Human Discovery and Settlement of the Remote Easter Island (SE Pacific)


Abstract
:1. Introduction
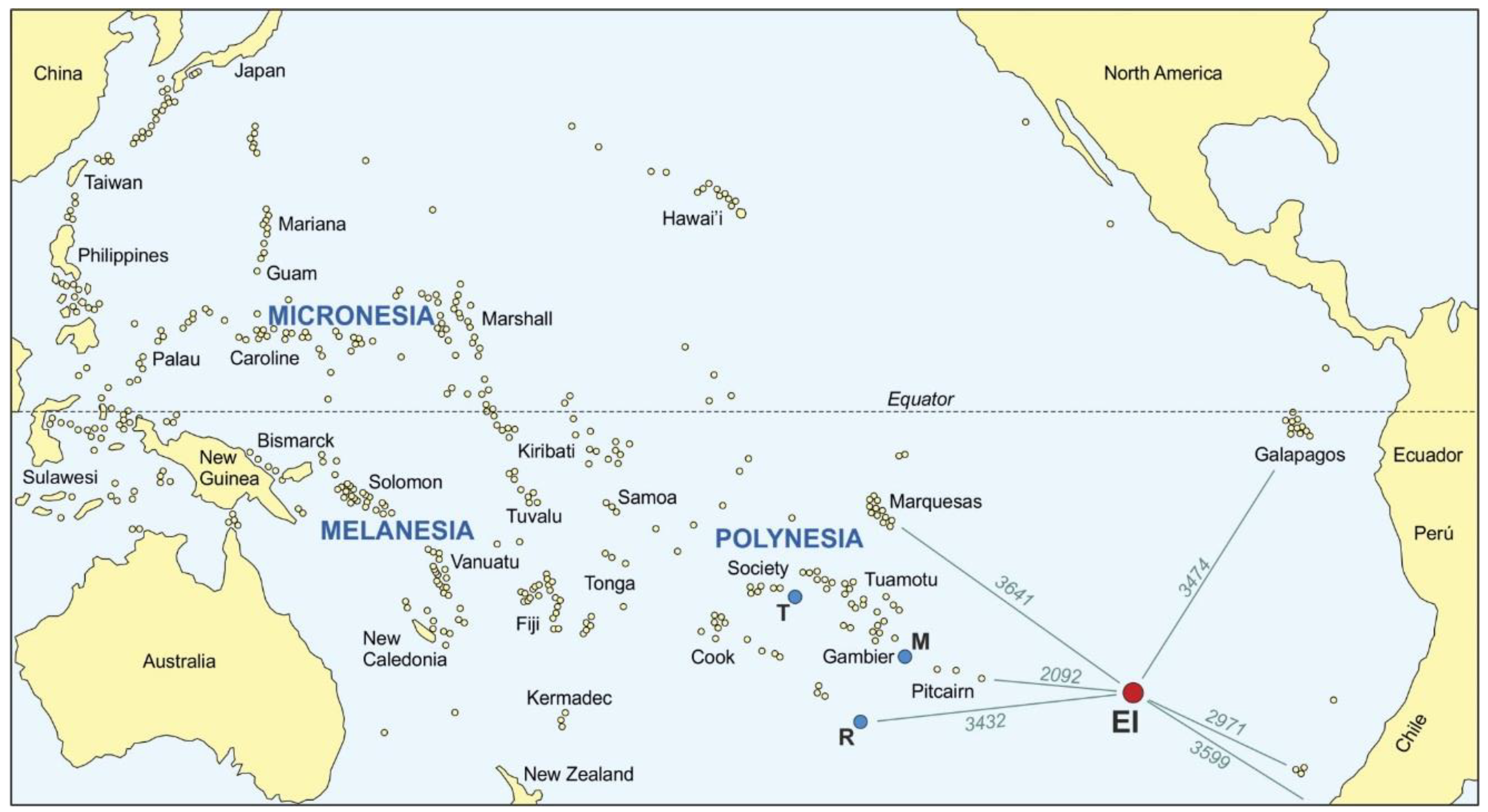
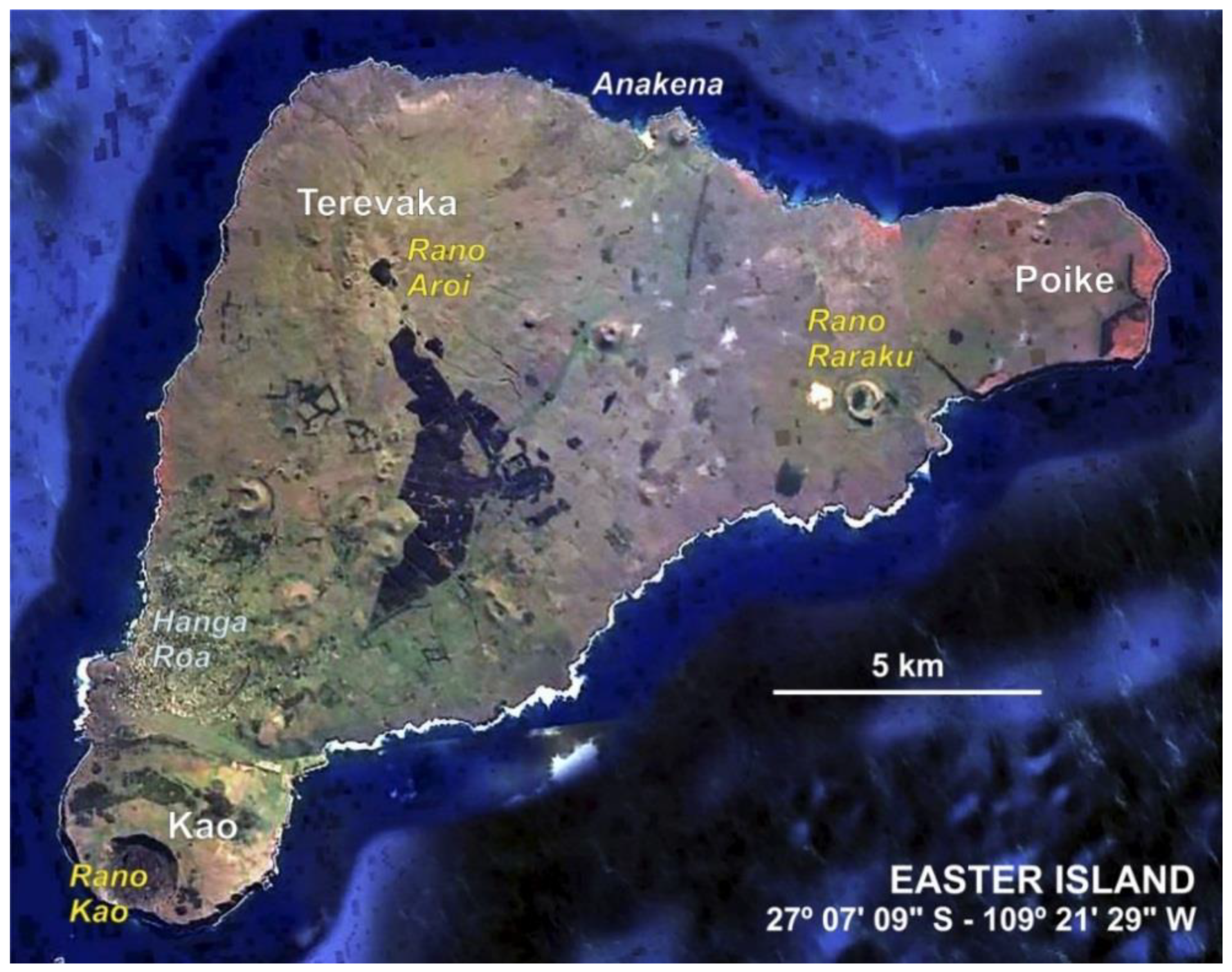
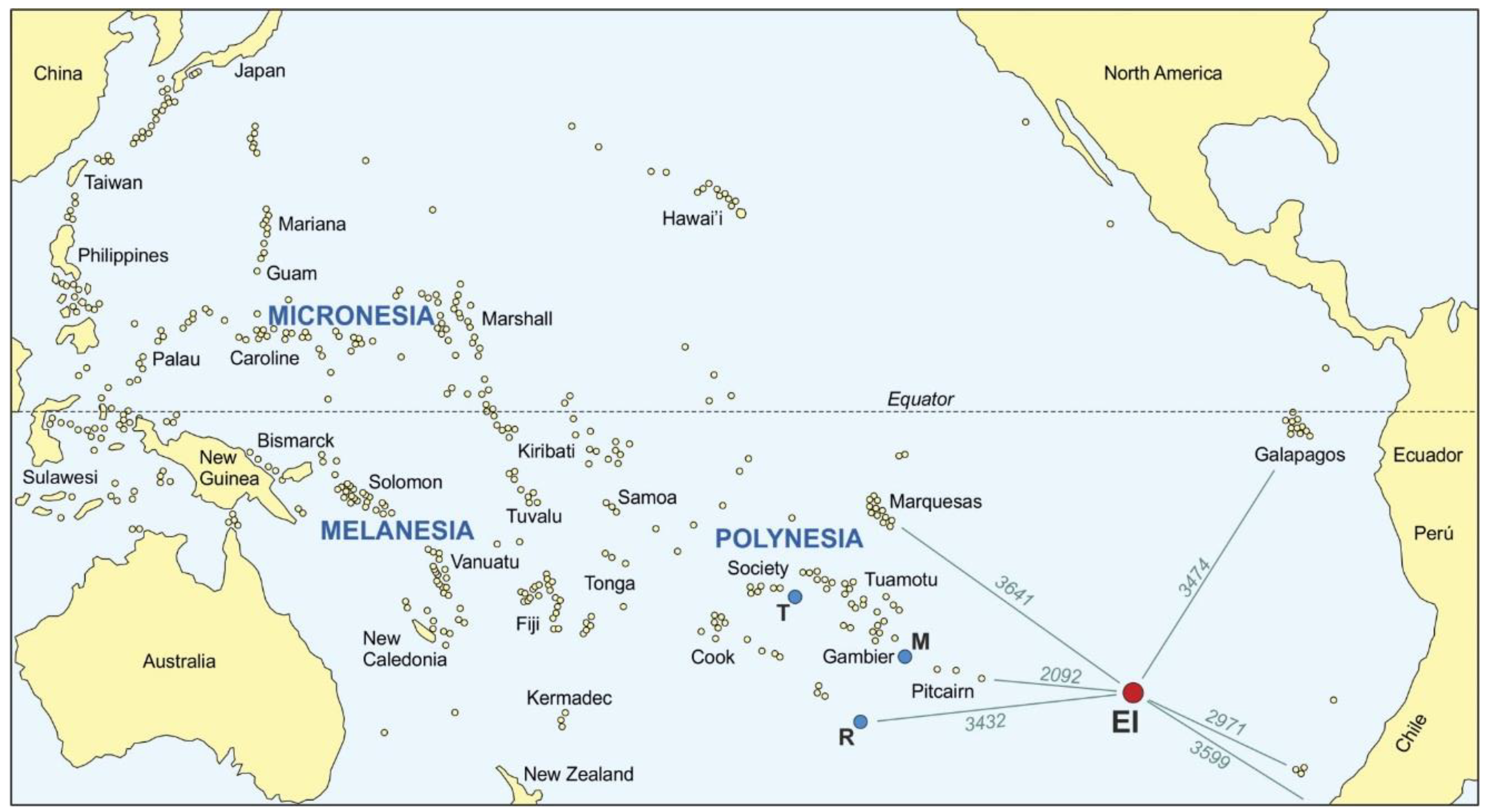
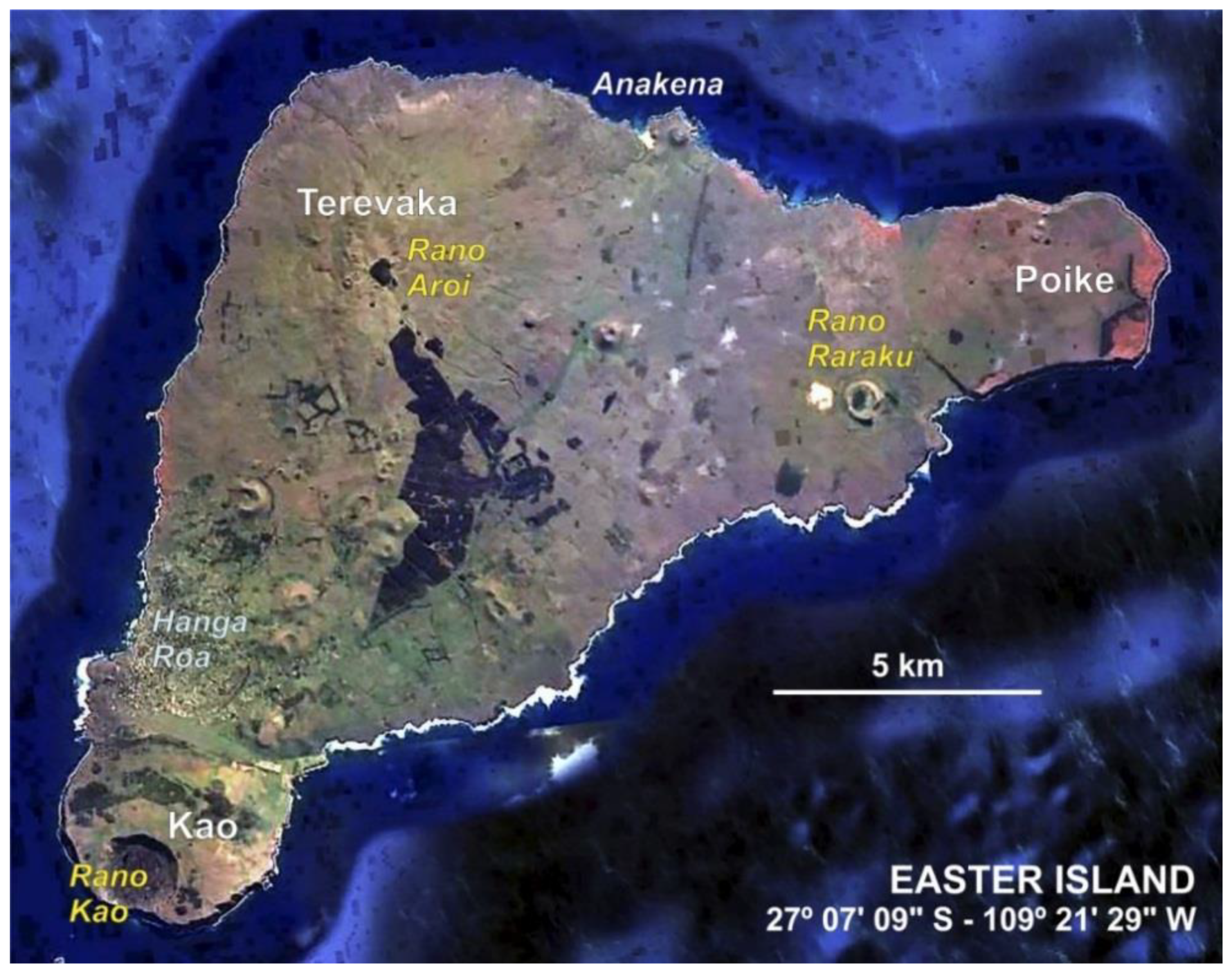
2. The Island Today
3. The East–West Debate
3.1. Rapanui Mythology
3.2. South American Origins
3.3. Polynesian Origins
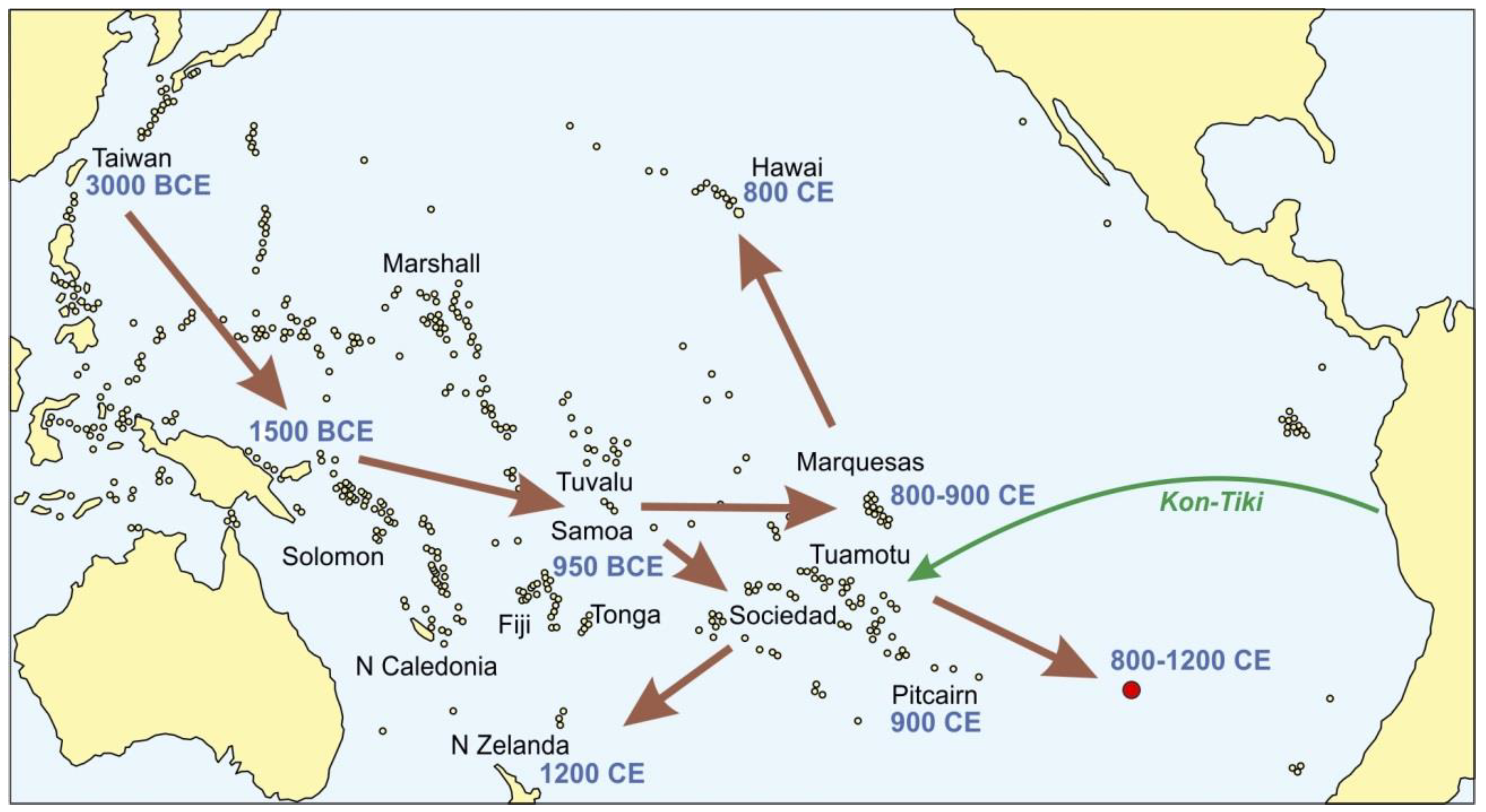
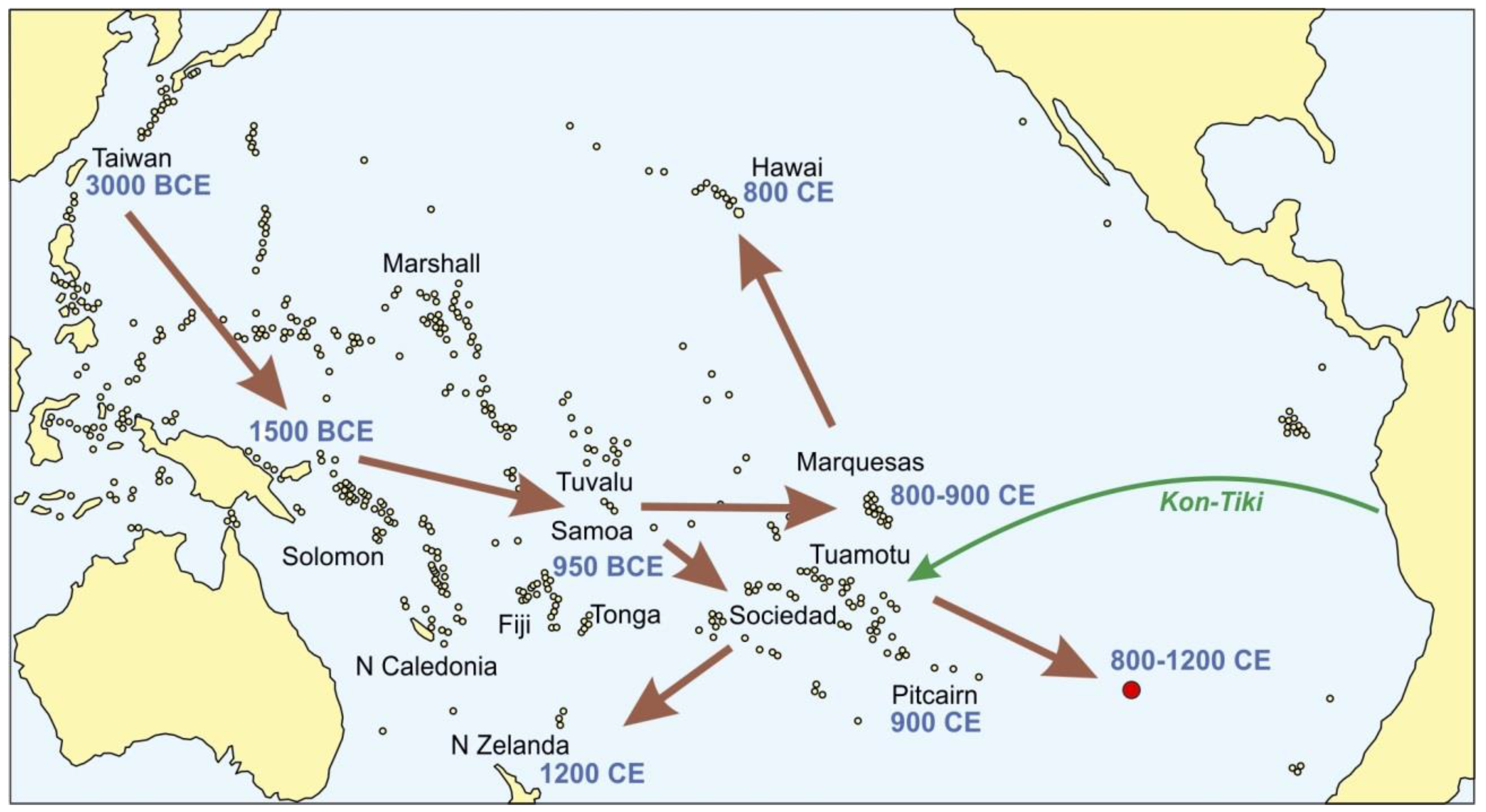
3.4. Peopling of the Pacific
4. The Sweet Potato Enigma
4.1. Long-Distance Dispersal Hypothesis
4.2. Back-and-Forth Hypothesis
4.3. Heyerdahl Hypothesis
4.4. Newcomers Hypothesis
5. New Evidence
5.1. Sweet Potato Phylogeography
5.2. Physical Anthropology
5.3. Paleoecology
6. Conclusions and Final Remarks
Conflicts of Interest
References
- Gillespie, R.G.; Clague, D.A. Encyclopedia of Islands; University California Press: Berkeley, CA, USA, 2009. [Google Scholar]
- McCall, G. Rapanui. Tradition and Survival on Easter Island; University Press Hawaii: Honolulu, HI, USA, 1981. [Google Scholar]
- Flenley, J.; Bahn, P. The Enigmas of Easter Island; Oxford University Press: Oxford, MS, USA, 2003. [Google Scholar]
- Fischer, S.R. Island at the End of the World. The Turbulent History of Easter Island; Reaktion Books: London, UK, 2005. [Google Scholar]
- Hunt, T.; Lipo, C. The Statues that Walked. Unraveling the Mystery of Easter Island; Free Press: New York, NY, USA, 2011. [Google Scholar]
- Rull, V. Natural and anthropogenic drivers of cultural change at Easter Island: Review and new insights. Quat. Sci. Rev. 2016, 150, 31–41. [Google Scholar] [CrossRef]
- Rull, V. La Isla de Pascua. Una Visión Científica; CSIC-La Catarata: Madrid, Spain, 2016. [Google Scholar]
- Rull, V. Strong Fuzzy EHLFS: A general conceptual framework to address past records of environmental, ecological and cultural change. Quaternary 2018, 1, 10. [Google Scholar] [CrossRef]
