Bacterial Consortiums Able to Use Metal-Cyanide Complexes as a Nitrogen Source †


 and
and {kind=link}
Abstract
:1. Introduction
2. Materials and Methods
3. Results
4. Discussion
Acknowledgments
Conflicts of Interest
References
- Akcil, A. Destruction of cyanide in gold mill effluents: Biological versus chemical treatments. Biotechnol. Adv. 2003, 21, 501–511. Available online: http://www.sciencedirect.com/science/article/pii/S0734975003000995 (accessed on 16 October 2018). [CrossRef]
- Vogel, S.N.; Sultan, T.R.; Ten Eyck, R.P. Cyanide Poisoning. Clin. Toxicol. 1981, 18, 367–383. Available online: https://doi.org/10.3109/15563658108990043 (accessed on 16 October 2018). [CrossRef] [PubMed]
- Rico, M.; Benito, G.; Salgueiro, A.R.; Díez-Herrero, A.; Pereira, H.G. Reported tailings dam failures: A review of the European incidents in the worldwide context. J. Hazard. Mater. 2008, 152, 846–852. Available online: http://www.sciencedirect.com/science/article/pii/S0304389407010837 (accessed on 16 October 2018). [CrossRef] [PubMed]
- Park, J.M.; Trevor Sewell, B.; Benedik, M.J. Cyanide bioremediation: The potential of engineered nitrilases. Appl. Microbiol. Biotechnol. 2017, 101, 3029–3042. [Google Scholar] [CrossRef] [PubMed]
- Luque-Almagro, V.M.; Huertas, M.J.; Martínez-Luque, M.; Moreno-Vivián, C.; Roldán, M.D.; García-Gil, L.J.; Castillo, F.; Blasco, R. Bacterial degradation of cyanide and its metal complexes under alkaline conditions. Appl. Environ. Microbiol. 2005, 71, 940–947. Available online: http://www.scopus.com/inward/record.url?eid=2-s2.0-13644269308&partnerID=40&md5=c1165cb3147cce49650bed7731dcac1d (accessed on 16 October 2018). [CrossRef] [PubMed]
- Sambrook, J.; Rusell, D.W. Molecular Cloning: A Laboratory Manual, 3rd ed.; Cold Spring Harbor Laboratory: New York, NY, USA, 2001. [Google Scholar]
 ) and the cyanide concentration in the culture supernatant (
) and the cyanide concentration in the culture supernatant (  ) were measured. The concentration of cyanide in a noninoculated flask containing culture medium (
) were measured. The concentration of cyanide in a noninoculated flask containing culture medium (  ) was measured at the same time.
) and the cyanide concentration in the culture supernatant ( ) were measured. The concentration of cyanide in a noninoculated flask containing culture medium ( ) was measured at the same time.
) was measured at the same time.
) and the cyanide concentration in the culture supernatant ( ) were measured. The concentration of cyanide in a noninoculated flask containing culture medium ( ) was measured at the same time.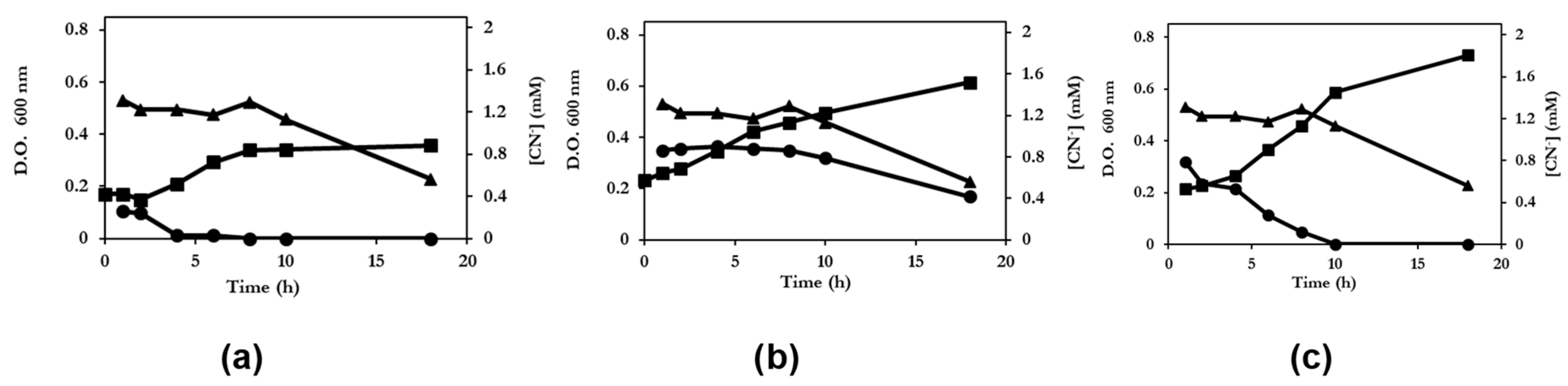
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Igeño, M.I.; Macías, D.; Guijo, M.I.; Sánchez-Clemente, R.; Población, A.G.; Merchán, F.; Blasco, R. Bacterial Consortiums Able to Use Metal-Cyanide Complexes as a Nitrogen Source. Proceedings 2018, 2, 1284. https://doi.org/10.3390/proceedings2201284
Igeño MI, Macías D, Guijo MI, Sánchez-Clemente R, Población AG, Merchán F, Blasco R. Bacterial Consortiums Able to Use Metal-Cyanide Complexes as a Nitrogen Source. Proceedings. 2018; 2(20):1284. https://doi.org/10.3390/proceedings2201284
Chicago/Turabian StyleIgeño, María Isabel, Daniel Macías, María Isabel Guijo, Rubén Sánchez-Clemente, Ana G. Población, Faustino Merchán, and Rafael Blasco. 2018. "Bacterial Consortiums Able to Use Metal-Cyanide Complexes as a Nitrogen Source" Proceedings 2, no. 20: 1284. https://doi.org/10.3390/proceedings2201284
APA StyleIgeño, M. I., Macías, D., Guijo, M. I., Sánchez-Clemente, R., Población, A. G., Merchán, F., & Blasco, R. (2018). Bacterial Consortiums Able to Use Metal-Cyanide Complexes as a Nitrogen Source. Proceedings, 2(20), 1284. https://doi.org/10.3390/proceedings2201284