Abstract
Small mammal remains obtained from the European localities dated to the Eemian (Mikulino) age have been analyzed for the first time at a regional scale based on the present biogeographical regionalization of Europe. The regional faunas dated to the warm interval in the first part of the Late Pleistocene display notable differences in fauna composition, species richness, and diversity indices. The classification of regional faunal assemblages revealed distinctive features of small mammal faunas in Eastern and Western Europe during the Eemian (=Mikulino, =Ipswichian) Interglacial. Faunas of the Iberian Peninsula, Apennine Peninsula, and Sardinia Island appear to deviate from the other regions. In the Eemian Interglacial, the maximum species richness of small mammals (≥40 species) with a relatively high proportion of typical forest species was recorded in Western and Central Europe and in the western part of Eastern Europe. The lowest species richness (5–14 species) was typical of island faunas and of those in the north of Eastern Europe. The data obtained make it possible to reconstruct the distribution of forest biotopes and open habitats (forest-steppe and steppe) in various regions of Europe. Noteworthy is a limited area of forests in the south and in the northeastern part of Europe. In these regions, it seems likely that under conditions of relatively high temperatures characteristic of the Last Interglacial and an insufficient moisture supply there could exist open forest stands or forest-steppe landscapes, as suggested by the presence of species indicative of forest-steppe and steppe north of the forest zone. The results obtained are useful in modeling changes in the mammal faunas as well as environmental changes in entire Europe due to global climatic changes (including the global warming recorded at present).
1. The History of Investigation of the Eemian Interglacial
The Eemian Interglacial was first recognized from boreholes in the vicinity of Amersfoort town, the Netherlands, by Harting [1] and named after the local Eems River. In the parastratotype of the Eemian Interglacial in the Amsterdam-Terminal section it was dated by infrared stimulated luminescence at 118 ± 6.3 ka BP [2].
In the north of Eastern Europe similar dates were obtained by 230Th/U for the Mikulino Interglacial deposits studied in Cheremoshnik key section (the Upper Volga drainage basin, Russia): 114.2 + 11.6/− 9.2 ka BP and 115.5 + 15.8/− 11.6 ka BP [3].
In the south of Eastern Europe, a lagoon and marine sequence including Karangatian mollusks and small mammal fauna has been described in the Eltigen section (eastern Crimea, Russia) and dated by 230Th/U to 127 ± 8.9 ka BP (cycle I) and 107 ± 7.7 ka (cycle II) [4]. The paleomagnetic measurements performed on those layers revealed a reverse polarity event identified as the Blake Event that correlates with the Eemian interglacial.
The unique drilling on the Greenland ice in 2007–2012—North Greenland Eemian Ice Drilling, NEEM-site (North-West Greenland, 77.45° N, 51.06° W) [5]—provided data on the duration (130–115 ka BP) of the Eemian Interglacial and on the chronological position of its optimum (126 ka BP). The interglacial is correlated with marine isotope stage (MIS) 5e.
As follows from abundant paleontological evidence (pollen assemblages, diatoms, mollusks, mammal fauna, etc.), the climate of the Eemian Interglacial was warmer than that of the recent Holocene interglacial. According to the scenario developed by Bakker et al. [6], the July temperature in the Northern Hemisphere could be 0.3 to 5.3 K above that of the present day. The warming also exerted an effect on the global (eustatic) sea level that was 5.5–9 m above that of today [7].
The interglacial is known as the Ipswichian in Great Britain, Riss-Würm in the Alps, and Sangamonian in North America. Specialists differ considerably in their estimates of the environments of that time. Some are of opinion that at the Eemian maximum (~126 ka BP) forests were widely spread over Europe as far north as North Cape (Norway, 71.16° N, 25.78° E) and the Barents Sea coast in Eastern Europe, including regions that are now covered with tundra vegetation [8]. According to other reconstructions [9], the most densely forested regions in the Northern Hemisphere (>60% of the total area) occurred approximately between 60° and 70° N. During Eemian optimum hippopotamus inhabited the Rhine and Thames river basins; this indicates a climate considerably warmer than today.
In Eastern Europe the interglacial is known as Mikulinian named after the Mikulino settlement in the Rudnya district, Smolensk Region, Russia (55.034° N, 31.108° E). The interglacial deposits located near the abovementioned Mikulino settlement, the city of Gadyach and Borok village near Bezhetsk are intercalated between two glacial tills—Dnieper and Valday; they were described by Markov [10]. The name “Mikulinian Interglacial” was suggested by Moskvitin [11].
2. Materials and Methods
There are voluminous data accumulated on the composition of local mammal faunas in Europe attributable to the Eemian (Mikulinian) Interglacial. Over the last 30 years the authors of this paper collected a huge amount of data with detailed information on the Pleistocene mammal localities in Europe and northern Asia. The remains of small mammals found in the Mikulino localities on the East European Plain were collected in the field and analyzed by the first author [12].
The studies provided data on the taxonomy of the European mammal faunas attributed to the Eemian Interglacial and revealed certain patterns in the spatial variability of the species composition in local faunas. They permitted to outline paleogeographical regions and to reconstruct paleoclimate and paleoenvironment in various regions of Europe based on paleontological information.
There are certain difficulties in dating the Eemian Interglacial faunas. Some of the recently studied faunas are dated with a high accuracy, using, for example, 230Th/U or infrared stimulated luminescence. Unfortunately, quite often the fauna localities attributed to that warm interval were dated only by geological, palynological, and archeological methods, undoubtedly giving less precise results. It should be noted, however, that the data on the small mammals attributed to the Eemian Interglacial usually display morphological and ecological characteristics (in particular taxonomic composition) that are clearly different from faunas dated to the preceding and succeeding glacial epochs.
The present work summarizes data from 91 small mammal localities (Figure 1; Table S1), including 51 localities with absolute dates and 40 ones with relative dates. In some cases the dates obtained on one locality vary over a wide range which hinders the fauna attribution to a certain interval.
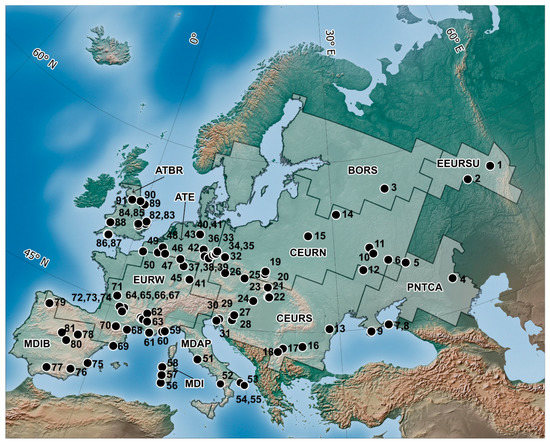
Figure 1.
Map of the Last Interglacial small mammal localities discussed in this paper and European bioregions (for more details see Table S1): 1-Bobylek (Russia) [16], 2-Krasnyi Bor (Russia) [17], 3-Cheremoshnik (Russia) [3,18], 4-ChernyYar (Russia) [19,20], 5-Shkurlat (Russia) (Shkurlat layers) [21], 6-Chernianka (Russia) [21], 7-El′tigen (Russia) [22], 8-Beglitsa (Russia) [23], 9-Kabazi II (Russia) [24,25], 10-Malutino (Russia) [26], 11-Mikhailovka 5 (Russia) [27], 12-Gadiach (Ukraine) [21], 13-Novonekrasovka (Ukraine) [28], 14-Borisova Gora (Belarus) [29], 15-Timoshkovichi (Belarus) [30], 16-Vârtop casa de Piatra (Romania) [31], 17-Kozarnika (Bulgaria) [32], 18-Mala Balanica (Serbia) [33], 19-Biśnik Cave (Poland) [34], 20-Nietoperzowa Cave (Poland) [35], 21-Gánovce-Hrádok (Slovenia) [36], 22-Kalman Lambrecht Cave (Hungary) [37], 23-Horváti-lik (Hungary) [38], 24-Süttő Travertine Complex (Hungary) [39], 25-Za Hájovnou Cave (Czech Republic) [40], 26-Chlupá čova sluj (Czech Republic) [41], 27-Vindija Cave (Croatia) [42,43], 28-Veternica (Croatia) [42,43], 29-Krapina (Croatia) [44], 30-Betalov podmol (Slovenia) [43], 31-Caverna degli Orsi (Italy) [45], 32-Schönfeld (Germany) [46], 33-Gröbern I (Germany) [46,47], 34-Grabschütz (Germany) [46], 35-Rabutz (Germany) [46], 36-Neumark-Nord 2/2 (Germany) [46,48], 37-Burgtonna I (Germany) [46], 38-Burgtonna II (Germany) [46], 39-BurgtonnaI-III (Germany) [46], 40-Taubach (Germany) [49], 41-Weimar-Ehringsdorf (Germany) [50,51,52], 42-Einhornhöhle (Unicorn Cave) (Germany) [44], 43-Lehringen (Germany) [47,53], 44-Biedermann travertine (Germany) [51,52], 45-Wallertheim (Germany) [53,54], 46-Tönchesberg (Germany) [53], 47-Scladina, Sclayn (Belgium) [55,56], 48-Zemst, Bosvan Aa (Belgium) [57], 49-Mesvin IV (Belgium) [58,59], 50-Caours (France) [60], 51-Castel di Guido (Italy) [61,62], 52-Grotta Grande of Scario (Italy) [63], 53-Avetrana (Italy) [64], 54-Cava Nuzzo (Italy) [65,66], 55-San Sidero 3 (Italy) [67,68], 56-Grotta dei Fiori (Italy) [69], 57-San Giovanni di Sinis (Italy) [70], 58-Dragonara Cave (Italy) [71], 59-Grotta del Principe (Monako) [72], 60-Lazaret (France) [73,74], 61-Cèdres (France) [75,76], 62-La Baume Moula-Guercy (France) [77,78], 63-Le Grand Abri aux Puces (France) [79], 64-Saint-Marcel (d′Ardeche) (France) [80], 65-Orgnac 3 (France) [81,82], 66-Baume Flandin (France) [83,84], 67-Abri-des-Pêcheurs (France) [85,86], 68-Grotte des Ramandils (France) [87,88], 69-Cova del Rinoceront (Spain) [89], 70-Grotte du Portel (France) [90,91], 71-Abri Bourgeois-Delaunay, La Chaise (France) [92], 72-Coudoulous I (France) [93], 73-Vaufrey (France) [94,95], 74-Bramefond (France) [96,97], 75-Bolomor (Spain) [98,99], 76-Cueva Negra (Spain) [100,101,102], 77-Cueva del Angel (Spain) [103], 78-Ambrona (Spain) [104,105], 79-Cova Eirós (Spain) [106,107], 80-Arenero de Arriaga (Spain) [108,109], 81-Villacastin (Spain) [110,111], 82-Swanton Morley (U.K.) [112], 83-Shropham Gravel Pit (U.K.) [113], 84-Barrington Chalk Pit (U.K.) [113], 85-Woolpack Farm beds (U.K.) [114], 86-Tornewton Cave (U.K.) [115,116], 87-Joint Mitnor Cave (U.K.) [117,118], 88-Bacon Hole (U.K.) [114,118,119], 89-Sewerby cliff (U.K.) [120,121], 90-Kirkdale Cave (U.K.) [122,123], and 91-Victoria Cave (U.K.) [113,124]. Bioregions: Mediterranean Islands (MDI), Mediterranean Iberian (MDIB), Mediterranean Apenninian (MDAP), Ponto-Caspian (PNTCA), West European (WEUR), Atlantic East (ATE), Atlantic Britain (ATBR), Central European South (CEURS), Central European North (CEURN), East European South (EEURS), Boreal South (BORS), and East European Southern Ural (EEURSU).
