Abstract
Structural features of the overstorey in managed and unmanaged forests can significantly influence plant community composition. Native Acacia species are common in temperate eucalypt forests in southeastern Australia. In these forests, intense disturbances, such as logging and wildfire, lead to high densities of regenerating trees, shrubs, and herbs. The tree layer is dominated by Acacia and Eucalyptus, that compete intensely for resources in the first decades after stand establishment. The relative abundance and size of Acacia and Eucalyptus varies widely due to stochastic factors such as dispersal, microsite variability, and weather and climatic conditions. This variability may influence the structure and composition of the herbaceous and shrub species. In the temperate forests of southeastern Australia, understorey plant diversity is assumed to be influenced by Acacia species density, rather than Eucalyptus density. To quantify the influence of Acacia and Eucalyptus density on plant community composition, we used remote sensing and machine learning methods to map canopy composition and then compare it to understorey composition. We combined unoccupied aerial vehicle (UAV or drone) imagery, supervised image classifications, and ground survey data of plant composition from post-logging regrowth forests in the Central Highlands of southeastern Australia. We found that aggregation and patch metrics of Eucalyptus and Acacia were strongly associated with understorey plant beta diversity. Increasing aggregation of Acacia and the number of Acacia patches had a significant negative effect on plant beta diversity, while the number of Eucalyptus patches had a positive influence. Our research demonstrates how accessible UAV remote sensing can be used to quantify variability in plant biodiversity in regrowth forests. This can help forest managers map patterns of plant diversity at the stand-scale and beyond to guide management activities across forested landscapes.
1. Introduction
Overstorey composition and structure are not static in time or space and are influenced by plant successional response to disturbance [1,2]. Understanding species turnover and changes in species composition in response to these stand dynamics enhances our capability to manage and conserve forest ecosystems, which is vital for achieving sustainable forest management. In many forested ecosystems, the diversity of understorey vegetation is driven by the structure and diversity of canopy trees [3,4] via effects on resource availability [5,6,7]. In particular, the spatial configuration of the overstorey can influence light transmission, microclimatic conditions, and soil nutrient availability [3,4,8,9,10,11].
Plant biodiversity is strongly linked to the spatial configuration and aggregation of tree canopies [12]. Post-disturbance legacies can significantly influence plant community composition [13]. In temperate Eucalyptus forests, native Acacia species are fast growing mid-storey species that often regenerate at high densities after stand-replacing disturbances [14,15,16]. High densities of Acacia species can typically be found in these forests after timber harvesting [17,18]. Acacia and Eucalyptus trees compete for the first decade after stand establishment, after which Eucalyptus typically outcompetes Acacia [14,19] and forms the overstorey, while Acacia remains in the midstorey at lower densities. In the temperate forests of southeastern Australia, understorey plant diversity is influenced by Acacia species density, rather than Eucalyptus density [15,16,20]. Given the importance of Acacia density in stand development and its influence on plant species composition, quantifying the effect of Acacia and Eucalyptus on understorey diversity is important for understanding how spatial heterogeneity in canopy composition might influence species turnover at the stand scale.
Remotely sensed data are increasingly being used to investigate stand structure and composition in forests over large areas as a practical and cost-effective alternative or complement to ground surveys [21,22,23,24,25,26]. Remote-sensing technologies, however, operate across a range of spatial resolutions and extents. For example, widely used satellite imagery usually covers broad extents and has a range of resolutions from coarse-grain (e.g., 30 m ground sampling distance in Landsat satellites) to fine-grain (5 m or less in, e.g., Hyperion and IKONOS sensors [27]). These data products may be either too coarse to study forest structure or too expensive to study forests at relevant scales. There are different sensors with different capabilities of spatial, temporal, spectral, and radiometric characteristics and the selection of appropriate sensors depends on the mapping objective. Unoccupied Aerial Vehicles (UAVs or drones) are a low-cost alternative to high-resolution, fee-for-service imagery or labor-intensive, ground-based inventories. UAVs have been used for biodiversity assessments and measurements of forest structural features such as tree height, volume, and density [24,28,29]. Today, UAV technology is inexpensive, efficient, and flexible. UAVs can be equipped with a range of sensors, increasing their utility and suitability for ecological research [22]. UAVs can also be used to map vegetation cover using supervised image classification [30], which allows remote assessments of the spatial configuration of forests [31,32]. As such, machine learning algorithms such as Random Forests [33] have been widely used to classify land cover classes and from that, carry out analyses of landscape metrics to quantify spatial aggregation and fragmentation [34]. Such machine learning approaches have, for example, been used to determine land use and delineate trees in forested [29,35] and urban environments [36,37], map canopy chemistry to determine foraging quality for arboreal folivores [38] or ground dwelling herbivores [39], and map canopy species richness or trait diversity with hyperspectral imagery [40,41].
The objective of this research was to use methods of supervised classification of UAV forest stand imagery to study how variability in Acacia and Eucalyptus canopy composition and density can influence understorey plant species diversity at the stand scale in temperate Eucalyptus forests. Quantifying the influence of Acacia and Eucalyptus composition and configuration on plant turnover using remote sensing will provide an important tool for mapping patterns of plant diversity in post-disturbance forests. To achieve this, we combined UAV imagery with ground survey data of plant composition from young post-logging regrowth forests in the Central Highlands of southeastern Victoria, Australia. We focused on young regrowth stands in which most species are present soon after disturbance with tree species dominating the canopy over time [20,42,43,44].
2. Materials and Methods
2.1. Study Area
Our study was conducted in mountain ash (Eucalyptus regnans) regrowth stands of the Central Highlands of Victoria, southeastern Australia (Figure 1). The region ranges in elevation between 75 and ~1500 m above sea level. Mean annual temperatures range between 5.8–15.2 °C, with precipitation between 564–2089 mm [45,46]. We selected stands that had been harvested using the clearfell, burn, and sow (CBS) silvicultural system that has been standard practice in these forests until recently [47]. We studied a total of 12 harvested and regenerated coupes, with stand ages between 10- and 16-years post timber harvesting (Figure 1).
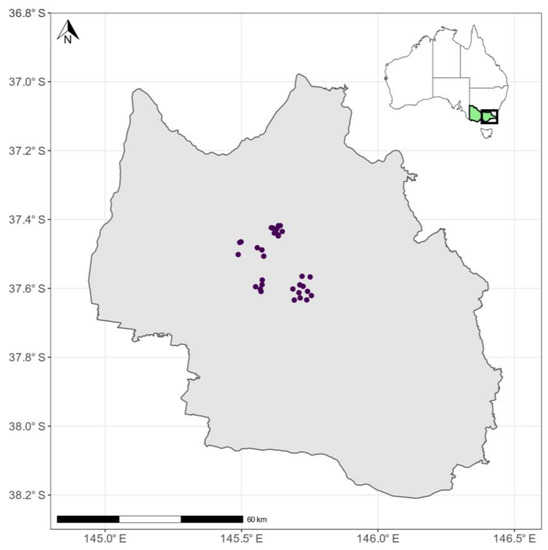
Figure 1.
