Revisiting the Foraging Ecology and Extinction History of Two Endemic Vertebrates from Tenerife, Canary Islands



Abstract
1. Introduction
1.1. Stable Isotopes in Bone Collagen
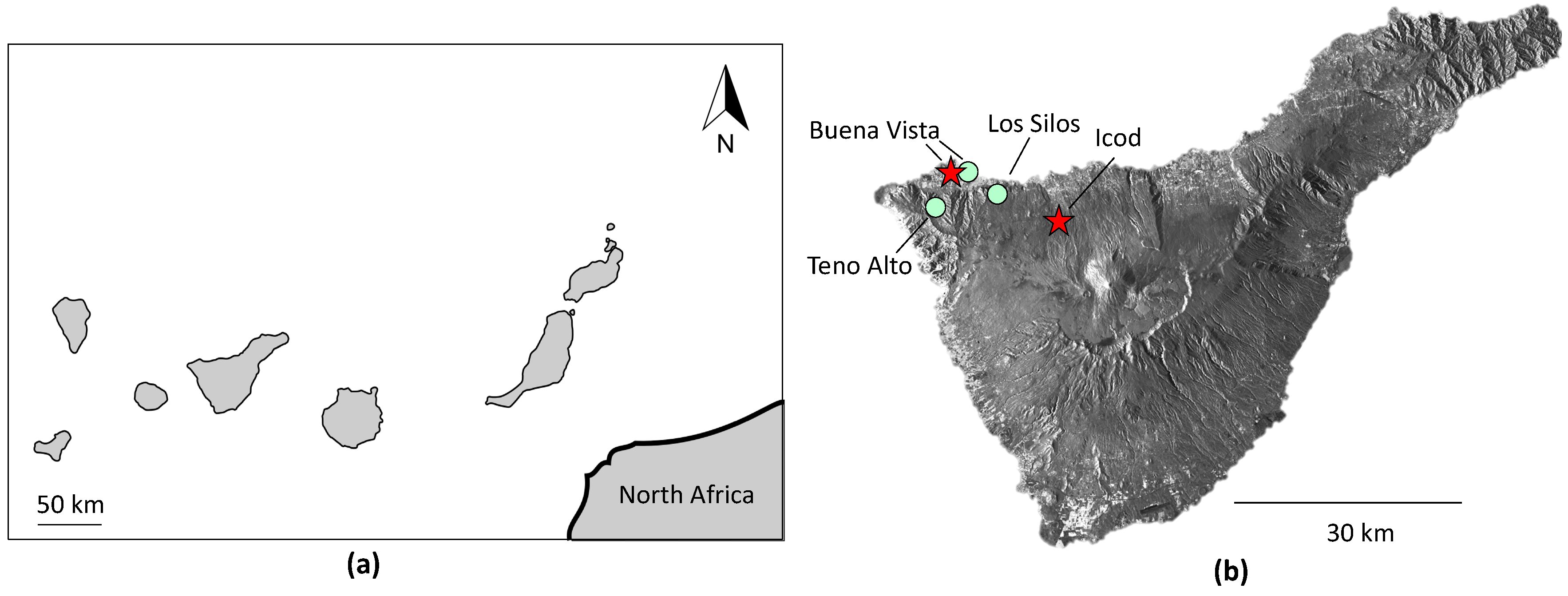
1.2. Regional Setting
2. Materials and Methods
2.1. Site Description and Sample Collection
2.2. Sample Preparation and Analysis
2.3. Data Analysis
3. Results
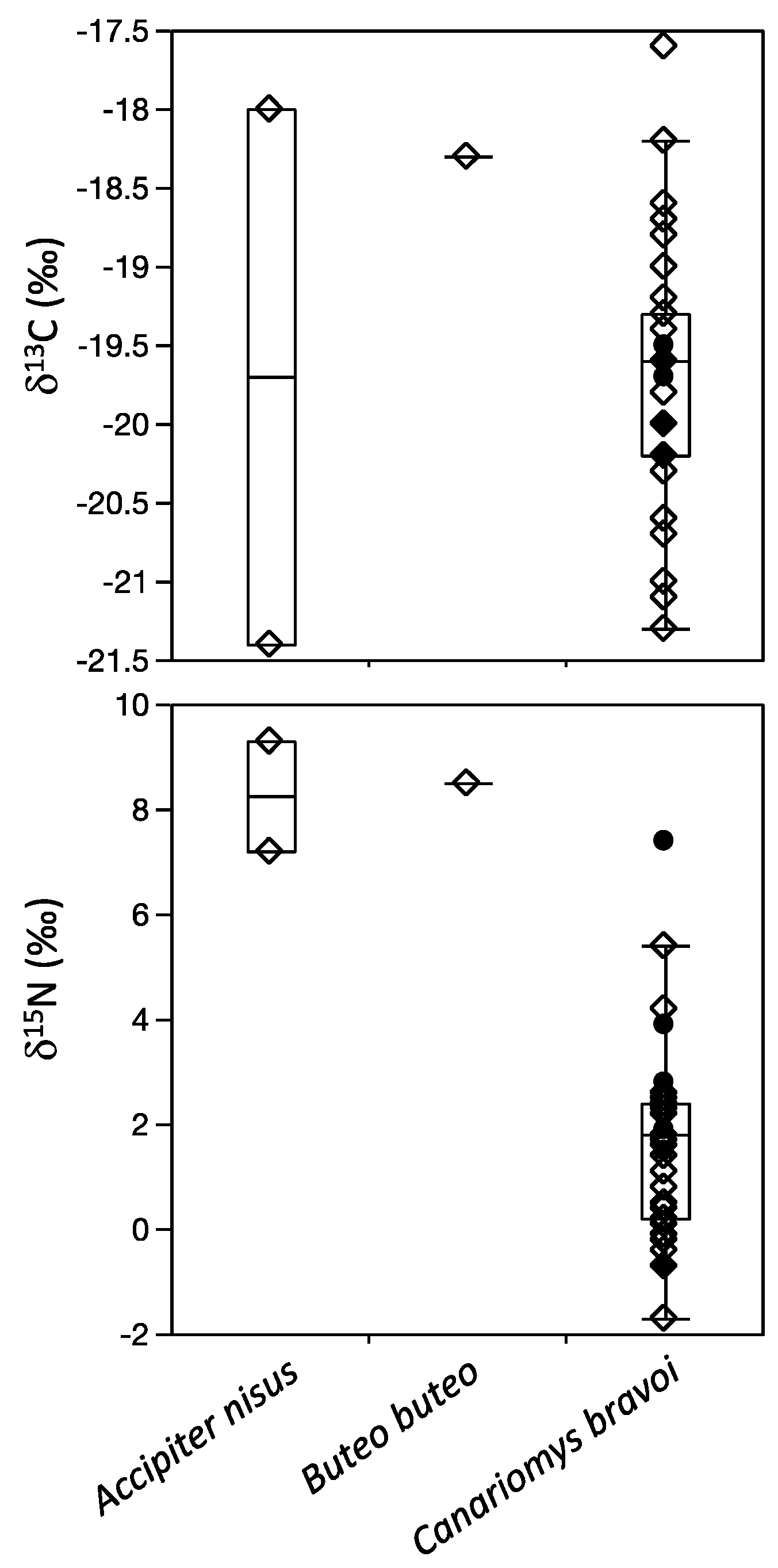
3.1. Icod Region
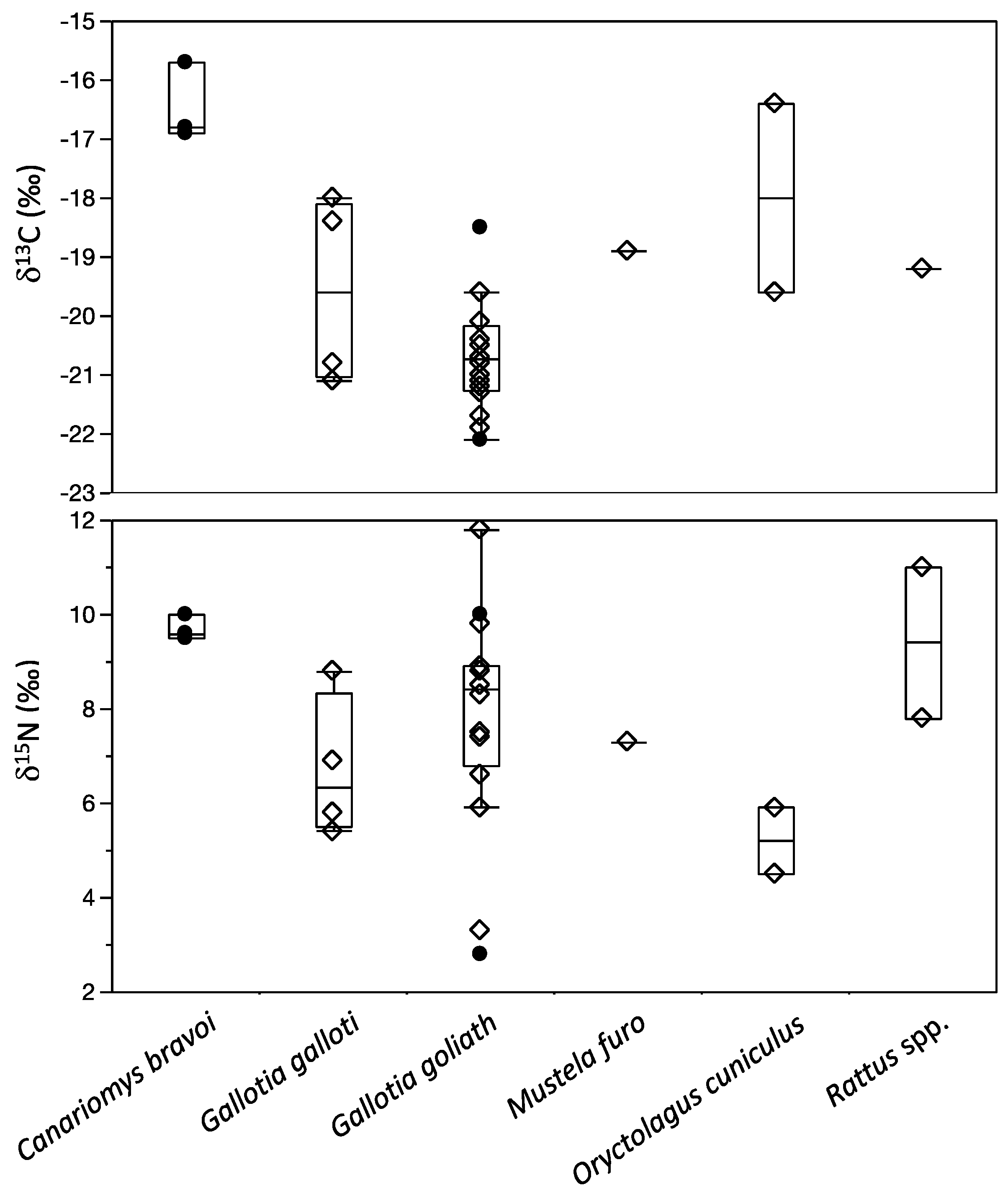
3.2. Buenavista del Norte
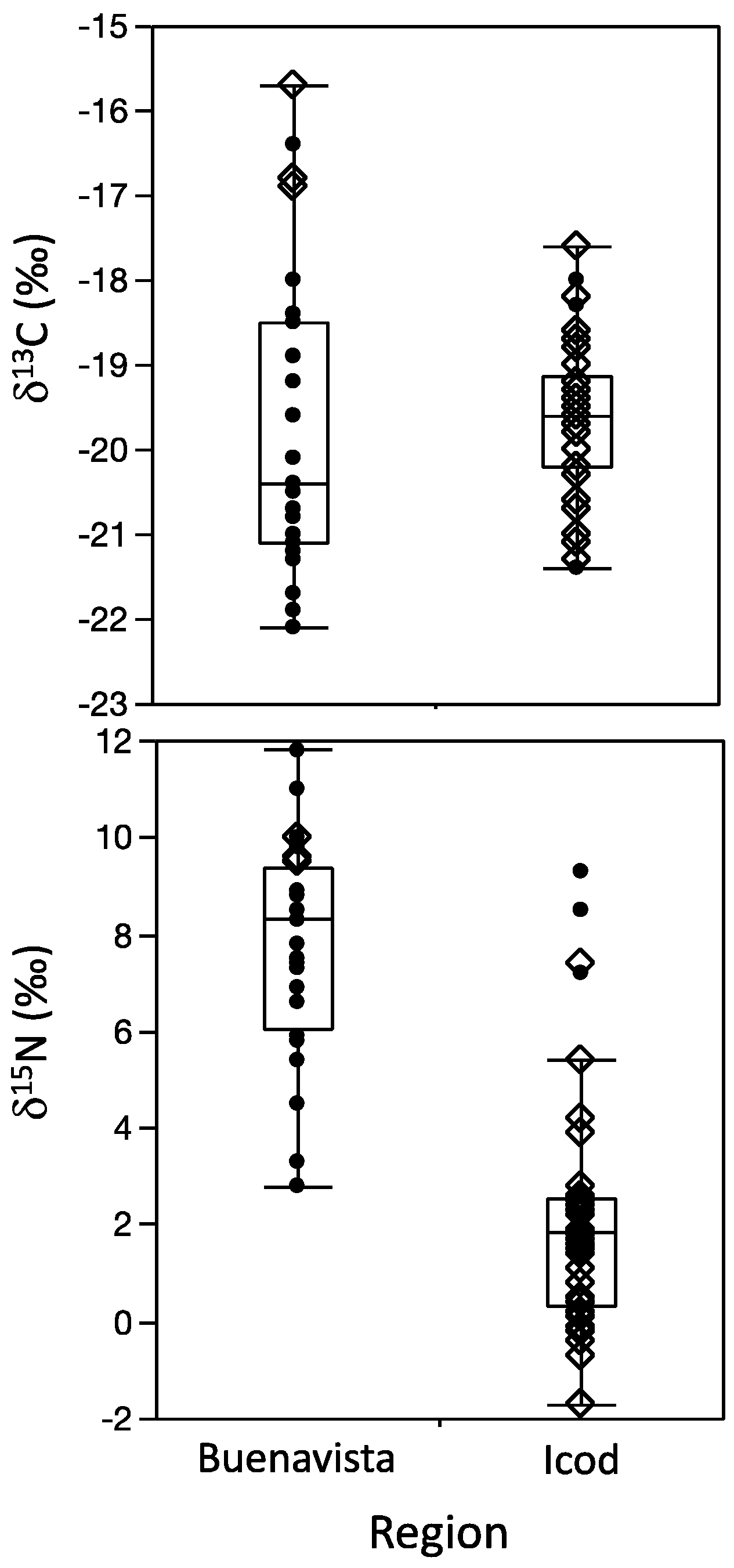
3.3. Regional Comparison
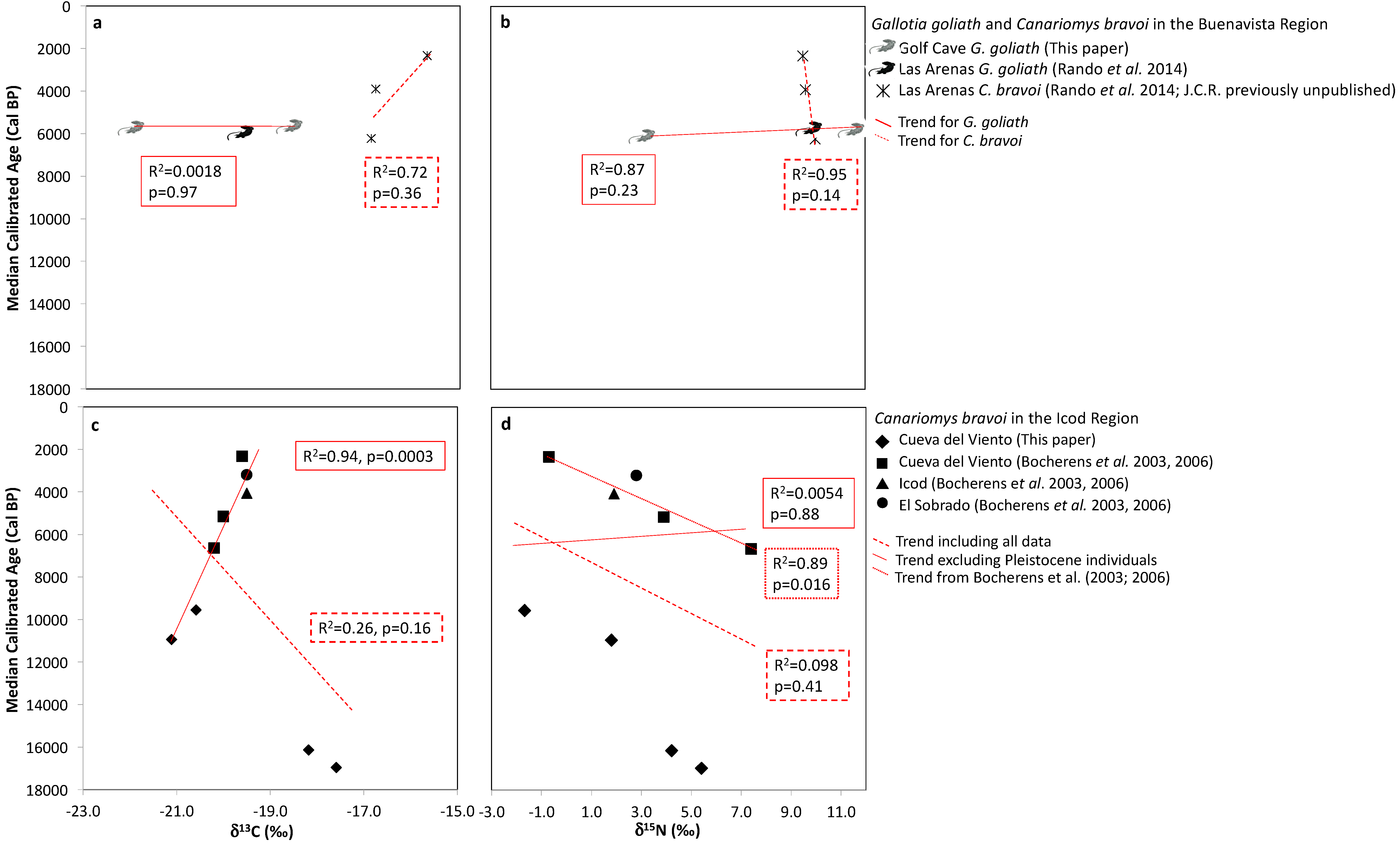
3.4. Temporal Trends
4. Discussion
4.1. Reconstructing Diet and Habitat for Now-Extinct Taxa
4.2. Extinction History and Temporal Isotopic Trends for Canariomys bravoi and Gallotia goliath
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Barnosky, A.D.; Matzke, N.; Tomiya, S.; Wogan, G.O.U.; Swartz, B.; Quental, T.B.; Marshall, C.; McGuire, J.L.; Lindsey, E.L.; Maguire, K.C.; et al. Has the Earth‘s sixth mass extinction already arrived? Nature 2011, 471, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Hooper, D.U.; Adair, E.C.; Cardinale, B.J.; Byrnes, J.E.; Hungate, B.A.; Matulich, K.L.; Gonzalez, A.; Duffy, J.E.; Gamfeldt, L.; O’Connor, M.I. A global synthesis reveals biodiversity loss as a major driver of ecosystem change. Nature 2012, 486, 105–108. [Google Scholar] [CrossRef] [PubMed]
- Dunne, J.A.; Williams, R.J.; Martinez, N.D. Network structure and biodiversity loss in food webs: Robustness increases with connectance. Ecol. Lett. 2002, 5, 558–567. [Google Scholar] [CrossRef]
- Bond, W.J.; Silander, J.A. Springs and wire plants; anacrhonistic defences against Madagascar’s extinct elephant birds. Proc. R. Soc. Lond. 2007, 274, 1985–1992. [Google Scholar] [CrossRef]
- Cerling, T.E.; Harris, J.M.; Leakey, M.G. Browsing and grazing in elephants: The isotope record of modern and fossil proboscideans. Oecologia 1999, 120, 364–374. [Google Scholar] [CrossRef] [PubMed]
- Crowley, B.E.; Godfrey, L.R. Why all those spines? Anachronistic defenses in the Didiereoideae against now extinct lemurs. S. Afr. J. Sci. 2013, 109, 70–76. [Google Scholar] [CrossRef]
- Gill, J.L.; Williams, J.W.; Jackson, S.T.; Donnelly, J.P.; Schellinger, G.C. Climatic and megaherbivory controls on late-glacial vegetation dynamics: A new, high-resolution, multi-proxy record from Silver Lake, Ohio. Quat. Sci. Rev. 2012, 34, 66–80. [Google Scholar] [CrossRef]
- Janzen, D.H.; Martin, P.S. Neotropical anachronisms: The fruits the gomphotheres ate. Science 1982, 215, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Newsome, S.D.; Etnier, M.A.; Gifford-Gonzalez, D.; Phillips, D.L.; van Tuinen, M.; Hadly, E.A.; Costa, D.P.; Kennett, D.J.; Guilderson, T.P.; Koch, P.L. The shifting baseline of northern fur seal ecology in the northeast Pacific Ocean. Proc. Natl. Acad. Sci. USA 2007, 104, 9709–9714. [Google Scholar] [CrossRef] [PubMed]
- Miller, G.H.; Fogel, M.L.; Magee, J.W.; Gagan, M.K.; Clarke, S.J.; Johnson, B.J. Ecosystem collapse in Pleistocene Australia and a human role in megafaunal extinction. Science 2005, 309, 287–290. [Google Scholar] [CrossRef]
- Pimm, S.L.; Jones, H.L.; Diamond, J. On the risk of extinction. Am. Nat. 1988, 132, 757–785. [Google Scholar] [CrossRef]
- Milberg, P.; Tyrberg, T. Naïve birds and noble savages: A review of man-caused prehistoric extinctions of island birds. Ecography 1993, 16, 229–250. [Google Scholar] [CrossRef]
- Blackburn, T.M.; Cassey, P.; Duncan, R.P.; Evans, K.L.; Gaston, K.L. Avian extinction and mammalian introductions on oceanic islands. Science 2004, 305, 1955–1958. [Google Scholar] [CrossRef] [PubMed]