- Herrera, C.; Custodio, E. Conceptual hydrological model of volcanic Easter Island (Chile) after chemical and isotopic surveys. Hydrogeol. J. 2008, 16, 1329–1348. [Google Scholar] [CrossRef]
- Etienne, M.; Michea, G.; Díaz, E. Flora, Vegetación y Potencial Pastoral de la Isla de Pascua; Facultad Ciencias Agrarias, Veterinarias y Forestales Universidad Chile: Santiago de Chile, Chile, 1982. [Google Scholar]
- Zizka, G. Flowering Plants of Easter Island; Palmarum Hortus Francofurtensis: Frankfurt am Main, Germany, 1991. [Google Scholar]
- Flenley, J.R.; King, S. Late Quaternary pollen records from Easter Island. Nature 1984, 307, 47–50. [Google Scholar] [CrossRef]
- Flenley, J.R.; King, A.S.M.; Jackson, J.; Chew, C. The Late Quaternary vegetational and climatic history of Easter Island. J. Quat. Sci. 1991, 6, 85–115. [Google Scholar] [CrossRef]
- Mieth, A.; Bork, H.R. Humas, climate or introduced rats-which is to blame for the woodland destruction of prehistoric Rapa Nui (Easter Island)? J. Archaeol. Sci. 2010, 37, 417–426. [Google Scholar] [CrossRef]
- Mann, D.; Edwards, J.; Chase, J.; Beck, W.; Reanier., R.; Mass, M.; Finney, B.; Loret, J. Drouhgt, vegetation change, and human history on Rapa Nui (Isla de Pascua, Easter Island). Quat. Res. 2008, 69, 16–28. [Google Scholar] [CrossRef]
- Hunt, T.L. Rethinking Easter Island’s ecological catastrophe. J. Archaeol. Sci. 2007, 34, 485–502. [Google Scholar] [CrossRef]
- Diamond, J. Collapse: How Societies Choose to Fail or Succeed; Viking: New York, NY, USA, 2005. [Google Scholar]
- Rull, V.; Cañellas-Boltà, N.; Sáez, A.; Margalef, O.; Bao, R.; Pla-Rabes, S.; Valero-Garcés, B.; Giralt, S. Challenging Easter Island’s collapse: The need for interdisciplinary synergies. Front. Ecol. Evol. 2013, 1, 3. [Google Scholar] [CrossRef]
- Rull, V.; Cañellas-Boltà, N.; Margalef, O.; Sáez, A.; Pla-Rabes, S.; Giralt, S. Late Holocene vegetation dynamics and deforestation in Rano Aroi: Implications for Easter Island’s ecological and cultural history. Quat. Sci. Rev. 2015, 126, 219–226. [Google Scholar] [CrossRef]