The Central Highlands (grey) of Victoria (green in inset map) in southeastern Australia. Points indicate the location of survey plots (three per coupe, n = 36, note, due to the scale, some points are overlapping) within the studied harvesting coupes (N = 12). Geographic coordinates for all sites and plots can be found as a spatial file in Supplementary Materials S2.
2.2. Field Observation Data
At each site we assessed plant community composition in a set of circular plots (40 m2 area). Three plots were established within each coupe (number of plots = 36), using a stratified random sampling design to record overstorey stem densities and number of plant species per plot. Plots were located at least 50 m from the forest edge and 100 m from each other. The post-harvest overstorey regeneration in these forests is dominated by E. regnans and/or Acacia dealbata, A. frigescens, and A. obliquinervia. Common understorey species included: Cassinia aculeata, Tetrarrhena juncea, and Pteridium esculentum. Plant community composition was assessed using the presence and absence of vascular plant species. We calculated beta diversity using the Jaccard Index, as it provides a robust measure of diversity from presence and absence data. Beta diversity is a measure of change in diversity between sites [48]. It is used to quantify the spatial turnover of species and is central to understanding processes resulting in species’ loss or gains, particularly in the context of conservation planning at landscape scales [49]. Beta diversity is expressed as the number of unique species (species only present in one of the sites) between sites [48]. To calculate beta diversity, we combined the number of species per site (sum of plant species from three plots per site). We used the package vegan [50] in R to calculate beta diversity.
2.3. UAV Imagery
Aerial imagery over each coupe was collected using an autopiloted Phantom 4 Pro V2 (DJI, Shenzhen, China) multirotor unoccupied aerial vehicle (UAV) in October and November 2019. This UAV captures visible light reflectance (red, green, and blue band, RGB) on a gimbal stabilized 1-inch 20 MP CMOS sensor. The sensor has a field of view of 84° with a 35 mm format equivalent lens allowing apertures between f/2.8–f/11. Using an electronic shutter, the sensor can reach shutter speeds of 8-1/8000 s. UAV flights were carried out over the course of three days in November 2020. We flew during overcast weather conditions for consistent ambient light in imagery from all sites. Flights were carried out at a maximum of two hours before or after noon (12 pm). For our imagery, we used an aperture of f/4.5, exposure of 1/200 s at ISO 100–500 depending on the light conditions on the day of imagery collection. Lens focal length was set to 9 mm for all flights. The flight paths for each coupe were planned and completed using the Ground Station Pro flight control software (DJI, Shenzhen, China). The flight altitude was at 100–120 m above the ground and within visible line of sight to the UAV operator, in accordance with the Australian Civil Aviation Safety Authority [51]. This flight altitude resulted in an average ground resolution of ~2.5 cm per pixel across all images captured. The UAV was launched and recovered from forest roads near the plot centers, which also ensured to capture sufficient ground imagery and accurately map ground elevation when using photogrammetry [52].
2.4. Imagery Processing
Aerial imagery was processed using Metashape 2.0.0 (Agisoft, St. Petersburg, Russia), which uses Structure from Motion (SfM) to derive three-dimensional (3D) structure from two-dimensional (2D) overlapping image sequences through photogrammetry processing (Ullman 1979). Processing resulted in 3D point clouds of each coupe, which were used to calculate orthomosaics of the three available color bands (red, green, and blue reflectance, Figure 2A). The resulting processed UAV imagery was analyzed using the raster, sf, and lidR packages [53,54,55] in R version 4.0.1 [56].

Figure 2.
Data products derived from UAV imagery used for canopy classification. (A) True-colour orthomosaic compiled from red, green, and blue reflectance bands, and (B) canopy height model derived from UAV point clouds, expressing tree height in meters above the ground.
For an additional independent variable for image classification, we generated a canopy height model (CHM) from point clouds for each coupe. Point clouds were normalized to a 0 m ground elevation. We then calculated digital terrain and digital surface models (DTM + DSM), expressing the ground elevation and point elevation above ground, respectively, using lidR. The CHM is the difference between DSM and DTM (CHM = DSM − DTM). It is expressed as a 2D raster in which each cell represents the respective canopy height above ground in meters. Resulting CHMs (Figure 2B) were smoothed using a sub-circle algorithm that added a 0.5 m disk around each cell to close empty pixels that would otherwise be a product of the point cloud density [57]. The final CHMs had a resolution of 0.5 m.
The three visible spectral bands (red, green, and blue light reflectance) from the UAV sensor, along with canopy height, were compiled as multi-layer raster stacks for each coupe and used as covariates to classify the imagery into areas of Acacia and Eucalyptus cover (see next section for details). These metrics were subsequently used in a model of plant beta diversity. To reduce processing times, we cropped each coupe’s raster stack to 2-hectare square areas (~141.5 × 141.5 m) around the ground survey plot center. Two plots had to be removed because the imagery only partially covered the 2-hectare square area. A total of 34 plots were included in the final classification analysis. To test the resulting plant diversity model on a range of potential stand configurations, we further extracted an additional 415 1-hectare blocks (100 × 100 m) from the coupe raster stacks that did not intersect with our plots or the image boundary.
2.5. Forest Cover Classification and Spatial Metrics
To predict the configuration of Acacia and Eucalyptus cover in each plot we used a supervised classification approach. We created training polygons to extract RGB reflectance and canopy height from the CHM to classify the plot imagery into three classes: (1) Eucalyptus cover, (2) Acacia cover, and (3) other features such as canopy gaps. The polygons were assigned to these cover classes using image interpretation. Up to 30 polygons for each cover class were placed on each 2-hectare square area (Figure 3A). The polygons were used as extraction masks on each plot’s raster stack to collate variable values describing the respective class. The extracted cell values for RGB reflectance and canopy height from the CHM were used as covariates to train a random forests machine learning model [33]. Analyses were carried out in R, using the packages randomForest and caret [58,59]. We extracted 1500 cell values per class (n = 4500 cells per plot, N = 153,000 cells), which were paired with their manually identified and assigned cover classes. To develop a universal model of cover classes across coupe and stand configurations, we combined extracted pixels from all plots. To build the model, extracted data was split into training (80%) and testing (20%) datasets. We trained the random forest model using the training dataset of the four UAV-derived covariates as predictors and the cover class as response variable [60,61]. Model performance was evaluated by predicting cover classes to the testing dataset (independent validation). We used model accuracy, based on average absolute model error and Cohen’s Kappa, a measure of model performance based on confusion matrices. To further quantify the accuracy of model performance, we used sensitivity (true positive rate—TPR) and specificity (true negative rate—TNR) and calculated the true skill statistic (). TSS values range from 0 to 1 and provide inference similar to the Kappa statistic, but without being dependent on prevalence [62,63]. We then used the model to predict to each plot’s 2-hectare raster stack, as well as the 415, 1-hectare stacks, creating classified maps of forest structure (Figure 3B) for further analysis of spatial configuration of Eucalyptus and Acacia cover and their influence on species beta diversity.
