Abstract
Rice is a major crop for half of the world’s population, and nitrogen (N) fertilizers play a crucial role in its production. However, imbalanced N fertilizer uses and traditional fertilization practices have led to low nitrogen use efficiency (NUE), increased N footprints, and reduced rice yields and farmers’ income. There are limited studies where the integration of both agronomic and molecular advancements to enhance NUE is discussed, particularly in developing countries. This review highlights novel agronomic and molecular strategies to enhance NUE, rice yields, and profitability, while minimizing environmental impact. The agronomic strategies include the 4R Nutrient Stewardship framework, enhanced efficiency nitrogen fertilizers (EENFs), nano-fertilizers, biochar-based fertilizers, biological N fixation, and sensor-based fertilizer management in major rice-growing countries. The molecular mechanisms focus on N uptake, assimilation, and utilization, highlighting the role of hormones, key genes, transcription factors (TFs), and regulatory pathways. Moreover, we examine promising rice genotypes and cultivars with improved NUE and grain yield. Additionally, this paper offers deep insights into recent advancements in molecular genetics, such as multi-omics approaches (transcriptomics, metabolomics, and metagenomics), the Genome-Wide Association Study (GWAS), Quantitative Traits Loci mapping (QTLs), Single Nucleotide Polymorphisms (SNPs) analysis, and Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR-Cas9)-mediated genome editing, which serve as valuable tools for developing rice cultivars with enhanced NUE and grain yield.
1. Introduction
Rice serves as a staple food for half of the world’s population, with an annual production of milled rice of around 480 million metric tons. About 90% of global rice is produced and consumed in Asia, making its production in this region crucial for global food security. Notably, China and India together account for approximately 50% of rice production and consumption. Together with China and India, other Asian countries such as Bangladesh, Vietnam, Myanmar, Thailand, the Philippines, Japan, Pakistan, Cambodia, the Republic of Korea, Nepal, and Sri Lanka account for 90% of global rice production. Moreover, rice is a significantly source of energy for 50% of people in Asia living in poverty [1,2,3,4]. Its significance extends beyond mere nourishment, intertwining with human civilizations, cultures, and traditions. Nitrogen (N) is crucial for synthesizing nucleotides, amino acids, and chlorophyll [5], making it an essential fertilizer for enhancing crop production. Nitrogen (N) fertilizer is intensively used in modern agriculture to increase crop production and meet growing food demands driven by rapid population growth, often without considering NUE and the associated environmental costs. Prolonged application of fertilizers and organic manures differentially influence mineral N and organic N fractions and total N in soils reduces NUE, leading to significant losses through volatilization, leaching, and runoff, emitting greenhouse gases like N₂O, which contribute to global warming and threaten food security [6,7,8].
Global nitrogen fertilizer consumption exceeds 110 million tons annually, with China and India accounting for nearly half [9]. Rice alone consumes “30 to 50%” of total agricultural nitrogen [10]. With the cost of chemical fertilizers rising and likely to increase further in the future, it is essential to prioritize improving nutrient usage efficiency through the adoption of best management practices (BMPs), resulting in reducing fertilizer costs and contributing to significant yield gaps [11]. Additionally, climate change increases crop N concentrations while reducing internal nitrogen use efficiency (iNUE). A meta-analysis of single-season irrigated rice across different zone and climatic conditions in China (northern middle and lower river basin, southwestern) found N concentrations rose by 5–9.6%, while iNUE dropped by 9–14%. To meet food demands, and as the population is predicted to be 9.7 billion [12,13] by 2050, iNUE must increase by 7.8–13.6% [14]. Another meta-analysis (1961 to 2013) in China and India showed N use in cereal production has increased significantly, ranging from 10 to 84%, but NUE declined to 20–24% by 2013. NUE in major rice-growing countries remains below 40%, lagging 15–20% behind the global average [15,16]. Despite decades of research on improving NUE, progress remains limited, primarily due to blanket fertilization recommendations that overlook site-specific needs, limited soil testing, imbalanced fertilization practices, fertilizer subsidies, and low adoption of BMPs across farming communities. Additionally, the lack of widespread adoption of proven research technologies has led to significant spatial-temporal variations in NUE in many developing countries. Despite a slight global NUE increase from 33% in 1999 to 35% in 2015, over 60% of applied N is still lost through leaching and emissions via volatilization and denitrification, which are primarily attributed to excessive application of N fertilizer and increased mechanization [9]. To meet global nitrogen sustainability goals, NUE must rise to 70–80% through precision nutrient management (PNM) and BMPs, reducing N losses, cutting costs, and improving yields [17].
Some of the promising and scalable fertilizer management practices to improve NUE include 4R Nutrient Stewardship (right time, right place, right method, and right proportion), enhanced efficiency fertilizers (EENFs), nano-fertilizers, biochar-enriched N fertilizers, and sensor-based precision nutrient management. Moreover, conservation agriculture (CA) practices, such as intercropping, green manuring, cover crops, and crop rotation, enhance biological nitrogen fixation (BNF) and reduce N input, ensuring climate-resilient production systems. Additionally, rice root architecture and genetic variability in NUE, particularly between indica and japonica varieties, offer opportunities for crop breeding and genetic improvement [18,19]. A combined agronomic and genetic approach is essential for sustainable NUE enhancement. Similarly, multi-disciplinary collaboration among researchers, farmers, and stakeholders is key to advancing smart fertilizer management technologies. Integrating genomic tools like GWAS, SNPs, genome editing, and multi-omics approaches can enhance NUE by identifying key traits for resilient rice cultivars, contributing to SDGs 2 (zero hunger) and 13 (climate action) [20].
In Asian countries, over 90% of rice is produced by smallholder farmers [21]. Fertilizer use among these farmers is heavily skewed toward excessive N application, while the use of phosphorous (P), potassium, K, and other secondary nutrients and micronutrients remain insufficient. This imbalance results in low NUE and is largely driven by high subsidies for urea and limited adoption of BMPs. Inefficient N fertilization is responsible for nearly 50–66% of the gap between actual and potential rice yields [22]. Additionally, the intensive use of inputs such as fertilizer and irrigation during the green revolution era has degraded soil health and reduced fertilizing efficiency. To narrow this yield gap and meet the growing food demands of a rising population, there is an urgent need to promote and scale efficient fertilizer management strategies that improve NUE and are widely adopted by farmers. Integrated studies on agronomic and molecular approaches are essential for enhancing NUE and yet such studies remain limited in many developing countries. Through this review, we aim to provide a comprehensive understanding of NUE in rice by synthesizing advancements from both agronomic and molecular perspectives. By bridging the knowledge gap between these two domains, this paper offers valuable insights into emerging innovations and outlines future research directions to support sustainable rice production. The structure of this review includes (i) an overview of nitrogen use, its efficiency, and yield gaps in rice production, (ii) innovative and scalable agronomic strategies to enhance NUE, (iii) molecular and genetic mechanisms of N uptake, assimilation and utilization, and (iv) molecular advancement and genomic tools that contribute to the development of superior rice cultivars aimed at enhancing NUE, yield, and sustainability. This review will serve as a valuable resource for researchers, academics, and stakeholders in the public–private sectors by presenting practical approaches and research-based strategies to enhance NUE in rice production systems.
