
Drone Surveys Are More Accurate Than Boat-Based Surveys of Bottlenose Dolphins (Tursiops truncatus)



Abstract
:1. Introduction
2. Materials and Methods
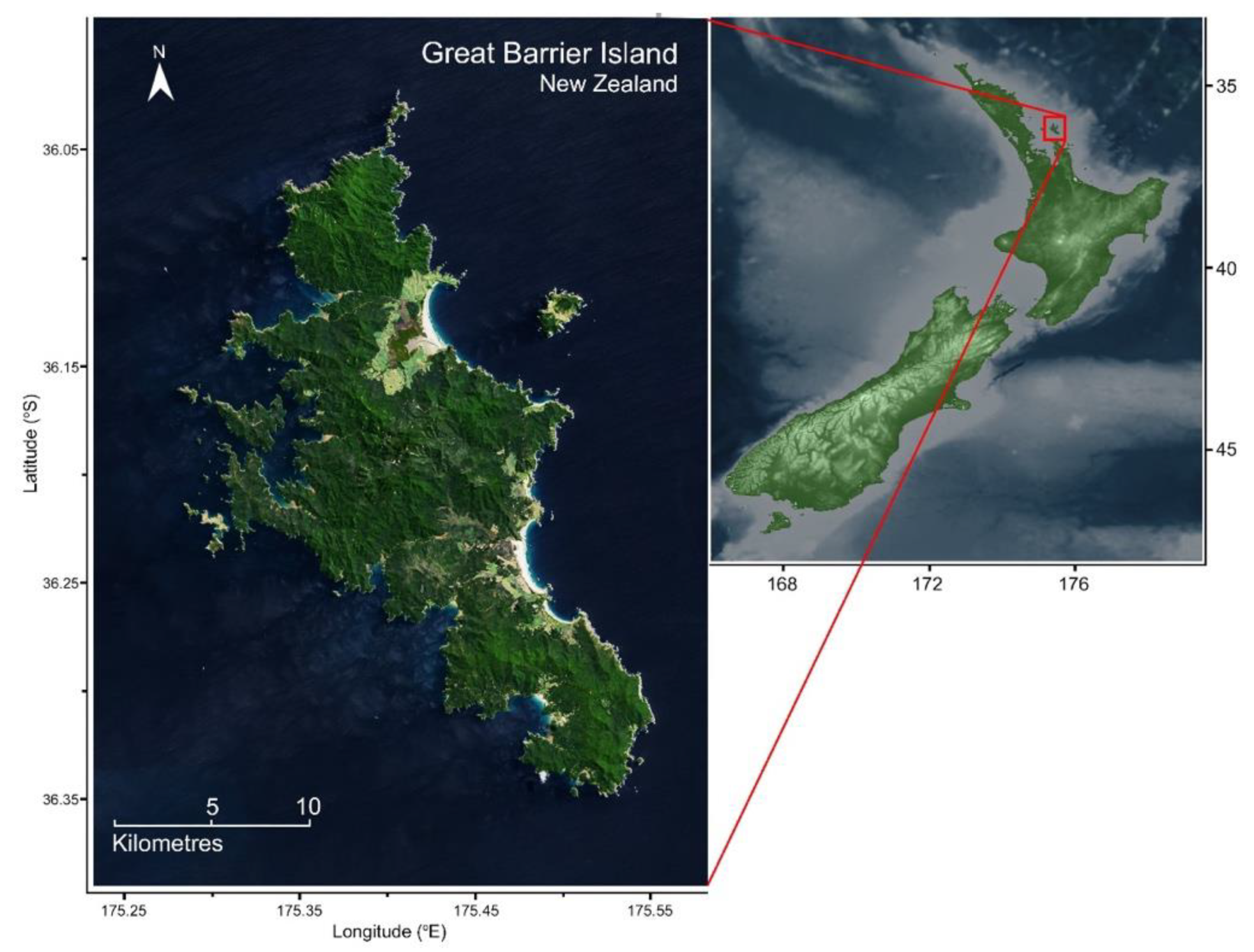
2.1. Group Counts
2.2. Behavioural Surveys
2.3. Data Analysis
3. Results
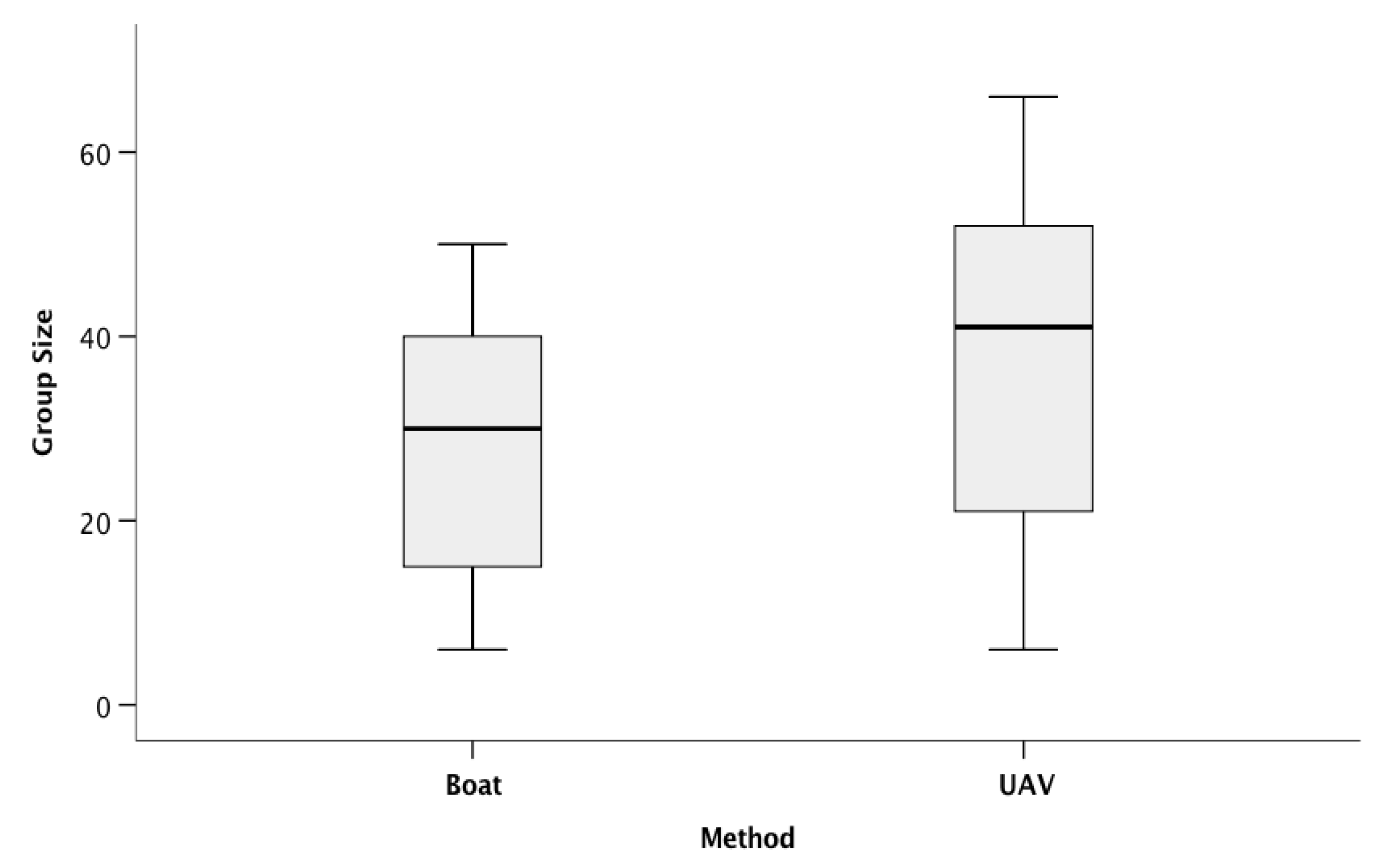
3.1. Group Counts
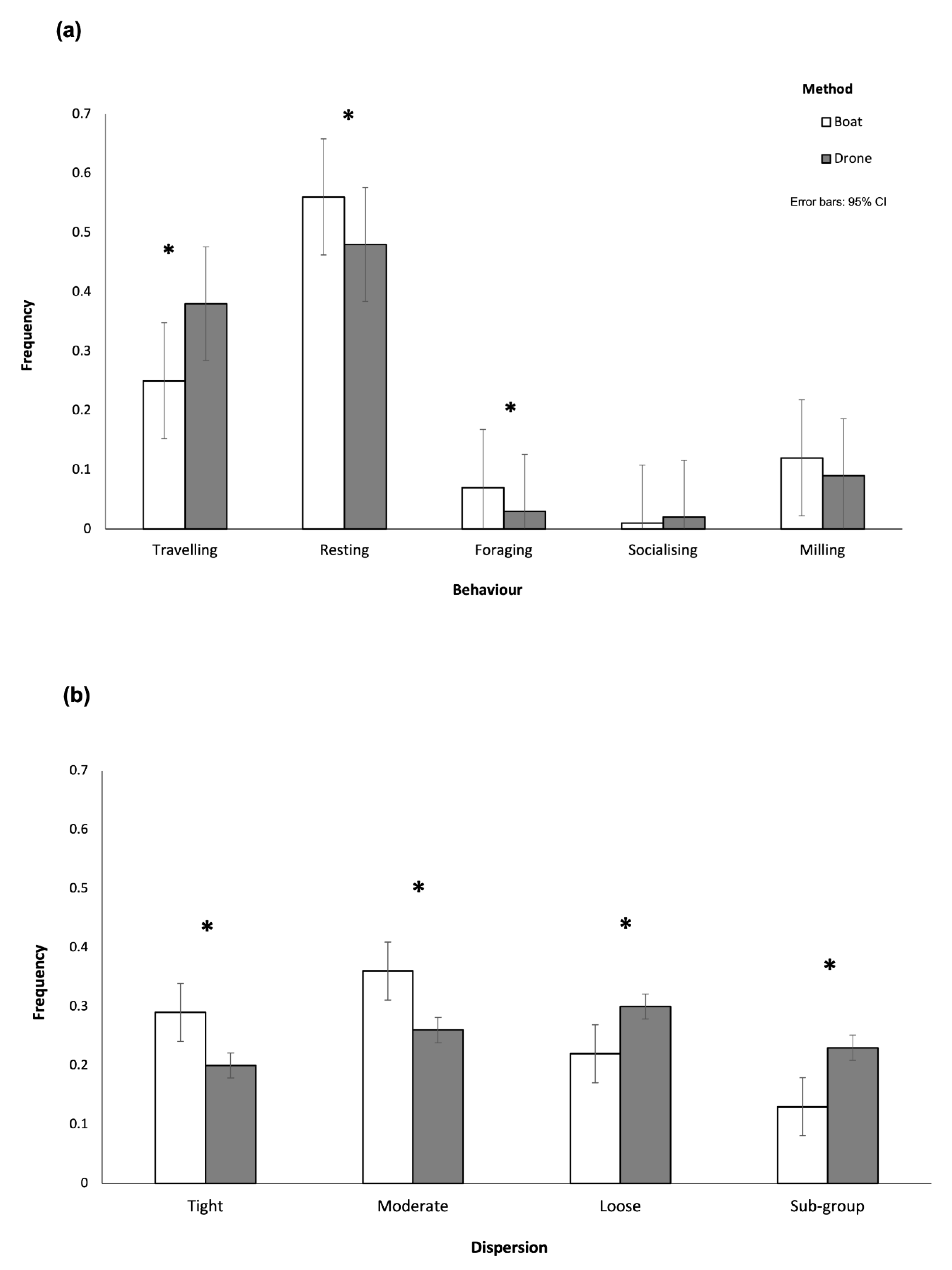
3.2. Behavioural Surveys
3.3. Additional Behavioural Events Recorded by the Drone
4. Discussion
4.1. Group Counts
4.2. Behavioural Observations
4.3. Additional Behavioural Events Detected from Drones
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Behavioural State | Boat-Based Frequency | Drone-Based Frequency | Difference (%) | p-Value |
|---|---|---|---|---|
| Resting | 0.56 | 0.48 | −15.1 | 0.024 |
| Travelling | 0.25 | 0.38 | 55.2 | <0.001 |
| Foraging | 0.07 | 0.03 | −58.3 | 0.014 |
| Socialising | 0.01 | 0.02 | 166.7 | 0.129 |
| Milling | 0.12 | 0.09 | −22.0 | 0.266 |
| Dispersion | Boat-Based Frequency | Drone-Based Frequency | Difference (%) | p-Value |
|---|---|---|---|---|
| Tight | 0.29 | 0.20 | −30.1 | 0.007 |
| Moderate | 0.36 | 0.26 | −27.0 | 0.006 |
| Loose | 0.22 | 0.30 | 35.4 | 0.017 |
| Sub-group | 0.13 | 0.23 | 80.4 | <0.001 |
References
- Fiori, L.; Martinez, E.; Bader, M.; Orams, M.B.; Bollard, B. Insights into the use of an unmanned aerial vehicle (UAV) to investigate the behavior of humpback whales (Megaptera novaeangliae) in Vava’u, Kingdom of Tonga. Mar. Mammal Sci. 2019, 36, 209–223. [Google Scholar] [CrossRef]
- Mann, J. Behavioral sampling methods for cetaceans: A review and critique. Mar. Mammal Sci. 1999, 15, 102–122. [Google Scholar] [CrossRef]
- Smultea, M.A.; Fertl, D.; Bacon, C.E.; Moore, M.R.; James, V.R.; Wursig, B. Cetacean Mother-Calf Behavior Observed from a Small Aircraft off Southern California. Anim. Behav. Cogn. 2017, 4, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Weir, J.S.; Fiori, L.; Orbach, D.N.; Piwetz, S.; Protheroe, C.; Würsig, B. Dusky Dolphin (Lagenorhynchus obscurus) Mother–Calf Pairs: An Aerial Perspective. Aquat. Mamm. 2018, 44, 603–607. [Google Scholar] [CrossRef]
- Wells, R.S.; Scott, M.D.; Irvine, A.B. The social structure of free-ranging bottlenose dolphins. In Current Mammalogy; Genoways, H.H., Ed.; Springer: Boston, MA, USA, 1987; pp. 247–305. [Google Scholar]
- Guerra, M.; Dawson, S.M.; Brough, T.E.; Rayment, W.J. Effects of boats on the surface and acoustic behaviour of an endangered population of bottlenose dolphins. Endanger. Species Res. 2014, 24, 221–236. [Google Scholar] [CrossRef] [Green Version]
- Dawson, S.; Wade, P.; Slooten, E.; Barlow, J. Design and field methods for sighting surveys of cetaceans in coastal and riverine habitats. Mamm. Rev. 2008, 38, 19–49. [Google Scholar] [CrossRef]
