Culture Medium Optimization for Production of Rhamnolipids by Burkholderia glumae



Abstract
1. Introduction
2. Materials and Methods
2.1. Microorganism and Inoculum Preparation
2.2. Quantification of Rhamnolipids and Mannitol
2.3. Biomass Measurement
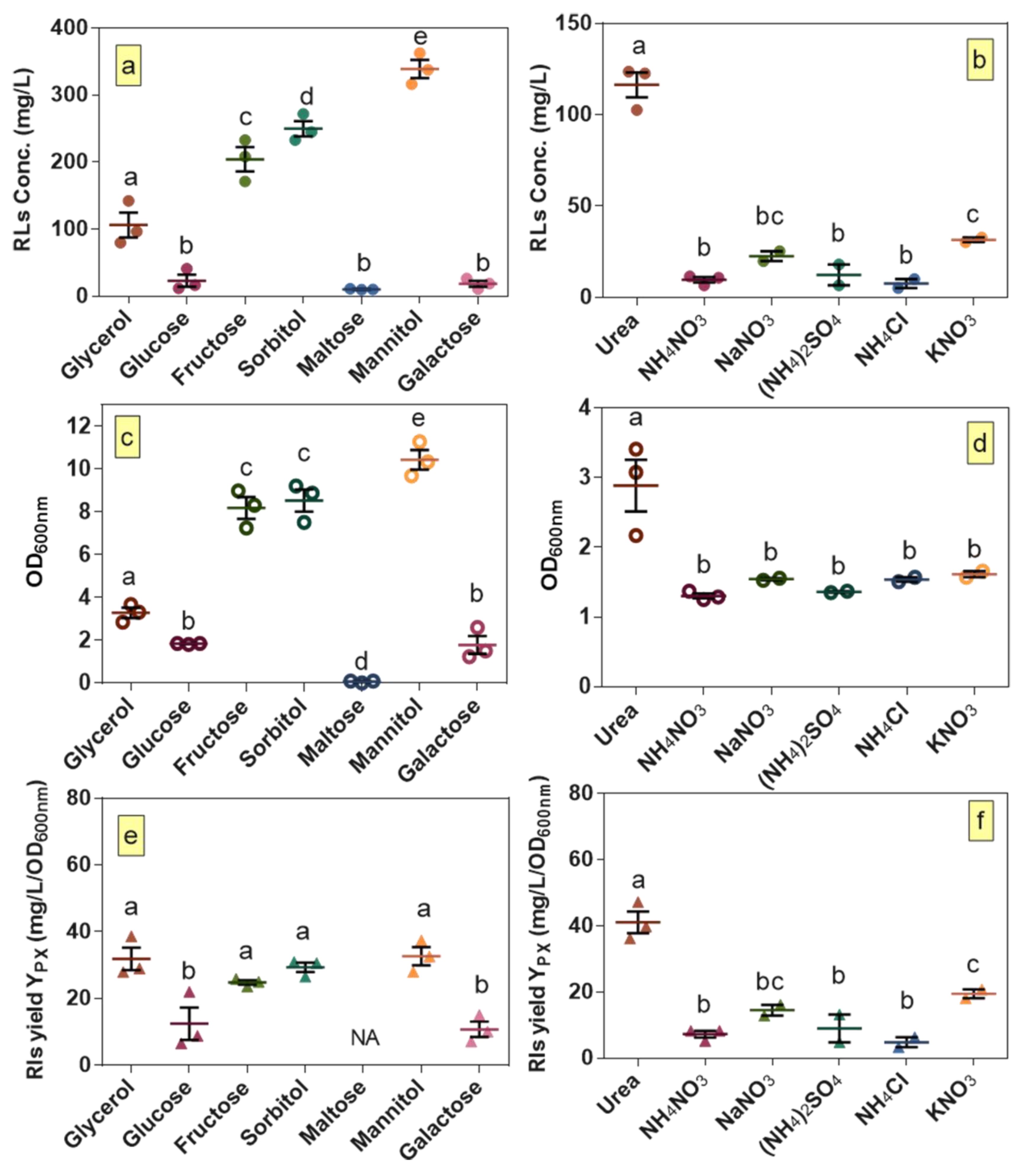
2.4. Selection of Optimal Carbon and Nitrogen Sources
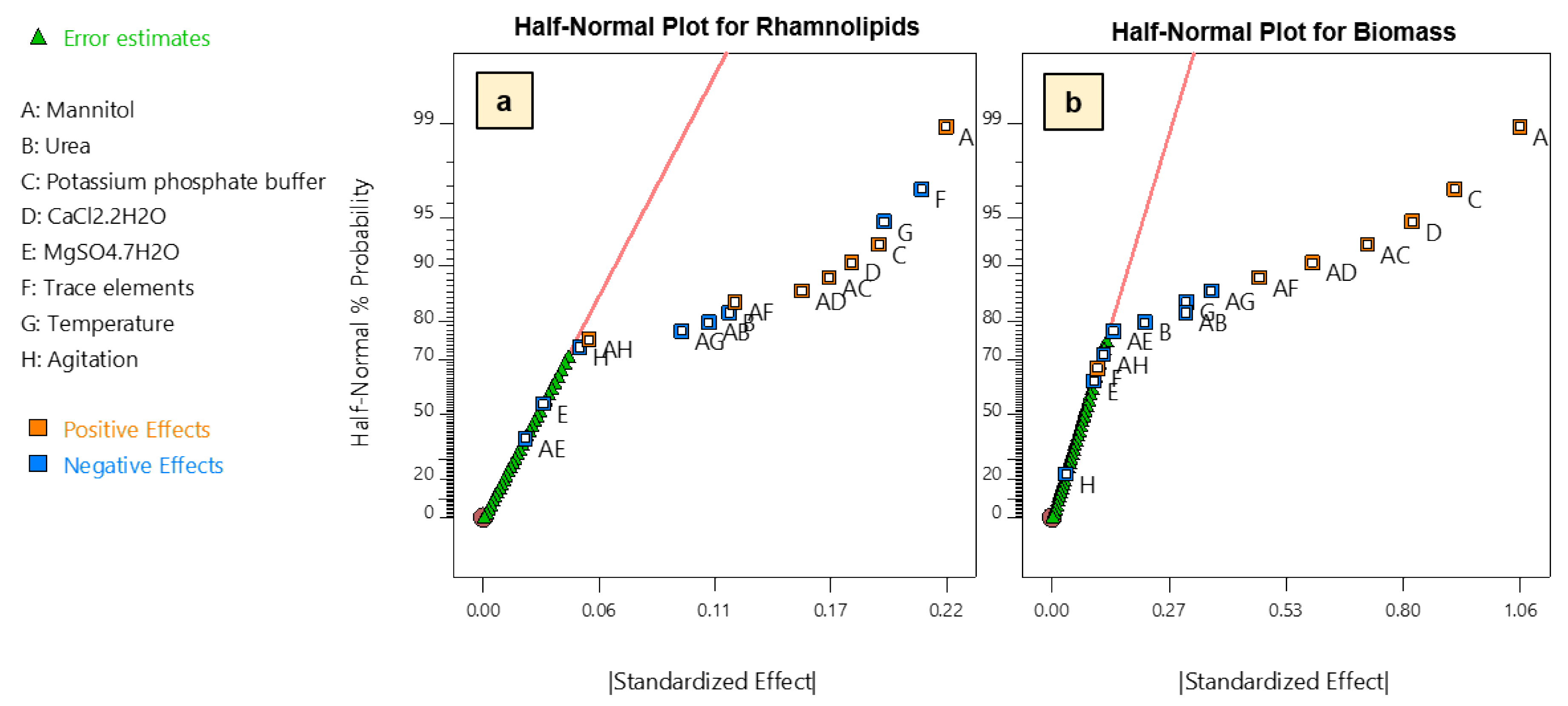
2.5. Fractional Factorial Design (FFD)
2.6. Response Surface Methodology (RSM)
2.7. Design Matrix and Data Analysis
2.8. Optimization and Model Verification
3. Results and Discussion
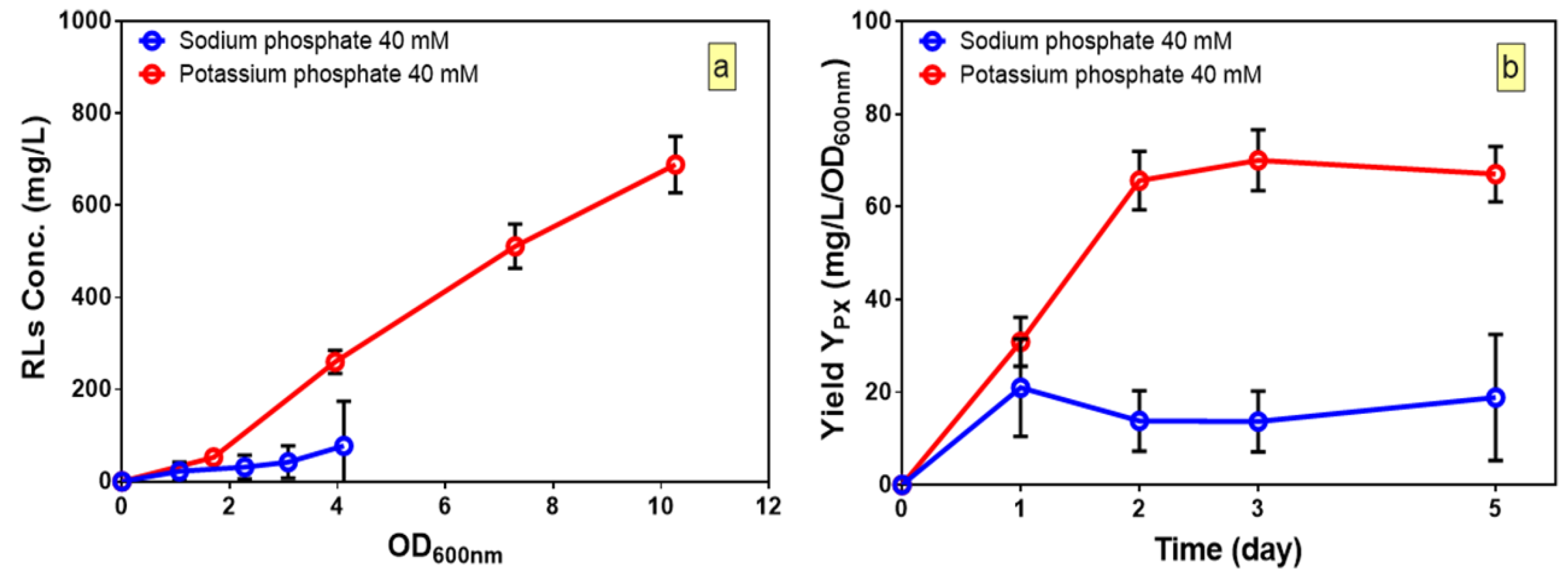
3.1. Selection of Optimal Carbon and Nitrogen Sources
3.2. Selection of Significant Parameters by Fractional Factorial Design
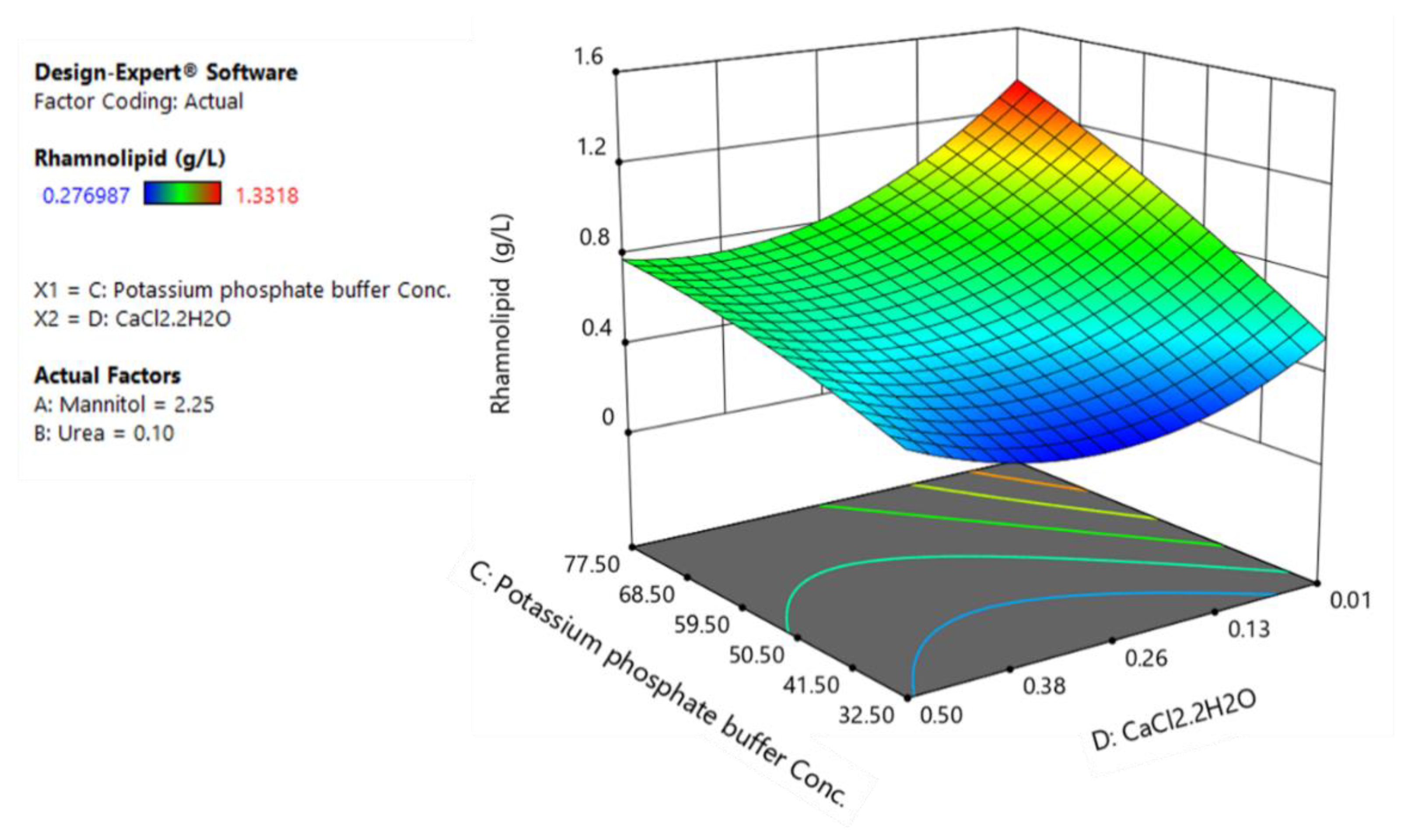
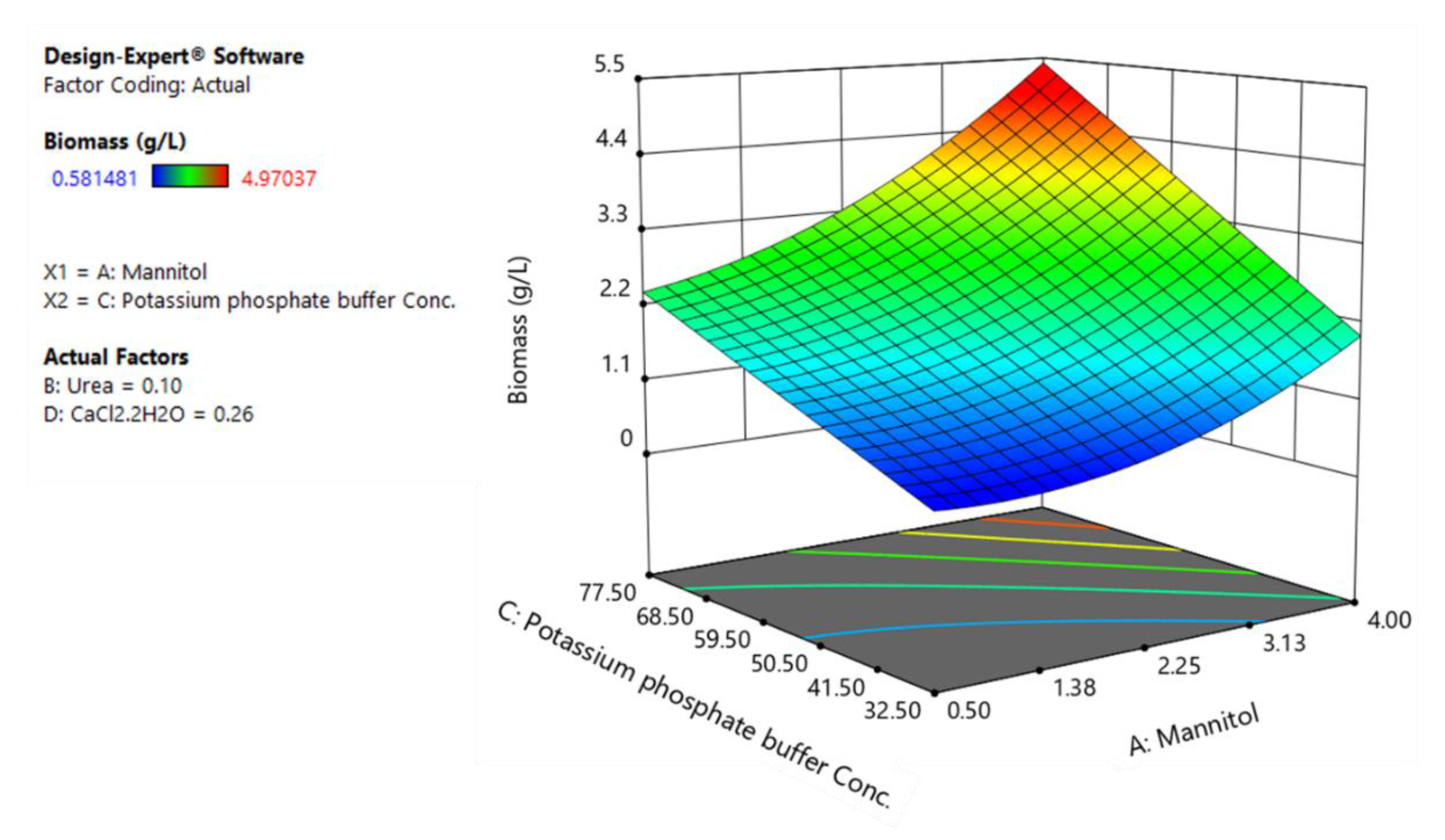
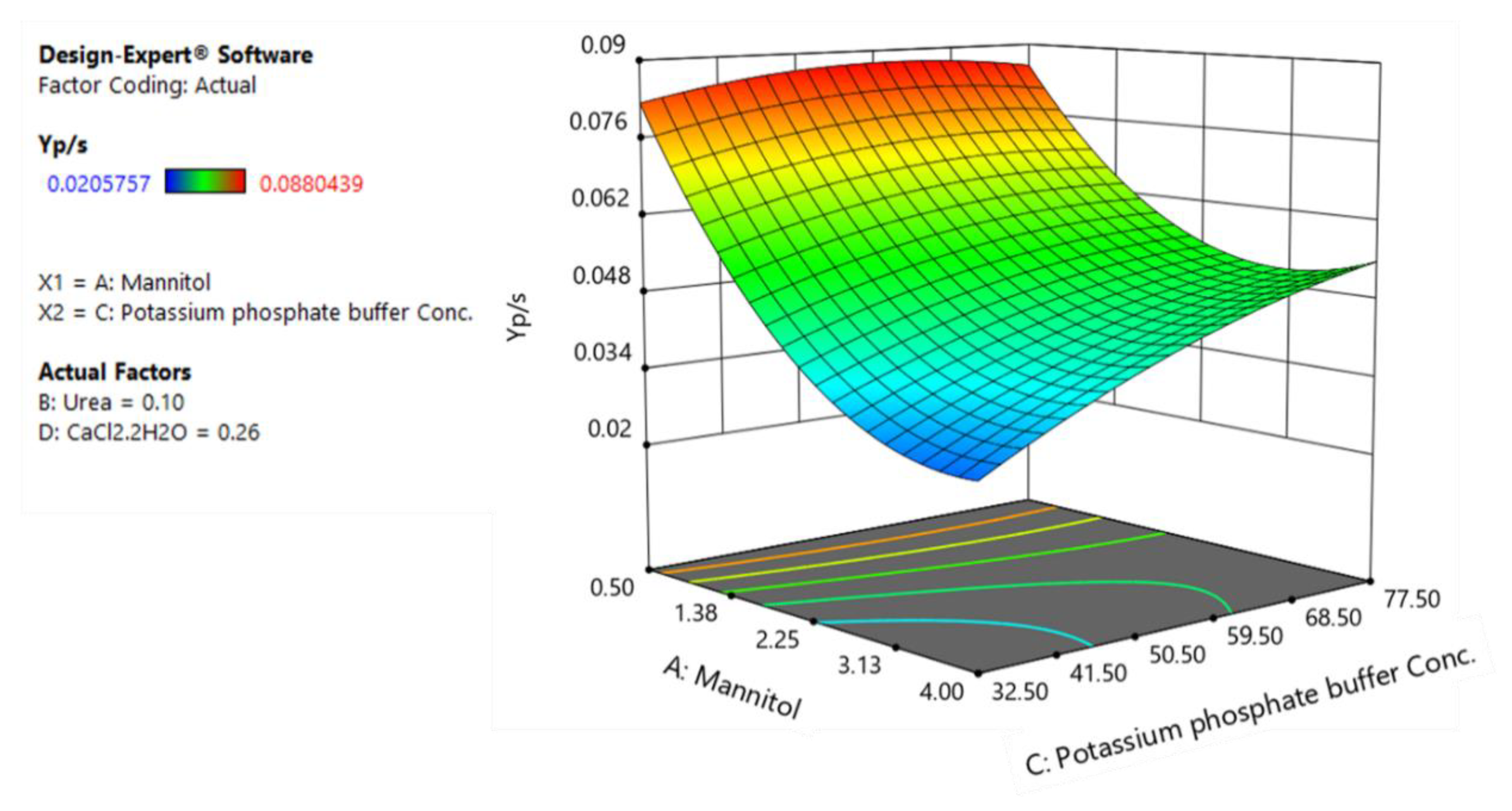
3.3. CCD and Fitted Regression Models for Production of Rhamnolipids by B. glumae BGR1
3.4. Optimization and Model Verification
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Neu, T.R. Significance of bacterial surface-active compounds in interaction of bacteria with interfaces. Microbiol. Rev. 1996, 60, 151–166. [Google Scholar] [PubMed]
- Banat, I.M.; Franzetti, A.; Gandolfi, I.; Bestetti, G.; Martinotti, M.G.; Fracchia, L.; Smyth, T.J.; Marchant, R. Microbial biosurfactants production, applications and future potential. Appl. Microbiol. Biotechnol. 2010, 87, 427–444. [Google Scholar] [CrossRef] [PubMed]
- Desai, J.D.; Banat, I.M. Microbial production of surfactants and their commercial potential. Microbiol. Mol. Biol. Rev. 1997, 61, 47–64. [Google Scholar] [PubMed]
- Muller, M.M.; Kugler, J.H.; Henkel, M.; Gerlitzki, M.; Hormann, B.; Pohnlein, M.; Syldatk, C.; Hausmann, R. Rhamnolipids—Next generation surfactants? J. Biotechnol. 2012, 162, 366–380. [Google Scholar] [CrossRef] [PubMed]
- Maier, R.M.; Soberon-Chavez, G. Pseudomonas aeruginosa rhamnolipids: Biosynthesis and potential applications. Appl. Microbiol. Biotechnol. 2000, 54, 625–633. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Mawgoud, A.M.; Lépine, F.; Déziel, E. Rhamnolipids: Diversity of structures, microbial origins and roles. Appl. Microbiol. Biotechnol. 2010, 86, 1323–1336. [Google Scholar] [CrossRef] [PubMed]