- Burney, D.A.; Flannery, T.F. Fifty millennia of catastrophic extinctions after human contact. Trends Ecol. Evol. 2005, 20, 395–401. [Google Scholar] [CrossRef] [PubMed]
- Gillespie, R.G.; Claridge, E.M.; Roderick, G.K. Biodiversity dynamics in isolated island communities: Interaction between natural and human-mediated processes. Mol. Ecol. 2008, 17, 45–57. [Google Scholar] [CrossRef]
- Crowley, B.E.; Godfrey, L.R.; Bankoff, R.J.; Perry, G.H.; Culleton, B.J.; Kennett, D.J.; Sutherland, M.R.; Samonds, K.E.; Burney, D.A. Island-wide aridity did not trigger recent megafaunal extinctions in Madagascar. Ecography 2017, 40, 901–912. [Google Scholar] [CrossRef]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; da Fonseca, G.A.B.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef]
- Bocherens, H.; Michaux, J.; Garcia Talavera, F.; Van der Plicht, J. Extinction of endemic vertebrates on islands: The case of the giant rat Canariomys bravoi (Mammalia, Rodentia) on Tenerife (Canary Islands, Spain). C. R. Palevol 2006, 5, 885–891. [Google Scholar] [CrossRef]
- Rando, J.C.; Alcover, J.A.; Navarro, J.F.; García-Talavera, F.; Hutterer, R.; Michaux, J. Chronology and causes of the extinction of the lava mouse, Malpaisomys insularis (Rodentia: Muridae) from the Canary Islands. Quat. Res. 2008, 70, 141–148. [Google Scholar] [CrossRef]
- Rando, J.C.; López, M.; Seguí, B. A new species of extinct flightless passerine (Emberizidae: Emberiza) from the Canary Islands. Condor 1999, 101, 1–13. [Google Scholar] [CrossRef]
- Rando, J.C.; Alcover, J.A. On the extinction of the dune shearwater (Puffinus holeae) from the Canary Islands. J. Ornithol. 2010, 151, 365–369. [Google Scholar] [CrossRef]
- Rando, J.C.; Alcover, J.A.; Michaux, J.; Huterrer, R.; Navarro, J.F. Late-Holocene asynchronous extinction of endemic mammals on the eastern Canary Islands. Holocene 2012, 22, 801–808. [Google Scholar] [CrossRef]
- Rando, J.C.; Alcover, J.A.; Galván, B.; Navarro, J.F. Reappraisal of the extinction of Canariomys bravoi, the giant rat from Tenerife (Canary Islands). Quat. Sci. Rev. 2014, 94, 22–27. [Google Scholar] [CrossRef]
- Fernández-Palacios, J.M.; Whittaker, R.J. The Canaries: An important biogeographical meeting place. J. Biogeogr. 2008, 35, 379–387. [Google Scholar] [CrossRef]
- Crusafont-Pairo, M.; Petter, F. Un murine géant fossile des Iles Canaries Canariomys bravoi Gen. nov., sp. nov. (Rongeurs, Muridés). Mamm. Biol. 1965, 28, 607–612. [Google Scholar] [CrossRef]
- López-Martinez, N.; López-Jurado, L.F. Un nuevo murido gigante del Cuaternario de Gran Canaria Canariomys tamarani nov. sp. (Rodentia Mammalia). Doñana Publ. Ocas. 1987, 2, 1–60. [Google Scholar]
- Michaux, J.; Hautier, L.; Hutterer, R.; Lebrun, R.; Guy, F.; García-Talavera, F. Body shape and life style of the extinct rodent Canariomys bravoi (Mammalia, Murinae) from Tenerife, Canary Islands (Spain). C. R. Palevol. 2012, 11, 485–494. [Google Scholar] [CrossRef]
- Lecompte, E.; Aplin, K.; Denys, C.; Catzeflis, F.; Chades, M.; Chevret, P. Phylogeny and biogeography of African Murinae based on mitochondrial and nuclear gene sequences, with a new tribal classification of the subfamily. BMC Evol. Biol. 2008, 8, 199. [Google Scholar] [CrossRef]
- Pàges, M.; Chevret, P.; Gros-Balthazard, M.; Hughes, S.; Alcover, J.A.; Hutterer, R.; Rando, J.C.; Michaux, J.; Haenni, C. Paleogenetic analyses reveal unsuspected phylogenetic affinities between mice and the extinct Malpaisomys insularis, an endemic rodent of the Canaries. PLoS ONE 2012, 7, e31123. [Google Scholar] [CrossRef]
- Firmat, C.; Rodrigues, H.G.; Renaud, S.; Claude, J.; Hutterer, R.; Garcia-Talavara, F.; Michaux, J. Mandible morphology, dental microwear, and diet of the extinct giant rats Canariomys (Rodentia: Murinae) of the Canary Islands (Spain). Biol. J. Linn. Soc. 2010, 101, 28–40. [Google Scholar] [CrossRef]