- May-Collado, L.J.; Quinones-Lebron, S.G. Dolphin changes in whistle structure with watercraft activity depends on their behavioral state. J. Acoust. Soc. Am. 2014, 135, 193–198. [Google Scholar] [CrossRef]
- Lusseau, D. The short-term behavioral reactions of Bottlenose dolphins to interactions with boats in Doubtful Sound, New Zealand. Mar. Mammal Sci. 2006, 22, 802–818. [Google Scholar] [CrossRef]
- Guerra, M.; Dawson, S.M. Boat-based tourism and bottlenose dolphins in Doubtful Sound, New Zealand: The role of management in decreasing dolphin-boat interactions. Tour. Manag. 2016, 57, 3–9. [Google Scholar] [CrossRef]
- Filby, N.E.; Stockin, K.A.; Scarpaci, C. Long-term responses of Burrunan dolphins (Tursiops australis) to swim-with dolphin tourism in Port Phillip Bay, Victoria, Australia: A population at risk. Glob. Ecol. Conserv. 2014, 2, 62–71. [Google Scholar] [CrossRef] [Green Version]
- Constantine, R.; Brunton, D.H.; Dennis, T. Dolphin-watching tour boats change bottlenose dolphin (Tursiops truncatus) behaviour. Biol. Conserv. 2004, 117, 299–307. [Google Scholar] [CrossRef]
- Dwyer, S.; Kozmian-Ledward, L.; Stockin, K. Short-term survival of severe propeller strike injuries and observations on wound progression in a bottlenose dolphin. N. Z. J. Mar. Freshw. Res. 2014, 48, 294–302. [Google Scholar] [CrossRef]
- Klemas, V.V. Coastal and Environmental Remote Sensing from Unmanned Aerial Vehicles: An Overview. J. Coast. Res. 2015, 31, 1260–1267. [Google Scholar] [CrossRef] [Green Version]
- Wich, S.; Dellatore, D.; Houghton, M.; Ardi, R.; Koh, L.P. A preliminary assessment of using conservation drones for Sumatran orang-utan (Pongo abelii) distribution and density. J. Unmanned Veh. Syst. 2016, 4, 45–52. [Google Scholar] [CrossRef]
- Sweeney, K.L.; Helker, V.T.; Perryman, W.L.; LeRoi, D.J.; Fritz, L.W.; Gelatt, T.S.; Angliss, R.P. Flying beneath the clouds at the edge of the world: Using a hexacopter to supplement abundance surveys of Steller sea lions (Eumetopias jubatus) in Alaska. J. Unmanned Veh. Syst. 2015, 4, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Jones, G.P.; Pearlstine, L.G.; Percival, H.F. An Assessment of Small Unmanned Aerial Vehicles for Wildlife Research. Wildl. Soc. Bull. 2006, 34, 750–758. [Google Scholar] [CrossRef]
- Clarke, P.J.; Cubaynes, H.C.; Stockin, K.A.; Olavarria, C.; de Vos, A.; Fretwell, P.T.; Jackson, J.A. Cetacean strandings from space: Challenges and opportunities of very high resolution satellites for the remote monitoring of cetacean mass strandings. Front. Mar. Sci. 2021, 8, 650735. [Google Scholar] [CrossRef]