2. Concepts of NUE
NUE, the ratio of grain yield to nitrogen applied, is determined by N uptake efficiency (NUpE) and N utilization efficiency (NUtE), where NUpE is the total amount of N taken up by the plant relative to the N supplied, and NUtE is the ratio of total grain yield to the N in the plant. NUE is influenced by various factors, and different forms of NUE can be calculated using the following equation [23].
- Recovery efficiency of N (REN) = (UNN − UN0)/(FN − FN0), kg N uptake per kg N applied;
- Agronomic efficiency of N (AEN) = (GYN − GY0)/(FN − FN0), kg grain yield increase per kg N applied;
- Partial factor productivity of N (PFPN) = GYN/FN, kg grain yield per kg N applied;
- Physiological efficiency of applied N (PEN) = (GYN − GY0)/(UNN − UN0), kg grain yield increase per kg N uptake;
- Internal efficiency of N (IEN) = GYN/UNN, kg grain per kg N uptake;
- Utilization efficiency (UEN) = Physiological efficiency × Apparent N recovery, kg kg−1, where GYN is the total grain yield when N is applied, FN is total N fertilizer applied, GY0 is total grain yield without N application, UNN is total N uptake with N treatment, and UN0 is total N uptake without nitrogen.
Generally, PEN ranges from 30 to 60 kg/kg, PFPN from 40 to 70 kg/kg, AEN from 10 to 30 kg/kg, and REN from 30 to 50% [24]. The recovery efficiency of applied nitrogen (REN) in rice varies from 35.4 to 55% (average 43.9%) in northern Thailand in acidic sandy soils under both irrigated and rainfed conditions [25] and 48% in Nepal in silty loam soils under rainfed conditions [23]. Moreover, meta-analysis 1333 field experiments conducted across varying soil types and biophysical factors in China revealed only 28.3% of REN in rice, which was primarily attributed to over-use of chemical fertilizer [26].
Blanket N applications, often surface-broadcasted, lead to low AEN and REN due to volatilization [27,28], especially when applied early during underdeveloped root stages [21]. NUE is influenced by various factors such as fertilizer type, rate, timing, rice variety, soil properties, crop management, and interaction with environmental factors.
The IRRI has developed strategies to improve NUE, including variety selection, biological N fixation (BNF), deep placement techniques, and chlorophyll meters for optimal N application [29]. Optical sensors, such as a chlorophyll meter like the Soil–Plant Analysis Development (SPAD) and crop reflectance sensors, enable optimized N application by allowing precise timing and dosages. For instance, the use of the SPAD meter in irrigated rice cultivation in India exhibited threshold values of 37 in south India and 37.5 in northwest India, indicating the plant nitrogen status and supporting precision nitrogen management strategies [30,31]. Moreover, in the context of BNF, free-living blue-green algae (BGA) and azolla are key sources for fixing atmospheric N and reducing the need for external chemical N fertilizer in rice fields, maintaining similar or even higher yields. Additionally, site-specific nutrient management (SSNM) remains crucial for increasing NUE, reducing N losses and enhancing productivity [32,33,34].
3. Efficient Fertilizer Management in Farmlands
In this section, some of the innovative, promising, and scalable agronomic N management strategies are highlighted (Figure 1) and discussed, along with their agronomic (NUE and yields) and economic benefits in Table 1.
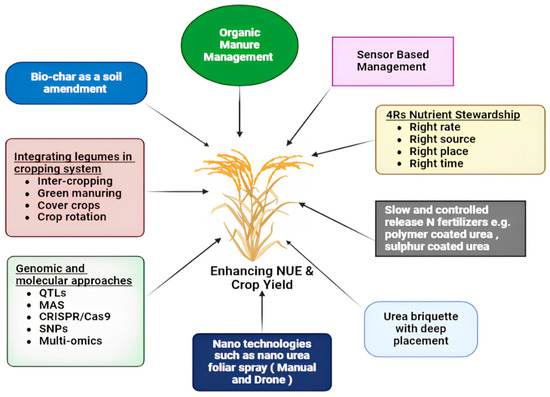
Figure 1.
Strategies and approaches to promote and scale efficient nitrogen management to enhance nitrogen use efficiency in rice.
3.1. The 4Rs Nutrient Stewardship Framework
Optimizing NUE and yields can be achieved by adopting the 4Rs technology: applying fertilizer at the right rate, source, place, and time, often termed as “fertilizer BMPs” [35,36,37]. This approach, as shown in Figure 1, integrates both organic and inorganic fertilizers, ensuring balanced fertilization based on soil, crops, and environmental factors.
Urea (46% N) is widely used in rice due to its high N content and affordability compared to ammonium sulfate (21% N). Multilocation and multiyear fertilizer trials with varying N rates, considering agro-ecological zones, soil types, indigenous nutrient supply, crop varieties, climate, and crop management, are crucial for identifying optimal N rates to achieve attainable yields and NUE [33,38]. Timely application also plays a key roles: Splitting a 100 kg N ha−1 application into three doses increased total N uptake by 42%, AEN by 146%, and rice yield (variety: Chenab basmati-2016) by 44% compared to a single basal application over two years in medium alkaline sandy clay loam soil under tropical conditions at the University of Agriculture, Faisalabad, Pakistan (Table 1) [39]. Application methods impact efficiency, with deep placement reducing N losses by 50% compared to surface broadcasting [40]. Field trials and demonstrations conducted across nine districts in Nepal ranging from tropical to sub-tropical regions have shown higher NUE, crop yields, income, and the growing interest of farmers in the adoption of 4Rs [35,41,42]. However, despite farmers’ understanding from farmer schools and training that the 4Rs can offset production cost through increased yields, widespread adoption is constrained by recurrent fertilizer shortages, limited farmers’ knowledge, and inadequate access to fertilizer-distributing cooperatives [42].
Although the 4Rs have been practiced for 30 years, the emerging 4R Nutrient Stewardship (4RNS) framework integrates cropping systems, landscapes, and sustainability metrics. The 4RNS framework is the adaptive management of plant mineral nutrients through the refinement of interconnected strategies based on location, soil, climate, and crop species. As 4RNS is still in the developmental stage, further research is needed to refine best practices for long-term soil health, water quality, and economic viability, emphasizing collaborative and multidisciplinary efforts [36,43].