When describing the faunal variability, the sites were grouped on the basis of their geographical location: Western Europe, Central Europe, Eastern Europe (including the East European Plain, Crimea, and the Urals), and the Mediterranean region (the Iberian Peninsula, Apennine Peninsula, and Sardinia Island). To describe the small mammal species diversity in more detail, the data were aggregated in smaller spatial units.
The biogeographical zones used in this study are based on modern terrestrial mammal distribution [13], combined with general approaches that have been described in literature [14,15]. All data on species ranges (presence (1), absence (0)) were aggregated on cell grid 150 × 150 km. The matrix of pairwise dissimilarities of grid cells was calculated using the Jaccard coefficient as the distance metric. Then, for the purpose of the cell ordination, the matrix was processed by means of nonmetric multidimensional scaling. Four first axes are used for cell clustering using the dichotomy of k-means clustering algorithms in order to form clusters of similar grid cell. At the 7th level of clustering, there are 12 European bioregions, which include Eemian Interglacial localities used in this study. The bioregions characterized here are as follows (Figure 1): Mediterranean Islands (MDI), Mediterranean Iberian (MDIB), Mediterranean Apenninian (MDAP), Ponto-Caspian (PNTCA), West European (WEUR), Atlantic East (ATE), Atlantic Britain (ATBR), Central European South (CEURS), Central European North (CEURN), East European South (EEURS), Boreal South (BORS), and East European Southern Ural (EEURSU).
For each bioregion, the number of species, Shannon (H) diversity index, and evenness (E) index were calculated. We used a permutation test [125] for testing the difference between diversity-indices yielded from any pairs of bioregions. All the statistical analyses were carried out using package PAST v. 3.12 (https://folk.uio.no/ohammer/past/) [126].
3. Results
3.1. The East European Plain, Crimea and the Urals
The small mammal data from Eastern Europe have been collected and studied by different paleontologists. The Eemian (Mikulinian) small mammal fauna data from Eastern Europe collected until the late 20th century have been summarized by A.K. Markova [12].
New data resulted in a revision of the stratigraphic status of the fauna from the Cherny Yar locality (Russia), formerly identified as a Middle Pleistocene Khozarian faunal assemblage [127,128]. New dates obtained using optically stimulated luminescence (OSL) techniques suggest a younger Mikulinian age of the locality [128]. The enamel thickness differentiation quotient (SDQ) of the water voles Arvicola in the Cherny Yar complex also permits a correlation of the faunal assemblages with Mikulinian faunas [129].
Absolute dates have been obtained for several localities with a Mikulino age, including Eltigen (Russia), Cheremoshnik (Russia), and Kabazi II (Russia). Most important among them are the dates obtained for the Cheremoshnik locality, which provided the corroborative evidence for its attribution to the Mikulinian Interglacial [3].
Detailed studies of the Paleolithic sites in Crimea performed by a team of Ukrainian archeologists proved an early age of the lowermost cultural layers in the multilayer site Kabazi II dated to the Middle Paleolithic [130]. The earliest layer of the site (Unit VI) with rich fauna of small mammals was attributed to the Mikulino Interglacial on the basis of its geological context and paleopedological data [23]. The absolute dates confirmed the stratigraphic position of the fauna.
The species composition of the East European small mammal faunas dated to the Mikulino Interglacial is given in Tables S2–S6.
It should be noted that the studied localities are widely spread and positioned in different natural zones than today; the appearance of small mammal faunas dating to the Mikulino age seems also to be affected by the diversified environments of that time. The interglacial fauna provides the possibility of reconstructing the natural zonality of the East European Plain and permits the identification of specific features of the environments in Crimea. The northernmost localities (Timoshkovichi (Belarus), Borisova Gora (Belarus), Konevich (Belarus), Niyatesos (Lithuania), Cheremoshnik (Russia), and Krasny Bor (Russia)) yielded a wide assortment of small mammal species (Eurasian red squirrel S. vulgaris, wood mouse/yellow necked mouse Apodemus ex gr. sylvaticus-flavicollis, bank vole Clethrionomys glareolus, field vole Microtus agrestis, and others) indicative of the forest zone. Steppe species are present in a small number. The Borisova Gora locality yielded remains of collared lemming Dicrostonyx torquatus. Motuzko [30] attributes their presence to the fact that the fauna belongs to the very beginning of the interglacial, being actually transitional from the preceding (Dnieper) glaciation.
Farther south, in Malyutino, Chernyanka, Mikhailovka 5, and Gadyach forest species are also present, for example wood mouse Apodemus sp., bank vole Cl. glareolus, Microtus (Terricola) subterraneus, and the field vole Microtus agrestis. Steppe dwellers are also well represented (steppe pica Ochotona pusilla, ground squirrel Spermophilus sp., mole rat Spalax, yellow steppe lemming Eolagurus, steppe lemming Lagurus, narrow-headed vole Lasiopodomys (Stenocranius) gregalis, and others). This kind of assemblage indicates a forest-steppe environments [26,131].
In the southeast of Eastern Europe, Shkurlat (Russia), Eltigen (Russia), Beglitsa, Mezin paleosol complex (Russia), and Cherny Yar are completely devoid of forest species. We consider, as an example, the Shkurlat stratotype locality (the recovered mammal remains formed the basis of the Shkurlat faunal complex corresponding to the Mikulino Interglacial [26]). Among small mammals the complex includes remains of steppe pika Ochotona pusilla, little ground squirrel Spermophilus pygmaeus, Pygerethmus (Alactagulus.) acontion, yellow steppe lemming Eolagurus luteus, and steppe lemming Lagurus lagurus [132]. The Cherny Yar fauna is also dominated by steppe animals: little ground squirrel S. pygmaeus, Northern mole-vole Ellobius talpinus, yellow steppe lemming E. luteus, and the steppe lemming L. lagurus [133]. Some species found in the localities prefer humid environments among them are the water vole Arvicola terrestris and the tundra vole Microtus oeconomus. The water voles from the Mikulino localities are characterized by their advanced (progressive) morphology, with SDQ values of ~100 [12,130].
The Mikulinian (Eemian) layers of the multilayered Middle Paleolithic site Kabazi II (Unit VI) in Crimea, dated by using absolute dating methods, include abundant small mammal remains characterized the different environmental conditions [24]. The fossil record includes remains of forest species: forest dormouse Dryomys nitedula, yellow-necked mouse A. flavicollis, pine vole M. (T.) subterraneus, and water vole A. ex gr. terrestris a species that indicates wet habitats. Steppe species are represented by S. pygmaeus, Spalax microphtalmus, Northern mole-vole Ellobius talpinus, yellow steppe lemming E. luteus, and the narrow-headed vole L. (Stenocranius) gregalis. This group is dominated by Altaian vole Microtus obscurus an inhabitant of meadow and steppe biotopes. The species composition of this particular fauna suggests highly diversified environments in the Crimean Mountains.
Layers 4 and 6 of the Bobylek Paleolithic site (Middle Urals), tentatively attributed to the Mikulino Interglacial (no absolute date is available), yielded small mammals of various environmental preferences: forest (Apodemus sp., Cl. ex gr. rutilus-glareolus, and M. agrestis), steppe pika Ochotona sp., ground squirrel Spermophilus sp., great jerboa Allactaga major, lesser fat-tailed jerboa Pygeretmus (A.) pumilio, black-bellied hamster Cricetus cricetus, narrow-headed vole L. (S.) gregalis), and inhabitants of wet areas (water vole A. ex gr. terrestris and tundra vole M. oeconomus) [134]. The fact of ecologically different species being found together is explained by the environmental diversity of the mountainous area and the altitudinal zonation. The remains were accumulated in the form of pellets from predatory birds hunting small animals within a few kilometers from the site at slopes varying in exposure and belonging to different altitudinal zones.
Materials of considerable importance were obtained from the Novonekrasovka site in the Danube drainage basin. The small mammal fauna were recovered from lagoonal and marine sediments of the Black Sea attributed to the Karangatian transgression (correlated with the Mikulino Interglacial and MIS 5e) based on the evidence retrieved mollusk fauna [28]. The SDQ index and size of the water vole Arvicola teeth agree with the data from other Mikulino localities [12]. The mammal fauna from the Eltigen section (eastern Crimea) is also confined to the Black Sea lagoonal-marine sediments and, in common with the Novonekrasovka sequence, includes Karangatian mollusk shells along with small mammal remains. That locality was dated by 230Th/U. In addition, paleomagnetic measurements revealed the presence of the Blake Event correlated with the Mikulino Interglacial [4].
The results obtained from the small mammal faunas of Eastern Europe indicate a natural zonation existence of the East European Plain, with clearly recognizable forest, forest-steppe, and steppe zones. The geographical position of the zones hardly differs from the present zonation, which may be attributed to the specificity of the dominant air mass transport from the Atlantic. Faunas of the Crimea mountains and the Urals suggest highly diversified environments within the mountain regions.
3.2. Central Europe
A considerable number of small mammal localities discovered in Central Europe are confidently correlated with the Eemian Interglacial. The species composition of a number of faunas is presented in Table S2. Various dating methods (U/Th), electron spin resonance dating (ESR), optically stimulated luminescence (OSL), and thermoluminescence dating (TL) have been applied. Many localities yielded remains of forest dwellers: European mole Talpa europaea, common shrew Sorex araneus, pygmy shrew Sorex minutus, Eurasian red squirrel S. vulgaris, bank vole Cl. glareolus, wood mouse A. sylvaticus, European pine vole M. (T.) subterraneus, and field vole M. agrestis. The species composition strongly suggests widely spread forest and shrub biotopes in the vicinities of the sites.
The paleo-landscapes of Central Europe dating to the Eemian Interglacial display a certain zonality in their spatial distribution. The sites located at latitudes ~48–51° N including Taubach (bed 11), Weimar-Eringsdorf (Upper Travertine), Burgtonna (lower layer in section Cm), Biśnik cave (layers 13 and 12), and some others yielded a considerable number of forest species: western hedgehog Erinaceus europaeus, European mole T. europaea, common shrew S. araneus, pygmy shrew S. minutus, bicoloured shrew C. leucodon, common white-toothed shrew Crocidura russula, lesser white-toothed shrew C. suaveolens, Eurasian beaver Castor fiber, fat dormouse Glis glis, forest dormouse D. nitedula, wood mouse A. sylvaticus, bank vole Cl. glareolus, and the European pine vole M. (T.) subterraneus [46,135,136]. A few small mammals found in those localities prefer open landscape habitats (Spermophilus citellus, Al. major, L. lagurus, and L. (S.) gregalis). Forest environments undoubtedly dominated in Central Europe at those latitudes.
Further east the Central European localities (Kozarnika, layers 9a–9b (Bulgaria), Kalman Lambreht cave (Hungary), Horvati-lik and Za Hájovnou cave (Czeck Republik), Vârtop casa de Piatra (Romania), and others) feature a noticeable presence of the open landscape species (steppe pika O. pusilla, ground squirrel Spermophilus sp., yellow steppe lemming E. luteus, steppe lemming L. lagurus, and the narrow-headed vole L. (S.) gregalis. Less common are golden hamster Mesocricetus auratus, Newton’ medium hamster Mesocricetus newtoni, hamster Allocricetus bursae, and the gray dwarf hamster Cricetulus migratorius) (e.g., [32,37,38]). Remains of those steppe species are usually found together with forest and eurytopic species. Specific species (water vole Arvicola, tundra vole M. oeconomus, Russian desman Desmana moschata, and the Eurasian water shrew Neomys fodiens) prefer wet biotopes; they were widely distributed over the European subcontinent. The obtained data suggest forest-steppe environments in the regions, both on the plains and in mountains.