- Rull, V.; Cañellas-Boltà, N.; Margalef, O.; Pla-Rabes, S.; Sáez, A.; Giralt, S. Three millennia of climatic, ecological and cultural change on Easter Island: A synthetic overview. Front. Ecol. Evol. 2016, 4, 29. [Google Scholar] [CrossRef]
- McLaughlin, S. The Complete Guide to Easter Island; Easter Island Foundation: Los Osos, CA, USA, 2007. [Google Scholar]
- Van Tilburg, J.A. Easter Island: Archaeology, Ecology and Culture; Smithsonian Institution Press: Washington, DC, USA, 1994. [Google Scholar]
- Vargas, P.; Cristino, C.; Izaurieta, R. 1000 Años en Rapa Nui. Arqueología del Asentamiento; Editorial Universitaria: Santiago, Chile, 2006. [Google Scholar]
- Routledge, K. The Mystery of Easter Island: The Story of an Expedition; Sifton Praed, Co.: London, UK, 1919. [Google Scholar]
- Métraux, A. Ethnology of Easter Island; Bishop Museum Press: Honolulu, HI, USA, 1940. [Google Scholar]
- Heyerdahl, T. Easter Island. The Mystery Solved; Random House: New York, NY, USA, 1989. [Google Scholar]
- Edwards, E.; Edwards, A. When the Universe Was an Island; Hangaroa Press: Hanga Roa, Chile, 2013. [Google Scholar]
- Heyerdahl, T. American Indians in the Pacific: The Theory Behind the Kon-Tiki Expedition; Allen & Unwin: London, UK, 1952. [Google Scholar]
- Heyerdahl, T.; Ferdon, E. Reports of the Norwegian Archaeological Expedition to Easter Island and the East Pacific. Vol. 1: The Archaeology of Easter Island; Allen & Unwin: London, UK, 1961. [Google Scholar]
- Martinsson-Wallin, H.; Crockford, S.J. Early settlement of Rapa Nui (Easter Island). Asian Perspect. 2002, 40, 244–278. [Google Scholar] [CrossRef]
- Kirch, P.V. Peopling of the Pacific: A holistic anthropological perspective. Annu. Rev. Anthropol. 2010, 39, 131–148. [Google Scholar]