3.2. Enhanced Efficiency Nitrogen Fertilizers (EENFs)
Enhanced efficiency nitrogen fertilizers (EENFs) are a key component of 4R Nutrient Stewardship, helping reduce N losses and improve NUE [44,45]. These fertilizers include slow and controlled-release coated fertilizers, nitrification, urease inhibitors, and urea briquettes (UB) with deep placement (UDP), etc. Coated fertilizers like polymer-coated urea (PCU) and sulfur-coated urea (SCU) enable slow-release mechanisms, providing nutrients to plants throughout the crop cycle and minimizing losses from volatilization, denitrification, and leaching [46,47,48,49,50,51]. In addition, EENFs are economically viable due to the single-time application, which saves time and labor cost compared to multiple splits of conventional urea [51]. One-time applications of PCU reduced labor requirements by six workers per hectare compared to two splits of urea application in rice fields in Nepal [49].
PCU has been shown to increase rice yield by 1.0–1.3 t ha−1 and agronomic NUE by 5.2–5.9 kg/kg compared to conventional urea. Additionally, it enhances root morphological traits [52]. Controlled-release urea led to enhanced NUE and grain yield in rice by increasing the activity of nitrogen metabolism enzymes such as Nitrate Reductase (NR), Glutamine Synthetase (GS), and Glutamate Synthase (GOGAT) by 50–200%, and upregulating key genes (OsNIA, OsGS1, OsGS2, OsGLN1.1, and OsGLN1.2) [53].
Urea briquette (UB) deep placement involves compressing prilled or granular urea into large briquettes (1–3 g) and placing them 7–10 cm below the soil surface [49]. This technique improves nitrogen retention, making it available during critical crop growth stages [23,54,55]. Studies show that UB can reduce N input by 20–35% while increasing crop yield by 25% and improving PFPN and AEN by 44% and 34%, respectively [23,56]. Research from Fujian Province, China, found that deep placement of slow-release fertilizers increased rice yield by 8.8–23.5% while cutting CH₄ emissions by 25.34% and N₂O emissions by 48.97%, as shown in Table 1 [57].
Controlled-release nitrogen fertilizers (CRNFs) offer both economic and environmental benefits. In southwest China, CRNFs improved environmental sustainability by 2.82–4.61%, NUE by up to 43.96%, and profitability by 5.21–11.44%. [58]. Similarly, in southeast China, slow-release fertilizer with deep placement increased NUE by 41.47%, economic benefits by 76.43%, net energy, and ecological benefits by 43.83%, while reducing cumulative CH4 accumulation by 25.34% [59].
3.3. Nanotechnology
Nanotechnology is an innovative and emerging technology to achieve SDGs by enhancing NUE and promoting sustainable agricultural practices under the circumstances of climate change and resource depletion. Nano-fertilizers are the nutrient carriers produced by encapsulating plant nutrients within nano-materials that are less than 100 nm in size [60,61]. Nano-fertilizers are considered more eco-friendly than conventional fertilizers as they are foliar spayed, which can minimize N losses through leaching and emissions, thereby reducing environmental pollution and ensuring sustainable production systems [60,62]. Nano-fertilizers improve nutrient absorption through the plant cuticle and are characterized by their excellent solubility and reactivity, enabling precise nutrient delivery to plant cells and efficient nitrogen utilization [63]. Nano-fertilizers have higher surface area to volume ratio and surface functionalization, which can reduce nutrient losses and release N for prolonged periods compared to conventional urea [64,65]. While the use of nano-fertilizers has shown promising results, their impact on specific crops and diverse agroecological zones remains unexplored [66]. Therefore, future studies should focus on understanding the mechanistic effects of nano-urea on various crops, including rice, across diverse biophysical conditions such as soil types, temperature, rainfall, and topography.
One of the most significant developments is the nano-urea produced by the Indian Farmers Fertilizer Cooperative (IFFCO). Given its higher NUE (80–90%), IFFCO claimed a 500 mL nano-urea bottle equivalent to a 45 kg sack of conventional urea. As nano-urea is applied in smaller quantities, it reduces the need for large quantities of urea, minimizing nitrogen losses, and improving crop productivity [67]. Results suggest that nano-fertilizers enhanced tomato yield by 30% and lowered nitrogen use by 50% compared to conventional urea, demonstrating tangible benefits for crop production [68]. Applying 75% of conventional nitrogen along with nano-nitrogen in wheat enhances NUE and crop yield, achieving a benefit–cost ratio of 5.51 [69]. Similarly, co-application of urea (50% as basal) and nano-urea two sprays at tillering and panicle initiation stages in rice significantly increased leaf area index, panicle weight, grain yield and NUE compared to other fertilizer treatments [70]. Similarly, in another study, urea 75% N and two sprays of nano-urea N showed significantly higher crop yields, NUE and reduced GHGs over 100 N [65].
The underlying mechanism is that the nano-urea particles are absorbed through stomatal uptake or surface absorption and the nutrients are slowly released and metabolized within the plant cells to support growth and development. These results demonstrate that nano-urea can reduce nitrogen input by 25 to 50%, fostering more sustainable agricultural practices. In addition to enhancing NUE, nano-urea is an eco-friendly product that supports improved soil health, reduces N leaching and emissions, and contributes to better air and water quality, thereby aiding in the mitigation of global warming compared to conventional urea. However, small amounts of nano-particles may leach from plant surfaces into the soil, although this is significantly lower than the N inputs from direct soil application of conventional urea in soil. This reduced N input may influence soil biogeochemical processes, altering nutrient dynamics and microbial activity. It is also more affordable to farmers compared to conventional urea, requiring less time and labor for application, which can reduce overall production cost and increase farmers’ income [71,72].
Although nanotechnology offers some promising results, it lacks comprehensive documentation on its utilization, potential, and application in agriculture. Additionally, it requires more careful consideration for its safe handling to minimize potential human health and environmental risks [61].
3.4. Digital and Sensor-Based Technologies for Enhancing Fertilizer Use Efficiency
The integration of digital and sensor-based technologies is transforming agriculture by enabling precision nutrient management and real-time fertilizer application, optimizing nitrogen (N) use, reducing nutrient losses, input costs, and enhancing crop yields. Technologies such as real-time sensors, drones, satellite remote sensing, GIS-GPS, and variable rate technology (VRT) provide precise fertilizer application during critical growth stages, boosting NUE and yields [73]. Proximal sensing and ICTs, like NDVI and Green Seeker, detect nitrogen stress and optimize N application rates [23,73,74]. Use of Green Seeker reduced N input by 54% without compromising rice yields and increased AEN, PFPN, REN, and UEN [23].