- Goebel, M.E.; Perryman, W.L.; Hinke, J.T.; Krause, D.J.; Hann, N.A.; Gardner, S.; LeRoi, D.J. A small unmanned aerial system for estimating abundance and size of Antarctic predators. Polar Biol. 2015, 38, 619–630. [Google Scholar] [CrossRef] [Green Version]
- Koski, W.R.; Gamage, G.; Davis, A.R.; Mathews, T.; LeBlanc, B.; Ferguson, S.H. Evaluation of UAS for photographic re-identification of bowhead whales, Balaena mysticetus. J. Unmanned Veh. Syst. 2015, 3, 22–29. [Google Scholar] [CrossRef]
- Watts, A.C.; Ambrosia, V.G.; Hinkley, E.A. Unmanned Aircraft Systems in Remote Sensing and Scientific Research: Classification and Considerations of Use. Remote Sens. 2012, 4, 1671–1692. [Google Scholar] [CrossRef] [Green Version]
- Landeo-Yauri, S.S.; Ramos, E.A.; Castelblanco-Martínez, D.N.; Niño-Torres, C.A.; Searle, L. Using small drones to photo-identify Antillean manatees: A novel method for monitoring an endangered marine mammal in the Caribbean Sea. Endanger. Species Res. 2020, 41, 79–90. [Google Scholar] [CrossRef] [Green Version]
- Durban, J.W.; Fearnbach, H.; Barrett-Lennard, L.G.; Perryman, W.L.; LeRoi, D.J. Photogrammetry of killer whales using a small hexacopter launched at sea. J. Unmanned Veh. Syst. 2015, 3, 131–135. [Google Scholar] [CrossRef]
- Martins, M.C.I.; Sette, L.; Josephson, E.; Bogomolni, A.; Rose, K.; Sharp, S.M.; Niemeyer, M.; Moore, M. Unoccupied aerial system assessment of entanglement in Northwest Atlantic gray seals (Halichoerus grypus). Mar. Mammal Sci. 2019, 35, 1613–1624. [Google Scholar] [CrossRef]
- Ramp, C.; Gaspard, D.; Gavrilchuk, K.; Unger, M.; Schleimer, A.; Delarue, J.; Landry, S.; Sears, R. Up in the air: Drone images reveal underestimation of entanglement rates in large rorqual whales. Endanger. Species Res. 2021, 44, 33–44. [Google Scholar] [CrossRef]
- Acevedo-Whitehouse, K.; Rocha-Gosselin, A.; Gendron, D. A novel non-invasive tool for disease surveillance of free-ranging whales and its relevance to conservation programs. Anim. Conserv. 2010, 13, 217–225. [Google Scholar] [CrossRef]
- Domínguez-Sánchez, C.A.; Acevedo-Whitehouse, K.A.; Gendron, D. Effect of drone-based blow sampling on blue whale (Balaenoptera musculus) behavior. Mar. Mammal Sci. 2018, 34, 841–850. [Google Scholar] [CrossRef]
- Centelleghe, C.; Carraro, L.; Gonzalvo, J.; Rosso, M.; Esposti, E.; Gili, C.; Bonato, M.; Pedrotti, D.; Cardazzo, B.; Povinelli, M.; et al. The use of Unmanned Aerial Vehicles (UAVs) to sample the blow microbiome of small cetaceans. PLoS ONE 2020, 15, e0235537. [Google Scholar] [CrossRef]
- Raudino, H.C.; Tyne, J.A.; Smith, A.; Ottewell, K.; McArthur, S.; Kopps, A.M.; Chabanne, D.; Harcourt, R.G.; Pirotta, V.; Waples, K. Challenges of collecting blow from small cetaceans. Ecosphere 2019, 10, e02901. [Google Scholar] [CrossRef] [Green Version]