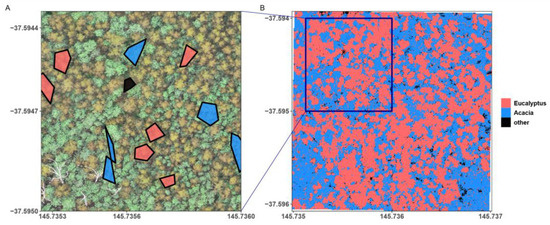
Figure 3.
(A) Example of training polygons placed using image interpretation on the RGB orthomosaic for the three target classes, and (B) spatial map of the forest canopy showing the configuration of Eucalyptus, Acacia, and other (i.e., gaps) cover classes as predicted by the final supervised classification model.
2.6. Relationship between Field and Imagery Data
To determine the relationship between beta diversity and forest structure derived from UAV imagery, we calculated the percentage cover for each class per plot and derived the number of Eucalyptus and Acacia patches, as well as the class spatial aggregation using the R package landscapemetrics [34]. In earlier research, we found that climate was a particularly strong predictor for plant species diversity in these forests [16]. Therefore, we added the average annual heat moisture index (AHMI) for each plot and additional 1-hectare testing areas from mean annual temperature (MAT) and annual precipitation (MAP, ), see Wang et al. [64], Paudel et al. [65]). AHMI was calculated from ~250 m resolution daily climate data extracted from Stewart et al. [45], Stewart and Nitschke [66,67], Fedrigo et al. [68]. We used generalized linear models (GLMs) to test the relationships between the derived metrics and beta diversity as expressed by the Jaccard index. All predictor variables were tested for intercorrelation before being used in the model and then log-transformed. We used a dredge function from the package MuMIn [69] to find the best performing model based on Akaike information criterion (AIC) and delta value.
3. Results
3.1. Observed Species Diversity
We recorded a total of 74 above-ground plant species with an average of 12.1 ± 3.4 and a range of 5–21 species per surveyed plot (Supplementary Materials S1: Table S1). The most frequent understorey species were Tetrarrhena juncea, Pteridium esculentum, and Pomaderris aspera.
3.2. Supervised Classification: Variable Importance and Model Performance
All four remotely sensed variables improved supervised classification performance. Canopy height derived from the unoccupied aerial vehicle (UAV) canopy height models (CHM) was the most important predictor variable based on both mean decrease accuracy and mean decrease Gini (Figure 4). Mean decrease accuracy refers to the loss in model accuracy if a variable was removed from the model. A Gini value of 0 refers to pure classification and the maximum value refers to random distribution across all predicted classes. Visible blue light reflectance and CHM were equally important in predicting forest cover classes while red light reflectance was the second most important predictor based on Gini.
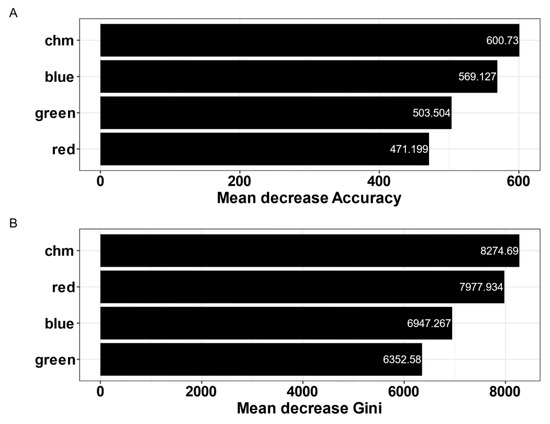
Figure 4.
Variable importance from the final random forests model of cover classification, according to mean decrease in model accuracy (A) and mean decrease Gini (B). chm = Tree height from the canopy height models; red, green, and blue refer to the visible reflectance bands collected from the UAV sensor.
Independent model validation suggested good classification performance. Overall model accuracy was 77% (p < 0.001), with a model Kappa of 0.65, confirming a substantial agreement between predicted and observed cover classes. Sensitivity and specificity for all three classes were high, leading to good true skill statistic values across classes (Supplementary Materials S1: Table S2). The best performing class was Acacia cover with a TSS of 0.71, followed by Eucalyptus cover with a TSS of 0.67, which also indicates substantial agreement.
3.3. Spatial Configuration and Arrangement
Across the 34 2-hectare plots for which we predicted forest cover, we observed a variety of forest overstorey configurations (Supplementary Materials S1: Figure S1). Eucalyptus cover ranged from 24.6–88.1% and Acacia cover from 6.3–61%. Other features (e.g., forest tracks or canopy gaps, class ‘other’) classified as ‘unknown’ covered 2.7–22.7%. Spatial aggregation was on average similar for Eucalyptus (0.77 ± 0.06; range: 0.59–0.87) and Acacia (0.76 ± 0.06; range: 0.54–0.85). The same patterns were observed for the number of patches of Eucalyptus and Acacia (Supplementary Materials S1: Figure S2). For the 415 1-hectare areas outside the circular plots, we observed a wider range for canopy cover, aggregation, and number of patches. Eucalyptus cover ranged from 11.0–96.2% and Acacia cover from 1.0–75.0%, while aggregation ranged from 0.45–0.87 and 0.46–0.85 for Eucalyptus and Acacia, respectively. Log-transformed mean number of patches was slightly higher for Acacia (4.41 ± 3.83) than Eucalyptus (4.02 ± 3.79).
3.4. Influence of Remotely Sensed Forest Structure on Beta Diversity
Using generalized linear models (GLMs), we found that derived forest configurations from the spatial classifications of Eucalyptus and Acacia cover metrics were useful in estimating understorey plant beta diversity of the forest. We found significant relationships between the aggregation metrics derived from UAV imagery, as well as site annual heat moisture index (AHMI), and beta diversity. The best model (R2 = 0.57) contained the log-transformed aggregation of Acacia, number of patches of both Eucalyptus and Acacia, and AHMI. Increasing AHMI, aggregation of Acacia, and number of Acacia patches had significant negative effects on beta diversity. In contrast, the number of patches of Eucalyptus had a positive influence on beta diversity (Table 1, Figure 5, Supplementary Materials S1: Table S3). Aggregation of Acacia was the most influential variable based on estimate comparison (Table 1, Supplementary Materials S1: Table S3).

Table 1.
Estimate, standard error, and significance level of predicted beta diversity from the best GLM model (based on AIC: −25.71) using aggregation of Acacia, number of Acacia and Eucalyptus from spatial predictions, and annual heat moisture index (AHMI) as predictor variables. All the variables were log-transformed.