- Wilmshurst, J.M.; Hunt, T.L.; Lipo, C.P.; Anderson, A.J. High-precision radiocarbon dating shows recent and rapid initial human colonization of East Polynesia. Proc. Natl. Acad. Sci. USA 2011, 108, 1815–1820. [Google Scholar] [CrossRef] [PubMed]
- Hunt, T.L.; Lipo, C.P. Late colonization of Easter Island. Science 2006, 311, 1603–1606. [Google Scholar] [CrossRef] [PubMed]
- Nunn, P.D. Climate, Environment and Society in the Pacific during the Last Millennium; Elsevier: Amsterdam, The Netherlands, 2007. [Google Scholar]
- Piperno, D.R.; Holst, I. The presence of starch grains on prehistoric stone tools from the humid Neotropics: Indications of early tuber use and agriculture in Panama. J. Archaeol. Sci. 1998, 25, 765–776. [Google Scholar] [CrossRef]
- Bovell-Benjamin, A.C. Sweet potato: A review of its past, present, and future role in human nutrition. Adv. Food Nutr. Res. 2007, 52, 1–59. [Google Scholar] [PubMed]
- Hather, J.; Kirch, P.V. Prehistoric sweet potato (Ipomoea batatas) from Mangaia Island, central Polynesia. Antiquity 1991, 65, 887–893. [Google Scholar] [CrossRef]
- Orliac, C.; Orliac, M. The disappearance of Easter Island’s forest: Over-exploitation or climatic catastrophe? In Easter Island in Pacific Context; Stevenson, C.M., Lee, G., Morin, F.J., Eds.; Easter Island Foundation: Los Osos, CA, USA, 1998; pp. 129–134. [Google Scholar]
- Orliac, C. The woody vegetation of Easter Island between the early 14th and the mid-17th centuries AD. In Easter Island Archaeology. Research on Early Rapanui Culture; Stevenson, C.M., Ayres, W.S., Eds.; Easter Island Foundation: Los Osos, CA, USA, 2000; pp. 211–220. [Google Scholar]
- McCall, G. Easter Island. In Encyclopedia of Islands; Gillespie, R.G., Clague, D.A., Eds.; University California Press: Berkeley, CA, USA, 2009; pp. 244–251. [Google Scholar]
- Carlquist, S. The biota of long-distance dispersal V. Plant dispersal to Pacific Islands. Bull. Torrey Bot. Club 1967, 44, 129–162. [Google Scholar] [CrossRef]
- Carlquist, S. Island Biology; Columbia University Press: New York, NY, USA, 1974. [Google Scholar]