- Lyczak, J.B.; Cannon, C.L.; Pier, G.B. Establishment of Pseudomonas aeruginosa infection: Lessons from a versatile opportunist. Microbes Infect. 2000, 2, 1051–1060. [Google Scholar] [CrossRef]
- Gunther, N.W.; Nunez, A.; Fett, W.; Solaiman, D.K.Y. Production of rhamnolipids by Pseudomonas chlororaphis, a nonpathogenic bacterium. Appl. Environ. Microbiol. 2005, 71, 2288–2293. [Google Scholar] [CrossRef] [PubMed]
- Tuleva, B.K.; Ivanov, G.R.; Christova, N.E. Biosurfactant production by a new Pseudomonas putida strain. Zeitschrift für Naturforschung C 2002, 57, 356–360. [Google Scholar] [CrossRef]
- Dubeau, D.; Déziel, E.; Woods, D.E.; Lépine, F. Burkholderia thailandensis harbors two identical rhl gene clusters responsible for the biosynthesis of rhamnolipids. BMC Microbiol. 2009, 9, 263. [Google Scholar] [CrossRef] [PubMed]
- Hörmann, B.; Müller, M.M.; Syldatk, C.; Hausmann, R. Rhamnolipid production by Burkholderia plantarii DSM 9509T. Eur. J. Lipid Sci. Technol. 2010, 112, 674–680. [Google Scholar] [CrossRef]
- Costa, S.G.V.A.O.; Déziel, E.; Lépine, F. Characterization of rhamnolipid production by Burkholderia glumae. Lett. Appl. Microbiol. 2011, 53, 620–627. [Google Scholar] [CrossRef] [PubMed]
- Bahia, F.M.; de Almeida, G.C.; de Andrade, L.P.; Campos, C.G.; Queiroz, L.R.; da Silva, R.L.V.; Abdelnur, P.V.; Correa, J.R.; Bettiga, M.; Parachin, N.S. Rhamnolipids production from sucrose by engineered Saccharomyces cerevisiae. Sci. Rep. 2018, 8, 2905. [Google Scholar] [CrossRef] [PubMed]
- Wittgens, A.; Tiso, T.; Arndt, T.T.; Wenk, P.; Hemmerich, J.; Muller, C.; Wichmann, R.; Kupper, B.; Zwick, M.; Wilhelm, S.; et al. Growth independent rhamnolipid production from glucose using the non-pathogenic Pseudomonas putida KT2440. Microb. Cell Fact. 2011, 10, 80. [Google Scholar] [CrossRef] [PubMed]