Digital tools such as digital soil maps (DSM) and apps like Crop Doctor offer precise fertilizer recommendations, improving NUE, farm profitability, and reducing environmental impact [75]. Real-time data obtained from various digital and sensor-based approaches can be fitted into the modeling applications to determine site-specific fertilizer management. For instance, soil fertility information in DSM can be utilized in the QUEFTS model (Quantitative Evaluation of Fertility of Tropical Soils) to determine site-specific fertilizer recommendations [76]. Variable-rate fertilizer application (VRFA) using a DSM in Kharagpur, India, was helpful in identifying optimum rates required for attainable yields while minimizing input cost and pollution [77]. However, sensor-based fertilizer management faces several challenges, including difficulties in sensor deployment, inconsistencies in data quality, and requirements for sustainable reliable energy sources, all of which underscore the complexity of integrating such technologies into precision farming systems [78]. Therefore, scaling up these digital and sensor-based methods at the farm level requires collaboration, hands-on training, and demonstration regarding its implementation among farmers and stakeholders to transition to precision nitrogen management and enhance NUE [73].
3.5. Conjoint Application of Biochar and Nitrogenous Fertilizers
Biochar, a carbon-rich material produced through pyrolysis, is globally known for enhancing soil fertility and mitigating climate change [79]. The combined application of biochar and N fertilizers has emerged as an effective strategy to enhance NUE, reduce N losses, and increase crop yields [80,81,82,83]. Studies have shown that biochar positively influences rice yield, harvest index, and straw production, with AEN increasing by 140% and grain nutrient recovery (REN) rising by 191% over two years [82].
Biochar’s high porosity, large surface area, negative surface charges, and high cation exchange capacity (CEC) contribute to enhanced N retention, improved NUE, and reduced N2O emissions. These effects are primarily due to biochar’s ability to promote electrostatic adsorption and retention of NH4+, while also minimizing NO3- leaching by increasing soil water-holding capacity and decreasing infiltration rates [84]. Moreover, biochar applications can mitigate N losses by reducing NH3 volatilization and N2O emissions by suppressing the enzymatic activities of urease and nitrate/nitrite reductase, inhibiting the activity of denitrifying microbes and enhancing N immobilization by soil microorganisms through the supply of labile carbon and the creation of a favorable microhabitat. Applying rice straw biochar (5 to 10 t ha−1) significantly improves soil N availability and nutrient levels, resulting in higher NUE and rice productivity [85]. Enriching biochar with organic and inorganic fertilizers increases its effectiveness [86,87,88,89], as seen in Nepalese soils where biochar enriched with cattle urine boosted NPK uptake and crop NUE [86]. The enhanced agronomic performance of nutrient-enriched biochar can be attributed to the organic coatings formed on biochar, which restrict water movement, enhance nutrient retention, and enable a slow release mechanism [90]. Additionally, biochar is economically viable, with a benefit–cost ratio of 1.47 in southern China, and it mitigates CO2 emissions by 6.22 tons per ton applied [91]. Unlike other organic and mineral fertilizers, biochar is typically applied only once, which helps reduce variable costs. Its recalcitrant nature allows it to persist in the soil for hundreds to thousands of years, offering long-term agronomic and environmental benefits.
3.6. Biological N Fixation
Biological nitrogen fixation (BNF) is the process by which specific microorganisms convert atmospheric nitrogen (N2) into ammonia (NH3) or similar compounds, making nitrogen available for plants. BNF contributes 17% of total N inputs in soil ecosystems globally [92], and can reduce reliance on chemical N fertilizers [93]. Microorganisms like diazotrophic iron-reducing bacteria play a key role in BNF, particularly in paddy fields. Cyanobacteria species such as Anabaena variabilis and Nostoc muscorum enhance soil N levels and NUE in rice by reducing ammonia volatilization [94]. Combined use of BNF and 50% chemical fertilizers can increase rice yields by 18.62% and NUE by 43.83% [94].
BNF practices like green manuring, cover cropping, and intercropping help reduce the need for chemical fertilizers, improve soil health, and increase crop productivity. Leguminous plants such as Sesbania rostrata and Crotalaria juncea are used as green manure, releasing nitrogen into the soil for subsequent crops. These leguminous green plants decompose and release nutrients, especially nitrogen for subsequent crops [95]. Approximately 40–60% of total N contained in legumes is available to succeeding crops, mostly in organic forms. The release of available N for successive crops largely depends on the presence of effective soil microorganisms, biological activity, C:N ratio, soil type, moisture, and temperature [96]. Most green manure N is mineralized within 4 weeks of incorporation [97].Green manuring can reduce nitrogen fertilizer use by 20–40%, minimizing nitrogen losses and fertilizer costs. For example, green manure with 20% less N (80 N) increased rice yield and nutrient recovery compared to full nitrogen application [98].
Another promising BNF practice is the adoption of intercropping, where cereals (main crops) are primarily combined with legumes (component crops) in definite cropping geometry to improve soil fertility and yields. In Asian region, farmers predominantly cultivate single crops, particularly cereals, throughout the year, leading to limited nutrient recycling, substantial soil nutrient depletion, and adverse effects on soil fertility and crop productivity. Due to rising population pressures and decreasing arable land, crop intensification is crucial for addressing food security issues and achieving SDGs. Intercropping offers benefits such as improved soil fertility, reduced fertilizer inputs, enhanced NUE, and increased yields [99,100]. For instance, intercropping rice with Neptunia oleracea reduced N inputs by 22% and improved yields compared to sole-cropping with conventional N [101]. Legumes fix N and contribute to nutrient recycling, thus supporting sustainable agriculture and improved food security [102]. While BNF is an innovative and profitable approach for crop production, its successful implementation depends on the availability of skilled labor, appropriate technology, and essential resources. However, labor shortages and limited access to key agri-inputs such as seeds, fertilizers, biofertilizers like azolla, and BGA poses significant barriers to the adoption of green manuring and intercropping in many developing countries.

Table 1.
Comparative analysis highlighting enhancements in NUE, grain yield, economic benefits, and N losses with the utilization of different technologies and major cereal crops across different countries.
Table 1.