- Horton, T.W.; Hauser, N.; Cassel, S.; Klaus, K.F.; Fettermann, T.; Key, N. Doctor Drone: Non-Invasive Measurement of Humpback Whale Vital Signs Using Unoccupied Aerial System Infrared Thermography. Front. Mar. Sci. 2019, 6, 466. [Google Scholar] [CrossRef] [Green Version]
- Dawson, S.M.; Bowman, M.H.; Leunissen, E.; Sirguey, P. Inexpensive Aerial Photogrammetry for Studies of Whales and Large Marine Animals. Front. Mar. Sci. 2017, 4, 366. [Google Scholar] [CrossRef] [Green Version]
- Christiansen, F.; Dawson, S.M.; Durban, J.W.; Fearnbach, H.; Miller, C.A.; Bejder, L.; Uhart, M.; Sironi, M.; Corkeron, P.; Rayment, W.; et al. Population comparison of right whale body condition reveals poor state of the North Atlantic right whale. Mar. Ecol. Prog. Ser. 2020, 640, 1–16. [Google Scholar] [CrossRef]
- Durban, J.W.; Moore, M.J.; Chiang, G.; Hickmott, L.S.; Bocconcelli, A.; Howes, G.; Bahamonde, P.A.; Perryman, W.L.; LeRoi, D.J. Photogrammetry of blue whales with an unmanned hexacopter. Mar. Mammal Sci. 2016, 32, 1510–1515. [Google Scholar] [CrossRef]
- Ramos, E.A.; Maloney, B.; Magnasco, M.O.; Reiss, D. Bottlenose Dolphins and Antillean Manatees Respond to Small Multi-Rotor Unmanned Aerial Systems. Front. Mar. Sci. 2018, 5, 316. [Google Scholar] [CrossRef]
- Oliveira-Da-Costa, M.; Marmontel, M.; Da-Rosa, D.S.X.; Coelho, A.; Wich, S.; Mosquera-Guerra, F.; Trujillo, F. Effectiveness of unmanned aerial vehicles to detect Amazon dolphins. Oryx 2019, 54, 696–698. [Google Scholar] [CrossRef] [Green Version]
- Orbach, D.N.; Eaton, J.; Fiori, L.; Piwetz, S.; Weir, J.S.; Würsig, M.; Würsig, B. Mating patterns of dusky dolphins (Lagenorhynchus obscurus) explored using an unmanned aerial vehicle. Mar. Mammal Sci. 2020, 36, 1097–1110. [Google Scholar] [CrossRef]
- Torres, L.G.; Nieukirk, S.L.; Lemos, L.; Chandler, T.E. Drone Up! Quantifying Whale Behavior from a New Perspective Improves Observational Capacity. Front. Mar. Sci. 2018, 5, 319. [Google Scholar] [CrossRef] [Green Version]
- Frouin-Mouy, H.; Tenorio-Hallé, L.; Thode, A.; Swartz, S.; Urbán, J. Using two drones to simultaneously monitor visual and acoustic behaviour of gray whales (Eschrichtius robustus) in Baja California, Mexico. J. Exp. Mar. Biol. Ecol. 2020, 525, 151321. [Google Scholar] [CrossRef]
- Torres, L.G.; Barlow, D.R.; Chandler, T.E.; Burnett, J.D. Insight into the kinematics of blue whale surface foraging through drone observations and prey data. PeerJ 2020, 8, e8906. [Google Scholar] [CrossRef] [Green Version]