- Wittgens, A.; Santiago-Schuebel, B.; Henkel, M.; Tiso, T.; Blank, L.M.; Hausmann, R.; Hofmann, D.; Wilhelm, S.; Jaeger, K.E.; Rosenau, F. Heterologous production of long-chain rhamnolipids from Burkholderia glumae in Pseudomonas putida-a step forward to tailor-made rhamnolipids. Appl. Microbiol. Biotechnol. 2018, 102, 1229–1239. [Google Scholar] [CrossRef] [PubMed]
- Müller, M.M.; Hausmann, R. Regulatory and metabolic network of rhamnolipid biosynthesis: Traditional and advanced engineering towards biotechnological production. Appl. Microbiol. Biotechnol. 2011, 91, 251–264. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.; Lee, T.H.; Nahm, B.H.; Choi, Y.D.; Kim, M.; Hwang, I. Complete genome sequence of Burkholderia glumae BGR1. J. Bacteriol. 2009, 191, 3758–3759. [Google Scholar] [CrossRef] [PubMed]
- Nickzad, A.; Lépine, F.; Déziel, E. Quorum sensing controls swarming motility of Burkholderia glumae through regulation of rhamnolipids. PLoS ONE 2015, 10, e0128509. [Google Scholar] [CrossRef] [PubMed]
- Knapp, A.; Voget, S.; Gao, R.; Zaburannyi, N.; Krysciak, D.; Breuer, M.; Hauer, B.; Streit, W.R.; Muller, R.; Daniel, R.; et al. Mutations improving production and secretion of extracellular lipase by Burkholderia glumae PG1. Appl. Microbiol. Biotechnol. 2016, 100, 1265–1273. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, S.; Das, P.; Sen, R. Towards commercial production of microbial surfactants. Trends Biotechnol. 2006, 24, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Montgomery, D.C. Design and Analysis of Experiments; John Wiley & Sons: Weinheim, Germany, 2008. [Google Scholar]
- Wei, Y.H.; Cheng, C.L.; Chien, C.C.; Wan, H.M. Enhanced di-rhamnolipid production with an indigenous isolate Pseudomonas aeruginosa J16. Process Biochem. 2008, 43, 769–774. [Google Scholar] [CrossRef]
- Deepika, K.V.; Kalam, S.; Sridhar, P.R.; Podile, A.R.; Bramhachari, P.V. Optimization of rhamnolipid biosurfactant production by mangrove sediment bacterium Pseudomonas aeruginosa KVD-HR42 using response surface methodology. Biocatal. Agric. Biotechnol. 2016, 5, 38–47. [Google Scholar] [CrossRef]
- Abalos, A.; Maximo, F.; Manresa, M.A.; Bastida, J. Utilization of response surface methodology to optimize the culture media for the production of rhamnolipids by Pseudomonas aeruginosa AT10. J. Chem. Technol. Biotechnol. 2002, 77, 777–784. [Google Scholar] [CrossRef]