Remains of a tundra species collared lemming Dicrostonyx gulielmi were described in two localities in Hungary and Poland (Horvati-lik and Biśnik cave, layers 12 and 13) [38,136]. The possibility must not be ruled out that those remains were redeposited from earlier layers dated to the Saale glaciation.
Of interest are findings of porcupine remains (Hystrix brachyura vinogradovi) in a few localities (Burgtonna, lower layer (Germany); Taubach, bed 11 (Germany); Weimar–Eringsdorf, upper travertine (Germany); Kalman-Lambrecht cave (Hungary)) [135,137,138,139,140]. That mammal is thermophilic and occurs nowadays mostly in subtropical areas. Its presence in the Eemian layers supports the view of the Eemian Interglacial climate was warmer than the present-day interglacial. Unlike the East European faunas, remains of arid open space inhabitants (such as pygmy ground squirrel S. pygmaeus, greater mole rat Spalax microphtalmus, Northern mole rat E. talpinus, fat-tailed jerboa P. (A.) pumilio, as well as Altaian vole M. obscures) have not been found in Central Europe. Their absence may be attributed to specific features of the fauna evolution there.
In summary, it can be stated that the composition of the Eemian faunas in Central Europe suggests a forest and forest-steppe landscape in the area. Steppe biotopes, widely spread over the southeast of Eastern Europe, are practically absent in the central regions of Europe.
3.3. Western Europe and the British Isles
In Western Europe there are two main groups of small mammal localities dated to the Eemian Interglacial. The localities in the northern part of the region (Biedermnann travertine and Tönchesberg, Zemst layer IIB (Germany), Scladina and Mesvin IV (Belgium), Caours (France)), mostly north of 47° N, yielded remains of Erinaceus sp., T. europaea, S. araneus, S. caecutiens, S. minutus, Crocidura leucodon, Lepus sp., O. pusilla (Biedermnann travertine locality), Sciurus vulgaris, Marmota marmota, Sicista betulina (Biedermnann travertine locality), C. cricetus, C. major, A. bursae, G. glis, A. sylvaticus, Cl. glareolus, A. terrestris-cantianus, M. (T.) subterraneus (Scladina I, locality l.5), M. agrestis and M. arvalis (e.g., [46,56,59,60,141,142] (Tables S4 and S5). Most of the localities have direct datings. The ecological characteristics of the fauna indicate a dominance of forest biotopes; species of open landscapes (pika and hamsters) are present in insignificant quantities. Some data must be verified, for example, the attribution of water vole from Tönchesberg locality to A. cantianus, which is characteristic for the Middle Pleistocene faunas [141].
Quite different in composition are faunas recovered from localities farther south (Baune Flandin (France), Cèrdes (France), Grotte Vaufrey, layer IV (France); Orgnac 3, layer 5a (France); Saint-Marcel, layer U (France); La Baume Moula-Guercy, layer XV (France); Lazaret (layers: C III 7-265, 236, CII 17-326) (France); Le Grand Abri aux Puces L. 2 (France), Grotte de Portel, L, K (France); Coudoulus I (France), Abri-des-Pêcheurs, ensemble 1 (France), Grotta del Principe (Monaco), etc.) [70,76,77,78,84,90,143,144,145,146,147,148]. The faunas of those localities, along with c Talpa europea, Sorex sp., C. leucodon, C. russula, Lepus sp., S. vulgaris, A. bursae, G. glis, A. sylvaticus, Cl. glareolus, M. (T.) subterraneus, L. (S.) gregalis (Grotte Vaufrey, layer IV; Orgnac 3, layer 5a, Moula, La Baume Moula-Guercy; Le Grand Abri aux Puces), and M. arvalis and M. agrestis, include also the southwestern water vole A. sapidus (the species characteristic of the Iberian Peninsula and the south of Western Europe), the vole Pliomys coronensis (a species extinct from Eastern and Central Europe as early as the beginning of the Middle Pleistocene), Cabrera’s vole Microtus (Iberomys) cabrerae brecciensis typical also of the Iberian Peninsula; voles Microtus (Terricola) duodecimcostatus and M. (T.) pyrenaicus (also commonly found on the Iberian Peninsula). La Baume Moula-Guercy [77,78,146] yielded remains of Balkan snow vole Dinaromys bogdanovi, the only representative of the Pliomyini tribe persisting today.
It may be concluded that the southern faunas of Western Europe are unique in having endemic species present and they are undoubtedly closely correlated to faunas of the Iberian Peninsula. Based on species composition, the region was dominated by forest and bush communities alternating with communities that inhabit open areas. The latter are characterized by steppe biotope species: A. bursae, L. (Stenocranius) gregalis, and L. lagurus (Orgnac 3, layer 5a [83]). A wide occurrence of marmots M. marmota is recorded in mountains.
On the whole, the Eemian faunas of Western Europe point to mild climate, which enabled the forest landscapes to expand widely on the continent. Only a few species recovered from the Eemian localities are inhabitants of open biotopes. Cold-tolerant animals were completely absent.
Few species of small mammals have been recovered from Eemian (Ipswichian) localities in the British Isles (U.K.), 15 taxa of Eulipotyphla, Lagomorpha, and Rodentia have been described in Bacon Hole, Barrington Chalk Pit, Joint Minor cave, Kirkdale Cave, Shropham Gravel Pit, Swanton Morley, Tornewton Cave (Hyaena stratum), Victoria cave, Woolpack Farm beds, and some others (Table S6) (e.g., [112,113,114,115,116,117,118,119,122,123,124]). Most of the species are inhabitants of forest and forest-steppe biotopes (T. europaea, Sorex araneus, S. minutus A. sylvaticus, Cl. glareolus, and M. agrestis). There are also some wet biotope dwellers (Castor fiber, N. fodiens, A. terrestris-cantianus, and M. oeconomus). Evidently, the small mammal fauna of the British Isles dated to the last interglacial suggests a mild climate beneficial for the expansion of forest biotopes. No cold-tolerant species are present. Among large mammal fauna there are some thermophilic species, such as Hippopotamus amphibious and Palaeoloxodon antiquus [149].
3.4. The Iberian Peninsula, Apennine Peninsula, and Sardinia Island
Small mammal faunas that inhabited the Mediterranean during the Eemian Interglacial were of a specific kind. The mammal remains of that age were recovered in Italy from Castel di Guido [63,64], Grotta Grande of Scario (Salerno) [63], Cava Nuzzo (Salento region) [65,66], and Avetrana (Taranto) (Table S6, Figure 1) [62]. The Castel di Guido fauna includes only a few small mammal species: Lepus europaeus, Lepus sp., and Castor sp. The remains of E. europaeus, L. europaeus, Oryctolagus cuniculus, and Microtus (Terricola) savii (Table S6) recovered from Cava Nuzzo, in the southeast Apennine Peninsula, suggest the prevalence of open biotopes. In the same region animal bones identified as E. europaeus, M. (T.) savii, Pleistocene porcupine H. b. vinogradovi were found in the Avetrana karst cavity (Table S6) [64]. The authors assigned that fauna to the last interglacial. It is extremely rare that Pleistocene porcupine remains are found in the deposits of that warm interval in Italy [150]. A considerable number of large and small mammals were recovered from Grotta Grande of Scario (Table S6), including M. (T.) cf. savii, A. terrestris, G. glis, and Crocidura suaveolens, three species of Talpa (T. caeca, T. romana, and T. europaea) and many others. The abundance of M. (Terricola) cf. savii seems to indicate open arid biotopes in the immediate vicinity of the site. The presence of Cl. glareolus, A. sylvaticus, and Eliomys quercinus and some other species of the same environmental preference suggests forest and shrub communities near the site. In the authors’ opinion, human presence at the site may be assigned to the Eemian Interglacial (before its optimum).
Eemian localities yielding small mammals are found in abundance on the Iberian Peninsula (in Spain): Cueva del Angel (layer I-XIII), Arenero de Arriaga (layer l.I), Ambrona (layer AS1), Cueva Negra del Estrecho (layer 15), Villacasin, Bolomor (layer II), Cova del Rinoceront (layers III a, b, c); Cova Eiros (layer 3) (Table S6, Figure 1). The southernmost faunas of the peninsula (Cueva del Angel, Cueva Negra del Estrecho, Bolomor) include E. europaeus, T. europaea, Sorex minutissimus, Neomys sp., Galemys sp., Prolagus sp., S. vulgaris, A. bursae, E. quercinus, A. sylvaticus, Arvicola sapidus, and M. (I.) cabrerae brecciensis. Specific features of the faunas are the presence of an Iberian Peninsula endemic—Pyrenean desman Galemys sp., southern water vole A. sapidus, as well as Cabrera vole M. (I.) cabrerae brecciensis, all occurring in the southern Mediterranean and on the Iberian Peninsula. Dominant in the fauna are small mammals, inhabitants of forest and shrub landscapes, those of wet biotopes are also present.
Some of the above-listed animals—E. europaeus, T. europaea, Neomys sp., Galemys sp., A. bursae, A. sapidus, and M. (I.) cabrerae brecciensis—occur farther north as well, at the center of the peninsula, in the localities (Spain) Ambrona, Arenero de Arriaga, and Villacastin (Figure 1). Additionally, there are S. minutus, bicoloured shrew C. leucodon, European rabbit O. cuniculus, porcupine Hystrix sp., Clethrionomys sp., and M. (T.) duodecimcostatus, which have not been found in the south [104,108,110]. Pliomys coronensis has not been found as yet in the local faunas of that period, though it has been recorded in the earlier and later Pleistocene faunas, as well as in layers dated to other stages of MIS 5 in the localities Valdegoba, Teixoneres cave, and Lezetxiki (Spain) [151,152,153].
The small mammals described in Cova Eiros locality (42.7° N), in the mountain region of Galicia in the NW of the peninsula (Figure 1), include E. europaea, Spanish mole Talpa occidentalis, wood mouse A. sylvaticus, Southern water vole A. sapidus, European snow vole Chionomys nivalis, Lusitanian pine vole M. (T.) lusitanicus, and M. arvalis [106]. The fauna is noted for the presence of endemic species, Spanish mole T. occidentalis, and Lusitanian pine vole M. (T.) lusitanicus. The snow vole C. nivalis is typical of the European mountains. The data available at present suggest forest and shrubland biotopes to be dominant in the low mountains of Galicia.
There are three localities attributable to the last interglacial described on Sardinia Island, namely San Giovani di Sinis, Dragonara Cave, and Grotta dei Fiona. The small mammals described include only a few species: Sardinian mole Talpa tyrrhenica, Sardinian shrew Episoriculus similis, Sardinian pika Prolagus sardus, large field mouse Rhagamus orthodon, and Tyrrhenian vole M. (Tyrrhenicola) henseli [69,70,81], all of them being endemic to Sardinia, which suggests a long-term isolation of the island from the continent. Abundant materials on the Plio-Pleistocene theriofaunas of Sardinia Island were summarized by Palombo [151].
The above overview shows essential differences in small mammal faunas of the bioregions. In the dendrogram (Figure 2a) there is a distinctly outlined insular bioregion MDI, its fauna consists entirely of endemics of Sardinia dominated by M. (Tyrrhenicola) henseli (Table 1).
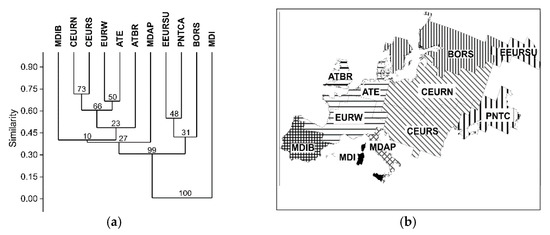
Figure 2.
