Paleoecología del Yacimiento de Galería; Díez, J.-C., Rosas, A., Carbonell, E., Eds.; Junta de Castilla y León: Valladolid, Spain, 1999; pp. 31–42. [Google Scholar]
- Vallverdú, J. Microfacies y Micromorfología de Las Unidades GII y GIII de Galería (Sierra de Atapuerca). In Ocupaciones Humanas y Paleoecología del Yacimiento de Galería; Díez, J.-C., Rosas, A., Carbonell, E., Eds.; Junta de Castilla y León: Valladolid, Spain, 2002; pp. 43–54. [Google Scholar]
- Grün, R.; Aguirre, E. Datación Por “ESR” y Por La Siere Del “U”, En Los Depositos Carsticos de Atapuerca. In El Hombre fósil de Ibeas y el Pleistoceno de la Sierra de Atapuerca; Aguirre, E., Carbonell, E., Bermúdez de Castro, J.M., Eds.; Junta de Castilla y León: Valladolid, Spain, 1987; pp. 201–204. [Google Scholar]
- Aguirre, E. Dépôts fossilifères du karst de atapuerca, premiers 20 ans. Anthropologie 2001, 105, 13–26. [Google Scholar] [CrossRef]
- Berger, G.W.; Pérez-González, A.; Carbonell, E.; Arsuaga, J.L.; Bermúdez de Castro, J.-M.; Ku, T.-L. Luminescence Chronology of Cave Sediments at the Atapuerca Paleoanthropological Site, Spain. J. Hum. Evol. 2008, 55, 300–311. [Google Scholar] [CrossRef]
- Falguères, C.; Bahain, J.-J.; Bischoff, J.L.; Pérez-González, A.; Ortega, A.I.; Ollé, A.; Quiles, A.; Ghaleb, B.; Moreno, D.; Dolo, J.-M.; et al. Combined ESR/U-Series Chronology of Acheulian Hominid-Bearing Layers at Trinchera Galería Site, Atapuerca, Spain. J. Hum. Evol. 2013, 65, 168–184. [Google Scholar] [CrossRef]
- Demuro, M.; Arnold, L.J.; Parés, J.M.; Pérez-González, A.; Ortega, A.I.; Arsuaga, J.L.; Bermúdez de Castro, J.M.; Carbonell, E. New Luminescence Ages for the Galería Complex Archaeological Site: Resolving Chronological Uncertainties on the Acheulean Record of the Sierra de Atapuerca, Northern Spain. PLoS ONE 2014, 9, e110169. [Google Scholar] [CrossRef]
- Arnold, L.J.; Demuro, M.; Parés, J.M.; Pérez-González, A.; Arsuaga, J.L.; Bermúdez de Castro, J.M.; Carbonell, E. Evaluating the Suitability of Extended-Range Luminescence Dating Techniques over Early and Middle Pleistocene Timescales: Published Datasets and Case Studies from Atapuerca, Spain. Quat. Int. 2015, 389, 167–190. [Google Scholar] [CrossRef]
- Falgéres, C.; Bahain, J.-J.; Yokoyama, Y.; Bischoff, J.L.; Arsuaga, J.L.; de Castro, J.M.B.; Carbonell, E.; Dolo, J.-M. Datation par RPE et U-TH des sites pléistocénes d’Atapuerca: Sima de los Huesos, Trinchera Dolina et Trinchera Galería. bilan géochronologique. Anthropologie 2001, 105, 71–81. [Google Scholar] [CrossRef]
- Bógalo, M.F.; Bradák, B.; Villalaín, J.J.; Calvo-Rathert, M.; González, M.I.; Heller, F.; Ortega, A.I.; Parés, J.M. High-Resolution Late Middle Pleistocene Paleoclimatic Record from the Galería Complex, Atapuerca Archaeological Site, Spain—An Environmental Magnetic Approach. Quat. Sci. Rev. 2021, 251, 106721. [Google Scholar] [CrossRef]
- Ortega, A.I.; Benito-Calvo, A.; Pérez-González, A.; Martín-Merino, M.A.; Pérez-Martínez, R.; Parés, J.M.; Aramburu, A.; Arsuaga, J.L.; Bermúdez de Castro, J.M.; Carbonell, E. Evolution of Multilevel Caves in the Sierra de Atapuerca (Burgos, Spain) and Its Relation to Human Occupation. Geomorphology 2013, 196, 122–137. [Google Scholar] [CrossRef]