- Zhao, F.; Mandlaa, M.; Hao, J.; Liang, X.; Shi, R.; Han, S.; Zhang, Y. Optimization of culture medium for anaerobic production of rhamnolipid by recombinant Pseudomonas stutzeri Rhl for microbial enhanced oil recovery. Lett. Appl. Microbiol. 2014, 59, 231–237. [Google Scholar] [CrossRef] [PubMed]
- Guerra-Santos, L.; Käppeli, O.; Fiechter, A. Dependence of Pseudomonas aeruginosa continous culture biosurfactant production on nutritional and environmental factors. Appl. Microbiol. Biotechnol. 1986, 24, 443–448. [Google Scholar] [CrossRef]
- Guerra-Santos, L.; Käppeli, O.; Fiechter, A. Pseudomonas aeruginosa biosurfactant production in continuous culture with glucose as carbon source. Appl. Environ. Microbiol. 1984, 48, 301–305. [Google Scholar] [PubMed]
- Soares Dos Santos, A.; Pereira, N., Jr.; Freire, D.M. Strategies for improved rhamnolipid production by Pseudomonas aeruginosa PA1. PeerJ 2016, 4, e2078. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.H.; Chou, C.L.; Chang, J.S. Rhamnolipid production by indigenous Pseudomonas aeruginosa J4 originating from petrochemical wastewater. Biochem. Eng. J. 2005, 27, 146–154. [Google Scholar] [CrossRef]
- Jeong, Y.; Kim, J.; Kim, S.; Kang, Y.; Nagamatsu, T.; Hwang, I. Toxoflavin produced by Burkholderia glumae causing rice grain rot is responsible for inducing bacterial wilt in many field crops. Plant Dis. 2003, 87, 890–895. [Google Scholar] [CrossRef]
- Abdel-Mawgoud, A.M.; Lépine, F.; Déziel, E. Liquid chromatography/mass spectrometry for the identification and quantification of rhamnolipids. Methods Mol. Biol. 2014, 1149, 359–373. [Google Scholar] [PubMed]
- Déziel, E.; Lepine, F.; Milot, S.; Villemur, R. rhlA is required for the production of a novel biosurfactant promoting swarming motility in Pseudomonas aeruginosa: 3-(3-hydroxyalkanoyloxy)alkanoic acids (HAAs), the precursors of rhamnolipids. Microbiology 2003, 149, 2005–2013. [Google Scholar] [CrossRef] [PubMed]