- Montenegro, A.; Avis, C.; Weaver, A. Modeling the prehistoric arrival of the sweet potato in Polynesia. J. Archaeol. Sci. 2008, 35, 355–367. [Google Scholar] [CrossRef]
- Bourke, R.M. Sweetpotato in Oceania. In The Sweetpotato; Loebenstein, G., Thottappilly, G., Eds.; Springer: Berlin, Germany, 2009; pp. 489–502. [Google Scholar]
- Yen, D.E. The Sweet Potato in Oceania: An Essay in Ethnobotany; Bishop Museum Press: Honolulu, HI, USA, 1974. [Google Scholar]
- Green, R.C. Origin for the Rapanui of Easter Island before European contact: Solutions from holistic anthropology to an issue no longer much of a mystery. Rapa Nui J. 2000, 14, 71–76. [Google Scholar]
- Green, R.C. Sweet potato transfers in Polynesian prehistory. In The Sweet Potato in Oceania: A Reappraisal; Ballard, C., Brown, P., Nourke, R.M., Harwood, T., Eds.; University of Sydney: Sydney, Australia, 2005; pp. 43–62. [Google Scholar]
- Lawler, A. Beyond Kon-Tiki: Did Polynesians sail to South America? Science 2010, 328, 1344–1347. [Google Scholar] [CrossRef]
- Jones, T.L.; Storey, A.A.; Matisoo-Smith, E.; Ramírez-Aliaga, J.M. Polynesians in America. Pre-Columbian Contacts with the New World; Altamira Press: Lanham, MD, USA, 2011. [Google Scholar]
- Roullier, C.; Benoit, L.; McKey, D.B.; Lebot, V. Historical collections reveal patterns of diffusion of sweet potato in Oceania obscured by modern plant movements and recombination. Proc. Natl. Acad. Sci. USA 2013, 110, 2205–2210. [Google Scholar] [CrossRef] [Green Version]
- Muñoz-Rodríguez, P.; Carruthers, T.; Wood, J.R.D.; Williams, B.R.M.; Weitemier, K.; Kronmiller, B.; Ellis, D.; Anglin, N.L.; Longway, L.; Harris, S.A.; et al. Reconciling conflicting phylogenies in the origin of sweet potato and dispersal to Polynesia. Curr. Biol. 2018, 28, 1246–1256. [Google Scholar] [CrossRef]