- Núñez-Lahuerta, C.; Galán, J.; Cuenca-Bescós, G.; García-Medrano, P.; Cáceres, I. A Bird Assemblage across the MIS 9/8 Boundary: The Middle Pleistocene of Galería (Atapuerca). Quat. Sci. Rev. 2022, 293, 107708. [Google Scholar] [CrossRef]
- Rosas, A.; Bermúdez de Castro, J.M. Descripción y Posición Evolutiva de La Mandíbula AT76-T1H Del Yacimiento de Galería (Sierra de Atapuerca). In Atapuerca: Ocupaciones Humanas y Paleoecología del Yacimiento de Galería; Díez, J.-C., Rosas, A., Carbonell, E., Eds.; Junta de Castilla y León: Valladolid, Spain, 1999; pp. 237–244. [Google Scholar]
- Arsuaga, J.L.; Gracia, A.; Lorenzo, C.; Martínez, I.; Pérez, P.J. Resto Craneal Humano de Galería/Cueva de Los Zarpazos (Sierra de Atapuerca, Burgos). In Atapuerca: Ocupaciones Humanas y Paleoecología del Yacimiento de Galería; Díez, J.-C., Rosas, A., Carbonell, E., Eds.; Junta de Castilla y León Burgos: Valladolid, Spain, 1999; Volume 7, pp. 233–235. [Google Scholar]
- Guthrie, R.D.; Matthews, J.V., Jr. The Cape Deceit Fauna—Early Pleistocene Mammalian Assemblage from the Alaskan Arctic. Quat. Res. 1971, 1, 474–510. [Google Scholar] [CrossRef]
- Van der Made, J.; Stefaniak, K.; Marciszak, A. The Polish Fossil Record of the Wolf Canis and the Deer Alces, Capreolus, Megaloceros, Dama and Cervus in an Evolutionary Perspective. Quat. Int. 2014, 326–327, 406–430. [Google Scholar] [CrossRef]
- Van Kolfschoten, T. The Middle Pleistocene (Saalian) and Late Pleistocene (Weichselian) Mammal Faunas from Maastricht-Belvédère,(Southern Limburg, The Netherlands). Meded. Rijks Geol. Dienst. 1985, 39, 45–74. [Google Scholar]
- Van der Made, J. Caune de l’Arago: Human Fossils, Archaeology and, Last but Not Least, a Long Sequence of Large Mammals Faunas. In Caune de l’Arago, Tautavel-en-Roussion, Pyrénées-Orientales, France. Tome VI. Individualisation des Unités Archéostratigraphiques; De Lumley, H., Ed.; CNRS Éditions: Paris, France, 2015; pp. 623–628. [Google Scholar]
- Soergel, W. Rentiere des deutschen Alt- und Mitteldiluviums. Palaontol. Z. 1941, 22, 387–420. [Google Scholar] [CrossRef]
- Koenigswald, W.; Heinrich, W.D. Mittelpleistozäne Säugetierfaunen Aus Mitteleuropa–Der Versuch Einer Biostratigraphischen Zuordnung. Kaupia 1999, 9, 53–112. [Google Scholar]
- Guérin, C. Les Rhinocéros (Mammalia, Perissodactyla) du Miocène Terminal au Pléistocène Supérieur en Europe Occidentale. Comparaison Avec les Espèces Actuelles (Fascicule 1). Trav. Doc. Lab. Géologie Lyon 1980, 79, 3–421. [Google Scholar]
- Lisiecki, L.E.; Raymo, M.E. A Pliocene-Pleistocene stack of 57 globally distributed benthic D18O records. Paleoceanography 2005, 20, PA1003. [Google Scholar]
- Obermaier, H. El Hombre Fósil, Memoria de La Comisión de Investigaciones Paleontológicas y Prehistóricas; Junta para Ampliación de Estudios e Investigaciones Científicas. Instituto Nacional de Ciencias Físico-Naturales: Madrid, Spain, 1916; Volume 9, p. 255. [Google Scholar]
- Del Pan, I. Paleogeografía de Los Mamíferos Cuaternarios de Europa y Norte de África, Memoria de la Comisión de Investigaciones Paleontológicas y Prehistóricas; Junta para Ampliación de Estudios e Investigaciones Científicas. Instituto Nacional de Ciencias Físico-Naturales: Madrid, Spain, 1918; Volume 21, pp. 1–90. [Google Scholar]