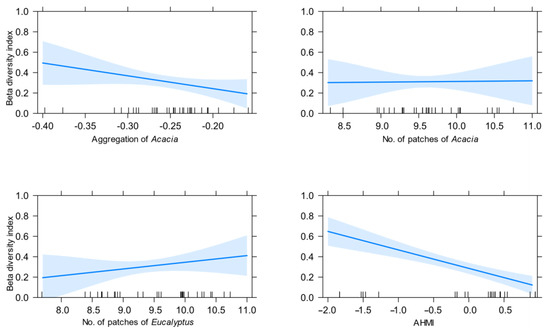
Figure 5.
Partial dependence plots for the best performing generalized linear model (GLM) of beta diversity using aggregation of Acacia, number of patches of Acacia and Eucalyptus from spatial predictions, and the annual heat moisture index (AHMI). All variables were log-transformed prior to modeling.
We used the spatial configuration of the additional 415 1-hectare areas to illustrate the relationships between the remotely sensed variables identified and plant beta diversity. Areas with large aggregations of Acacia and few patches of Eucalyptus had the lowest beta diversity, whereas a patchy distribution of Eucalyptus with lower or dispersed Acacia cover was associated with the highest beta diversity (Figure 6, Supplementary Materials S1: Figure S3). AHMI had a negative influence on beta diversity (Table 1; Figure 5, Supplementary Materials S1: Figure S3).
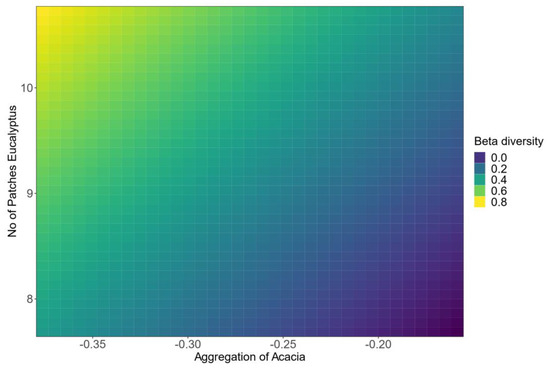
Figure 6.
Prediction matrix from model testing on 415 1-hectare forest areas, illustrating the influence of (log-transformed) spatial aggregation of Acacia and number of patches of Eucalyptus on understorey beta diversity. With increasing aggregation of Acacia, beta diversity decreases, while the reverse holds true for the number of patches of Eucalyptus. The more patches of Eucalyptus present, the greater beta diversity of understorey plant species.
4. Discussion
Spatial variability in canopy structure and composition should influence patterns of plant diversity in the understorey by altering resource availability. However, linking the canopy structure to understorey composition presents a number of logistical challenges, particularly over broader scales. Here we demonstrate that the relative abundance and spatial aggregation of a canopy tree species in the stand reduces beta diversity in the understorey of young forests. We also show that commercial unoccupied aerial vehicles (UAVs) provide an efficient and cost-effective tool for exploring these relationships over large areas. We found that the spatial patterns of Acacia and Eucalyptus have opposite effects on plant beta diversity. Increased aggregation of Acacia in regrowth forests leads to lower species beta diversity, suggesting Acacia dominance has a homogenizing influence on plant diversity. In contrast, Eucalyptus dominance promoted beta diversity. While aridity was found to be an important predictor of beta diversity at the landscape level [70], spatial patterns in forest structure affect stand scale diversity, which has important implications for understanding how different stand development pathways can shape plant diversity in the temperate Eucalyptus forests of southeastern Australia [16,43,71]. We show that using UAVs to map plant diversity at the stand-scale can be a cost-effective alternate and readily applicable approach for assessments of plant species diversity in regrowth forests.
4.1. Spatial Aggregation of Acacia
The importance of arrangement and aggregation of Acacia highlights the need for a deeper understanding of the patterns and processes that shape plant diversity in forests affected by stand-replacing disturbances [16,70,71,72,73]. The main objective of the clearfell, burn, and sow (CBS) silvicultural system in the study area is to promote a dense, homogeneous cohort of Eucalyptus recruits [74]. In some of the studied coupes this did not occur due to failures in Eucalyptus regeneration; this resulted in Acacia dominating the stands [70,71]. Our earlier work demonstrated that seasonal climate impacts Eucalyptus regeneration, leading to lower occupancy during climatically unfavourable years [70,71]. This can favor the recruitment of Acacia in areas where eucalypts have failed to recruit, as Acacia species form persistent soil seedbanks that promote rapid regeneration in response to disturbance [17,18,75,76]. Our results suggest that under these conditions, areas with highly aggregated Acacia develop, which leads to lower plant beta diversity. Bauhus et al. [77] found that Acacia growing in mixtures with Eucalyptus did not form viable crowns, whereas when grown in pure stands, they were able to form full crowns that drastically reduced light transmission to the understorey. Acacia are likely shaping plant diversity through light limitation in areas where crown aggregation occurs in the absence of competition with Eucalyptus (Figure 7, Supplementary Materials S1: Figure S1). Trouvé et al. [71] found that the CBS system typically leads to homogeneous regeneration (in both size and density) for eucalypts. However, in our study, we found that there was great variability in stand structures in young post-logging regrowth (Figure 7). Variability in regeneration patterns created heterogeneity in stand density and overstorey composition within these stands. These heterogenous patterns promote increased alpha diversity within stands but also increase turnover (i.e., beta diversity) as species occurrence is shaped at fine scales by changes in overstorey composition. These findings highlight how spatial patterns in canopy composition, which originate at the time of regeneration, can have long-term impacts on plant community composition. In our study sites, eucalypt dominance facilitated turnover while Acacia dominance reduced it.
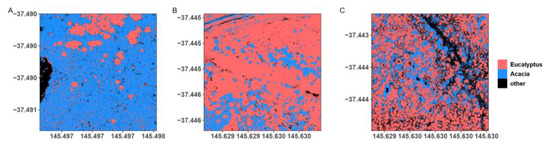
Figure 7.
Different configurations of the Acacia, Eucalyptus, and ‘other’ classification, predicted to 1-hectare squares within the studied coupe areas. The three configurations have distinct effects on plant beta diversity: A forest dominated by dense Acacia cover (A) results in a low understorey diversity, while a stand with low Acacia cover, but high Eucalyptus aggregation (B) results in a medium understorey. Low Acacia cover and a patchy Eucalyptus overstorey (C) results in the highest plant beta diversity, according to the final model.