(a) Bioregion classification based on the composition of the small mammal faunas (method: unweighted pair group method with arithmetic mean clustering (UPGMA); similarity: Sorensen coefficient; bootstrap support values (%, 1000 repeats) are shown at the branch nodes); (b) the sketch map of bioregions’ clustering.

Table 1.
Dominant and subdominant taxa of bioregions.
The ensuing analysis permits recognition of a cluster of bioregions in the east of Europe, of these the regions EEURSU and PNTCA being closest to each other. A common feature of the two regions consists in a considerable proportion of xerophilous species (С. cricetus, S. pygmaeus, L. lagurus, E. luteus, and L. (S.) gregalis). The dominance of these is most pronounced in the SSE of Europe, in the PNTCA bioregion. At the same time, a wide occurrence of yellow-necked mouse (A. flavicollis) in the EEURSU regional fauna suggests widespread broadleaf forests in the Southern Urals during the Last Interglacial.
Boreal South region is rather close to the EEURSU in the fauna composition. In spite of its position farther north (57.12° N; Figure 2b), forest-steppe and steppe species are present including marmot (Marmota), mole rat (Spalax), hamster (С. cricetus), narrow-headed vole (L. (S.) gregalis), yellow steppe lemming (E. luteus), etc.
The southern Mediterranean bioregions (MDAP and MDIB) appear to differ considerably from each other in the fauna composition. The small mammal faunas of the Iberian and Apennine peninsulas are also distinctly different from those of continental Europe. As our materials show, the most common species in the MDAP region are Apennine hare Lepus corsicanus and/or L. europaeus, Savi’s pine vole M. (T.) savii, and M. (T.) brachycercus [154]. The most widespread in MDIB bioregion was A. sylvaticus, with A. sapidus and M. (Iberomys) cabrerae brecciensis as its codominants.
East European bioregions (СEURN and CEURS) differ distinctly enough from West European ATE and EURW. It is quite possible that the demarcation between them during the Last Interglacial was close to its present position. Dominant in the СEURN fauna was the field vole (M. agrestis), its co-dominants T. europaea, Cl. glareolus, and others (Table 1). In the southern bioregion CEURS small mammal assemblages seem to be polydominant. Along with species typical of forest, forest-steppe and meadows (Cl. glareolus and M. arvalis), the proportion of xerophilous species of open biotopes (S. citellus and L. lagurus) was considerable. The European mole (T. europaea) and beaver (Castor fiber) occur most often in the localities in the north and northwest of Western Europe (ATE region), with Sorex araneus, M. (T.) subterraneus, and M. arvalis as codominants.
The small mammal fauna of the British Isles is similar to continental fauna in species composition, though the number of species is insignificant. The most common animals are the water vole (A. terrestris), wood mouse (A. sylvaticus), and the field vole (M. agrestis), no specific taxa has been recorded.
There is an essential difference in the species richness of individual bioregions (Table 2). Faunas of the three regions (EURW, CEURN, and CEURS) include at least 40 species. This is lower in the ATE region (~29 species), as well as in regions MDIB and PNTCA (no less than 24 taxa). The “Island effect” makes itself evident in a low diversity of faunas; that may be clearly seen in the faunas of the Apennine Peninsula (MDAP, at least 17–19 taxa), Great Britain (ATBR, 14 taxa), and Sardinia Island (MDI, 5 taxa). A low diversity index denotes the East European bioregion BORS.
Table 2.
Species richness and diversity indices (Shannon diversity index (H) and evenness diversity index (E)) of bioregions and their approximate 95 percent confidence intervals computed with a bootstrap procedure.
The diversity indices (H, E) display specific features of species occurrence, including the degree of individual taxa predominance (Table 2 and Table 3). The Shannon index varies regularly from 3.57 (EURW) to 1.52 (MDI). Higher values of the H index are typical for the regions marked with high species richness (CEURN and CEURS). A low diversity of small mammal fauna is characteristic, apart from the Sardinia Island fauna, also for Great Britain (ATBR), as well as the BORS and MDAP bioregions. Relatively high dominance of a few taxa (index E) have been documented in faunas with a high (EURW) and a low (ATBR) species richness. The interregional differences of the Shannon index are statistically more significant than the differences revealed by the evenness index (Table 2).
Table 3.
Pairwise comparisons Shannon diversity index (above diagonal) and evenness diversity index (under diagonal) using permutation test (probability of a no difference).
4. Summary and Conclusions
This work presents the first overview of the small mammal faunas of Europe dating to the last (Mikulinian/Eemian/Ipswichian) interglacial at a sub-continent scale. The wide spatial coverage of the studies revealed principal differences in the faunas, most probably resulting from the nonuniform climate in various regions of Europe, as well as from the geographic position of the regions, their physiographic characteristics, and the degree of isolation (the most important in the case of island faunas).
The analysis of the small mammal faunas revealed a considerable number of geographical variants, each with its own specificity in species composition and species richness. A distinct geographic zonation is reflected in the fauna composition, species richness, and the widespread dominance of species.
The faunas of Eastern Europe (in the south of the East European platform and in Crimea), include, in addition to forest species, a considerable number of inhabitants of open landscapes. Typically, xerophilous species of Asian provenance are dominant (or present to a considerable degree) in the south, east, and even in the north of Eastern Europe. These data give evidence for reconstructing a zonal pattern: forest landscapes were spread over the CEURN bioregion (Figure 2b) and most probably including mixed and broadleaf forests; farther south (bioregion CEURS and particularly PNTCA) there were forest-steppe and steppe (including dry steppe) environment. Xero- to mesophytic forest-steppe landscapes existed in the north and northeast of Eastern Europe up to the Urals. The Southern Ural Mountains were covered by montane broadleaf and coniferous-broadleaf forests and forest-steppe and steppe in the forelands. On the whole, such a zonation differs essentially from the modern zonality; in all probability it was related to the specific distribution of rainfall in the region combined with the relatively high mean annual temperatures. It should be noted that, judging from palynological data, open forests and xerophytic forest-steppes developed in the north of the East European Plain during the Last Interglacial, probably due to the absence of conditions for the tundra zone formation.
The paleo-landscapes of Central Europe also display a certain zonation. Small mammal sites located at ~48–51° N indicate forest environments in the north (bioregions ATE and CEURN). Localities in the east of Central Europe (bioregion CEURS) are noted for a noticeable amount of open landscape species, though forest animals are also present. The data suggest forest-steppe environments in these regions.
The micromammal materials from the northern and central regions of Western Europe (bioregions ATE and EURW) strongly suggest the dominance of forest ecosystems. Judging from the dominant mammal species, the landscapes were mostly mixed coniferous-broadleaf forests. In the south they gave way to broadleaf forests (except for mountains). The wide occurrence of M. arvalis in the north (bioregion ATE) suggests a predominance of open habitats including meadows.
The local faunas of the West European south show a noticeable influence from the Mediterranean fauna. Some West European relicts were present; one of them, P. coronensis, should be mentioned. The species became extinct in Central and Eastern Europe as early as the beginning of Middle Pleistocene.
Few species have been identified in the small mammal assemblages from the British Isles. In species composition those faunas are similar to (though much poorer than) those of continental Europe (bioregion ATE); no endemic species are found. The fauna is dominated by inhabitants of forest and wet biotopes.
The small mammal fauna of the Iberian Peninsula, with exception of its northernmost part (bioregion MDIB), was distinct from the fauna of the EURW bioregion bordering MDIB on the north. The dominance of A. sylvaticus and the wide occurrence of typical species such as the southwestern water vole (A. sapidus) and the Cabrera’s vole (M. (I.) cabrerae brecciensis) are indicative of a mild climate with sufficient moisture supply; the aridity increased gradually from north to south. The faunal composition suggests the predominance of forest and shrub vegetation alternating with open sites, the proportion of the latter increasing southwards. Specific features of the MDIB bioregion fauna are the presence of Pyrenean desman (Galemys pyrenaicus), Pyrenean pine vole (Microtus (Terricola) pyrenaicus), Mediterranean pine vole (M. (T.) duodecimcostatus), and some others.
The fauna of Sardinia Island differs from faunas of all the other regions. The Eemian localities of the island contain only endemic forms of small mammals, indicative of a long-term isolation of the island from the continent during the Pleistocene.
The micromammal fauna of the Apennine Peninsula (bioregion MDAP) is not highly diversified in composition. The notable presence of endemic Savi’s pine vole species (at least in the south of peninsula) suggests the wide occurrence of open and relatively arid landscapes.
All the Mikulinian (Eemian, Ipswichian) faunas in Europe show signs of a considerably warmer climate. Under those conditions the rainfall could be important for the development of forests. The climate was favorable for the territorial afforestation in the west of Europe and over a considerable part of Central and Eastern Europe. At the same time the south of Central Europe, as well as considerable areas in the south, southeast, and east of Eastern Europe, could suffer from insufficient moisture supply, which accounts for wide distribution of open xerophytic steppes and mesophytic forest-steppes. In contrast, the southwestern margins of Europe received precipitation in greater abundance in the Last Interglacial than at present and the forest assemblages were widespread in the region, in its middle part in particular. A similar situation presumably existed on the Apennine Peninsula, though this hypothesis cannot be tested as yet due to insufficient data on the small mammals in its northern part. By analogy, we may assume rather arid conditions and open biotope prevalence over a greater part of the Balkan Peninsula at the considered time interval.
To obtain more detailed regional characteristics of the faunas we used the data on the European regions based on the modern ranges of mammal species. Those biogeographic units appear to be distinctly differentiated with respect to the composition of local small mammal faunas of the Last Interglacial age forming “natural” well-interpretable clusters. This is an argument in support of a relative stability of biogeographic units during the Late Pleistocene as had been suggested on the basis of the analysis of mammal species ranges in MIS 3–MIS 1 [155].
Supplementary Materials
The following are available online at http://www.mdpi.com/2571-550X/1/2/9/s1. Table S1: The list of localities used in this work, Table S2: The Eemian small mammals faunas of Eastern Europe, Table S3: The Eemian small mammal faunas of Central Europe, Table S4: The Eemian small mammal faunas of Western Europe, Table S5: The Eemian small mammal faunas of the British Isles, Table S6: The Eemian small mammal faunas of Northern Mediterranean (Apennine and Iberian Peninsulas and Sardinia Island).
Author Contributions
The individual contributions are conceptualization; A.M., methodology, data analyzing, and writing the text; A.M. and A.P.
Funding
These researches were in part funded by the Russian Foundation for Basic Research (RFBR) grant number (18-05-00076) and by the Institute of Geography of Russian Academy of Sciences theme 69.2. “Mammal faunas of Northern Eurasia in the Late Pleistocene—Holocene: taxonomy, diversity, and evolution”.