- Schmidberger, A.; Henkel, M.; Hausmann, R.; Schwartz, T. Influence of ferric iron on gene expression and rhamnolipid synthesis during batch cultivation of Pseudomonas aeruginosa PAO1. Appl. Microbiol. Biotechnol. 2014, 98, 6725–6737. [Google Scholar] [CrossRef] [PubMed]
- Ochsner, U.A.; Hembach, T.; Fiechter, A. Production of rhamnolipid biosurfactants. Adv. Biochem. Eng. Biotechnol. 1996, 53, 89–118. [Google Scholar] [PubMed]
- Santos, A.S.; Sampaio, A.P.; Vasquez, G.S.; Santa Anna, L.M.; Pereira, N., Jr.; Freire, D.M. Evaluation of different carbon and nitrogen sources in production of rhamnolipids by a strain of Pseudomonas aeruginosa. Appl. Biochem. Biotechnol. 2002, 98–100, 1025–1035. [Google Scholar] [CrossRef]
- Nickzad, A.; Deziel, E. Adaptive Significance of Quorum Sensing-Dependent Regulation of Rhamnolipids by Integration of Growth Rate in Burkholderia glumae: A Trade-Off between Survival and Efficiency. Front. Microbiol. 2016, 7, 1215. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experimental Values | |||
|---|---|---|---|
| Variables | Units | Lower | Higher |
| Mannitol | % w/v | 0.5 | 2 |
| Urea | M | 0.02 | 0.2 |
| Potassium phosphate buffer, pH 7 | mM | 10 | 40 |
| CaCl2·2H2O | g/L | 0.01 | 0.1 |
| MgSO4·7H2O | g/L | 0.1 | 0.4 |
| Trace elements solution | mL/L | 0 | 2 |
| Temperature | °C | 30 | 37 |
| Agitation | rpm | 100 | 250 |
| Range and Levels | ||||||
|---|---|---|---|---|---|---|
| Variables | Symbol Coded | −2 | −1 | 0 | +1 | +2 |
| Mannitol (% w/v) | X1 | 0.5 | 1.375 | 2.25 | 3.125 | 4 |
| Urea (M) | X2 | 0.01 | 0.0575 | 0.105 | 0.1525 | 0.2 |
| Potassium phosphate buffer, pH 7 (mM) | X3 | 10 | 32.5 | 55 | 77.5 | 100 |
| CaCl2·2H2O (g/L) | X4 | 0.01 | 0.1325 | 0.255 | 0.3775 | 0.5 |
| Run | Mannitol (%) | Urea (M) | Potassium Phosphate Buffer (mM) | CaCl2·2H2O (g/L) | MgSO4·7H2O (g/L) | Trace Elements | Temperature (°C) | Agitation (rpm) | RLs Production (mg/L) (Mean ± SD) | Biomass (g/L) (Mean ± SD) |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 0.5 | 0.02 | 10 | 0.01 | 0.1 | − | 30 | 100 | 415.9 ± 77.0 | 0.25 ± 0.01 |
| 2 | 2 | 0.02 | 10 | 0.01 | 0.1 | + | 37 | 250 | ND 1 | 0.24 ± 0.11 |
| 3 | 0.5 | 0.2 | 10 | 0.01 | 0.4 | − | 37 | 250 | 201.1 ± 79.4 | 0.54 ± 0.03 |