4.2. Implications for Forest Management and Future Directions
Trouvé et al. [71] found that patterns of eucalypt regeneration after catastrophic fire disturbances and the timber harvesting differed. They identified that different patterns in regeneration post-fire or post-harvesting could impact future stand dynamics and structure. Forests affected by wildfire were more likely to have high-density clumps of Eucalyptus regeneration. These clumps were separated by areas with little or no Eucalyptus regeneration and often abundant regeneration of other species (including Acacia) [71]. This initial pattern of recruitment appears to generate spatially variable patterns in plant species composition and turnover within and between sites. Higher densities of Acacia therefore have implications for plant diversity in young stands that establish after severe disturbances. Differences in the relative abundance and spatial patterning of Eucalyptus and Acacia regrowth after disturbances may put stands on different development trajectories during the early stages of stand development. We previously found that density and occurrence of Eucalyptus regeneration was influenced by climate at a landscape scale, with drought conditions leading to lower levels of regeneration [70]. Our findings suggest that changes in Eucalyptus regeneration could also lead to changes in plant composition if areas that have failed to recruit and are colonized and dominated by Acacia. Blair et al. [17] and Bowd et al. [18] found that Acacia density was greater in post-logging regrowth compared to post-fire regrowth. The greater abundance of Acacia in post-logging regrowth forests may act as a filter, encouraging the recruitment of some species and reducing the recruitment of others depending on soil nutrition, moisture, and climate.
If a warming and drying climate leads to more frequent recruitment failure by Eucalyptus regnans in these forests, it may facilitate the development of a canopy with a greater proportion of Acacia species. The resultant increase in Acacia in the forest canopy over long periods and large areas could cause a more general decline in understorey plant diversity [78]. The Acacia may act as a filter, favouring species with particular functional traits. For example, the denser canopy cover under clumps of Acacia would likely favour slower-growing species that are more shade tolerant and have higher wood density, at the expense of species adapted to open canopy features of dominant Eucalyptus trees [16,19]. Understanding the impact of Acacia on understorey diversity in regrowth forests and at later stages of stand development in areas impacted by disturbance is an important area for future research.
4.3. Limitations, Implications, and Future Directions
The approach demonstrated here was carried out using a small, commercial UAV, that can be used with few civil aviation restrictions and can therefore be readily integrated into forest monitoring programs. Due to its size however, the UAV is limited to stand-scale assessments of ~10–20 ha per flight, which is smaller than the usual management unit in the study area (maximum coupe sizes are 40 ha in E. regnans dominated forests and 60 ha in mixed species forests [79]). This presents a limitation for management decisions at the relevant scale. Future research should focus on implementing the described methods with larger UAVs which can readily cover up to 100 ha per flight [22,80,81,82,83,84,85].
Our research highlights how cost-effective remote sensing tools can be used to explore functional linkages between canopy and understorey diversity. Commercial visible-light UAV imagery can be used to map and classify forest areas with different canopy compositions, and avoid errors associated with extrapolating plot level survey measures to broader spatial contexts. This may help forest managers to quantify and map patterns of plant diversity at the stand-scale and beyond. Such methods provide essential information on the impacts of post-harvest forest structure on plant diversity, and can identify areas where management interventions could be implemented to promote plant species diversity where high Acacia aggregation occurs. For example, thinning could be used to reduce Acacia dominance to promote increased light availability at early stages of stand development; however, such approaches must also consider the importance of Acacia as a habitat for the critically endangered Leadbeater’s possum [19,86]. The strong relationship between aridity and aggregation of Acacia and understorey beta diversity allows assessments of species occurrence to be conducted across regrowth forests in both post-fire and harvested forests. Remotely sensed mapping of biodiversity at the stand-scale can provide important insights into the impacts of climate, disturbance, and management on patterns of plant diversity in the Eucalyptus regnans forests in southeastern Australia. Determining the role of both climate and stand structure on plant community assembly during the stand initiation and stem exclusion stages of stand development [87] may provide key insights into how changing climatic conditions and management strategies will influence forest structure over time and space.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/drones7030155/s1, Supplementary Material S1 (Word document with figures and tables) and Supplementary Material S2 (geopackage of site and plot locations).
Author Contributions
A.S., P.J.B., S.K. and C.R.N. designed the project. A.S., C.R.N. and B.W. collected and analyzed the data. A.S. and B.W. drafted the manuscript with contributions from all co-authors. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the Australian Research Council (LP140100580 to C.R.N. and P.J.B., FT120100715 to P.J.B.). A.S. and B.W. were supported by the University of Melbourne through a Melbourne Research Scholarship. C.R.N. and S.K. were additionally funded through the Integrated Forest Ecosystem Research (IFER) program of the Victorian government’s Department of Energy, Environment and Climate Action. Additional funding was generously provided to A.S., S.K. and C.R.N. by Eucalypt Australia.
Data Availability Statement
Codes and tabular data supporting this research are available on the following GitHub repository: https://github.com/BennyWag/betadivdrones (accessed on 5 January 2023). As UAV imagery and resulting raster predictions are too large for online storage, it will be available upon request to the corresponding author.
Acknowledgments
We acknowledge the traditional owners of the lands on which this research was conducted, the Wurundjeri, Taungurung, and Gunaikurnai people. We pay our respects to their elders, past and present. We thank Ben Smith for fieldwork and plant ID support on this research. UAV flights were planned and executed in accordance with Australian Civil Aviation Safety Authority regulations at a maximum flight altitude of 120 m above ground and within visible line of sight. UAVs used were registered under ARN: 1044739.
Conflicts of Interest
The authors have no competing interests to declare. All authors conceived the study.