- Baker, C.S.; Bore, L.; Childerhouse, S.; Constantine, R.; van Helden, A.; Lundquist, D.; Rayment, W.; Rolfe, J.R. Conservation status of New Zealand marine invertebrates, 2019. N. Z. J. Mar. Freshw. Res. 2010, 44, 129–148. [Google Scholar] [CrossRef]
- Tezanos-Pinto, G.; Constantine, R.; Brooks, L.; Jackson, J.; Mourão, F.; Wells, S.; Scott Baker, C. Decline in local abundance of bottlenose dolphins (Tursiops truncatus) in the Bay of Islands, New Zealand. Mar. Mammal Sci. 2013, 29, 390–410. [Google Scholar] [CrossRef]
- Tezanos-Pinto, G.; Baker, C.S.; Russell, K.; Martien, K.; Baird, R.W.; Hutt, A.; Stone, G.; Mignucci-Giannoni, A.A.; Caballero, S.; Endo, T.; et al. A Worldwide Perspective on the Population Structure and Genetic Diversity of Bottlenose Dolphins (Tursiops truncatus) in New Zealand. J. Hered. 2009, 100, 11–24. [Google Scholar] [CrossRef] [Green Version]
- Constantine, R. The Behavioural Ecology of the Bottlenose Dolphins (Tursiops truncatus) of Northeastern New Zealand: A Population Exposed to Tourism; The University of Auckland: Auckland, New Zealand, 2002. [Google Scholar]
- Dwyer, S.L.; Tezanos-Pinto, G.; Visser, I.N.; Pawley, M.D.M.; Meissner, A.M.; Berghan, J.; Stockin, K.A. Overlooking a potential hotspot at Great Barrier Island for the nationally endangered bottlenose dolphin of New Zealand. Endanger. Species Res. 2014, 25, 97–114. [Google Scholar] [CrossRef] [Green Version]
- Whitehead, H.; Reeves, R.R.; Tyack, P.L. Science and the Conservation, Protection, and Management of Wild Cetaceans. In Cetacean Societies: Field Studies of Dolphins and Whales; Mann, J., Connor, R.C., Tyack, P.L., Whitehead, H., Eds.; The University of Chicago Press: Chicago, IL, USA, 2000; pp. 308–332. [Google Scholar]
- Berger-Tal, O.; Polak, T.; Oron, A.; Lubin, Y.; Kotler, B.P.; Saltz, D. Integrating animal behavior and conservation biology: A conceptual framework. Behav. Ecol. 2011, 22, 236–239. [Google Scholar] [CrossRef] [Green Version]
- Barros, N.B.; Wells, R.S. Prey and feeding patterns of resident Bottlenose dolphins (Tursiops truncatus) in Sarasota Bay, Florida. J. Mammal. 1998, 79, 1045–1059. [Google Scholar] [CrossRef]
- Rogan, E.; Ingram, S.; Holmes, B.; O’Flanagan, C.; Institute, M. A Survey of Bottlenose Dolphins (Tursiops truncatus) in the Shannon Estuary; National University of Ireland: Cork, Ireland, 2000. [Google Scholar]
- Whitehead, H. Analysis of animal movement using opportunistic individual identification: Application to sperm whales. Ecology 2001, 82, 1417–1732. [Google Scholar] [CrossRef]
- Fiori, L.; Doshi, A.; Martinez, E.; Orams, M.B.; Bollard-Breen, B. The Use of Unmanned Aerial Systems in Marine Mammal Research. Remote Sens. 2017, 9, 543. [Google Scholar] [CrossRef] [Green Version]