Acknowledgments
The authors are very grateful to Lutz Maul, Gloria Cuenca-Bescós, Irina Kirillova, and Pavel Kosintcev for their help during the preparation of this paper. We are also very thankful to the reviewers of our article for their very useful and detailed comments. Our special thanks to Claire Fuller Martin (Associate Curator in Anthropology Illinois State Museum, USA) for her great help in editing the original English text.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Harting, P. Le système Éemien. Arch Néerlandaises Sci Exactes Nat la Soc Hollandaise des Sci (Harlem) 1875, 10, 443–454. [Google Scholar]
- Kruk, R.W. U-Series Radiometric Dating of Eemian Molluscs from Amsterdam Basin. About the Last Period That Was Really Warm in Amsterdam; Faculty of Earth Sciences, Free University: Amsterdam, The Netherlands, 1988. [Google Scholar]
- Rusakov, A.; Nikonov, A.; Savelieva, L.; Simakova, A.; Sedov, S.; Maksimov, F.; Kuznetsov, V.; Korkka, M.; Savenko, V.; Starikova, A. Landscape Evolution in the Periglacial Zone of Eastern Europe since MIS5: Proxies from Paleosols and Sediments of the Cheremoshnik Key Site (Upper Volga, Russia). Quat. Int. 2015, 365, 26–41. [Google Scholar] [CrossRef]
- Dodonov, A.E.; Tchepalyga, A.L.; Mihailescu, C.D.; Zhou, L.P.; Markova, A.K.; Trubikhin, V.M.; Simakova, A.N.; Konikov, E.G. Last-Interglacial Records from Central Asia to the Northern Black Sea Shoreline: Stratigraphy and Correlation. Geol. Mijnbouw/Neth. J. Geosci. 2000, 79, 303–311. [Google Scholar] [CrossRef]
- Dahl-Jensen, D.; Albert, M.R.; Aldahan, A.; Azuma, N.; Balslev-Clausen, D.; Baumgartner, M.; Berggren, A.M.; Bigler, M.; Binder, T.; Blunier, T.; et al. Eemian Interglacial Reconstructed from a Greenland Folded Ice Core. Nature 2013, 493, 489–494. [Google Scholar] [CrossRef] [PubMed]
- Bakker, P.; Stone, E.J.; Charbit, S.; Groger, M.; Krebs-Kanzow, U.; Ritz, S.P.; Varma, V.; Khon, V.; Lunt, D.J.; Mikolajewicz, U.; et al. Last Interglacial Temperature Evolution—A Model Inter-Comparison. Clim. Past 2013, 9, 605–619. [Google Scholar] [CrossRef]
- Dutton, A.; Lambeck, K. Ice Volume and Sea Level during the Last Interglacial. Science 2012, 337, 216–219. [Google Scholar] [CrossRef] [PubMed]
- Gerasimov, I.P. Degradation of the Last European Ice Sheet. In Quaternary Geology and Climate 16; Wright, H.E.J., Ed.; National Academy of Science Publ.: Washington, DC, USA, 1969; pp. 72–78. [Google Scholar]
- Nikolova, I.; Yin, Q.; Berger, A.; Singh, U.K.; Karami, M.P. The Last Interglacial (Eemian) Climate Simulated by LOVECLIM and CCSM3. Clim. Past 2013, 9, 1789–1806. [Google Scholar] [CrossRef]
- Gerasimov, I.P.; Markov, K.K. The Ice Age in the Territory of USSR; Tr. In-ta Geogr. AN SSSR, No 38; Academy of Sciences USS: Moscow and Leningrad, Russia, 1939. (In Russian) [Google Scholar]
- Moskvitin, A.N. Mologa-Sheksna Interglacial Lake. Proc. Inst. Geol. Sci. AS 1947, 88, 5–18. (In Russian) [Google Scholar]
- Markova, A.K. The Mikulino (=Eemian) Mammal Faunas of the Russian Plain and Crimea. Geol. Mijnbouw/Neth. J. Geosci. 2000, 79, 293–301. [Google Scholar] [CrossRef]
- The IUCN Red List of Threatened Species. Spatial Data Download. Available online: http://www.iucnredlist.org/download_spatial_data (accessed on 20 July 2017).
- Heikinheimo, H.; Fortelius, M.; Eronen, J.; Mannila, H. Biogeography of European Land Mammals Shows Environmentally Distinct and Spatially Coherent Clusters. J. Biogeogr. 2007, 34, 1053–1064. [Google Scholar] [CrossRef]
- Kreft, H.; Jetz, W. A Framework for Delineating Biogeographical Regions Based on Species Distributions. J. Biogeogr. 2010, 37, 2029–2053. [Google Scholar] [CrossRef]
- Kosintsev, P.A.; Gasilin, V.V.; Gimranov, D.O.; Bachura, O.P. Carnivores (Mammalia, Carnivora) of the Urals in the Late Pleistocene and Holocene. Quat. Int. 2016, 420, 145–155. [Google Scholar] [CrossRef]
- Yakovlev, A.G. Small mammals of the Pleistocene and Holocene of the Bashkir Urals and the western slope of the Southern Urals. Ph.D. Thesis, Institute of Geology, Ufa Federal Research Centre of the RAS, Ekaterinburg, Russia, 1996. [Google Scholar]
- Agadjanian, A.K.; Erbaeva, M.A. Late Cenozoic Rodents and Lagomorphs of USSR Territory; Nauka: Moscow, Russia, 1983. (In Russian) [Google Scholar]
- Titov, V.V.; Golovachev, M.V. The Skeleton of Mammoth Mammuthus trogontherii cf. chosaricus Dubrovo, 1966 from Terminal Middle Pleistocene of the Lower Volga Region (Russia). Russ. J. Theriol. 2017, 16, 15–29. [Google Scholar] [CrossRef]
- Kirillova, I.V.; Svitoch, A.A. New Middle Pleistocene small mammal finds in the Cherny Yar section (Lower Volga basin) and its stratigraphic significance. Dokl. Akad. Nauk 1994, 334, 731–734. [Google Scholar]
- Markova, A.K. Small mammals of Mikulino Interglacial (by the materials of the Russian Plain). Dokl. Akad. Nauk 1985, 284, 718–721. [Google Scholar]
- Krokhmal’, A.I. Microtheriofauna from the Pleistocene marine and estuary sediments of Ukraine. In Geology of Seas and Oceans, Proceedings of the XXII International Conference on Marine Geology, Moscow, Russia, 20–26 November 2017; Lisitzin, A.P., Politova, N.V., Shevchenko, V.P., Eds.; IO RAS: Moscow, Russia, 2017; Volume 1, pp. 192–194. [Google Scholar]
- Tesakov, A.S.; Titov, V.V.; Baygusheva, V.S.; Velichko, A.A.; Timiryova, S.N.; Simakova, A.N.; Borisova, O.K.; Morozova, T.D.; Semenov, V.V.; Syromyatnikova, E.V.; et al. Guide to field excursions. In Fundamental Problems of Quaternary, Results and Main Trends of Future Studies, Proceedings of the VIII All-Russian Conference on Quaternary Research, Rostov-on-Don, Russia, 10–15 June 2013; SSC RAS Publishers: Rostov-on-Don, Russia, 2013; pp. 1–48. (In Russian) [Google Scholar]
- Markova, A. Small mammals from the Palaeolithic site Kabazi II, Western Crimea. In Kabazi II: Last Interglacial Occupation, Environment and Subsistence. The Palaeolithic Sites of Crimea, 1; Chabai, V., Richter, J., Uthmeier, T., Eds.; Shlyakh: Simferopol, Ukraine; Cologne, Germany, 2005; pp. 59–73. [Google Scholar]
- Patenaude, B. Faunal Exploitation at the Middle Palaeolithic Site Kabazi II (Western Crimea). Ph.D. Thesis, Université de Montréal, Montréal, QC, Canada, 2010. [Google Scholar]
- Alekseeva, L.I. The Late Pleistocene Theriofauna of Eastern Europe (Large Mammals); Nauka: Moscow, Russia, 1989. (In Russian) [Google Scholar]
- Agadjanian, A.K.; Glushankova, N.I. Mikhailovka Is the Key Pleistocene Section of the Central Russian Plain; Moscow State University: Moscow, Russia, 1986. (In Russian) [Google Scholar]
- Markova, A.K.; Mikhailesku, C.D. The new locality of mammals and mollusks in Mikulino deposits of lower Danube basin. Bull. Commiss. Quat. Investig. 1990, 59, 94–101. [Google Scholar]
- San’ko, A.F.; Motuzko, A.N. The mollusks and mammals from the limestone tufa of Borisova Gora on Zapadnaya Dvina River. Dokl. Belorussian AN SSR 1991, 35, 1004–1008. [Google Scholar]
- Motusko, A.N. Anthropogene rodents of Belorussia and adjacent territories. In The Problems of Pleistocene; Nauka i Tekhnika: Minsk, Belarus, 1985; pp. 173–188. (In Russian) [Google Scholar]
- Carciumaru, M.; Angelinu, M.; Nitu, E.-C.; Cosac, M.; Muratoreanu, G. Géo-Archéologie du Paléolithique moyen, Paléolithique Supérieur, Épipaléolithique et Mésolithique en Roumanie; Cetatea de Scaun: Târgovişte, Romania, 2007; pp. 1–187. [Google Scholar]
- Popov, V.; Marinska, M. An almost one million year long (Early to Late Pleistocene) small mammal succession from the archaeological layers of Kozarnika Cave in Northern Bulgaria. Cour. Forsch-Inst. Senckenberg. 2007, 259, 79–92. [Google Scholar]
- Rink, W.J.; Mercier, N.; Mihailović, D.; Morley, M.W.; Thompson, J.W.; Roksandic, M. New Radiometric Ages for the BH-1 Hominin from Balanica (Serbia): Implications for Understanding the Role of the Balkans in Middle Pleistocene Human Evolution. PLoS ONE 2013, 8. [Google Scholar] [CrossRef]
- Socha, P. Rodent Palaeofaunas from Biśnik Cave (Kraków-Czestochowa Upland, Poland): Palaeoecological, Palaeoclimatic and Biostratigraphic Reconstruction. Quat. Int. 2014, 326–327, 64–81. [Google Scholar] [CrossRef]
- Nadachowski, A. Late Quaternary Rodents of Poland with Special Reference to Morphotype Dentition Analysis of Voles; Wydawnictwo Naukowe PWN: Warszawa and Kraków, Poland, 1982. [Google Scholar]
- Sabol, M.; Slyšková, D.; Bodoriková, S.; Čejka, T.; Čerňanský, A.; Ivanov, M.; Joniak, P.; Kováčová, M.; Tóth, C. Revised floral and faunal assemblages from Late Pleistocene deposits of the Gánovce-Hrádok Neanderthal site—Biostratigraphic and palaeoecological implications. Fossil Imprint 2017, 73, 182–196. [Google Scholar] [CrossRef]
- Ivanov, M.; Vöröš, D. Middle Pleistocene Voles and Lemmings (Rodentia: Arvicolinae) from Za Hájovnou Cave (Javoříčko Karst). Acta Musei Natl. Pragae, Ser. B Hist. Nat. 2014, 70, 43–54. [Google Scholar] [CrossRef]
- Pazonyi, P.; Kordos, L. Late Eemian (Late Pleistocene) Vertebrate Fauna from the Horváti-Lik (Uppony, NE Hungary). Fragm. Palaeontol. Hung. 2004, 22, 107–117. [Google Scholar]