References
- Bowman, D.M.; Kolden, C.A.; Abatzoglou, J.T.; Johnston, F.H.; van der Werf, G.R.; Flannigan, M. Vegetation fires in the Anthropocene. Nat. Rev. Earth Environ. 2020, 1, 500–515. [Google Scholar] [CrossRef]
- Johnstone, J.F.; Allen, C.D.; Franklin, J.F.; Frelich, L.E.; Harvey, B.J.; Higuera, P.E.; Mack, M.C.; Meentemeyer, R.K.; Metz, M.R.; Perry, G.L. Changing disturbance regimes, ecological memory, and forest resilience. Front. Ecol. Environ. 2016, 14, 369–378. [Google Scholar] [CrossRef]
- Bratton, S.P. Resource division in an understory herb community: Responses to temporal and microtopographic gradients. Am. Nat. 1976, 110, 679–693. [Google Scholar] [CrossRef]
- Barbier, S.; Gosselin, F.; Balandier, P. Influence of tree species on understory vegetation diversity and mechanisms involved—A critical review for temperate and boreal forests. For. Ecol. Manag. 2008, 254, 1–15. [Google Scholar] [CrossRef]
- Cole, D.; Rapp, M. Elemental cycling in forest ecosystems. Dyn. Prop. For. Ecosyst. 1981, 23, 341–409. [Google Scholar]
- Wilson, S.D.; Tilman, D. Plant competition and resource availability in response to disturbance and fertilization. Ecology 1993, 74, 599–611. [Google Scholar] [CrossRef]
- Eilts, J.A.; Mittelbach, G.G.; Reynolds, H.L.; Gross, K.L. Resource heterogeneity, soil fertility, and species diversity: Effects of clonal species on plant communities. Am. Nat. 2011, 177, 574–588. [Google Scholar] [CrossRef]
- Chen, H.Y.; Légaré, S.; Bergeron, Y. Variation of the understory composition and diversity along a gradient of productivity in Populus tremuloides stands of northern British Columbia, Canada. Can. J. Bot. 2004, 82, 1314–1323. [Google Scholar] [CrossRef]
- Bartemucci, P.; Messier, C.; Canham, C.D. Overstory influences on light attenuation patterns and understory plant community diversity and composition in southern boreal forests of Quebec. Can. J. For. Res. 2006, 36, 2065–2079. [Google Scholar] [CrossRef]
- Bartels, S.F.; Chen, H.Y. Interactions between overstorey and understorey vegetation along an overstorey compositional gradient. J. Veg. Sci. 2013, 24, 543–552. [Google Scholar] [CrossRef]
- Bartels, S.F.; Chen, H.Y. Is understory plant species diversity driven by resource quantity or resource heterogeneity? Ecology 2010, 91, 1931–1938. [Google Scholar] [CrossRef]
- Getzin, S.; Wiegand, K.; Schöning, I. Assessing biodiversity in forests using very high-resolution images and unmanned aerial vehicles. Methods Ecol. Evol. 2012, 3, 397–404. [Google Scholar] [CrossRef]
- Seidl, R.; Rammer, W.; Spies, T.A. Disturbance legacies increase the resilience of forest ecosystem structure, composition, and functioning. Ecol. Appl. 2014, 24, 2063–2077. [Google Scholar] [CrossRef]
- Ashton, D. The development of even-aged stands of Eucalyptus regnans F. Muell. in central Victoria. Aust. J. Bot. 1976, 24, 397–414. [Google Scholar] [CrossRef]
- Ashton, D.; Martin, D.G. Regeneration in a pole-stage forest of Eucalyptus regnans subjected to different fire intensities in 1982. Aust. J. Bot. 1996, 44, 393–410. [Google Scholar] [CrossRef]
- Singh, A. Environmental Filtering Shapes Plant Turnover and Species Occurrence in Post-Logging Regrowth Forest in Southeastern Australia; The University of Melbourne: Melbourne, Australia, 2021. [Google Scholar]
- Blair, D.P.; McBurney, L.M.; Blanchard, W.; Banks, S.C.; Lindenmayer, D.B. Disturbance gradient shows logging affects plant functional groups more than fire. Ecol. Appl. 2016, 26, 2280–2301. [Google Scholar] [CrossRef]
- Bowd, E.J.; Lindenmayer, D.B.; Banks, S.C.; Blair, D.P. Logging and fire regimes alter plant communities. Ecol. Appl. 2018, 28, 826–841. [Google Scholar] [CrossRef]
- Trouvé, R.; Nitschke, C.R.; Andrieux, L.; Willersdorf, T.; Robinson, A.P.; Baker, P.J. Competition drives the decline of a dominant midstorey tree species. Habitat implications for an endangered marsupial. For. Ecol. Manag. 2019, 447, 26–34. [Google Scholar] [CrossRef]
- Kasel, S.; Bennett, L.T.; Aponte, C.; Fedrigo, M.; Nitschke, C.R. Environmental heterogeneity promotes floristic turnover in temperate forests of south-eastern Australia more than dispersal limitation and disturbance. Landsc. Ecol. 2017, 32, 1613–1629. [Google Scholar] [CrossRef]
- Jones, H.G.; Vaughan, R.A. Remote Sensing of Vegetation: Principles, Techniques, and Applications; Oxford University Press: Oxford, UK, 2010. [Google Scholar]
- Anderson, K.; Gaston, K.J. Lightweight unmanned aerial vehicles will revolutionize spatial ecology. Front. Ecol. Environ. 2013, 11, 138–146. [Google Scholar] [CrossRef]
- Heaphy, M.; Watt, M.S.; Dash, J.P.; Pearse, G.D. UAVs for data collection-plugging the gap. N. Z. J. For. 2017, 62, 23–30. [Google Scholar]
- Jayathunga, S.; Owari, T.; Tsuyuki, S.; Hirata, Y. Potential of UAV photogrammetry for characterization of forest canopy structure in uneven-aged mixed conifer-broadleaf forests. Int. J. Remote Sens. 2020, 41, 53–73. [Google Scholar] [CrossRef]
- Swinfield, T.; Lindsell, J.A.; Williams, J.V.; Harrison, R.D.; Agustiono; Habibi; Gemita, E.; Schönlieb, C.B.; Coomes, D.A. Accurate Measurement of Tropical Forest Canopy Heights and Aboveground Carbon Using Structure From Motion. Remote Sens. 2019, 11, 928. [Google Scholar] [CrossRef]
- Nevalainen, O.; Honkavaara, E.; Tuominen, S.; Viljanen, N.; Hakala, T.; Yu, X.; Hyyppä, J.; Saari, H.; Pölönen, I.; Imai, N.; et al. Individual Tree Detection and Classification with UAV-Based Photogrammetric Point Clouds and Hyperspectral Imaging. Remote Sens. 2017, 9, 185. [Google Scholar] [CrossRef]
- Xie, Y.; Sha, Z.; Yu, M. Remote sensing imagery in vegetation mapping: A review. J. Plant Ecol. 2008, 1, 9–23. [Google Scholar] [CrossRef]
- Dash, J.P.; Watt, M.S.; Pearse, G.D.; Heaphy, M.; Dungey, H.S. Assessing very high resolution UAV imagery for monitoring forest health during a simulated disease outbreak. ISPRS J. Photogramm. Remote Sens. 2017, 131, 1–14. [Google Scholar] [CrossRef]
- Mohan, M.; Silva, C.A.; Klauberg, C.; Jat, P.; Catts, G.; Cardil, A.; Hudak, A.T.; Dia, M. Individual tree detection from unmanned aerial vehicle (UAV) derived canopy height model in an open canopy mixed conifer forest. Forests 2017, 8, 340. [Google Scholar] [CrossRef]