- Shane, S.H. Behavior and ecology of the bottlenose dolphin at Sanibel Island, Florida. In The Bottlenose Dolphin; Leatherwood, S., Reeves, R.R., Eds.; Academic Press, Inc.: New York, NY, USA, 1990; pp. 245–265. [Google Scholar]
- Fettermann, T.; Fiori, L.; Bader, M.; Doshi, A.; Breen, D.; Stockin, K.A.; Bollard, B. Behaviour reactions of bottlenose dolphins (Tursiops truncatus) to multirotor Unmanned Aerial Vehicles (UAVs). Sci. Rep. 2019, 9, 8558. [Google Scholar] [CrossRef] [Green Version]
- Altmann, J. Observational study of behavior: Sampling methods. Behaviour 1974, 49, 227–267. [Google Scholar] [CrossRef] [Green Version]
- Lusseau, D. Why do dolphins jump? Interpreting the behavioural repertoire of bottlenose dolphins (Tursiops sp.) in Doubtful Sound, New Zealand. Behav. Process. 2006, 73, 257–265. [Google Scholar] [CrossRef]
- Shane, S.H.; Wells, R.S.; Wursig, B. Ecology, behavior and social organization of the bottlenose dolphin: A review. Mar. Mammal Sci. 1986, 2, 34–63. [Google Scholar] [CrossRef]
- Kelaher, B.P.; Peddemors, V.M.; Hoade, B.; Colefax, A.P.; Butcher, P.A. Comparison of sampling precision for nearshore marine wildlife using unmanned and manned aerial surveys. J. Unmanned Veh. Syst. 2019, 8, 30–43. [Google Scholar] [CrossRef]
- Sorrell, K.; Clarke, R.H.; Holmberg, R.; McIntosh, R.R. Remotely piloted aircraft improve precision of capture–mark–resight population estimates of Australian fur seals. Ecosphere 2019, 10, e02812. [Google Scholar] [CrossRef] [Green Version]
- Hodgson, J.C.; Baylis, S.M.; Mott, R.; Herrod, A.; Clarke, R.H. Precision wildlife monitoring using unmanned aerial vehicles. Sci. Rep. 2016, 6, 22574. [Google Scholar] [CrossRef] [Green Version]
- Wilson, B.; Hammond, P.S.; Thompson, P.M. Estimating size and assessing trends in a coastal bottlenose dolphin population (Tursiops truncatus). Ecol. Appl. 1999, 9, 288–300. [Google Scholar] [CrossRef]
- Hupman, K.; Stockin, K.A.; Pollock, K.; Pawley, M.D.M.; Dwyer, S.L.; Lea, C.; Tezanos-Pinto, G. Challenges of implementing Mark-recapture studies on poorly marked gregarious delphinids. PLoS ONE 2018, 13, e0198167, Correction in PLoS ONE 2018, 13, e0203356. [Google Scholar] [CrossRef]
- Hooker, S.K.; Gerber, L.R. Marine Reserves as a Tool for Ecosystem-Based Management: The Potential Importance of Megafauna. Bioscience 2004, 54, 27–39. [Google Scholar] [CrossRef]
- Mann, J.; Connor, R.C.; Tyack, P.; Whitehead, H. Field studies of dolphins and whales. In Cetacean Societies; The University of Chicago Press: Chicago, IL, USA, 2000; p. 433. [Google Scholar]
- Peters, C.H.; Stockin, K.A. Response of Bottlenose Dolphin (Tursiops truncatus) to Vessel Activity in Northland, New Zealand; Massey University: Auckland, New Zealand, 2016. [Google Scholar]