- Tromp, M.; Dudgeon, J.V. Differentiating dietary and non-dietary microfossils extracted from human dentaal calculus: The importance of sweet potato to ancient diet on Rapa Nui. J. Archaeol. Sci. 2015, 54, 54–63. [Google Scholar] [CrossRef]
- Moreno-Mayar, J.V.; Rasmussen, S.; Seguin-Orlando, A.; Rasmussen, M.; Liang, M.; Flåm, S.T.; Lie, B.A.; Gilfillan, G.D.; Nielsssen, R.; Thorsby, E.; et al. Genome-wide ancestry patterns in Rapanui suggest pre-European admixture with Native Americans. Curr. Biol. 2014, 24, 2518–2525. [Google Scholar] [CrossRef]
- Thorsby, E. Genetic evidence for a contribution of native Americans to the early settlement of Rapa Nui (Easter Island). Front. Ecol. Evol. 2016, 4, 118. [Google Scholar] [CrossRef]
- Fehren-Scmitz, L.; Jarman, C.L.; Harkins, K.M.; Kayser, K.M.; Popp, B.N. Genetic ancestry of Rapanui before and after European contact. Curr. Biol. 2017, 27, 3209–3215. [Google Scholar] [CrossRef]
- Storey, A.A.; Ramírez, J.M.; Quiroz, D.; Burley, D.V.; Addison, D.J.; Walter, R.; Anderson, A.J.; Hunt, T.; Athens, J.S.; Huynen, L.; et al. Radiocarbon and DNA evidence for a pre-Columbian introduction of Polynesian chickens to Chile. Proc. Natl. Acad. Sci. USA 2007, 104, 10335–10339. [Google Scholar] [CrossRef] [Green Version]
- Thomson, V.A.; Lebrasseur, O.; Austin, J.J.; Hunt, T.L.; Burney, D.A.; Denham, T.; Rawlence, N.J.; Wood, J.R.; Flink, L.G.; Linderholm, A.; et al. Using ancient DNA to study the origins and dispersal of ancestral Polynesian chickens across the Pacific. Proc. Natl. Acad. Sci. USA 2014, 111, 4826–4831. [Google Scholar] [CrossRef] [Green Version]
- Matisoo-Smith, E.; Ramirez, J.M. Human skeletal evidence of Polynesian presence in South America? Metric analyses of six crania from Mocha Island, Chile. J. Pac. Archaeol. 2010, 1, 76–88. [Google Scholar]
- Malaspinas, A.-S.; Lao, O.; Schroeder, H.; Rasmussen, M.; Raghavan, M.; Moltke, I.; Campos, P.F.; Sagredo, F.S.; Rasmussen, S.; Gonçalves, V.F.; et al. Two ancient human genomes reveal Polynesian ancestry among the indigenous Botocudos of Brazil. Curr. Biol. 2014, 24, R1035–R1037. [Google Scholar] [CrossRef] [Green Version]