Comparative analysis highlighting enhancements in NUE, grain yield, economic benefits, and N losses with the utilization of different technologies and major cereal crops across different countries.
| S.N. | Technologies | Country | Crop | NUE Increment | Grain Yield Increment | Economic Benefits | Reduction in Emissions | References |
|---|---|---|---|---|---|---|---|---|
| 1. | 4Rs Technology (right timing- three spilt doses) | Pakistan | Rice | 42 (N uptake), 146 (AFN), | 44% | Reduction in cost of production (31–43%) | [39] | |
| 2. | 4Rs Technology (right timing) | Ethiopia | Rice | 31.4 (PFPN) | 5.6% | Reduction in N fertilizer use by 31.4% | [40] | |
| 3. | 4Rs | Nepal | Maize | 35% | 40% | [35] | ||
| 4. | Polymer-coated urea | China | Rice | 5.2 to 5.9 kg/kg | 1.0 to 1.3 t ha−1 | [52] | ||
| 5. | Urea briquette (UB) | Nepal | Rice | 25% | [23] | |||
| 6. | Deep placement of urea super granules (USGs) | Bangladesh | Rice | NUE −13%, agronomic NUE- 20%, recovery N use efficiency −19% | 5.22% | [103] | ||
| 7. | Slow-release fertilizer | China | Rice | N recovery efficiency by 62.50–91.57% and 24.38–64.24% (two rice cultivars Yongxian15 and Yongyou1540, respectively) | 6.30–11.64% and 6.23–13.11% (two rice cultivars Yongxian15 and Yongyou1540, respectively) | CH4 emissions by 25.34% and N20 emissions by 48.97% | [57] | |
| 8. | Controlled release urea + conventional urea | China | Rice | 24.4% | 25.3% | [104] | ||
| 9. | Polymer-coated urea | China | Wheat | 3–34% | Reduce N volatilization loss (23–62%), ammonia emissions (51.3–91.3%) | [105] | ||
| 10. | Controlled release urea | China | Rice | 0.65–43.96% | 5.21–11.44% | [58] | ||
| 11. | Controlled release urea with deep placement | China | Rice | 41.47% | 76.43% | CH4 accumulation by 25.34% | [59] | |
| 12. | Joint use of PCU and urea briquette | Nepal | Maize and rice | 46% | 23% | [49] | ||
| 13. | Nano-urea | India | Maize, wheat | 80–90% | [67] | |||
| 14. | Application of nano-urea in flooding-irrigated conditions | Bangladesh | Rice | 95.38% | 4.91% | [106] | ||
| 15. | Application of N 150 kg/ha along with nano-urea (4%) in wheat | India | Wheat | 7.8% | Gross return of $2542 and cost–benefit of 2.01 | [107] | ||
| 16. | Co-joint application of biochar (8 t/ha) | India | Peas rabi season and maize kharif season | Benefit–cost ratio of 1.47 | One ton of biochar mitigates 6.22 tonnes of CO2 | [91] | ||
| 17. | NFC combined with 50% chemical fertilizers | China | Rice | 18.62% | 43.83% | NH3 volatilization losses by 41.63–45.34% | [94] |
4. Molecular Mechanisms of Nitrogen Uptake and Assimilation
4.1. Root System Architecture (RSA) and Genes Associated with Improving RSA
Root functions influence NUE in plants to absorb nitrogen from soil, and the distribution and availability of nitrogen play a crucial role in shaping root system architecture (RSA) [108,109]. Rice fibrous root systems consist of seminal and adventitious roots. Primary roots extend deeper into the soil to acquire nitrogen, while lateral roots spread out to cover a larger surface area, encompassing a greater soil volume [110]. Moreover, rice root morphology includes the number of roots, weight, total length, surface area, root branches, root diameter, and angle [29]. These root morphological traits, as extended root length, greater volume, high density, and increased surface area, significantly increase nitrogen absorption [111]. Nitrogen absorption from the soil is actively facilitated by specialized transporter proteins responsible for nitrogen uptake. The inorganic nitrogen form, NO3− (nitrate), is primarily located in the rhizosphere, where its concentration is significantly higher than that of NH4+ (ammonium) [112]. Nitrogen absorption efficiency is directly proportional to the size and shape of roots. It has been observed that nitrates, a common form of nitrogen in soil, are highly prone to leaching. So, improving the root system’s architecture, such as its depth and density, enhances the nitrogen absorption rate [113]. NUE is significantly affected by plant root’s ability to uptake and utilize nitrogen from external sources.
The availability and distribution of these sources alter crop RSA to optimize nitrogen utilization. RSA and specific gene expression possess a significant role in nitrogen uptake and grain yield in low nitrogen conditions. For example, overexpression of the OsAMT1;1 and OsAMT1;2 genes increased root length and surface area and encoded ammonium transporters enhancing nitrogen accumulation capability, leading to higher grain productivity in rice variety YD6 [114]. Research suggests that RNR10, F-box protein, is a crucial gene that controls nitrogen-responsive RSA in rice, and provides new tools to improve NUE by manipulating the RNR10-DNR1-auxin module [109]. Quantitative trait locus (QTL) qRDWN6XB provides new insights for a potential genetic resource for improving rice RSA through marker-assisted selection (MAS) under low nitrogen conditions, thereby enhancing NUE and grain yield [115]. The OsMADS57 gene elongates roots and modulates nitrate translocation from rice roots to shoots under nitrogen-deprived conditions. Results suggest that the over-expression of this gene increases nitrate translocation twofold, thereby enhancing NUE [116]. Upregulation of the OsNPF2.4 gene enhances root surface area, length, and volume, and carbon and nitrogen translocation, synergistically improving NUE and grain yield in the dep1 rice line [117]. Application of biochar in the seedling stage of rice showed up-regulation of N-metabolism genes such as OsAMT1.1, OsGS1.1, and OsGS2, enhancing root system and nitrate uptake by 33.60% [118]. The use of slow-release nitrogen fertilizer (sulfur-coated urea) in rice exhibited significant improvement in root development, leaf structure, NUE, and grain yield [119].
4.2. Hormonal Regulation for Enhancing NUE
Comprehensive transcriptomic analysis of the OsAAP1 gene suggests that OsAAP1 may have a crucial role in nitrogen absorption and metabolism through signaling pathways of auxin, cytokinin, and strigolactone, which are essential for regulating tillering in rice [120]. An expression of DNR1 protein in indica rice varieties has antagonistic effects on auxin regulation, NUE, and crop yield. For instance, lower DNR1 abundance enhances auxin synthesis, activating nitrate transporter and nitrogen metabolism-related genes through the auxin response factor (ARF) mechanisms, enhancing both NUE and grain yield [121]. Similarly, results suggest that auxin plays a crucial role in root growth, N uptake, and assimilation in maize. The application of naphthalene acetic acid (NAA, 0.1 mmol/L) exhibited improvements in plant photosynthetic rate and biomass by 58.6% and 91.7%, respectively. Moreover, the activities of key N assimilation enzymes NR, GS, and GOGAT were enhanced by 32.3%, 22.9%, and 16.2% [122]. Enhanced lateral root elongation through auxin signaling under lower N conditions in wheat increases NUE by boosting crop nitrate acquisition capacity [123].