- Pazonyi, P.; Kordos, L.; Magyari, E.; Marinova, E.; Fükoh, L.; Venczel, M. Pleistocene Vertebrate Faunas of the Sütto Travertine Complex (Hungary). Quat. Int. 2014, 319, 50–63. [Google Scholar] [CrossRef]
- Lundberg, J.; Musil, R.; Sabol, M. Sedimentary History of Za Hájovnou Cave (Moravia, Czech Republic): A Unique Middle Pleistocene Palaeontological Site. Quat. Int. 2014, 339–340, 11–24. [Google Scholar] [CrossRef]
- Wagner, J. The Updated Record of Cave Bear and Other Members of the Genus Ursus on the Territory of Czech Republic (Registro Actualizado de Oso de Las Cavernas y Otras Especies Del Género Ursus En La República Checa). Cad. Lab. Xeolóxico Laxe Coruña. 2001, 26, 447–455. [Google Scholar]
- Miracle, P.T.; Mauch Lenardić, J.; Brajković, D. Last Glacial Climates, “Refugia”, and Faunal Change in Southeastern Europe: Mammalian Assemblages from Veternica, Velika Pećina, and Vindija Caves (Croatia). Quat. Int. 2010, 212, 137–148. [Google Scholar] [CrossRef]
- Musil, R. The Environment of the Middle Palaeolithic Sites in Central and Eastern Europe. Stud. Archaeol. 2010, 3207, 121–179. [Google Scholar]
- Döppes, D.; Kempe, S.; Rosendahl, W. Dated Paleontological Cave Sites of Central Europe from Late Middle Pleistocene to Early Upper Pleistocene (OIS 5 to OIS 8). Quat. Int. 2008, 187, 97–104. [Google Scholar] [CrossRef]
- Berto, C.; Rubinato, G. The Upper Pleistocene Mammal Record from Caverna Degli Orsi (San Dorligo Della Valle—Dolina, Trieste, Italy): A Faunal Complex between Eastern and Western Europe. Quat. Int. 2013, 284, 7–14. [Google Scholar] [CrossRef]
- Van Kolfschoten, T. The Eemian Mammal Fauna of Central Europe. Geol. Mijnbouw/Neth. J. Geosci. 2000, 79, 269–281. [Google Scholar] [CrossRef]
- Gaudzinski, S. A Matter of High Resolution? The Eemian Interglacial (OIS 5e) in North-Central Europe and Middle Palaeolithic Subsistence. Int. J. Osteoarchaeol. 2004, 14, 201–211. [Google Scholar] [CrossRef]
- Strahl, J.; Krbetschek, M.R.; Luckert, J.; Machalett, B.; Meng, S.; Oches, E.A.; Rappsilber, I.; Wansa, S.; Zöller, L. Geologie, Paläontologie Und Geochronologie Des Eem-Beckens Neumark-Nord 2 Und Vergleich Mit Dem Becken Neumark-Nord 1 (Geiseltal, Sachsen-Anhalt). Eiszeitalter Gegenwart (Quat. Sci. J.) 2010, 59, 120–167. [Google Scholar] [CrossRef]
- Bratlund, B. Taubach Revisited. Jahrb. Römisch-Germanischen Zentralmuseums 1999, 46, 61–174. [Google Scholar]
- Blackwell, B.; Schwarcz, H.P. U-Series Analyses of the Lower Travertine at Ehringsdorf, DDR. Quat. Res. 1986, 25, 215–222. [Google Scholar] [CrossRef]
- Schäfer, D.; Heinrich, W.D.; Böhme, G.; Steiner, W. Aspects of the Geology, Palaeontology and Archaeology of the Travertine Site of Weimar-Ehringsdorf (Thuringia, Central Germany). Cour. Forsch.-Inst. Senckenberg 2007, 259, 141–147. [Google Scholar]
- Frank, N.; Braum, M.; Hambach, U.; Mangini, A.; Wagner, G. Warm Period Growth of Travertine during the Last Interglaciation in Southern Germany. Quat. Res. 2000, 54, 38–48. [Google Scholar] [CrossRef]
- Conard, N.J.; Prindiville, T.J.; Adler, D.S. Refitting Bones and Stones as a Means of Reconstructing Middle Paleolithic Subsistence in the Rhineland. In Proceedings of the Economie préhistorique: Les Comportements de subsistance au Paléolithique: Actes des Rencontres, Antibes, France, 23–25 October 1997; Brugal, J-P., Meignen, L., Patou-Mathis, M., Eds.; APDCA: Sophia Antipolis, France, 1998; pp. 273–290. [Google Scholar]
- Kolfschoten, T.; van Conard, N.J.; Preuss, J.; Langohr, R.; Haesaerts, P.; Forrest, D.T.; Kaszas, P.J.; Becze-Deak, J.; Rebholz, A. New Archaeological and Geological Research at the Palaeolithic Locality of Wallertheim in Rheinhessen. Archäologisches Korrespondenzblatt 1994, 24, 1–14. [Google Scholar]
- Bonjean, D.; di Modica, K.; Abrams, G.; Pirson, S.; Otte, M. Le Paléolithique moyen en Belgique. Bull. Cherch. Wallonie 2011, 17/4, 323–334. [Google Scholar] [CrossRef]
- Cordy, J.-M. Bio- et chronostratigraphie des dépôts quaternaires de la grotte Scladina (Province de Namur, Belgique) à partir des micromammifères. Et. et rech. arch. de l’Univ. de Liège. 1992, 27, 79–125. [Google Scholar]
- Germonpré, M. Mammoth Taphonomy of Two Fluvial Sites from the Flemish Valley, Belgium. Deinsea 2003, 9, 171–183. [Google Scholar]
- Van Neer, W. La Faune Saalienne Du Site Paléolithique Moyen de Mesvin IV (Hainaut, Belgique). Suppl. Bull. Ass. Fr. Et. Quat. 1986, 26, 103–111. [Google Scholar]
- Cahen, D.; Michel, J. Le Site Paléolithique Moyen Ancien de Mesvin IV (Hainaut, Belgique). Suppl. Bull. Ass. Fr. Et. Quat. 1986, 26, 89–102. [Google Scholar]
- Antoine, P.; Limondin-Lozouet, N.; Auguste, P.; Locht, J.-L.; Galheb, B.; Reyss, J.-L.; Escude, É.; Carbonel, P.; Mercier, N.; Bahain, J.-J.; et al. Le Tuf de Caours (Somme, France): Mise En Évidence d’une Séquence Eemienne et d’un Site Paléolithique Associé. Quaternaire 2006, 17, 281–320. [Google Scholar] [CrossRef]
- Mariani-Costantini, R.; Ottini, L.; Caramiello, S.; Palmirotta, R.; Mallegni, F.; Rossi, A.; Frati, L.; Capasso, L. Taphonomy of the Fossil Hominid Bones from the Acheulean Site of Castel Di Guido near Rome, Italy. J. Hum. Evol. 2001, 41, 211–225. [Google Scholar] [CrossRef] [PubMed]
- Michel, V.; Boschian, G.; Valensi, P. Datation ESR de Dents d’aurochs Du Site Paléolithique Inférieur de Castel Di Guido (Italie). Archeosciences 2008, 32, 51–58. [Google Scholar] [CrossRef]
- Ronchitelli, A.; Boscato, P.; Surdi, G.; Masini, F.; Petruso, D.; Accorsi, C.A.; Torri, P. The Grotta Grande of Scario (Salerno, Italy): Archaeology and Environment during the Last Interglacial (MIS 5) of the Mediterranean Region. Quat. Int. 2011, 231, 95–109. [Google Scholar] [CrossRef]
- Petronio, C.; Bellardini, F.; Arzarello, M.; Bedetti, C.; Bellucci, L.; Cipullo, A.; Di Stefano, G.; Pandolfi, L.; Pavia, M.; Petrucci, M.; et al. The Deposit of the Late Pleistocene from Avetrana (Taranto, Southern Italy): Biocrhonology and Palaeoecology. Quat. Ital. J. Quat. Sci. 2008, 21, 409–422. [Google Scholar]
- Pandolfi, L.; Mannino, M.A.; Talamo, S.; Salari, L.; Sansò, P.; Saponetti, S.S.; Vacca, E.; Vicari, D.; Richards, M.P.; Petronio, C. A Reassessment of the Infills and Faunal Assemblages of Karst Cavities Known as Ventarole in Salento (Apulia, Southern Italy): A Multidisciplinary Investigation on Cava Donno (Corigliano d’Otranto, Lecce). Alp. Mediterr. Quat. 2017, 30, 25–40. [Google Scholar]
- Bologna, P.; Di Stefano, G.; Manzi, G.; Petronio, C.; Sardella, R.; Squazzini, E. Late Pleistocene Mammals from the Melpignano (LE) «Ventarole»: Preliminary Analysis and Correlations. Bull. Soc. Paleontol. Ital. 1994, 33, 265–274. [Google Scholar]
- Houben, C. Die Wirbeltierfauna Aus Dem Letzten Interglazial von Lehringen (Niedersachsen, Deutschland). Eiszeitalter Gegenwart (Quat. Sci. J.) 2003, 52, 25–39. [Google Scholar]
- De Giuli, C. Le Faune Pelistoceniche Del Salento. 1. La Fauna Di San Sidero 3. Quad. Mus. Paleontol. Maglie 1983, I, 45–84. [Google Scholar]
- Melis, R.T.; Ghaleb, B.; Boldrini, R.; Palombo, M.R. The Grotta Dei Fiori (Sardinia, Italy) Stratigraphical Successions: A Key for Inferring Palaeoenvironment Evolution and Updating the Biochronology of the Pleistocene Mammalian Fauna from Sardinia. Quat. Int. 2013, 288, 81–96. [Google Scholar] [CrossRef]
- Chesi, F.; Delfino, M.; Abbazzi, L.; Carboni, S.; Lecca, L.; Rook, L. New Fossil vertebrate Remains from San Giovanni Di Sinis (Late Pleistocene, Sardinia): The Last Mauremys (Reptilia, Testudines) in the Central Mediterranean. Riv. Ital. di Paleontol. e Stratigr 2007, 113, 287–297. [Google Scholar]
- Malatesta, A. On the Genus Cyrnaonyx Helbing, 1935 (Mustelidae, Lutrinae) and Its Type Species. Quat. XX 1978, 20, 109–116. [Google Scholar]
- Shen, G. U-Series Dating of the Deposits from the Prince Cave, Northern Italy. Archaeometry 1986, 28, 179–184. [Google Scholar]
- Valensi, P.; Psathi, E. Faunal Exploitation during the Middle Palaeolithic in South-Eastern France and North-Western Italy. Int. J. Osteoarchaeol. 2004, 14, 256–272. [Google Scholar] [CrossRef]
- Valensi, P.; Abbassi, M. Quaternaire Reconstitution de Paléoenvironnements Quaternaires Par l’utilisation de Diverses Méthodes Sur Une Communauté de Mammifères—Application à La Grotte Du Lazaret. Quaternaire 1998, 9, 291–302. [Google Scholar] [CrossRef]
- Valladas, H.; Mercier, N.; Falguères, C.; Bahain, J. Contribution Des Méthodes Nucléaires à La Chronologie Des Cultures Paléolithiques Entre 300 000 et 35 000 Ans BP. Gall. Préhistoire 1999, 41, 153–166. [Google Scholar] [CrossRef]
- Defleur, A.; Crégut-Bonnoure, E.; Radulescu, C.; Villette, P. La Grotte Des Cèdres, Commune Du Plan d’Aups (Var). Bull. Société Préhistorique Française 1990, 87, 270–274. [Google Scholar] [CrossRef]
- Valensi, P.; Crégut-Bonnoure, E.; Defleur, A. Archaeozoological Data from the Mousterian Level from Moula-Guercy (Ardèche, France) Bearing Cannibalised Neanderthal Remains. Quat. Int. 2012, 252, 48–55. [Google Scholar] [CrossRef]
- Foury, Y.; Desclaux, E.; Daujeard, C.; Defleur, A.; Moncel, M.-H.; Raynal, J.-P. Evolution Des Faunes de Rongeurs En Moyenne Vallée Du Rhône (Rive Droite, Ardèche, France) Au Cours Du Pléistocène Moyen Final et Du Pléistocène Supérieur Ancien, Du Mis 6 Au Mis 4. Quaternaire 2016, 27, 55–79. [Google Scholar] [CrossRef]