- Maca-Meyer, N.; Carranza, S.; Rando, J.C.; Arnold, E.N.; Cabrera, V.M. Status and relationships of the extinct giant Canary Island lizard Gallotia goliath (Reptilia: Lacertidae), assessed using ancient mtDNA from its mummified remains. Biol. J. Linn. Soc. 2003, 80, 659–670. [Google Scholar] [CrossRef]
- Barahona, F.; Evans, S.E.; Mateo, J.A.; García-Márquez, M.; López-Jurado, L.J. Endemism, gigantism and extinction in island lizards: The genus Gallotia on the Canary Islands. J. Zool. Lond. 2000, 250, 373–388. [Google Scholar] [CrossRef]
- Van Damme, R. Evolution of herbivory in lacertid lizards: Effects of insularity and body size. J. Herpetol. 1999, 33, 663–674. [Google Scholar] [CrossRef]
- López-Jurado, L.F.; Mateo, J.A. Origin, colonization, adaptive radiation, intrainsular evolution and species substitution processes in the fossil and living lizards of the Canary Islands. In Scienta Herpetologica; Llorente, G.A., Montori, A., Santos, X., Carretero, M.A., Eds.; Asociación Herpetológica Española: Barcelona, Spain, 1995; pp. 81–91. [Google Scholar]
- Mateo, J.A.; Lopez-Jurado, L.F. Study of dentition in lizards from Gran Canaria Island (Canary Islands) and its ecological and evolutionary significance. Biol. J. Linn. Soc. 1992, 46, 39–48. [Google Scholar] [CrossRef]
- Naranjo, J.J.; Nogales, M.; Quilis, V. Sobre la presencia de Gallotia stehlini en la isla de Fuerte- ventura (Canarias) y datos preliminares de su alimentación. Rev. Española De Herpetol. 1991, 6, 45–48. [Google Scholar]
- Valido, A.; Nogales, M. Frugivory and seed dispersal by the lizard Gallotia galloti (Lacertidae) in a xeric habitat of the Canary Islands. Oikos 1994, 70, 403–411. [Google Scholar] [CrossRef]
- Rodríguez, A.; Nogales, M.; Rumeu, B.; Rodríguez, B. Temporal and spatial variation in the diet of the endemic lizard Gallotia galloti in an insular Mediterranean scrubland. J. Herpetol. 2008, 42, 213–222. [Google Scholar] [CrossRef]
- Valido, A.; Nogales, M.; Medina, F.M. Fleshy fruits in the diet of Canarian lizards Gallotia galloti (Lacertidae) in a xeric habitat of the island of Tenerife. J. Herpetol. 2003, 37, 741–747. [Google Scholar] [CrossRef]
- Valido, A.; Nogales, M. Digestive ecology of two omnivorous Canarian lizard species (Gallotia, Lacertidae). Amphib. -Reptil. 2003, 24, 331–344. [Google Scholar] [CrossRef]
- Nogales, M.; Medina, F.M. Trophic ecology of feral cats (Felis silvestris f. catus) in the main environments of an oceanic archipelago (Canary Islands): An updated approach. Mamm. Biol. 2009, 74, 169–181. [Google Scholar] [CrossRef]
- Nogales, M.; Vidal, E.; Medina, F.M.; Bonnaud, E.; Tershy, B.R.; Campbell, K.J.; Zavaleta, E.S. Feral cats and biodiversity conservation: The urgent prioritization of island management. BioScience 2013, 63, 804–810. [Google Scholar] [CrossRef]
- Tieszen, L.L.; Matzner, S.; Buseman, S.K. Dietary reconstruction based on stable isotopes (13C, 15N) of the Guanche, pre-hispanic Tenerife, Canary Islands. In Proceedings of the I World Congress on Mummy Studies, Tenerife, Spain, 1992; pp. 41–57. [Google Scholar]
- Bocherens, H.; Michaux, J.; Billiou, D.; Castanet, J.; Garcia-Talavera, F. Contribution of collagen stable isotope biogeochemistry to the paleobiology of extinct endemic vertebrates from Tenerife (Canary Islands, Spain). Isot. Environ. Health Stud. 2003, 39, 197–210. [Google Scholar] [CrossRef] [PubMed]
- Arnay-de-la-Rosa, M.; González-Reimers, E.; Yanes, Y.; Velasco-Vázquez, J.; Romanek, C.S.; Noakes, J.E. Paleodietary analysis of the prehistoric population of the Canary Islands inferred from stable isotopes (carbon, nitrogen and hydrogen) in bone collagen. J. Archaeol. Sci. 2010, 37, 1490–1501. [Google Scholar] [CrossRef]