- Rull, V.; Lara, A.; Rubio-Inglés, M.J.; Giralt, S.; Gonçalves, V.; Raposeiro, P.; Hernández, A.; Sánchez-López, G.; Vázquez-Loureiro, D.; Bao, R.; et al. Vegetation and landscape dynamics under natural and anthropogenic forcing on the Azores Islands: A 700-year pollen record from the São Miguel Island. Quat. Sci. Rev. 2017, 159, 155–168. [Google Scholar] [CrossRef] [Green Version]
- Rull, V. The EIRA database: Last Glacial and Holocene radiocarbon ages from Easter Island’s sedimentary records. Front. Ecol. Evol. 2016, 4, 44. [Google Scholar] [CrossRef]
- Haberle, S.G.; Atkin, G. Needles in a stayhack: Searching for the sweet potato (Ipomoea batatas (L.) Lam.) in the fossil pollen record. In The Sweet Potato in Oceania: A Reappraisal; Ballard, C., Brown, P., Nourke, R.M., Harwood, T., Eds.; University of Sydney: Sydney, Australia, 2005; pp. 25–33. [Google Scholar]
- Horrocks, M.; Wozniak, J.A. Plant microfossil analysis reveals disturbed forest and a mixed-crop, dryland production system at Te Nui, Easter Island. J. Archaeol. Sci. 2008, 35, 126–142. [Google Scholar] [CrossRef]
- Horrocks, M.; Baisden, T.; Flenley, J.; Feek, D.; Love, C.; Haoa-Cardinali, S.; González Nualart, L.; Edmunds Gorman, T. Pollen, phytolith and starch analyses of dryland soils from Easter Island (Rapa Nui) show widespread vegetation clearance and Polynesian-introduced crops. Palynology 2016, 41, 339–350. [Google Scholar] [CrossRef]
- Cummings, L.S. A review of recent pollen and phytolith studies from various contexts on Easter Island. In Easter Island in Pacific Context; Stevenson, C.M., Lee, G., Morin, F.J., Eds.; Easter Island Foundation: Los Osos, CA, USA, 1998; pp. 100–106. [Google Scholar]
- Horrocks, M.; Baisden, W.T.; Nieuwoudt, M.K.; Flenley, J.; Feek, D.; González Nualart, L.; Haoa-Cardinali, S.; Edmunds Gorman, T. Microfossils of Polynesian cultigens in lake sediment cores from Rao Kau, Easter Island. J. Paleolimnol. 2012, 47, 185–204. [Google Scholar] [CrossRef]