- Van der Made, J.; Rodríguez-Alba, J.J.; Martos, J.A.; Gamarra, J.; Rubio-Jara, S.; Panera, J.; Yravedra, J. The Fallow Deer Dama celiae sp. nov. with Two-Pointed Antlers from the Middle Pleistocene of Madrid, a Contemporary of Humans with Acheulean Technology. Archaeol. Anthropol. Sci. 2023, 15, 41. [Google Scholar] [CrossRef]
- Kurtén, B.; Anderson, E. Pleistocene Mammals of North America; Columbia University Press: New York, NY, USA, 1980. [Google Scholar]
- Webb, S.D. Cervidae and Bovidae. In Evolution of Tertiary Mammals in North America; Janis, C.M., Scott, K.M., Jacobs, L.L., Eds.; Cambridge University Press: Cambridge, UK, 1998; Volume 1, pp. 508–510. [Google Scholar]
- Webb, S. A Cranium of Navahoceros and Its Phylogenetic Place among New World Cervidae. Ann. Zool. Fenn. 1991, 28, 401–410. [Google Scholar]
- Harington, C.R. Pleistocene Vertebrates of the Yukon Territory. Quat. Sci. Rev. 2011, 30, 2341–2354. [Google Scholar] [CrossRef]
- Martin, R.A.; Sneed, J.M. Late Pleistocene Records of Caribou and Elk from Georgia and Alabama. Ga. J. Sci. 1989, 47, 117–122. [Google Scholar]
- Kahlke, H.D. Die Cerviden-Reste Aus Den Kiesen von Süssenborn Bei Weimar; Akademie-Verlag: Berlin, Germany, 1969. [Google Scholar]
- de Lumley, H. (Ed.) Caune de l’Arago: Tautavel-En-Roussillon, Pyrénées-Orientales, France Tome VI; CNRS éditions: Paris, France, 2014; pp. 1–641. [Google Scholar]
- Sardella, R.; Caloi, L.; Di Stefano, G.; Palombo, M.R.; Petronio, C.; Abbazzi, L.; Azzaroli, A.; Ficcarelli, G.; Mazza, P.; Mezzabotta, C.; et al. Mammal Faunal Turnover in Italy from the Middle Pliocene to the Holocene. In The Dawn of the Quaternary; T. van Kolfschoten, P.L.G., Ed.; Mededelingen Nederlands Instituut voor Toegepaste Geowetenschappen: Haarlem, The Netherlands, 1998; pp. 499–512. [Google Scholar]
- Soergel, W. Der Klimacharakter Der Als Nordisch Geltenden Säugetiere Des Eiszeitalters. In Sitzungsberichte der Heidelberger Akademie der Wissenschaften, Mathematisch-Naturwissenschaftliche Klasse; Heidelberg Academy of Sciences: Heidelberg, Germany, 1943; Volume 1941, pp. 3–36. [Google Scholar]
- Mattioli, S. Family Cervidae (Deer). In Handbook of the Mammals of the World; Wilson, D.E., Mittermeier, R.A., Eds.; Lynx: Barcelona, Spain, 2011; Volume 2, pp. 350–443. [Google Scholar]
- Geist, V. Deer of the World: Their Evolution, Behaviour, and Ecology; Stackpole Books: Mechanicsburg, PA, USA, 1998; Volume 272. [Google Scholar]
- Kahlke, H.-D. Die Rhinocerotiden-Reste Aus Den Kiesen von Süssenborn Bei Weimar. Paläontologische Abh. A 1969, 3, 667–709. [Google Scholar]
- Kahlke, H.D. Die Ovibos-Reste Aus Den Kiesen von Süssenborn Bei Weimar. Paläontologische Abh. 1969, 3, 521–529. [Google Scholar]
- Kahlke, H.-D. Die Soergelia-Reste Aus Den Kiesen von Süssenborn Bei Weimar. Paläontologische Abh. 1969, 3, 531–545. [Google Scholar]
- Fejfar, O. Die Nager Aus Den Kiesen von Süßenborn Bei Weimar. Paläontologische Abh. Abt. A 1969, 3, 761–770. [Google Scholar]
- Aguirre, E.; Lhenaff, R.; Zazo, C. Nuevos Fósiles de Elefantes En Andalucía. Estud. Geológicos 1973, 29, 295–306. [Google Scholar]