- Arnay-de-la-Rosa, M.; González-Reimers, E.; Yanes, Y.; Romanek, C.S.; Noakes, J.E.; Galindo-Martín, L. Paleonutritional and paleodietary survey on prehistoric humans from Las Cañadas del Teide (Tenerife, Canary Islands) based on chemical and histological analysis of bone. J. Archaeol. Sci. 2011, 38, 884–895. [Google Scholar] [CrossRef]
- O’Leary, M.H. Carbon isotopes in photosynthesis. BioScience 1988, 38, 328–336. [Google Scholar] [CrossRef]
- Kohn, M.J. Carbon isotope compositions of terrestrial C3 plants as indicators of (paleo)ecology and (paleo)climate. Proc. Natl. Acad. Sci. USA 2010, 107, 19691–19695. [Google Scholar] [CrossRef] [PubMed]
- Koch, P.L. Isotopic reconstruction of past continental environments. Annu. Rev. Earth Planet. Sci. 1998, 26, 573–613. [Google Scholar] [CrossRef]
- Fox-Dobbs, K.; Bump, J.K.; Peterson, R.O.; Fox, D.L.; Koch, P.L. Carnivore-specific stable isotope variables and variation in the foraging ecology of modern and ancient wolf populations: Case studies from Isle Royale, Minnesota, and La Brea. Can. J. Zool. 2007, 85, 458–471. [Google Scholar] [CrossRef]
- Cormie, A.B.; Schwarcz, H.P. Stable isotopes of nitrogen and carbon of North American white-tailed deer and implications for paleodietary and other food web studies. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1994, 107, 227–241. [Google Scholar] [CrossRef]
- Crowley, B.E.; Thorén, S.; Rasoazanabary, E.; Vogel, E.R.; Barrett, M.A.; Zohdy, S.; Blanco, M.B.; McGoogan, K.C.; Arrigo-Nelson, S.J.; Irwin, M.T.; et al. Explaining geographical variation in the isotope composition of mouse lemurs (Microcebus). J. Biogeogr. 2011, 38, 2106–2121. [Google Scholar] [CrossRef]
- Hartman, G. Are elevated δ15N values in herbivores in hot and arid environments caused by diet or animal physiology? Funct. Ecol. 2011, 25, 122–131. [Google Scholar] [CrossRef]
- van der Merwe, N.J.; Medina, E. The canopy effect, carbon isotope ratios and foodwebs in Amazonia. J. Archaeol. Sci. 1991, 18, 249–259. [Google Scholar] [CrossRef]
- Broadmeadow, M.S.J.; Griffiths, H.; Maxwell, C.; Borland, A.M. The carbon isotope ratio of plant organic material reflects temporal and spatial variations in CO2 within the tropical forest formations in Trinidad. Oecologia 1992, 89, 435–441. [Google Scholar] [CrossRef] [PubMed]
- Cerling, T.E.; Hart, J.A.; Hart, T.B. Isotope ecology in the Ituri forest. Oecologia 2004, 138, 5–12. [Google Scholar] [CrossRef]
- Carlson, B.A.; Crowley, B.E. Variation in carbon isotope values among chimpanzee foods at Ngogo, Kibale National Park and Bwindi Impenetrable National Park, Uganda. Am. J. Primatol. 2016, 78, 1031–1040. [Google Scholar] [CrossRef]
- Britton, K.; Müldner, G.; Bell, M. Stable isotope evidence for salt-marsh grazing in the Bronze Age Severn Estuary, UK: Implications for palaeodietary analysis at coastal sites. J. Archaeol. Sci. 2008, 35, 2111–2118. [Google Scholar] [CrossRef]
- Reid, R.E.; Gifford-Gonzalez, D.; Koch, P.L. Coyote (Canis latrans) use of marine resources in coastal California: A new behavior relative to their recent ancestors. Holocene 2018, 28, 1781–1790. [Google Scholar] [CrossRef]
- Fox-Dobbs, K.; Stidham, T.A.; Bowen, G.J.; Emslie, S.D.; Koch, P.L. Dietary controls on extinction versus survial among avian megafauna in the late Pleistocene. Geology 2006, 34, 685–688. [Google Scholar] [CrossRef]
- Sparks, J.M.; Crowley, B.E. Where did people forage in prehistoric Trinidad? Testing the utility of a multi-isotope approach for tracking the origins of terrestrial prey. J. Archaeol. Sci. 2018, 19, 968–978. [Google Scholar] [CrossRef]
- Fernández-Palacios, J.M. Climatic responses of plant species on Tenerife, the Canary Islands. J. Veg. Sci. 1992, 3, 595–602. [Google Scholar] [CrossRef]