- Slimak, L.; Lewis, J.E.; Crégut-Bonnoure, E.; Metz, L.; Ollivier, V.; André, P.; Chrzavzez, J.; Giraud, Y.; Jeannet, M.; Magnin, F. Le Grand Abri Aux Puces, a Mousterian Site from the Last Interglacial: Paleogeography, Paleoenvironment, and New Excavation Results. J. Archaeol. Sci. 2010, 37, 2747–2761. [Google Scholar] [CrossRef]
- Crégut-Bonnoure, E.; Boulbes, N.; Daujeard, C.; Fernandez, P.; Valensi, P. Nouvelles Données Sur La Grande Faune de l’Éemien Dans Le Sud-Est de La France. Quaternaire 2010, 21, 227–248. [Google Scholar] [CrossRef]
- Michel, V.; Shen, G.; Shen, C.C.; Wu, C.C.; Vérati, C.; Gallet, S.; Moncel, M.H.; Combier, J.; Khatib, S.; Manetti, M. Application of U/Th And40Ar/39Ar Dating to Orgnac 3, a Late Acheulean and Early Middle Palaeolithic Site in Ardèche, France. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [PubMed]
- Moncel, M.-H. Les Niveaux Profonds Du Site Pléistocène Moyen d’Orgnac 3 (Ardèche, France): Habitat, Repaire, Aven-Piège? L’exemple Du Niveau 6. Bull. Soc. Préhistorique Française 1996, 93, 470–481. [Google Scholar] [CrossRef]
- Moncel, M.H. Baume Flandin et Abri Du Maras: Deux Exemples de Débitage Laminaire Du Début Du Pléistocène Supérieur Dans La Vallée Du Rhône (Sud-Est, France). Anthropologie 2005, 109, 451–480. [Google Scholar] [CrossRef]
- Moncel, M.H.; Crégut-Bonnoure, É.; Daujeard, C.; Lartigot, A.S.; Lebon, M.; Puaud, S.; Boulbes, N.; Croizet, S. Le Site de La Baume Flandin (Commune d’Orgnac-l’Aven): Nouvelles Données Sur Ce Gisement Du Paléolithique Moyen. C. R. Palevol 2008, 7, 315–325. [Google Scholar] [CrossRef]
- Moncel, M.H. L’abri Des Pêcheurs (Ardèche, Sud-Est France). Des Occupations Néandertaliennes Dans Une «grotte En Forme de Fissure ». Ann. d’Universite Valahia Targoviste, Sect. d’Archeologie d’Histoire 2010, 12, 7–21. [Google Scholar]
- Masaoudi, H.; Falgueres, C.; Bahain, J.J.; Yokoyama, Y.; Lhomme, G. Datation d’ossements et de Planchers Stalagmitiques Provenant de l’Abri Des Pecheurs (Ardeche) Par La Methode Des Desequilibres Des Familles de l’uranium (U-Th). Quaternaire 1994, 5, 79–83. [Google Scholar] [CrossRef]
- Yokoyama, Y.; Nguyen, H.V.; Shen, G.; Quaegebeur, J.P.; Koumba, N. Datation par les méthodes des familles de l'uranium et la résonance de spin électronique (ESR) du remplissage de la grotte des Ramandils à Port-la-Nouvelle (Aude, France). Cypsela VI 1987, 151–154. [Google Scholar]
- Gerber, J.P. Le Site Des Ramandils Une Faune Du Début Du Würmien II En Languedoc-Méditerranéen. Bull. l’Association Française Pour L’étude du Quat 1973, 10, 7–12. [Google Scholar] [CrossRef]
- Daura, J.; Sanz, M.; Julià, R.; García-Fernández, D.; Fornós, J.J.; Vaquero, M.; Allué, E.; López-García, J.M.; Blain, H.A.; Ortiz, J.E.; et al. Cova Del Rinoceront (Castelldefels, Barcelona): A Terrestrial Record for the Last Interglacial Period (MIS 5) in the Mediterranean Coast of the Iberian Peninsula. Quat. Sci. Rev. 2015, 114, 203–227. [Google Scholar] [CrossRef]
- Marquet, J.-C.; Vezian, R.; Gardeisen, A. Le Portel-Ouest. Associations Fauniques et Paléoenvironnements Sur La Frange Septentrionale Des Pyrénées Ariégeoises Au Würm Ancien. Quaternaire 1998, 9, 303–314. [Google Scholar] [CrossRef]
- Bahain, J.J.; Laurent, M.; Falguères, C.; Voinchet, P.; Farkh, S.; Tissoux, H. Datation Par Résonnance Paramagnétique Électronique (RPE) Des Formations Fluviatiles Pléistocènes et Des Gisements Archéologiques Ou Paléontologiques Associés. Quaternaire 2002, 13, 91–103. [Google Scholar] [CrossRef]
- David, P.; Prat, F. Considérations Sur Les Faunes de La Chaise (Commune de Vouthon, Charente). Abris Suard et Bourgeois-Delaunay. Bull. L’association Française Pour L’étude Quat. 1965, 2, 222–231. [Google Scholar] [CrossRef]
- Jaubert, J.; Kervazo, B.; Bahain, J.-J.; Brugal, J.-P.; Chalard, P.; Falguères, C.; Jarry, M.; Jeannet, M.; Lemorini, C.; Louchart, A.; et al. Coudoulous I (Tour-deFaure, Lot), site du Pléistocène moyen en Quercy: Bilan pluridisciplinaire. In Données récentes sur les peuplements du Paléolithique inférieur et moyen en Europe; Molines, N., Monnier, J.-L., Moncel, M.-H., Eds.; Actes du Colloque de Rennes, Université de Rennes: Rennes, France, 2005; Volume 1364, pp. 227–251. [Google Scholar]
- Brugal, J.-P.; Jaubert, J. Les Gisements Paléontologiques Pléistocènes à Indices de Fréquentation Humaine: Un Nouveau Type de Comportement de Prédation? Paléo 1991, 3, 15–41. [Google Scholar] [CrossRef]
- Grayson, D.K.; Delpech, F. The Evidence for Middle Palaeolithic Scavenging from Couche VIII, Grotte Vaufrey (Dordogne, France). J. Archaeol. Sci. 1994, 21, 359–375. [Google Scholar] [CrossRef]
- Hoffmann, D.; Mangini, A. A Method for Coupled ESR/U-Series Dating of Teeth Showing Post-Depositional U-Loss. Quat. Sci. Rev. 2003, 22, 1367–1372. [Google Scholar] [CrossRef]
- Costamago, S.; Vigier, S.; Astruc, J.-G.; Drieux, M. Le Gisement Paleotologique de Bramenfond (Autoroute A20, Section 4, Brive-Souillac, Souillac, Lot). Bull. Société Préhistorique Française 1998, 95, 423–435. [Google Scholar]
- Sanchis, A.; Fernández Peris, J. Procesado y Consumo Antrópico de Conejo En La Cova de Bolomor (Tavernes de La Valldigna, Valencia): El Nivel XVIIc (ca 350 Ka). Complutum 2008, 19, 25–46. [Google Scholar]
- Blasco, R.; Fernández Peris, J. A Uniquely Broad Spectrum Diet during the Middle Pleistocene at Bolomor Cave (Valencia, Spain). Quat. Int. 2012, 252, 16–31. [Google Scholar] [CrossRef]
- Pérez, M.; Vicente, J.; Serra, A.S.; Margalef, C.R.; Ripoll, M.P.; Emili, J.; Tortosa, A.; Bonilla, V.V. Evidences of Interaction Homo-Cuon in Three Upper Pleistocene Sites of the Iberian Mediterranean Central Region. J. Taphon. 2012, 10, 3–4. [Google Scholar]
- Walker, M.J.; Gibert, J.; Sánchez, F.; Lombardi, A.V.; Serrano, I.; Gómez, A.; Eastham, A.; Ribot, F.; Arribas, A.; Cuenca, A.; et al. Excavations at New Sites of Early Man in Murcia: Sima de Las Palomas Del Cabezo Gordo and Cueva Negra Del Estrecho Del Río Quípar de La Encarnación. Hum. Evol. 1999, 14, 99–123. [Google Scholar] [CrossRef]
- Lopez-Garcia, J.M. Evolución de La Diversidad Taxonómica de Los Micromamíferos En La Península Ibérica y Cambios Paleoambientales Durante El Pleistoceno Superior. Ph.D. Thesis, Universitat Rovira i Virgili, Tarragona, Spain, April 2008. [Google Scholar]
- Barroso Ruíz, C.; Botella Ortega, D.; Caparrós, M.; Moigne, A.M.; Celiberti, V.; Testu, A.; Barsky, D.; Notter, O.; Riquelme Cantal, J.A.; Rodríguez, M.P.; et al. The Cueva Del Angel (Lucena, Spain): An Acheulean Hunters Habitat in the South of the Iberian Peninsula. Quat. Int. 2011, 243, 105–126. [Google Scholar] [CrossRef]
- Falguères, C.; Bahain, J.J.; Pérez-González, A.; Mercier, N.; Santonja, M.; Dolo, J.M. The Lower Acheulian Site of Ambrona, Soria (Spain): Ages Derived from a Combined ESR/U-Series Model. J. Archaeol. Sci. 2006, 33, 149–157. [Google Scholar] [CrossRef]
- Soto, E.; Sesé, C.; Pérez-González, A.; Santonja, M. Mammal Fauna with Elephas (Palaeoloxodon) antiquus from the Lower Levels of Ambrona (Soria, Spain). In Proceedings of the 1st International Congress: The World of Elephants. Workshop: Stone tools and elephant remains in the Lower and Middle Palaeolithic of the Iberian Peninsula, Rome, Italy, October 2001; CNR: Rome, Italy, 2001; pp. 607–610. [Google Scholar]
- De Lombera-Hermida, A.; Rodríguez, X.P.; Fábregas-Valcarce, R. Cova Eirós Archaeo-Palaeontological Site, Triacastela, Lugo. In Pleistocene and Holocene Hunter-Gatherers in Iberia and the Gibraltar Strait the Current Archaeological Record; Universidad de Burgos, Fundación Atapuerca: Burgos, Spain, 2014; pp. 18–25. [Google Scholar]
- Rodríguez Álvarez, X.P.; de Lombera Hermida, A.; Fábregas Valcarce, R.; Lazuén Fernández, T. The Upper Pleistocene Site of Cova Eirós (Triacastela, Lugo, Galicia). In To the West of Spanish Cantabria: The Palaeolithic Settlement of Galicia; de Lombera Hermida, A., Fábregas Valcarce, R., Eds.; Archaeopress: Oxford, UK, 2011; pp. 123–132. ISBN 9781407308609. [Google Scholar]
- Sesé, C.; López Martínez, N. Nuevos Datos Paleontológicos Del Pleistoceno En El Valle Del Manzanares (Madrid, España): Los Micromamíferos Del Yacimiento Del Arenero de Arriaga. Estud. Geol. 2013, 69, 271–282. [Google Scholar] [CrossRef]
- Panera, J.; Rubio-Jara, S.; Yravedra, J.; Blain, H.A.; Sesé, C.; Pérez-González, A. Manzanares Valley (Madrid, Spain): A Good Country for Proboscideans and Neanderthals. Quat. Int. 2014, 326–327, 329–343. [Google Scholar] [CrossRef]
- Arribas, A. El Yacimiento Mesopleistoceno de Villacastin (Segovia, Espana): Geologia y Paleontologia de Micromamiferos. Bol. Geol. Min. 1994, 105, 146–166. [Google Scholar]
- Arribas, A. Consideraciones Cronológicas, Tafonómicas y Paleoecológicas Del Yacimiento Cuaternario de Villacastín (Segovia, España). Bol. Geol. Min. 1995, 106, 3–22. [Google Scholar]
- Coxon, P.; Hall, A.R.; Lister, A.; Stuart, A.J. New Evidence on the Vertebrate Fauna, Stratigraphy and Palaeobotany of the Interglacial Deposits at Swanton Morley, Norfolk. Geol. Mag. 1980, 117, 525–546. [Google Scholar] [CrossRef]