- Ferreras, P.; Aguirre, E. Rinocerontes Lanudos En La Provincia de Madrid (Coelodonta antiquitatis Blumenbach). Boletín Real Soc. Españolade Hist. Natural. Sección Geológica 1979, 77, 23–59. [Google Scholar]
- Buffon, G.L.L. Les Époques de la Nature; De l’Imprimerie royale: Paris, France, 1780; Volume 1. [Google Scholar]
- Matthew, W.D. Climate and Evolution. Revised and Enlarged. Spec. Publ. N.Y. Acad. Sci. 1939, 1, 1–223. [Google Scholar]
- Penck, A.; Brückner, E. Die Alpen Im Eiszeitalter; Tauchnitz: Leipzig, Germany, 1909. [Google Scholar]
- Penck, A. Säugetierfauna Und Paläolithikum Des Jüngeren Pleistozäns in Mitteleuropa; Verlag der Akademie der Wissenschaften: Berlin, Germany, 1938. [Google Scholar]
- Soergel, W. Zur Biologischen Beurteilung Diluvialer Sängetierfaunen; Sitzungsberichte der Heidelberger Akademie der Wissenschaften, Mathematisch-naturwissenschaftliche Klasse: Heidelberg, Germany, 1940; Volume 4, pp. 3–39. [Google Scholar]
- Carbonell, E.; Bermúdez de Castro, J.M.; Arsuaga, J.L.; Díez, J.C.; Rosas, A.; Cuenca-Bescós, G.; Sala, R.; Mosquera, M.; Rodríguez, X.P. Lower Pleistocene Hominids and Artifacts from Atapuerca-TD6 (Spain). Science 1995, 269, 826–830. [Google Scholar] [CrossRef]
- Tixier, J.; Roe, D.; Turq, A.; Gibert, J.; Martinez, B.; Arribas, A. Présence d’industries Lithiques Dans Le Pléistocène Inférieur de La Région d’Orce (Grenade, Espagne): Quel Est l’état de La Question? Comptes Rendus De L’académie Des sciences. Série 2. Sci. De La Terre Des Planètes 1995, 321, 71–78. [Google Scholar]
- Arzarello, M.; Marcolini, F.; Pavia, G.; Pavia, M.; Petronio, C.; Petrucci, M.; Rook, L.; Sardella, R. Evidence of Earliest Human Occurrence in Europe: The Site of Pirro Nord (Southern Italy). Sci. Nat. 2007, 94, 107–112. [Google Scholar] [CrossRef]
- Díez Fernández-Lomana, J.-C.; Rosas, A.; Carbonell, E. Atapuerca: Ocupaciones Humanas y Paleoecología Del Yacimiento de Galería; Junta de Castilla y León, Consejería de Educación y Cultura: Valladolid, Spain, 1999; ISBN 9788478468768. [Google Scholar]
- Parfitt, S.A.; Ashton, N.M.; Lewis, S.G.; Abel, R.L.; Coope, G.R.; Field, M.H.; Gale, R.; Hoare, P.G.; Larkin, N.R.; Lewis, M.D.; et al. Early Pleistocene Human Occupation at the Edge of the Boreal Zone in Northwest Europe. Nature 2010, 466, 229–233. [Google Scholar] [CrossRef]
- Parfitt, S.; Barendregt, R.; Breda, M.; Candy, I.; Collins, M.; Coope, G.; Durbidge, P.; Field, M.; Lee, J.R.; Lister, A.; et al. The Earliest Record of Human Activity in Northern Europe. Nature 2005, 438, 1008–1012. [Google Scholar] [CrossRef]
- Despriée, J.; Gageonnet, R. La Très Haute Nappe Alluviale d’âge Pléistocène Inférieur de La Vallée de La Creuse à Eguzon (Indre): Figures de Cryoturbation, Habitats Préhistoriques et Datations Absolues. Bull. Société Géologique Fr. 2003, 174, 383–400. [Google Scholar]
- Urciuoli, A.; Martínez, I.; Quam, R.; Arsuaga, J.L.; Keeling, B.A.; Diez-Valero, J.; Conde-Valverde, M. Semicircular Canals Shed Light on Bottleneck Events in the Evolution of the Neanderthal Clade. Nat. Commun. 2025, 16, 972. [Google Scholar] [CrossRef] [PubMed]