- Hartel, E.F.; Constantine, R.; Torres, L.G. Changes in habitat use patterns by bottlenose dolphins over a 10-year period render static management boundaries ineffective. Aquat. Conserv. Mar. Freshw. Ecosyst. 2014, 25, 701–711. [Google Scholar] [CrossRef]
- Tayler, C.K.; Saayman, G.S. The social organisation and behaviour of dolphins (Tursiops aduncus) and baboons (Papio ursinus): Some comparisons and assessments. Ann. Cape Prov. Museums 1972, 9, 11–49. [Google Scholar]
- Leatherwood, S.; Evans, W.E. Some recent uses and potentials of radiotelemetry in field studies of cetaceans. In Behaviour of Marine Mammals: Current Perspectivies in Research; Olla, H.E.W., Olla, B.L., Eds.; Plenum Press: New York, NY, USA, 1979; Volume 3. [Google Scholar]
- Bearzi, G.; Politi, E. Diurnal behavior of free-ranging bottlenose dolphins in the Kvarneric (Northern Adriatic Sea). Mar. Mammal Sci. 1999, 15, 1065–1097. [Google Scholar] [CrossRef]
- Hodgson, A.; Peel, D.; Kelly, N. Unmanned aerial vehicles for surveying marine fauna: Assessing detection probability. Ecol. Appl. 2017, 27, 1253–1267. [Google Scholar] [CrossRef]
- Constantine, R. Increased avoidance of swimmers by wild Bottlenose dolphins (Tursiops truncatus) due to long-term exposure to swim-with-dolphin tourism. Mar. Mammal Sci. 2001, 17, 689–702. [Google Scholar] [CrossRef]
- Nowacek, D.P.; Christiansen, F.; Bejder, L.; Goldbogen, J.A.; Friedlaender, A.S. Studying cetacean behaviour: New technological approaches and conservation applications. Anim. Behav. 2016, 120, 235–244. [Google Scholar] [CrossRef] [Green Version]
- Giles, A.B.; Butcher, P.A.; Colefax, A.P.; Pagendam, D.E.; Mayjor, M.; Kelaher, B.P. Responses of bottlenose dolphins (Tursiops spp.) to small drones. Aquat. Conserv. Mar. Freshw. Ecosyst. 2020, 31, 677–684. [Google Scholar] [CrossRef]
- Hartman, K.; van der Harst, P.; Vilela, R. Continuous Focal Group Follows Operated by a Drone Enable Analysis of the Relation Between Sociality and Position in a Group of Male Risso’s Dolphins (Grampus griseus). Front. Mar. Sci. 2020, 7, 283. [Google Scholar] [CrossRef]
- Nielsen, M.L.K.; Sprogis, K.R.; Bejder, L.; Madsen, P.T.; Christiansen, F. Behavioural development in southern right whale calves. Mar. Ecol. Prog. Ser. 2019, 629, 219–234. [Google Scholar] [CrossRef]
- Booth, C.G.; Sinclair, R.R.; Harwood, J. Methods for Monitoring for the Population Consequences of Disturbance in Marine Mammals: A Review. Front. Mar. Sci. 2020, 7, 115. [Google Scholar] [CrossRef] [Green Version]
- Erbe, C. Effects of underwater noise on marine mammals. Adv. Exp. Med. Biol. 2012, 730, 17–22. [Google Scholar] [CrossRef]