- Horrocks, M.; Baisden, W.T.; Flenley, J.; Feek, D.; González Nualart, L.; Haoa-Cardinali, S.; Edmunds Gorman, T. Fossil plant remains at Rano Raraku, Easter Island’s statue quarry: Evidence for past elevated lake level and ancient Polynesian agriculture. J. Paleolimnol. 2012, 48, 1–17. [Google Scholar] [CrossRef]
- Cañellas-Boltà, N.; Rull, V.; Sáez, A.; Margalef, O.; Bao, R.; Pla-Rabes, S.; Blaauw, M.; Valero-Garcés, B.; Giralt, S. Vegetation changes and human settlement of Easter Island during the last millennia: A multiproxy study of the Lake Raraku sediments. Quat. Sci. Rev. 2013, 72, 36–48. [Google Scholar] [CrossRef]
- Butler, K.R.; Flenley, J.R. The Rano Kau 2 pollen diagram: Palaeoecology revealed. Rapa Nui J. 2010, 24, 5–10. [Google Scholar]
- Chamberlin, T.C. The method of multiple working hypotheses. Science (Old Ser.) 1980, 15, 92–96, reprinted 1965, 148, 754–759. [Google Scholar]
- Platt, J.R. Strong inference. Science 1964, 146, 347–353. [Google Scholar] [CrossRef] [PubMed]
- Popper, K.R. The Logic of Scientific Discovery; Basic Books: New York, NY, USA, 1959. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
© 2019 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rull, V. Human Discovery and Settlement of the Remote Easter Island (SE Pacific). Quaternary 2019, 2, 15. https://doi.org/10.3390/quat2020015
Rull V. Human Discovery and Settlement of the Remote Easter Island (SE Pacific). Quaternary. 2019; 2(2):15. https://doi.org/10.3390/quat2020015
Chicago/Turabian StyleRull, Valentí. 2019. "Human Discovery and Settlement of the Remote Easter Island (SE Pacific)" Quaternary 2, no. 2: 15. https://doi.org/10.3390/quat2020015
APA StyleRull, V. (2019). Human Discovery and Settlement of the Remote Easter Island (SE Pacific). Quaternary, 2(2), 15. https://doi.org/10.3390/quat2020015