- Stuart, A.J. Insularity and Quaternary Vertebrate Faunas in Britain and Ireland. Geol. Soc. Lond. Spec. Publ. 1995, 96, 111–125. [Google Scholar] [CrossRef]
- Gao, C.; Keen, D.H.; Boreham, S.; Coope, G.R.; Pettit, M.E.; Stuart, A.J.; Gibbard, P.L. Last Interglacial and Devensian Deposits of the River Great Ouse at Woolpack Farm, Fenstanton, Cambridgeshire, UK. Quat. Sci. Rev. 2000, 19, 787–810. [Google Scholar] [CrossRef]
- Gilmour, M.; Currant, A.; Jacobi, R.; Stringer, C. Recent TIMS Dating Results from British Late Pleistocene Vertebrate Faunal Localities: Context and Interpretation. J. Quat. Sci. 2007, 22, 793–800. [Google Scholar] [CrossRef]
- Stuart, A.J. Pleistocene Vertebrates in the British Isles; Longman Group (Far East), Limited: London, UK; New York, NY, USA, 1982. [Google Scholar]
- Currant, A.; Jacobi, R. A Formal Mammalian Biostratigraphy for the Late Pleistocene of Britain. Quat. Sci. Rev. 2001, 20, 1707–1716. [Google Scholar] [CrossRef]
- Sutcliffe, A. Joint Mitnor Cave, Buckfastleigh. Trans. Torquay Nat. Hist. Soc. 1960, 13, 1–26. [Google Scholar]
- Stringer, C.B.; Currant, A.P.; Schwarcz, H.P.; Collcutt, S.N. Age of Pleistocene Faunas from Bacon Hole, Wales. Nature 1986, 320, 59–62. [Google Scholar] [CrossRef]
- Bateman, M.D.; Catt, J.A. An Absolute Chronology for the Raised Beach and Associated Deposits at Sewerby, East Yorkshire, England. J. Quat. Sci. 1996, 11, 389–395. [Google Scholar] [CrossRef]
- Boylan, P.J. The Pleistocene Mammalia of the Sewerby-Hessle Buried Cliff, East Yorkshire. Proc. Yorksh. Geol. Soc. 1967, 36, 115–125. [Google Scholar] [CrossRef]
- McFarlane, D.A.; Ford, D.C. The Age of the Kirkdale Cave Palaeofauna. Cave Karst Sci. 1998, 25, 3–6. [Google Scholar]
- Boylan, P.J. A New Revision of the Pleistocene Mammalian Fauna of Kirkdale Cave, Yorkshire. Proc. Yorksh. Geol. Soc. 1981, 43, 253–280. [Google Scholar] [CrossRef]
- Gascoyne, M.; Currant, A.P.; Lord, T.C. Ipswichian Fauna of Victoria Cave and the Marine Palaeoclimatic Record. Nature 1981, 294, 652–654. [Google Scholar] [CrossRef]
- Manly, B.F.J. Randomization, Bootstrap and Monte Carlo Methods in Biology; Chapman and Hall/CRC: London, UK, 2007. [Google Scholar]
- Hammer, Ø.; Harper, D.A.T.A.T.; Ryan, P.D. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar] [CrossRef]
- Gromov, V.I. Paleontological and Archaeological Substantiation of the Stratigraphy of Continental Deposits of the Quaternary Period in the Territory of the USSR. Proc. Geol. Inst. USSR Acad. Sci. 1948, 17, 1–521. (In Russian) [Google Scholar]
- Alexandrova, L.P. Rodents of the Anthropogene of the European Part of the USSR. Proc. Geol. Inst. USSR Acad. Sci. 1976, 291, 1–96. (In Russian) [Google Scholar]
- Zastrozhnov, A.; Danukalova, G.; Murray, A. New Data on the Age of the Neopleistocene Deposits of the Lower Volga Sites According to the OSL Method Results. In Quaternary: The Results of Investigation and the Main Directions in the Future; GEOS: Moscow, Russia, 2017; pp. 139–140. [Google Scholar]
- Kirillova, I.V.; Tesakov, A.S. Water Vole (Arvicola, Arvicolinae, Rodentia) from the Khozar Deposits of Lower Volga Basin. In Quaternary and Modern Ecology: Nature and Man. Volgograd-Astrakhan, September 24–27, 2004; Gumanistika Press: Volgograd, Russia, 2004; pp. 138–139. (In Russian) [Google Scholar]
- Markova, A.K. Pleistocene Rodents of the Russian Plain; Nauka: Moscow, Russia, 1982. (In Russian) [Google Scholar]
- Chabai, V.; Richter, J.; Uthmeier, T. (Eds.) Kabazi II: Last Intergacial Occupation, Environment and Subsistence; Simferopol-Cologne: Shlyakh, Ukraine, 2005. [Google Scholar]
- Markova, A.K. Pleistocene Microtheriofauna of Eastern Europe. In Stratigraphy and Paleogeography of Quaternary of Eastern Europe; Velichko, A.A., Shik, S.M., Eds.; Institute of Geography RAS: Moscow, Russia, 1992; pp. 50–94. (In Russian) [Google Scholar]
- Cyrek, K.; Socha, P.; Stefaniak, K.; Madeyska, T.; Mirosław-Grabowska, J.; Sudoł, M.; Czyźewski, Ł. Palaeolithic of Biśnik Cave (Southern Poland) within the Environmental Background. Quat. Int. 2010, 220, 5–30. [Google Scholar] [CrossRef]
- Smirnov, N.G. Small Mammals of the Middle Urals in the Late Pleistocene; Nauka: Ekaterinburg, Russia, 1993. [Google Scholar]
- Maul, L.C. The Quaternary Small Mammal Faunas of Thuringia (Central Germany). In The Quaternary of Central Germany (Thuringia and Surroundings). Field Guide; Meyrick, R.A., Schreveand, D.C., Eds.; London Quaternary Research Association: London, UK, 2002; pp. 79–95. [Google Scholar]
- Heinrich, W.-D.; Janossy, D. Insektivoren Und Rodentier Aus Dem Travertin von Burgtonna in Thüringen. Quartärpaläontologie 1977, 2, 401–411. [Google Scholar]
- Heinrich, W.-D.; Janossy, D. Insektivoren Und Rodentier Aus Dem Travertin von Burgtonna in Thüringen. Quartärpaläontologie 1978, 3, 167–170. [Google Scholar]
- Von Koenigswald, W.; Tobien, H. Important Arvicolid Faunas from the Late Pliocene to Early Holocene in Western Germany (FRG). In International Symposium Evolution, Phylogeny Biostratigraphy of Arvicolids; Fejfar, O., Heinrich, W.-D., Eds.; Pfeil: München, Germany, 1990; pp. 231–253. [Google Scholar]
- Grün, R.; Brunnacker, K.; Hennig, G. 230Th/234U-Daten Mittel- Und Jungpleistozäner Travertine im Raum Stuttgart. Jahresberichte Mitteilungen Oberrheinischen Geol. Vereins 1982, 64, 201–211. [Google Scholar] [CrossRef]
- Schmidt, E.D.; Frechen, M.; Murray, A.S.; Tsukamoto, S.; Bittmann, F. Luminescence Chronology of the Loess Record from the Tönchesberg Section: A Comparison of Using Quartz and Feldspar as Dosimeter to Extend the Age Range beyond the Eemian. Quat. Int. 2011, 234, 10–22. [Google Scholar] [CrossRef]
- Blain, H.-A.; López-García, J.M.; Cordy, J.-M.; Pirson, S.; Abrams, G.; Di Modica, K.; Bonjean, D. New Contribution to the Study of the Small-Vertebrate Assemblages of Scladina Cave (Late Pleistocene, Province of Namur, Belgium. In Book of Abstracts. Middle Paleolithic in North-West Europe: Multidisciplinary approaches, Namur, Moulins de Beez, Belgium, 20–21 March 2014; p. 50. [Google Scholar]
- Delpech, F. L’environnement Animal Des Moustériens Quina Du Périgord. Paléo 1996, 8, 31–46. [Google Scholar] [CrossRef]
- Valensi, P. The Archaeozoology of Lazaret Cave (Nice, France). Int. J. Osteoarchaeol. 2000, 10, 357–367. [Google Scholar] [CrossRef]
- Tissoux, H.; Vezian, R.; De Lumley, H.; Bahain, J.-J.; Falguères, C. Datation Par Les Séries de l’uranium (U-Th) et La Résonance Paramagnétique Électronique (R.P.E.) Combinées Du Gisement Paléolithique de La Grotte Du Portel-Ouest (Commune de Loubens, Ariège, France). Résultats Préliminaires. In BAR International, S1270, Acts of the XIVth UISPP Congress-Section 2—General Sessions and Posters; Archaeopress: Oxford, UK, 2004; pp. 1–8. [Google Scholar]
- Crégut-Bonnoure, É.; Boulbes, N.; Guérin, C.; Pernaud, J.; Tavoso, A.; Cammas, R. Le Contexte Géomorphologique et Faunique de l’homme de Montmaurin (Haute-Garonn). Préhistoires Méditerranéennes 2010, 1, 35–85. [Google Scholar]
- Richter, D.; Dibble, H.; Goldberg, P.; McPherron, S.P.; Niven, L.; Sandgathe, D.; Talamo, S.; Turq, A. The Late Middle Palaeolithic in Southwest France: New TL Dates for the Sequence of Pech de l’Azé IV. Quat. Int. 2013, 294, 160–167. [Google Scholar] [CrossRef]
- Hanquet, C. Evolution Des Paléoenvironnements et Des Paléoclimats Au Pléistocène Moyen, En Europe Méridionale, d’après Les Faunes de Micromammifères. ABES 2011. [Google Scholar] [CrossRef]
- Stuart, A.J. The History of the Mammal Fauna during the Ipswichian/Last Interglacial in England. Philos. Trans. R. Soc. B Biol. Sci. 1976, 276, 221–250. [Google Scholar] [CrossRef]
- Salari, L.; Sardella, R. The Pleistocene Porcupine Hystrix vinogradovi Argyropulo, 1941 in Italy. Boll. Della Soc. Paleontol. Ital. 2009, 48, 123–127. [Google Scholar]
- Palombo, M.R. The Plio-Pleistocene Terrestrial Mammals of Sardinia: Biochronology and Faunal Renewals. Hantkeniana 2006, 49–55. [Google Scholar]
- Fernández-García, M.; López-García, J.M.; Lorenzo, C. Palaeoecological Implications of Rodents as Proxies for the Late Pleistocene-Holocene Environmental and Climatic Changes in Northeastern Iberia. C. R. Palevol. 2016, 15, 707–719. [Google Scholar] [CrossRef]
- Díez, C.; García, M.A.; Gil, E.; Jordá Pardo, J.F.; Ortega, A.; Sánchez, A.; Sánchez, B. La Cueva de Valdegoba (Burgos). Primera Campaña de Excavaciones. Zephyrus Rev. Prehist. Arqueol. 1988, 41–42, 55–74. [Google Scholar]
- Bezerra, A.M.R.; Annesi, F.; Aloise, G.; Amori, G.; Giustini, L.; Castiglia, R. Integrative Taxonomy of the Italian Pine Voles, Microtus savii Group (Cricetidae, Arvicolinae). Zool. Scr. 2016, 45, 225–236. [Google Scholar] [CrossRef]
- Puzachenko, A.Y.; Markova, A.K. Space-Time Dynamycs of European Mammal Diversity (Late Pleistocene-Holocene). In Evolution of European Ecosystems during Pleistocene-Holocene Transition (24–8 kyr BP); Markova, A.K., van Kolfschoten, T., Eds.; KMK Scientific Press: Moscow, Russia, 2008; pp. 275–298. ISBN 978-5-87317-456-0. [Google Scholar]
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).