Gibberellins (GAs) influence the expression of genes involved in nitrogen assimilation, such as NR and GS, which play vital roles in nitrogen metabolism and enhance NUE [124]; in addition, GAs have recently been reported to be involved in saline–alkaline stress management in rice through increased ammonium uptake [125]. GAs regulates the expression of ZmNRT2.1 and ZmNRT2.2 in maize, thereby promoting N uptake [126]. The expression of primary NPF (nitrate transporter1/peptide transporter family) genes, specifically NPF6.3 and NRT2.1, regulated by auxin, ethylene, and cytokinin play a role in influencing nitrogen uptake [126,127]. Cytokinin modifies root behavior in rice in response to high ammonium levels [128] as the overexpression of the cytokinin activation enzyme-like gene OsLOGL5 has a vital role in primary root growth, tiller number, yield, and NUE under various conditions, including irrigated conditions, water stress, nitrogen sufficiency, nitrogen deficiency, etc. [129,130]. Brassinosteroids (BRs) play a key role in plant growth, development, and responses to environmental stresses and nutrient utilization in maize. Research suggests that BRs regulate the physiological response, and nitrogen uptake in maize enhances the growth of primary roots and plant biomass and up-regulates the expression of nitrogen transporter genes [131,132]. A nitrate, GA, and ABA transporter gene OsNPF3.1 are reported to regulate NUE [133].
4.3. Mechanisms of Nitrogen Uptake and Assimilation Through Transporters
Nitrate (NO3-) transporters are the most important components for facilitating the uptake, transport, assimilation, and re-mobilization of nitrogen, resulting in enhanced NUE. These transporters belong to four major gene families: NRT1/NPF (Nitrate transporter 1 or peptide transporter), NRT2/NNP (Nitrate transporter 2/Nitrate–Nitrite porter), CLC (Chloride channel), and SLAC1/SLAH (slow anion channel-associated homologs) [134,135]. The physiology of NO3− uptake can be described by low- and high-affinity transport. Popularly known as the high-affinity transport system (HATS) and low-affinity transport system (LATS), both of these mechanisms have been observed in Arabidopsis (Arabidopsis thaliana). For instance, AtNRT2.1 is a HATS transporter that facilitates the uptake of nitrate from the soil, particularly when nitrate concentrations are low. On the other hand, the AtNRT1.1 transporter can operate both low and high nitrogen affinity depending upon the environmental conditions. Similarly, chlorate-resistant 1/nitrate transporter1 (CHL1/NRT1.1), NPF6.3, also operates as both a low- and high-affinity transporter [134,136,137]. The NRT2 family, in contrast to the NRT1 family, consists of high-affinity NO3- transporters. Among the eight members of the NRT2 family, seven have been characterized. NRT2.1, NRT2.2, NRT2.4, and NRT2.5 are actively involved in the influx of NO3- into Arabidopsis root cells [138,139]. Similarly, in the high-affinity nitrate transporter family, OsNRT2.3 (Table 2 generates two transcripts, OsNRT2.3A, and OsNRT2.3B, via alternative splicing primarily expressed in the plant’s phloem. OsNRT2.3A mediates the long-distance transportation of nitrate from the root to the stem and encodes a phloem protein located at amino acid position 516 [140,141]. Similarly, research elucidates that OsNRT1.1B, OsSPX4, and OsNLP3 genes have a significant role in regulating nitrate uptake from the soil, influencing rice productivity [142].
Ammonia (NH4+) predominantly exists in flooded and acidic soil conditions [143]. High-affinity transporters mainly located in the root hairs and epidermis, known as ammonia-mediated transporters (AMT), regulate its transport [144]. The Arabidopsis genome contains six AMT, each coding for high-affinity NH4+ transporters upregulated in low nitrogen conditions [145]. Among the six AMTs, AMT1;1, AMT1;2, and AMT1;3 play critical roles in the high-affinity NH4+ absorption, showing that plants deploy diverse NH4+ transporters to efficiently uptake NH4+ under conditions of low nitrogen availability [144]. AtAMT1;1, AtAMT1;2, and AtAMT1;3 are predominantly found in roots, whereas AtAMT1;1 and AtAMT1;2 can also be expressed in shoots, with higher expression observed in leaves. The enhanced uptake of ammonia is co-related to the up-regulation of AtAMT1;1 and AtAMT1;3 under nitrogen deficiency [146,147]. OsAMT1;1 is expressed on both roots and shoots. The mutants of OsAMT1;1 showed a decrease in ammonium uptake as well as impeded growth of roots and shoots. OsAMT1;2 expressed exclusively on the roots and OsAMT1;3 on nitrogen deficiency roots. Moreover, the expression of OsAMT1 genes does not depend on endogenous ammonium levels but its expression is regulated by glutamine concentration, where glutamine synthase is a key enzyme converting inorganic ammonium into glutamine [147,148,149,150]. Moreover, Nin-like protein 4 (NLPs) and circadian clock genes have been reported in transcriptional regulation of the OsAMT1 gene, which is associated with N uptake under N-deficient conditions in rice [151].
4.4. Nitrogen Assimilation and Utilization Pathways
The first step in nitrate assimilation includes the activation of nitrate reductase (NR) and its involvement in the reduction and conversion of nitrate into biologically active forms like nucleic acids, amino acids, and other nitrogen-containing biomolecules within cytoplasm. Along with NR, a molybdenum cofactor (MoCo) also plays a crucial role in catalyzing nitrate reduction. The next step includes the conversion of nitrate into ammonia (NO2− to NH4+) by nitrite reductase in the chloroplast [143,152,153,154].
The GS-GOGAT (Figure 2) cycles are major regulatory pathways that have a significant role in nitrogen assimilation and remobilization, with various metabolic processes leading to enhanced NUE and crop yield [155]. Ammonium can be absorbed by plant roots, formed through the conversion of nitrates, or released during the breakdown of amino acids or through photorespiration. The breakdown and conversion processes are facilitated by two major enzymes, GS and GOGAT, interactively known as the GS/GOGAT cycle. GS catalyzes the conversion of ammonia and glutamate to glutamine, GOGAT transfers the amide amino group from glutamine to 2-oxoglutarate, forming glutamate. Cytosolic glutamine synthetase (GS1) is crucial in assimilating inorganic nitrogen from the soil, such as nitrate or NH4+. Furthermore, GS1 plays a role in N reassimilation from NH4+ and enhanced expression during senescence. An integration of the chimeric alfalfa NADH-GOGAT gene into tobacco showed higher total carbon and nitrogen content in shoots and increased shoot dry weight at the flowering stage, reflecting a higher capacity to assimilate nitrogen [155,156,157].