| 4 | 2 | 0.2 | 10 | 0.01 | 0.4 | + | 30 | 100 | ND | 0.27 ± 0.03 |
| 5 | 0.5 | 0.02 | 40 | 0.01 | 0.4 | + | 37 | 100 | ND | 0.19 ± 0.02 |
| 6 | 2 | 0.02 | 40 | 0.01 | 0.4 | − | 30 | 250 | 736.1 ± 158.4 | 1.96 ± 0.12 |
| 7 | 0.5 | 0.2 | 40 | 0.01 | 0.1 | + | 30 | 250 | ND | 0.26 ± 0.04 |
| 8 | 2 | 0.2 | 40 | 0.01 | 0.1 | − | 37 | 100 | 215.4 ± 12.8 | 0.93 ± 0.15 |
| 9 | 0.5 | 0.02 | 10 | 0.1 | 0.4 | + | 30 | 250 | ND | 0.25 ± 0.02 |
| 10 | 2 | 0.02 | 10 | 0.1 | 0.4 | − | 37 | 100 | 359.6 ± 56.6 | 0.95 ± 0.13 |
| 11 | 0.5 | 0.2 | 10 | 0.1 | 0.1 | + | 37 | 100 | ND | 0.26 ± 0.04 |
| 12 | 2 | 0.2 | 10 | 0.1 | 0.1 | − | 30 | 250 | 482.27 ± 93.7 | 1.23 ± 0.17 |
| 13 | 0.5 | 0.02 | 40 | 0.1 | 0.1 | − | 37 | 250 | 268.53 ± 75.4 | 0.82 ± 0.10 |
| 14 | 2 | 0.02 | 40 | 0.1 | 0.1 | + | 30 | 100 | 972.72 ± 271.3 | 4.09 ± 0.12 |
| 15 | 0.5 | 0.2 | 40 | 0.1 | 0.4 | − | 30 | 100 | 444.08 ± 76.8 | 0.82 ± 0.06 |
| 16 | 2 | 0.2 | 40 | 0.1 | 0.4 | + | 37 | 250 | 404.32 ± 212.6 | 2.49 ± 1.11 |
| Source | Sum of Squares | Degrees of Freedom | Mean Square | F Value | p-Value (Prob > F) |
|---|---|---|---|---|---|
| Model | 3.76 | 11 | 0.34 | 20.16 | <0.0001 |
| A-Mannitol | 0.64 | 1 | 0.64 | 37.48 | <0.0001 |
| B-Urea | 0.19 | 1 | 0.19 | 11.18 | 0.0019 |
| C-Potassium phosphate buffer | 0.47 | 1 | 0.47 | 27.69 | <0.0001 |
| D-CaCl2·2H2O | 0.35 | 1 | 0.35 | 20.55 | <0.0001 |
| F-Trace elements | 0.57 | 1 | 0.57 | 33.71 | <0.0001 |
| G-Temperature | 0.48 | 1 | 0.48 | 28.39 | <0.0001 |
| AB | 0.16 | 1 | 0.16 | 9.51 | 0.0039 |
| AC | 0.36 | 1 | 0.36 | 21.40 | <0.0001 |
| AD | 0.26 | 1 | 0.26 | 15.18 | 0.0004 |
| AF | 0.16 | 1 | 0.16 | 9.22 | 0.0044 |
| AG | 0.13 | 1 | 0.13 | 7.46 | 0.0097 |
| Residual | 0.61 | 36 | 0.017 | ||
| Lack of Fit | 0.071 | 4 | 0.018 | 1.05 | 0.3980 |
| Pure Error | 0.54 | 32 | 0.017 | ||
| Cor Total | 4.37 | 47 |
| Source | Sum of Squares | Degrees of Freedom | Mean Square | F Value | p-Value (Prob > F) |
|---|---|---|---|---|---|
| Model | 50.65 | 10 | 5.07 | 41.78 | <0.0001 |
| A-Mannitol | 14.41 | 1 | 14.41 | 118.82 | <0.0001 |
| B-Urea | 0.72 | 1 | 0.72 | 5.94 | 0.0197 |