- Balasse, M. Potential biases in sampling design and interpretation of intra-tooth isotope analysis. Int. J. Osteoarchaeol. 2003, 13, 3–10. [Google Scholar] [CrossRef]
- Fernández-Palacios, J.M.; María, J.; de Nicolás, J.P. Altitudinal pattern of vegetation variation on Tenerife. J. Veg. Sci. 1995, 6, 183–190. [Google Scholar] [CrossRef]
- del-Arco, M.; Pérez-de-Paz, P.L.; Acebes, J.R.; González-Mancebo, J.M.; Reyes-Betancort, J.A.; Bermejo, J.A.; de-Armas, S.; González-González, R. Bioclimatology and climatophilous vegetation of Tenerife (Canary Islands). Ann. Bot. Fenn. 2006, 43, 167–192. [Google Scholar] [CrossRef]
- Yanes, Y.; Yapp, C.J.; Ibáñez, M.; Alonso, M.R.; De la Nuez, J.; Quesada, M.L.; Castillo, C.; Delgado, A. Pleistocene-Holocene environmental change in the Canary Archipelago as inferred from stable isotopes of land snail shells. Quat. Res. 2011, 65, 658–669. [Google Scholar] [CrossRef]
- Gasse, F. Hydrological changes in the African tropics since the Last Glacial Maximum. Quat. Sci. Rev. 2000, 19, 189–211. [Google Scholar] [CrossRef]
- deMenocal, P.; Ortiz, J.; Guilderson, T.; Adkins, J.; Sarnthein, M.; Baker, L.; Yarusinsky, M. Abrupt onset and termination of the African Humid Period: Rapid climate responses to gradual insolation forcing. Quat. Sci. Rev. 2000, 19, 347–361. [Google Scholar] [CrossRef]
- Ramos-Román, M.J.; Jiménez-Moreno, G.; Camuera, J.; García-Alix, A.; Anderson, R.S.; Jiménez-Espejo, F.J.; Sachse, D.; Toney, J.L.; Carrión, J.S.; Webster, C.; et al. Millennial-scale cyclical environment and climate variability during the Holocene in the western Mediterranean region deduced from a new multi-proxy analysis from the Padul record (Sierra Nevada, Spain). Glob. Planet. Chang. 2018, 168, 35–53. [Google Scholar] [CrossRef]
- Alcover, J.A.; Rando, J.C.; García-Talavera, F.; Hutterer, R.; Michaux, J.; Trias, M.; Navarro, J.F. A reappraisal of the stratigraphy of Cueva del Llano (Fuerteventura) and the chronology of the house mouse (Mus musculus) introduction into the Canary Islands. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2009, 277, 184–190. [Google Scholar] [CrossRef]
- Rando, J.C.; Alcover, J.A. Evidence for a second western Palaearctic seabird extinction during the last Millennium: The lava shearwater Puffinus olsoni. Ibis 2008, 150, 188–192. [Google Scholar] [CrossRef]
- de Nascimento, L.; Willis, K.J.; Fernández-Palacios, J.M.; Criado, C.; Whittaker, R.J. The long-term ecology of the lost forests of La Laguna, Tenerife (Canary Islands). J. Biogeogr. 2009, 36, 499–514. [Google Scholar] [CrossRef]
- Morales, J.; Rodríguez, A.; Alberto, V.; Machado, C.; Criado, C.; Rando, J.C. The impact of human activities on the natural environment of the Canary Islands (Spain) during the pre-Hispanic stage (3rd–2nd Century BC to 15th Century AD): An overview. Environ. Archaeol. 2009, 14, 27–36. [Google Scholar] [CrossRef]
- Dickson, J.H.; Rodriguez, J.C.; Machado, A. Invading plants at high altitudes on Tenerife especially in the Teide National Park. Bot. J. Linn. Soc. 1987, 95, 155–179. [Google Scholar] [CrossRef]
- Francisco-Ortega, J.; Santos-Guerra, A.; Kim, S.-C.; Crawford, D.J. Plant genetic diversity in the Canary Islands: A conservation perspective. Am. J. Bot. 2000, 87, 900–919. [Google Scholar] [CrossRef]
- Nogales, N.; Rodríguez-Luengo, J.L.; Marrero, P. Ecological effects and distribution of invasive non-native mammals on the Canary Islands. Mamm. Rev. 2006, 36, 49–65. [Google Scholar] [CrossRef]
- Carrete, M.; Serrano, D.; Illera, J.C.; López, G.; Vögeli, M. Goats, birds, and emergent diseases: Apparent and hidden effects of exotic species in an island environment. Ecol. Appl. 2009, 19, 840–853. [Google Scholar] [CrossRef] [PubMed]