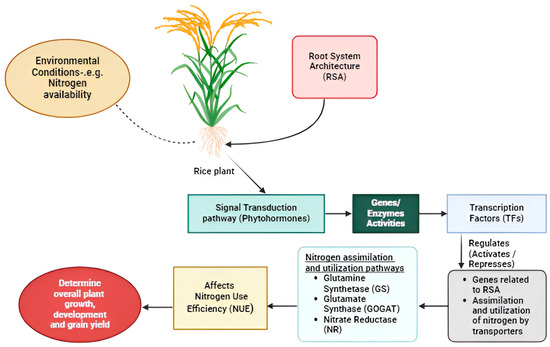
Figure 2.
Molecular mechanisms of N uptake, assimilation, and utilization in rice.
NR assimilates nitrogen in plants via post-translational phosphorylation. NIA1 protein dephosphorylation on rice growth and metabolism under different nitrogen sources reveals that specific phosphorylation site mutations enhanced nitrogen assimilation and tolerance to ammonium nitrogen deficiency. When ammonium nitrate was used as the main nitrogen source, there was weak growth of NIA1 phosphorylation site-directed mutant lines by excessive nitrite accumulation. Conversely, in the case of potassium nitrate, the assimilation rates of nitrate, nitrite, and ammonium salt increased significantly in NIA1 phosphorylation site-directed mutant lines [158]. OsNLP4, one homologous gene of Arabidopsis NLP7, has a crucial role in nitrogen uptake, assimilation, signaling, and utilization in rice. Overexpression of OsNLP4 significantly increased NUE, crop yield, and plant biomass. Under a nitrate-dependent rice cultivation system, OsNLP4 can influence plant growth by affecting NR activity, particularly in OsNLP4 mutant lines controlling the expression patterns of some nitrate signaling genes. Moreover, OsNLP4 showed a shoot reduction of 30%, suggesting enhanced translocation of N from the roots, resulting in enhanced NUE (47%) and grain yield (30%) [159,160]. Partial nitrate nutrition (PNN) enhances NUE in rice by increasing nitric oxide (NO) production mediated by NR. This mechanism adapts rice to fluctuating nitrate supplies, improving nitrogen acquisition and utilization [161].
4.5. Key Genes and Transcription Factors (TFs) Associated with Enhanced NUE in Rice
Japonica and indica are two important cultivated rice subspecies. Japonica varieties are well-suited to high-altitude, temperate regions due to their tolerance to cold and fertilizers. In contrast, indica varieties thrive in subtropical and tropical climates characterized by high temperatures, humidity, and light intensity, but display lesser tolerance to cold and fertilizers. Both varieties exhibit notable genetic and environmental differences in nitrogen absorption and utilization. Compared to japonica rice varieties, indica have higher NUE. Research indicates that indica varieties demonstrate a 14.1% greater NUE than japonica varieties. Additionally, hybrid rice varieties exhibit enhanced NUE over traditional strains. Specifically, hybrid indica varieties show a 22.8% increase in NUE compared to conventional indica, and hybrid japonica varieties achieve a 16.4% improvement over their traditional counterparts [19]. Also, some research findings indicate that indica varieties exhibit 30–40% higher NUE than japonica varieties. The genes NRT1.1B, ARE1, and NR2 display functional differences between these rice varieties [162]. The NRT1.1B gene is crucial for the absorption and movement of nitrogen within plants. This gene variant, identified in indica rice and known as NRT1.1B-Indica, boosts nitrogen uptake and activates genes responsive to nitrate. When NRT1.1B-Indica (Table 2) is introduced into japonica rice varieties, it enhances the grain yield and NUE by 10% with normal nitrogen levels and 30% when the nitrogen supply is halved [163]. Decreasing the expression of OsCBL1 in rice leads to a higher expression of the high-affinity nitrate gene OsNRT2.2. This effect could stem from suppressing the OsCCA1 TF, which results in improved NUE under low nitrogen conditions [164].
The presence of DNR1 in indica varieties underscores the variance in nitrate absorption and assimilation compared to japonica types. The OsNPF3.1 gene, found in the aerial parts of rice, influences plant height, the timing of the heading, and grain weight. It also regulates NUE in rice. Variations in this gene contribute to the differences in NUE observed between indica and japonica rice varieties [162].
The MYB TFs OsMYB305 (shown in Table 2) enhances nitrogen uptake and assimilation, shoot dry weight, and tiller number significantly under low N conditions in rice by encoding a transcriptional activator and its expression [165]. Similarly, the indica allele of MYB61 augments biomass and grain yield under a limited nitrogen supply. This finding is significant because MYB61 is pivotal in governing cellulose synthesis and nitrogen utilization [10].
BT1 and BT2 (bric-a-brac/tramtrack/broad) are a prominent group of TFs that play a crucial role in forming a network for nitrate assimilation by acting on multiple genes. Functional analysis has revealed that these TFs are responsible for regulating NUE in Arabidopsis orthologs, particularly in rice and other cereal crops. The expression of the nitrate transporter genes NRT2.1 and NRT2.4 was higher in the bt1/bt2 double mutant compared to wild-type plants, accompanied by a 65% increase in nitrate uptake under low nitrate conditions. In rice, NUE increased by 20% compared to wild types in low nitrogen conditions [166]. OsDOF18 TFs play a crucial role in ammonium uptake in rice by regulating the expression of ammonium transporter genes and influencing NUE (Table 2) [167].
Table 2.
Regulatory genes and transcription factors with their functions controlling NUE in plants.
Table 2.
Regulatory genes and transcription factors with their functions controlling NUE in plants.