| C-Potassium phosphate buffer | 10.77 | 1 | 10.77 | 88.82 | <0.0001 |
| D-CaCl2·2H2O | 7.37 | 1 | 7.37 | 60.78 | <0.0001 |
| G-Temperature | 1.38 | 1 | 1.38 | 11.35 | 0.0018 |
| AB | 1.37 | 1 | 1.37 | 11.29 | 0.0018 |
| AC | 6.74 | 1 | 6.74 | 55.57 | <0.0001 |
| AD | 3.73 | 1 | 3.73 | 30.78 | <0.0001 |
| AF | 2.30 | 1 | 2.30 | 18.93 | 0.0001 |
| AG | 1.88 | 1 | 1.88 | 15.47 | 0.0004 |
| Residual | 4.49 | 37 | 0.12 | ||
| Lack of Fit | 0.40 | 5 | 0.081 | 0.63 | 0.6763 |
| Pure Error | 4.08 | 32 | 0.13 | ||
| Cor Total | 55.14 | 47 |
| Source | Sum of Squares | Degrees of Freedom | Mean Square | F Value | p-Value (Prob > F) |
|---|---|---|---|---|---|
| Block | 0.12 | 1 | 0.12 | ||
| Model | 4.12 | 9 | 0.46 | 35.76 | <0.0001 |
| A-Mannitol | 0.24 | 1 | 0.24 | 18.56 | <0.0001 |
| B-Urea | 0.11 | 1 | 0.11 | 8.42 | 0.0052 |
| C-Potassium phosphate buffer | 3.08 | 1 | 3.08 | 240.57 | <0.0001 |
| D-CaCl2·2H2O | 0.34 | 1 | 0.34 | 26.47 | <0.0001 |
| AB | 0.14 | 1 | 0.14 | 10.62 | 0.0019 |
| BC | 0.098 | 1 | 0.098 | 7.65 | 0.0076 |
| CD | 0.14 | 1 | 0.14 | 10.67 | 0.0018 |
| C2 | 0.081 | 1 | 0.081 | 6.33 | 0.0147 |
| D2 | 0.19 | 1 | 0.19 | 15.01 | 0.0003 |
| Residual | 0.74 | 58 | 0.013 | ||
| Lack of Fit | 0.22 | 14 | 0.016 | 1.32 | 0.2320 |
| Pure Error | 0.52 | 44 | 0.012 | ||
| Cor Total | 4.98 | 68 |
| Source | Sum of Squares | Degrees of Freedom | Mean Square | F Value | p-Value (Prob > F) |
|---|---|---|---|---|---|
| Block | 0.010 | 1 | 0.010 | ||
| Model | 79.62 | 5 | 15.92 | 130.96 | <0.0001 |
| A-Mannitol | 14.23 | 1 | 14.23 | 117.02 | <0.0001 |
| C-Potassium phosphate buffer | 61.92 | 1 | 61.92 | 509.29 | <0.0001 |
| D-CaCl2·2H2O | 3.39 | 1 | 3.39 | 27.92 | <0.0001 |
| AC | 1.07 | 1 | 1.07 | 8.81 | 0.0048 |
| A2 | 0.79 | 1 | 0.79 | 6.53 | 0.0141 |
| Residual | 5.47 | 45 | 0.12 | ||
| Lack of Fit | 2.57 | 17 | 0.15 | 1.46 | 0.1818 |
| Pure Error | 2.90 | 28 | 0.10 | ||
| Cor Total | 85.10 | 51 |
| Source | Sum of Squares | Degrees of Freedom | Mean Square | F Value | p-Value (Prob > F) |
|---|---|---|---|---|---|
| Block | 7.163 × 10−5 | 1 | 7.163 × 10−5 | ||
| Model | 0.013 | 10 | 1.316 × 10−3 | 27.01 | <0.0001 |