- Wikimedia Commons. Available online: https://commons.wikimedia.org/wiki/File:Blank_Map-Canary_Islands.svg (accessed on 20 December 2018).
- Airbus Defence and Space. Available online: http://www.intelligence-airbusds.com/en/67-base-maps (accessed on 20 December 2018).
- Gangoso, L.; Donázar, J.A.; Scholz, S.; Palacios, C.J.; Hiraldo, F. Contradiction in conservation of island ecosystems: Plants, introduced herbivores and avian scavengers in the Canary Islands. Biodivers. Conserv. 2006, 15, 2231–2248. [Google Scholar] [CrossRef]
- Palomar, C.J. El ratonero común (Buteo buteo insularum) en Fuerteventura, islas Canarias (Aves, Accipitridae). Vieraea 2005, 33, 1–10. [Google Scholar]
- Post, P.; Götmark, F. Prey selection by sparrowhawks, Accipiter nisus: Relative predation risk for breeding passerine birds in relation to their size, ecology and behaviour. Philos. Trans. Biol. Sci. 1996, 351, 1559–1577. [Google Scholar]
- Delgado, G.; Martin, A.; Quilis, V.; Emmerson, K. Diet of the sparrowhawk (Accipiter nisus) on the island of Tenerife. Doñana. Acta Vertebr. 1988, 15, 193–199. [Google Scholar]
- Medina, F.M.; Martín, A. A new invasive species in the Canary Islands: A naturalized population of ferrets Mustela furo in La Palma Biosphere Reserve. Oryx 2009, 44, 41–44. [Google Scholar] [CrossRef]
- Smith, G.P.; Ragg, J.R.; Moller, H.; Waldrup, K.A. Diet of ferrets (Mustela furo) from pastoral habitats in Otago and Southland, New Zealand. N. Z. J. Zool. 1995, 22, 363–369. [Google Scholar] [CrossRef]
- Ragg, J.R. Intraspecific and seasonal differences in the diet of feral ferrets (Mustela furo) in a pastoral habitat, East Otago, New Zealand. N. Z. J. Ecol. 1998, 22, 113–119. [Google Scholar]
- Bodey, T.W.; Bearhop, S.; McDonald, R.A. The diet of an invasive nonnative predator, the feral ferret Mustela furo, and implications for the conservation of ground-nesting birds. Eur. J. Wildl. Res. 2011, 57, 107–177. [Google Scholar] [CrossRef]
- Cole, F.R.; Loope, L.L.; Medeiros, A.C.; Howe, C.E.; Anderson, L.J. Food habits of introduced rodents in high-elevation shrubland of Haleakala National Park, Maui, Hawaii. Pac. Sci. 2000, 54, 313–329. [Google Scholar]
- Towns, D.R.; Atkinson, I.A.E.; Dagherty, C.H. Have the harmful effects of introduced rats on islands been exaggerated? Biol. Invasions 2006, 8, 863–891. [Google Scholar] [CrossRef]
- Shiels, A.B.; Flores, C.A.; Khamsing, A.; Krushelnycky, P.D.; Mosher, S.M.; Drake, D.R. Dietary niche differentiation among three species of invasive rodents (Rattus rattus, R. exulans, Mus musculus). Biol. Invasions 2013, 15, 1037–1048. [Google Scholar] [CrossRef]
- Chapuis, J.L. Comparison of the diets of two sympatric lagomorphs, Lepus europaeus (Pallas) and Oryctolagus cuniculus (L.) in an agro-ecosystem of the Ile-de-France. Z. Für Säugetierkunde 1990, 55, 176–185. [Google Scholar]
- Martins, H.; Milne, J.A.; Rego, F. Seasonal and spatial variation in the diet of the wild rabbit (Oryctolagus cuniculus L.) in Portugal. J. Zool. 2002, 258, 395–404. [Google Scholar] [CrossRef]
- France, C.M.; Giaccai, J.A.; Cano, N. The effects of PVAc treatment and organic solvent removal on δ13C, δ15N, and δ18O values of collagen and hydroxyapatite in a modern bone. J. Archaeol. Sci. 2011, 38, 3387–3393. [Google Scholar] [CrossRef]
- Ambrose, S.H. Preparation and characterization of bone and tooth collagen for isotopic analysis. J. Archaeol. Sci. 1990, 17, 431–451. [Google Scholar] [CrossRef]
- Skrypzpek, G. Normalization procedures and reference material selection in stable HCNOS isotope analyses: An overview. Anal. Bioanal. Chem. 2013, 405, 2815–2823. [Google Scholar] [CrossRef] [PubMed]
- Stuiver, M.; Reimer, P.J.; Reimer, R.W. CALIB 7.1. Available online: http://calib.org (accessed on 2 January 2019).