- Ma, L.; Li, M.; Ma, X.; Cheng, L.; Du, P.; Liu, Y. A review of supervised object-based land-cover image classification. ISPRS J. Photogramm. Remote Sens. 2017, 130, 277–293. [Google Scholar] [CrossRef]
- He, H.S.; DeZonia, B.E.; Mladenoff, D.J. An aggregation index (AI) to quantify spatial patterns of landscapes. Landsc. Ecol. 2000, 15, 591–601. [Google Scholar] [CrossRef]
- Wagner, B.; Baker, P.J.; Nitschke, C.R. The influence of spatial patterns in foraging habitat on the abundance and home range size of a vulnerable arboreal marsupial in southeast Australia. Conserv. Sci. Pract. 2021, 3, e566. [Google Scholar] [CrossRef]
- Breiman, L. Random forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Hesselbarth, M.H.K.; Sciaini, M.; With, K.A.; Wiegand, K.; Nowosad, J. landscapemetrics: An open-source R tool to calculate landscape metrics. Ecography 2019, 42, 1648–1657. [Google Scholar] [CrossRef]
- Araujo, R.F.; Chambers, J.Q.; Celes, C.H.S.; Muller-Landau, H.C.; Santos, A.; Emmert, F.; Ribeiro, G.; Gimenez, B.O.; Lima, A.J.N.; Campos, M.A.A.; et al. Integrating high resolution drone imagery and forest inventory to distinguish canopy and understory trees and quantify their contributions to forest structure and dynamics. PLoS ONE 2020, 15, e0243079. [Google Scholar] [CrossRef]
- Egerer, M.; Wagner, B.; Lin, B.B.; Kendal, D.; Zhu, K. New methods of spatial analysis in urban gardens inform future vegetation surveying. Landsc. Ecol. 2020, 35, 761–778. [Google Scholar] [CrossRef]
- Feng, Q.L.; Liu, J.T.; Gong, J.H. UAV Remote Sensing for Urban Vegetation Mapping Using Random Forest and Texture Analysis. Remote Sens. 2015, 7, 1074–1094. [Google Scholar] [CrossRef]
- Wagner, B.; Baker, P.J.; Moore, B.D.; Nitschke, C.R. Mapping canopy nitrogen-scapes to assess foraging habitat for a vulnerable arboreal folivore in mixed-species Eucalyptus forests. Ecol. Evol. 2021, 11, 18401–18421. [Google Scholar] [CrossRef]
- Olsoy, P.J.; Forbey, J.S.; Shipley, L.A.; Rachlow, J.L.; Robb, B.C.; Nobler, J.D.; Thornton, D.H. Mapping foodscapes and sagebrush morphotypes with unmanned aerial systems for multiple herbivores. Landsc. Ecol. 2020, 35, 921–936. [Google Scholar] [CrossRef]
- Asner, G.P.; Martin, R.E.; Knapp, D.E.; Tupayachi, R.; Anderson, C.B.; Sinca, F.; Vaughn, N.R.; Llactayo, W. Airborne laser-guided imaging spectroscopy to map forest trait diversity and guide conservation. Science 2017, 355, 385–389. [Google Scholar] [CrossRef]
- Baldeck, C.A.; Asner, G.P.; Martin, R.E.; Anderson, C.B.; Knapp, D.E.; Kellner, J.R.; Wright, S.J. Operational Tree Species Mapping in a Diverse Tropical Forest with Airborne Imaging Spectroscopy. PLoS ONE 2015, 10, e0118403. [Google Scholar] [CrossRef]
- Attiwill, P.M. The disturbance of forest ecosystems: The ecological basis for conservative management. For. Ecol. Manag. 1994, 63, 247–300. [Google Scholar] [CrossRef]
- Kasel, S.; Nitschke, C.R.; Baker, S.C.; Pryde, E.C. Concurrent assessment of functional types in extant vegetation and soil seed banks informs environmental constraints and mechanisms of plant community turnover in temperate forests of south-eastern Australia. For. Ecol. Manag. 2022, 519, 120321. [Google Scholar] [CrossRef]
- Vickers, H.; Kasel, S.; Duff, T.; Nitschke, C. Recruitment and growth dynamics of a temperate forest understorey species following wildfire in southeast Australia. Dendrochronologia 2021, 67, 125829. [Google Scholar] [CrossRef]
- Stewart, S.B.; Fedrigo, M.; Roxburgh, S.H.; Kasel, S.; Nitschke, C. Climate Victoria: Precipitation (9 Second, Approx. 250 m); v1; CSIRO, 1 ed.; The University of Melbourne: Melbourne, Australia, 2020. [Google Scholar] [CrossRef]
- Stewart, S.B.; Nitschke, C.R. Improving temperature interpolation using MODIS LST and local topography: A comparison of methods in south east Australia. Int. J. Climatol. 2017, 37, 3098–3110. [Google Scholar] [CrossRef]
- Lutze, M.T.; Campbell, R.G.; Fagg, P.C. Development of silviculture in the native State forests of Victoria. Aust. For. 1999, 62, 236–244. [Google Scholar] [CrossRef]
- Anderson, M.J.; Crist, T.O.; Chase, J.M.; Vellend, M.; Inouye, B.D.; Freestone, A.L.; Sanders, N.J.; Cornell, H.V.; Comita, L.S.; Davies, K.F.; et al. Navigating the multiple meanings of β diversity: A roadmap for the practicing ecologist. Ecol. Lett. 2011, 14, 19–28. [Google Scholar] [CrossRef]
- Socolar, J.B.; Gilroy, J.J.; Kunin, W.E.; Edwards, D.P. How Should Beta-Diversity Inform Biodiversity Conservation? Trends Ecol. Evol. 2016, 31, 67–80. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package, R Package Version 2.2-0; R Foundation for Statistical Computing: Vienna, Austria, 2020.
- CASA. Flying in Public Spaces. Available online: https://www.casa.gov.au/drones/rules/public-spaces (accessed on 10 January 2020).
- Dandois, J.P.; Olano, M.; Ellis, E.C. Optimal altitude, overlap, and weather conditions for computer vision UAV estimates of forest structure. Remote Sens. 2015, 7, 13895–13920. [Google Scholar] [CrossRef]
- Roussel, J.-R.; Auty, D. lidR: Airborne LiDAR Data Manipulation and Visualization for Forestry Applications; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Pebesma, E. Simple features for R: Standardized support for spatial vector data. R J. 2018, 10, 439–446. [Google Scholar] [CrossRef]
- Hijmans, R.J. Raster: Geographic Data Analysis and Modeling, 3.6-14; R Foundation for Statistical Computing: Vienna, Austria, 2019.
- R Core Development Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- Khosravipour, A.; Skidmore, A.K.; Isenburg, M.; Wang, T.J.; Hussin, Y.A. Generating Pit-free Canopy Height Models from Airborne Lidar. Photogramm. Eng. Remote Sens. 2014, 80, 863–872. [Google Scholar] [CrossRef]
- Wing, J.; Kuhn, M.; Weston, S.; Williams, A.; Keefer, C.; Engelhardt, A.; Cooper, T.; Mayer, Z.; Kenkel, B.; R Core Development Team; et al. Caret: Classification and Regression Training, 6.0-93; R Foundation for Statistical Computing: Vienna, Austria, 2019.