- Castaños, P.; Castaños, J. Kiputz IX: A Rich Simple of Reindeer (Rangifer tarandus Linnaeus, 1758) from the Late Pleistocene of the Iberian Peninsula. Rev. Paléobiologie Genève 2018, 37, 409–420. [Google Scholar]
- Costamagno, S.; Liliane, M.; Cédric, B.; Bernard, V.; Bruno, M. Les Pradelles (Marillac-Le-Franc, France): A Mousterian Reindeer Hunting Camp? J. Anthropol. Archaeol. 2006, 25, 466–484. [Google Scholar] [CrossRef]
- Weinstock, J. Late Pleistocene Reindeer Population in Middle and Western Europe. BioArchaeologia 2000, 3, 1–307. [Google Scholar]
- Stefaniak, K. Neogene and Quaternary Cervidae from Poland; Institute of Systematics and Evolution of Animals Polish Academy of Sciences: Kraków, Poland, 2015; pp. 1–204. [Google Scholar]
- Kahlke, H.D. Die Cerviden-Reste aus den Tonen von Voigtstedt in Thüringen. Paläontologische Abhanlungen 1965, A2, 379–426, pls. 13-22. [Google Scholar]
- Kahlke, H.D. Die Cerviden-Reste aus dem Untepleistozän von Untermassfeld. In Das Pleistozän von Untermassfeld bei Meiningen (Thüringen); Kahlke, R.D., Ed.; Dr Rudolf Habelt GMBH: Bonn, Germany, 1997; pp. 181–275, pls. 34-44. [Google Scholar]



| Taxon | GIIa | GIIb | GIIIa | GIIIb |
|---|---|---|---|---|
| Homo | X? | X | ||
| Acheulean industry | X | X | X | X |
| Vulpes vulpes | X | X | X | |
| Canis lupus | X | |||
| Cuon alpinus europaeus | X | X | X | |
| Mustela nivalis | X | X | X | |
| Mustela putorius | X | X | X | |
| Meles meles | X | X | ||
| Ursus spelaeus | X | |||
| Lynx pardinus spelaeus | X | X | X | |
| Panthera leo | X | X | X | |
| Felis sylvestris | X | |||
| Stephanorhinus hemitoechus | X | X | X | X |
| Equus hydruntinus | X | X | ||
| Equus torralbae | X | X | X | X |
| Capreolus priscus | X | X | ||
| Rangifer | X | |||
| Megaceroides | X | X | X | |
| Dama clactoniana | X | X | X | X |
| Cervus elaphus | X | X | X | X |
| Bison sp. | X | X | X | X |
| Hemitragus bonali | X | X | X | X |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
van der Made, J.; Lazagabaster, I.A.; García-Medrano, P.; Cáceres, I. Southernmost Eurasian Record of Reindeer (Rangifer) in MIS 8 at Galería (Atapuerca, Spain): Evidence of Progressive Southern Expansion of Glacial Fauna Across Climatic Cycles. Quaternary 2025, 8, 43. https://doi.org/10.3390/quat8030043
van der Made J, Lazagabaster IA, García-Medrano P, Cáceres I. Southernmost Eurasian Record of Reindeer (Rangifer) in MIS 8 at Galería (Atapuerca, Spain): Evidence of Progressive Southern Expansion of Glacial Fauna Across Climatic Cycles. Quaternary. 2025; 8(3):43. https://doi.org/10.3390/quat8030043
Chicago/Turabian Stylevan der Made, Jan, Ignacio A. Lazagabaster, Paula García-Medrano, and Isabel Cáceres. 2025. "Southernmost Eurasian Record of Reindeer (Rangifer) in MIS 8 at Galería (Atapuerca, Spain): Evidence of Progressive Southern Expansion of Glacial Fauna Across Climatic Cycles" Quaternary 8, no. 3: 43. https://doi.org/10.3390/quat8030043
APA Stylevan der Made, J., Lazagabaster, I. A., García-Medrano, P., & Cáceres, I. (2025). Southernmost Eurasian Record of Reindeer (Rangifer) in MIS 8 at Galería (Atapuerca, Spain): Evidence of Progressive Southern Expansion of Glacial Fauna Across Climatic Cycles. Quaternary, 8(3), 43. https://doi.org/10.3390/quat8030043