- Reimer, P.; Bard, E.; Bayliss, A.; Beck, J.W.; Blackwell, P.G.; Bronk Ramsey, C.; Buck, C.E.; Cheng, H.; Edwards, R.L.; Friedrich, M.; et al. IntCal13 AND Marine13 radiocarbon age calibration curves 0–50,000 years cal BP. Radiocarbon 2013, 55, 1869–1887. [Google Scholar] [CrossRef]
- Mosher, S.G.; Crowley, B.E.; Diefendorf, A.D.; Brone, R.; Yanes, Y. Characterizing spatial variability in foliar carbon and nitrogen isotopes on Tenerife, Canary Islands. Oecologia, in preparation.
- Francey, R.J.; Allison, C.E.; Etheridge, D.M.; Trudinger, C.M.; Enting, I.G.; Leuenberger, M.; Langenfelds, R.L.; Michel, E.; Steele, L.P. A 1000-year high precision record of δ13C in atmospheric CO2. Tellus B 1999, 51, 170–193. [Google Scholar] [CrossRef]
- Leuenberger, M.; Siegenthaler, U.; Langway, C. Carbon isotope composition of atmospheric CO2 during the last ice age from an Antarctic core. Nature 1992, 357, 488–490. [Google Scholar] [CrossRef]
- Elsig, J.; Schmitt, J.; Leuenberger, D.; Schneider, R.; Eyer, M.; Leuenberger, M.; Joos, F.; Fischer, H.; Stocker, T.F. Stable isotope constraints on Holocene carbon cycle changes from an Antarctic ice core. Nature 2009, 461, 507–510. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, J.; Schneider, R.; Elsig, J.; Leuenberger, D.; Lourantou, A.; Chappellaz, J.; Köhler, P.; Joos, F.; Stocker, T.F.; Leuenberger, M.; et al. Carbon isotope constraints on the deglacial CO2 rise from ice cores. Science 2012, 336, 711–714. [Google Scholar] [CrossRef]
- Indermühle, A.; Stocker, T.F.; Joos, F.; Fischer, H.; Smith, H.J.; Wahlen, M.; Deck, B.; Mastroianni, D.; Tschumi, J.; Blunier, T.; et al. Holocene carbon-cycle dynamics based on CO2 trapped in ice at Taylor Dome, Antarctica. Nature 1999, 398, 121–126. [Google Scholar] [CrossRef]
- Strauss, D.; Sadler, P.M. Classical confidence intervals and Bayesian probability estimates for ends of local taxon ranges. Math. Geol. 1989, 21, 411–427. [Google Scholar] [CrossRef]
- McFarlane, D.A. A comparison of methods for the probabilistic determination of vertebrate extinction chronologies. In Extinction in Near Time; MacPhee, R.D.E., Ed.; Kluwer/Plenum: New York, NY, USA, 1999; pp. 95–103. [Google Scholar]
- Marshall, C.R. Estimation of taxonomic ranges from the fossil record. In Analytical PaleoBiology; Gilinksy, N.L., Signor, P.W., Eds.; University of Tennessee: Knoxville, TN, USA, 1991; pp. 19–38. [Google Scholar]
- Crowley, B.E. A refined chronology of prehistoric Madagascar and the demise of the megafauna. Quat. Sci. Rev. 2010, 29, 2591–2603. [Google Scholar] [CrossRef]
- Michaux, J.; López-Martínez, N.; Hernández-Pacheco, J.J. A 14C dating of Canariomys bravoi (Mammalia Rodentia), the extinct giant rat from Tenerife (Canary Islands, Spain), and the recent history of endemic mammals in the archipelago. Vie Milieu 1996, 46, 261–266. [Google Scholar]
- Bearhop, S.; Adams, C.E.; Waldron, S.; Fuller, R.A.; MacLeod, H. Determining trophic niche width: A novel approach using stable isotope analysis. J. Anim. Ecol. 2004, 73, 1007–1012. [Google Scholar] [CrossRef]
- Moller, H.; Alterio, N. Home range and spatial organisation of stoats (Mustela erminea), ferrets (Mustela furo) and feral house cats (Felis catus) on coastal grasslands, Otago Peninsula, New Zealand: Implications for yellow-eyed penguin (Megadyptes antipodes) conservation. N. Z. J. Zool. 1999, 26, 165–174. [Google Scholar] [CrossRef]
- Yanes, Y.; Romanek, C.S.; Delgado, A.; Brant, H.A.; Noakes, J.E.; Alonso, M.R.; Ibáñez, M. Oxygen and carbon stable isotopes of modern land snail shells as environmental indicators from a low-latitude oceanic island. Geochim. Cosmochim. Acta 2009, 73, 4077–4099. [Google Scholar] [CrossRef]
- Codron, J.; Codron, D.; Lee-Thorp, J.A.; Sponheimer, M.; Bond, W.J.; de Ruiter, D.; Grant, R. Taxonomic, anatomical, and spatio-temporal variations in the stable carbon and nitrogen isotopic compositions of plants from an African savanna. J. Archaeol. Sci. 2005, 32, 1757–1772. [Google Scholar] [CrossRef]
- Cernusak, L.A.; Tcherkez, G.; Keitel, C.; Cornwell, W.K.; Santiago, L.S.; Knohl, A.; Barbour, M.M.; Williams, D.G.; ReichI, P.B.; Ellsworth, D.S.; et al. Why are non-photosynthetic tissues generally 13C enriched compared with leaves in C3 plants? Review and synthesis of current hypotheses Funct. Plant. Biol. 2009, 36, 199–213. [Google Scholar] [CrossRef]
- Blumenthal, S.A.; Rothman, J.M.; Chiritz, K.L.; Cerling, T.E. Stable isotopic variation in tropical forest plants for applications in primatology. Am. J. Primatol. 2016, 78, 1041–1054. [Google Scholar] [CrossRef]
- Marzol-Jaén, M.V.; Sànchez-Megía, J.; García-Santos, G. Effects of fog on climate conditions at a subtropical montane cloud forest site in northern Tenerife (Canary Islands, Spain). In Tropical Montane Cloud Forests: Science for Conservation and Management; Bruijnzeel, L.A., Scatena, F.N., Hamilton, L.S., Eds.; Cambridge University Press: Cambridge, UK, 2010; pp. 359–364. [Google Scholar]
- Parker, W.G.; Yanes, Y.; Surge, D.; Mesa-Hernandez, E. High-resolution paleoclimate reconstruction over the last two millennia in the eastern subtropical Atlantic Ocean. Gsa Abstr. Programs 2018, 50, 86. [Google Scholar] [CrossRef]