| A-Mannitol | 5.156 × 10−3 | 1 | 5.156 × 10−3 | 105.78 | <0.0001 |
| B-Urea | 8.171 × 10−4 | 1 | 8.171 × 10−4 | 16.76 | 0.0002 |
| C-Potassium phosphate buffer | 1.737 × 10−3 | 1 | 1.737 × 10−3 | 35.64 | <0.0001 |
| D-CaCl2·2H2O | 2.535 × 10−3 | 1 | 2.535 × 10−3 | 52.00 | <0.0001 |
| AC | 2.523 × 10−4 | 1 | 2.523 × 10−4 | 5.18 | 0.0280 |
| AD | 4.779 × 10−4 | 1 | 4.779 × 10−4 | 9.80 | 0.0031 |
| BC | 3.271 × 10−4 | 1 | 3.271 × 10−4 | 6.71 | 0.0130 |
| A2 | 7.420 × 10−4 | 1 | 7.420 × 10−4 | 15.22 | 0.0003 |
| B2 | 2.165 × 10−4 | 1 | 2.165 × 10−4 | 4.44 | 0.0409 |
| C2 | 3.383 × 10−4 | 1 | 3.383 × 10−4 | 6.94 | 0.0117 |
| Residual | 2.096 × 10−3 | 43 | 4.875 × 10−5 | ||
| Lack of Fit | 7.169 × 10−4 | 13 | 5.515 × 10−5 | 1.20 | 0.3266 |
| Pure Error | 1.379 × 10−3 | 30 | 4.597 × 10−5 | ||
| Cor Total | 0.015 | 54 |
| Mannitol (% w/v) | Urea (M) | Potassium Phosphate Buffer (mM) | CaCl2·2H2O (g/L) | Desirability |
|---|---|---|---|---|
| 0.5 | 0.01 | 100 | 0.01 | 0.107 |
| Response | Predicted Value | 95% PI Low | Mean Experimental Value | 95% PI High |
|---|---|---|---|---|
| Rhamnolipid (g/L) | 2.02 | 1.67 | 1.66 | 2.38 |
| Biomass (g/L) | 2.78 | 1.96 | 0.85 | 3.61 |
| YP/S (g/g) | 0.12 | 0.09 | 0.34 | 0.15 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nickzad, A.; Guertin, C.; Déziel, E. Culture Medium Optimization for Production of Rhamnolipids by Burkholderia glumae. Colloids Interfaces 2018, 2, 49. https://doi.org/10.3390/colloids2040049
Nickzad A, Guertin C, Déziel E. Culture Medium Optimization for Production of Rhamnolipids by Burkholderia glumae. Colloids and Interfaces. 2018; 2(4):49. https://doi.org/10.3390/colloids2040049
Chicago/Turabian StyleNickzad, Arvin, Claude Guertin, and Eric Déziel. 2018. "Culture Medium Optimization for Production of Rhamnolipids by Burkholderia glumae" Colloids and Interfaces 2, no. 4: 49. https://doi.org/10.3390/colloids2040049
APA StyleNickzad, A., Guertin, C., & Déziel, E. (2018). Culture Medium Optimization for Production of Rhamnolipids by Burkholderia glumae. Colloids and Interfaces, 2(4), 49. https://doi.org/10.3390/colloids2040049