- Liaw, A.; Wiener, M. Classification and Regression by randomForest. R News 2002, 2, 18–22. [Google Scholar]
- Gislason, P.O.; Benediktsson, J.A.; Sveinsson, J.R. Random forests for land cover classification. Pattern Recognit. Lett. 2006, 27, 294–300. [Google Scholar] [CrossRef]
- Pal, M. Random forest classifier for remote sensing classification. Int. J. Remote Sens. 2005, 26, 217–222. [Google Scholar] [CrossRef]
- Allouche, O.; Tsoar, A.; Kadmon, R. Assessing the accuracy of species distribution models: Prevalence, kappa and the true skill statistic (TSS). J. Appl. Ecol. 2006, 43, 1223–1232. [Google Scholar] [CrossRef]
- Fielding, A.H.; Bell, J.F. A review of methods for the assessment of prediction errors in conservation presence/absence models. Environ. Conserv. 1997, 24, 38–49. [Google Scholar] [CrossRef]
- Wang, T.; Hamann, A.; Yanchuk, A.; O’Neill, G.A.; Aitken, S.N. Use of response functions in selecting lodgepole pine populations for future climates. Glob. Change Biol. 2006, 12, 2404–2416. [Google Scholar] [CrossRef]
- Paudel, S.K.; Waeber, P.O.; Simard, S.W.; Innes, J.L.; Nitschke, C.R. Multiple factors influence plant richness and diversity in the cold and dry boreal forest of southwest Yukon, Canada. Plant Ecol. 2016, 217, 505–519. [Google Scholar] [CrossRef]
- Stewart, S.B.; Nitschke, C. Climate Victoria: Maximum Temperature (3DS; 9 Second, Approx. 250 m); v2; CSIRO; R Foundation for Statistical Computing: Vienna, Austria, 2017. [Google Scholar] [CrossRef]
- Stewart, S.B.; Nitschke, C. Climate Victoria: Minimum Temperature (3DS-TM; 9 Second, Approx. 250 m); v2; CSIRO; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar] [CrossRef]
- Fedrigo, M.; Stewart, S.B.; Roxburgh, S.H.; Kasel, S.; Bennett, L.T.; Vickers, H.; Nitschke, C.R. Predictive Ecosystem Mapping of South-Eastern Australian Temperate Forests Using Lidar-Derived Structural Profiles and Species Distribution Models. Remote Sens. 2019, 11, 93. [Google Scholar] [CrossRef]
- Barton, K. MuMIn: Multi-Model Inference, R package version 1.0.0; R Foundation for Statistical Computing: Vienna, Austria, 2009. Available online: http://r-forge.r-project.org/projects/mumin/2009 (accessed on 23 November 2019).
- Singh, A.; Baker, P.J.; Kasel, S.; Trouvé, R.; Stewart, S.B.; Nitschke, C.R. The role of climatic variability on Eucalyptus regeneration in southeastern Australia. Glob. Ecol. Conserv. 2021, 32, e01929. [Google Scholar] [CrossRef]
- Trouvé, R.; Sherriff, R.M.; Holt, L.M.; Baker, P.J. Differing regeneration patterns after catastrophic fire and clearfelling: Implications for future stand dynamics and forest management. For. Ecol. Manag. 2021, 498, 119555. [Google Scholar] [CrossRef]
- Bowman, D.M.; Murphy, B.P.; Neyland, D.L.; Williamson, G.J.; Prior, L.D. Abrupt fire regime change may cause landscape-wide loss of mature obligate seeder forests. Glob. Change Biol. 2014, 20, 1008–1015. [Google Scholar] [CrossRef]
- Doherty, M.D.; Lavorel, S.; Colloff, M.J.; Williams, K.J.; Williams, R.J. Moving from autonomous to planned adaptation in the montane forests of southeastern Australia under changing fire regimes. Austral Ecol. 2017, 42, 309–316. [Google Scholar] [CrossRef]
- Lutze, M.; Ades, P.; Campbell, R. Review of measures of site occupancy by regeneration. Aust. For. 2004, 67, 164–171. [Google Scholar] [CrossRef]
- May, B. Silver Wattle (Acacia dealbata): Its Role in the Ecology of the Mountain Ash Forest and the Effect of Alternative Silvicultural Systems on Its Regeneration. Ph.D. Thesis, Department of Botany, The University of Melbourne, Melbourne, Australia, 1999. [Google Scholar]
- Bowd, E.J.; McBurney, L.; Blair, D.P.; Lindenmayer, D.B. Temporal patterns of forest seedling emergence across different disturbance histories. Ecol. Evol. 2021, 11, 9254–9292. [Google Scholar] [CrossRef] [PubMed]
- Bauhus, J.; Van Winden, A.P.; Nicotra, A.B. Aboveground interactions and productivity in mixed-species plantations of Acacia mearnsii and Eucalyptus globulus. Can. J. For. Res. 2004, 34, 686–694. [Google Scholar] [CrossRef]
- Bowd, E.J.; McBurney, L.; Lindenmayer, D.B. The characteristics of regeneration failure and their potential to shift wet temperate forests into alternate stable states. For. Ecol. Manag. 2023, 529, 120673. [Google Scholar] [CrossRef]
- DAWE. Victorian Regional Forest Agreements Major Event Review of the 2019–20 Bushfires; Australian Government—Department of Agriculture, Water and the Environment: Melbourne, Australia, 2021.
- Goodbody, T.R.; Coops, N.C.; Marshall, P.L.; Tompalski, P.; Crawford, P. Unmanned aerial systems for precision forest inventory purposes: A review and case study. For. Chron. 2017, 93, 71–81. [Google Scholar] [CrossRef]
- Adão, T.; Hruška, J.; Pádua, L.; Bessa, J.; Peres, E.; Morais, R.; Sousa, J.J. Hyperspectral imaging: A review on UAV-based sensors, data processing and applications for agriculture and forestry. Remote Sens. 2017, 9, 1110. [Google Scholar] [CrossRef]
- Krůček, M.; Král, K.; Cushman, K.C.; Missarov, A.; Kellner, J.R. Supervised Segmentation of Ultra-High-Density Drone Lidar for Large-Area Mapping of Individual Trees. Remote Sens. 2020, 12, 3260. [Google Scholar] [CrossRef]
- Popescu, S.C.; Wynne, R.H. Seeing the trees in the forest: Using lidar and multispectral data fusion with local filtering and variable window size for estimating tree height. Photogramm. Eng. Remote Sens. 2004, 70, 589–604. [Google Scholar] [CrossRef]
- Hakkenberg, C.R.; Peet, R.K.; Urban, D.L.; Song, C. Modeling plant composition as community-continua in a forest landscape with LiDAR and hyperspectral remote sensing. Ecol. Appl. 2017, 28, 177–190. [Google Scholar] [CrossRef]
- Bunting, P.; Lucas, R. The delineation of tree crowns in Australian mixed species forests using hyperspectral Compact Airborne Spectrographic Imager (CASI) data. Remote Sens Environ. 2006, 101, 230–248. [Google Scholar] [CrossRef]
- Baker, P.J.; Nitschke, C.R.; Trouvé, R.; Robinson, A.P. Forest Stand Dynamics Drive a Conservation Conundrum for the Critically Endangered Leadbeater’s Possum. In Forests as Complex Social and Ecological Systems: A Festschrift for Chadwick D. Oliver; Baker, P.J., Larsen, D.R., Saxena, A., Eds.; Springer International Publishing: Cham, Switzerland, 2022; pp. 93–113. [Google Scholar]
- Oliver, C.D.; Larson, B.C. Forest Stand Dynamics: Updated Edition; John Wiley and Sons: Hoboken, NJ, USA, 1996. [Google Scholar]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).