- Pirotta, E.; Merchant, N.D.; Thompson, P.M.; Barton, T.R.; Lusseau, D. Quantifying the effect of boat disturbance on bottlenose dolphin foraging activity. Biol. Conserv. 2015, 181, 82–89. [Google Scholar] [CrossRef]



| Behavioural State | Definition |
|---|---|
| Socialising | Dolphins observed chasing, copulating, and/or engaged in any other physical contact with another dolphin, such as rubbing and touching (excluding mother-calf pairs). Aerial behavioural events, such as horizontal and vertical jumps, may occur. |
| Milling | Dolphins exhibited non-directional movements, with frequent changes in bearing that prevented dolphins from making headway in any specific direction. Most of the time appears to be ‘transition’ behaviour between behavioural states. |
| Foraging | Dolphins involved in any effort to pursue, capture, and/or consume prey. Diving for long periods of time, showing repeated unsynchronised dives in different directions in a determined location, exhibiting behaviour such as fluke out dives. |
| Resting | Dolphins observed in a tight group, engaged moving slowly and in a constant direction. Surfacing is generally more predictable, often more synchronous than observed in other behavioural states. |
| Travelling | Dolphins engaged in persistent, directional movement, making noticeable headway along a specific compass bearing. Group space varies and individuals swim with short and relatively constant dive intervals. |
| Group Dispersion | Definition |
|---|---|
| Tight | Less than one body length between individuals. |
| Moderate | One to five body lengths between individuals. |
| Loose | More than five body lengths between individuals. |
| Sub-group | Dolphins are divided in two or more groups but act as part of a single pod. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fettermann, T.; Fiori, L.; Gillman, L.; Stockin, K.A.; Bollard, B. Drone Surveys Are More Accurate Than Boat-Based Surveys of Bottlenose Dolphins (Tursiops truncatus). Drones 2022, 6, 82. https://doi.org/10.3390/drones6040082
Fettermann T, Fiori L, Gillman L, Stockin KA, Bollard B. Drone Surveys Are More Accurate Than Boat-Based Surveys of Bottlenose Dolphins (Tursiops truncatus). Drones. 2022; 6(4):82. https://doi.org/10.3390/drones6040082
Chicago/Turabian StyleFettermann, Ticiana, Lorenzo Fiori, Len Gillman, Karen A. Stockin, and Barbara Bollard. 2022. "Drone Surveys Are More Accurate Than Boat-Based Surveys of Bottlenose Dolphins (Tursiops truncatus)" Drones 6, no. 4: 82. https://doi.org/10.3390/drones6040082
APA StyleFettermann, T., Fiori, L., Gillman, L., Stockin, K. A., & Bollard, B. (2022). Drone Surveys Are More Accurate Than Boat-Based Surveys of Bottlenose Dolphins (Tursiops truncatus). Drones, 6(4), 82. https://doi.org/10.3390/drones6040082