- Gieger, T.; Leuschner, C. Altitudinal change in needle water relations of Pinus canariensis and possible evidence of a drought-induced alpine timberline on Mt. Teide, Tenerife. Flora 2004, 199, 100–109. [Google Scholar] [CrossRef]
- Anderson, R.L.; Byrne, R.; Dawson, T. Stable isotope evidence for a foggy climate on Santa Cruz Island, California at ~16,600 cal. yr. B.P. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2008, 262, 176–181. [Google Scholar] [CrossRef]
- Johnstone, J.A.; Dawson, T.E. Climatic context and ecological implications of summer fog decline in the coast redwood region. Proc. Natl. Acad. Sci. USA 2010, 107, 4533–4538. [Google Scholar] [CrossRef]
- del-Arco, M.C.; del-Arco, M.M.; Atiénzar, E.; Atoche, P.; Martín, M.; Rodríguez, C.; Rosario, C. Dataciones absolutas en la prehistoria de Tenerife. In Homenaje a Celso Martín Guzmán; Millares, A., Carbrera, M.L., Peña, P.A., Eds.; Universidad De Las Palmas De Gran Canaria: Las Palmas de Gran Canaria, Spain, 1997; pp. 65–77. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Municipality | Site | Genus and Species | Common Name | δ13C (‰) | δ15N (‰) | Source 1 | |||
|---|---|---|---|---|---|---|---|---|---|
| N | Mean ± 1σ | Median | Mean ± 1σ | Median | |||||
| Icod | |||||||||
| Cueva del Viento | |||||||||
| Accipiter nisus | Macaronesian sparrowhawk | 2 | −19.7 ± 2.4 | −19.7 | 8.3 ± 1.5 | 8.3 | 1 | ||
| Buteo buteo | Common buzzard | 1 | -18.3 | −18.3 | 8.5 | 8.5 | 1 | ||
| Canariomys bravoi | Extinct giant rat | 31 | −19.7 ± 0.8 | −19.6 | 1.6 ± 1.8 | 1.8 | 1, 2 | ||
| Icod | Canariomys bravoi | Extinct giant rat | 1 | −19.5 | −19.5 | 2.8 | 2.8 | 2 | |
| El Sobrado | Canariomys bravoi | Extinct giant rat | 3 | −19.6 ± 0.1 | −19.7 | 1.8 ± 1.9 | 1.7 | 2 | |
| Buenavista del Norte | |||||||||
| Golf Cave | Gallotia goliath | Extinct giant lizard | 14 | -20.8 ± 0.7 | −20.8 | 8.0 ± 2.0 | 8.4 | 1 | |
| Golf Cave | Gallotia galloti | Tenerife lizard | 4 | −19.6 ± 1.6 | −19.6 | 6.7 ± 1.5 | 6.4 | 1 | |
| Golf Cave | Oryctolagus cuniculus | Rabbit | 2 | -18.0 ± 2.3 | −18.0 | 5.2 ± 1.0 | 5.2 | 1 | |
| Golf Cave | Rattus sp. | Rat | 2 | −19.2 2 | −19.2 2 | 9.4 ± 2.3 | 9.4 | ||
| Golf Cave | Mustela furo | Ferret | 1 | −18.9 | −18.9 | 7.3 | 7.3 | 1 | |
| Las Arenas 3 | Canariomys bravoi | Extinct giant rat | 3 | −16.5 ± 0.7 | −16.8 | 9.7 ± 0.3 | 9.6 | 3, 4 | |
| Las Arenas 3 | Gallotia goliath | Extinct giant lizard | 2 | −20.3 ± 2.5 | −20.3 | 6.4 ± 5.1 | 6.4 | 2–4 |
| Taxon | Locality | Lab ID 1 | 14C years BP | 2σ Calibrated Date (Cal BP) | Mean ±1σ Calibrated Date (Cal BP) | Median Calibrated Date (Cal BP) | Source 2 |
|---|---|---|---|---|---|---|---|
| Canariomys bravoi | |||||||
| Cueva del Viento | CAMS 167035 | 13,980 ± 130 | 16,525–17,395 | 16,960 ± 435 | 16,965 | 1 | |
| Cueva del Viento | UCI 184465 | 13,410 ± 70 | 15,885–16,355 | 16,120 ± 235 | 16,135 | 1 | |
| Cueva del Viento | OxA-54502 | 12,230 ± 140 | 13,755–14,820 | 14,290 ± 535 | 14,200 | 2 | |
| Cueva del Viento | CAMS 167036 | 9590 ± 70 | 10,720–11,170 | 10,945 ± 225 | 10,940 | 1 | |
| Cueva del Viento | UCI 184466 | 8595 ± 35 | 9500–9630 | 9565 ± 65 | 9545 | 1 | |
| Cueva del Viento | GrA 22661 | 5840 ± 50 | 6500–6750 | 6625 ± 125 | 6655 | 3 | |
| Cueva del Viento | GrA 22658 | 4515 ± 45 | 5040–5315 | 5180 ± 140 | 5160 | 3 | |
| Cueva del Viento | GrA 22680 | 2305 ± 40 | 2155–2380 | 2270 ± 113 | 2330 | 3 | |
| Icod | GrA 22656 | 3010 ± 45 | 3065–3345 | 3205 ± 140 | 3200 | 3 | |
| El Sobrado | GrA 22657 | 3720 ± 45 | 3960–4160 | 4060 ± 100 | 4065 | 3 | |
| Arenas 3 | KIA 40849 | 5400 ± 30 | 6180–6285 | 6235 ± 55 | 6230 | 4 | |
| Arenas 1 | KIA 40846 | 3605 ± 50 | 3825–4085 | 3955 ± 130 | 3915 | 4 | |
| Arenas 3 | KIA 47428 | 2315 ± 55 | 2155–2490 | 2325 ± 170 | 2335 | 4 | |
| La Fortaleza | WK-28571 | 2304 ± 32 | 2180–2360 | 2270 ± 90 | 2335 | 4 | |
| Bco. Moraditas | KIA-36264 | 2275 ± 25 | 2180–2350 | 2265 ± 85 | 2315 | 4 | |
| Upper 95% CI | -1180 | ||||||
| Upper 50% CI | 1570 | ||||||
| Gallotia goliath | |||||||
| Golf Cave | CAMS 166991 | 5265 ± 35 | 5935–6180 | 6060 ± 130 | 6050 | 1 | |
| Golf Cave | CAMS 166990 | 4975 ± 40 | 5605–5880 | 5745 ± 140 | 5700 | 1 | |
| Arenas 3 | KIA 47429 | 4895 ± 40 | 5585–5715 | 5650 ± 65 | 5630 | 4 | |
| Upper 95% CI | 4170 | ||||||
| Upper 50% CI | 5455 | ||||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Crowley, B.E.; Yanes, Y.; Mosher, S.G.; Rando, J.C. Revisiting the Foraging Ecology and Extinction History of Two Endemic Vertebrates from Tenerife, Canary Islands. Quaternary 2019, 2, 10. https://doi.org/10.3390/quat2010010
Crowley BE, Yanes Y, Mosher SG, Rando JC. Revisiting the Foraging Ecology and Extinction History of Two Endemic Vertebrates from Tenerife, Canary Islands. Quaternary. 2019; 2(1):10. https://doi.org/10.3390/quat2010010
Chicago/Turabian StyleCrowley, Brooke Erin, Yurena Yanes, Stella Grace Mosher, and Juan Carlos Rando. 2019. "Revisiting the Foraging Ecology and Extinction History of Two Endemic Vertebrates from Tenerife, Canary Islands" Quaternary 2, no. 1: 10. https://doi.org/10.3390/quat2010010
APA StyleCrowley, B. E., Yanes, Y., Mosher, S. G., & Rando, J. C. (2019). Revisiting the Foraging Ecology and Extinction History of Two Endemic Vertebrates from Tenerife, Canary Islands. Quaternary, 2(1), 10. https://doi.org/10.3390/quat2010010