| Category | Gene | Descriptions | Function | Crops | References |
|---|---|---|---|---|---|
| Root System Architecture | OsAMT1;1 | Ammonium transporter 1 member 1 | Increase root length and surface area and encode ammonium transporters, enhancing nitrogen accumulation. | Rice | [114] |
| OsAMT1;2 | Ammonium transporter 1 member 2 | ||||
| RNR10 | Ribonucleoside-diphosphate reductase | Nitrogen-responsive RSA. | [109] | ||
| Nitrate (NO3−) Transporters | AtNRT1.1 | Nitrate transporter 1.1 | Uptake of nitrate NO3− in both low and high nitrogen availability. | Arabidopsis | [134,136,137] |
| NPF6.3 | Nitrate/chlorate transporter | ||||
| AtNRT2.1 | High-affinity nitrate transporter 2.1 | Uptake of nitrate (NO3−) when nitrate is low in soil. | |||
| NRT2.4 | High-affinity nitrate transporter 3.1 | Influx of NO3− into root. | [138,139] | ||
| NRT2.5 | High-affinity nitrate transporter 2.5 | ||||
| OsNRT2.3 | High-affinity nitrate transporter 2.3 | Transportation of nitrate from the root to the stem. | Rice | [140,141] | |
| Ammonia (NH4+) Transporters | AtAMT1;1 | Ammonium transporter 1 member 1 | Decreased uptake of ammonia (NH4+), impedes root and shoot growth. | Arabidopsis | [144,147,148] |
| AtAMT1;2 | Ammonium transporter 1 member 2 | Uptake of ammonia (NH4+), found in root and shoot (leaves). | |||
| AtAMT1;3 | Ammonium transporter 1 member 3 | Uptake of ammonia (NH4+), found in root in low nitrogen availability. | |||
| Nitrogen Assimilation and Utilization | GLN1-1 | Glutamine synthetase cytosolic isozyme 1-1 | Catalyzes synthesis of glutamine from ammonium and glutamate to glutamine synthesis in roots. Ammonium assimilation. | Arabidopsis and Rice | [168,169] |
| NADH-GOGAT 1 | Glutamate synthase 1 [NADH], chloroplast | Glutamate biosynthesis and ammonium ions assimilation in roots. Reutilization of glutamine in developing organs. Development of tillers. | Rice | [170] | |
| GLU2 | Ferredoxin-dependent glutamate synthase 2, chloroplast | Nitrogen assimilation in roots. | Arabidopsis | [171] | |
| Transcription Factors | OsNLP4 | Transcription factor NLP4 | Nitrogen uptake, assimilation, signaling, and utilization in rice. | Rice | [159,160] |
| OsMYB305 | MYB transcription factor | Enhance nitrogen uptake and assimilation, shoot dry weight, and tiller number significantly under low N conditions in rice. | Rice | [165] | |
| MYB61 | Transcription factor MYB61 | Increase biomass and grain yield, under limited nitrogen. | Rice | [10] | |
| OsDOF18 | Dof family transcription factor | Ammonium uptake by regulating the expression of ammonium transporter genes and influencing NUE. | [167] |
5. Genomics and Recent Innovations for Improving NUE in Rice
5.1. Multi-Omics Approaches for Improving NUE in Crops
Integrated transcriptomics and physiological analysis of NUE in rice under drought conditions showed that water scarcity led to a notable reduction in NUE. Similarly, expression of the GAPA and GAPB genes at the vegetative stage and GLO4, GLO5, and OsGDCH reproductive phase facilitates nitrogen utilization and enhancement, making them suitable genetic tools to develop specific crop traits under drought conditions. Moreover, glyoxylate dicarboxylate metabolism and carbon fixation in photosynthetic organisms are major regulatory pathways governing NUE in rice under water-scarce conditions [172]. Integrated multi-omics (Figure 3) in rice regarding low nitrogen (LN) tolerance in the flowering stage revealed three genes (LOC_Os02g40010, LOC_Os11g25260, and LOC_Os11g47300) involved in purine metabolism and genes (LOC_Os02g36880 and LOC_Os08g05830) and metabolites 3-phosphoglyceric acid and sedoheptulose 7-phosphate) associated with glycolysis and the pentose phosphate pathway. Moreover, results demonstrate that purine metabolism is the major pathway governing LN during flowering in rice and five candidate genes can be utilized for molecular breeding strategies for LN tolerance in rice during flowering [173]. Integrative transcriptomics and proteomics revealed that activation of nitrogen transporters (NRT2.1, NRT2.2, NRT2.3, NRT2.4, NAR2.1, AMT1.3, AMT1.2, and AMT3.3) and nitrogen assimilators (like NR2, GS1;1, GS1;2, GS1;3, NADH-GOGAT2, and AS2), along with key metabolic pathways such as phenylpropanoid biosynthesis and glutathione metabolism, optimize nitrogen use efficiency in rice under low nitrogen conditions [174]. Bulk segregant analysis sequencing (BSA-Seq) and RNA-Seq identified candidate genes (BnaC02g14540D, BnaA03g20760D, and BnaA05g01330D) related to NUE in rapeseed (Brassica napus L.) [175].
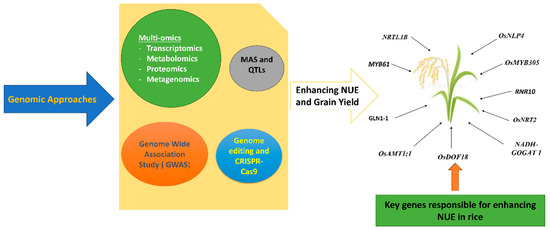
Figure 3.
Genomic approaches employed for enhancing NUE in rice along with key genes associated with increasing N uptake, assimilation, and utilization.
5.2. MAS and QTLs Mediated NUE Enhancements in Crops
Integrating traditional breeding methods with molecular genetics facilitates understanding crop inheritance patterns, leading to improvements in specific crop traits. Advances in rice breeding, such as MAS, QTLs (Figure 3), gene discovery, and CRISPR/Cas9 technologies, enhance NUE and contribute to sustainable crop production. For instance, 190 out of 293 QTLs (64.8%) were found for the nutrient uptake and utilization of N, P, and K in wheat [143,176,177]. A meta-analysis of 506 QTLs from 18 studies has identified 12 meta-QTLs associated with NUE and grain yield per plant under nitrogen-deficit conditions in rice. The mQTLs were found on four rice chromosomes: 1, 3, 4, and 8. These mQTLs are widely distributed throughout the rice germplasm, suggesting the potential to use them as MAS in developing genotypes that have more NUE. This could be an innovative breeding tool for screening and developing rice varieties requiring less nitrogen [178]. QTLs identified 13 loci on various chromosomes linked to root system architecture and nitrogen-deficiency tolerance in rice. Particularly, a locus on chromosome 1 showed a significant correlation with increased root depth and nitrogen-deficiency tolerance, directly impacting grain yield. These identified QTLs could serve as valuable assets in rice breeding programs aiming to enhance RSA traits, ultimately improving rice performance under nitrogen deficiency [179]. Similarly, advancements in genetic tools like QTLs, genes linked to nitrogen metabolism, proteomics, association mapping, microarrays, miRNA, genetic transformation, and transcriptomic approaches can improve crop NUE [180].
SNPs from indica and japonica rice varieties can be crucial for improving NUE in rice. Precisely replacing the japonica NRT-1B allele with the indica allele in a single generation using CRISPR/Cas9 technology illustrates that substituting any gene with superior alleles with desired traits can enhance specific characteristics in crops, such as NUE, leading to increased crop production and productivity [181]. Deploying genome-editing technology can develop cultivars with higher yields and improved nitrogen uptake and utilization efficiency. This technology can be precisely modified in the plant genome with specific genes related to NUE. Using tools like CRISPR-Cas9, researchers can edit genes that regulate nitrogen uptake, assimilation, and utilization in cereals. Moreover, the application of genome editing in cereals has the potential to address the challenges created by N pollution and environmental degradation due to the non-scientific use of N in crop production contributing to sustainable agriculture production [182]. Researchers have been working to identify germplasms that efficiently utilize nutrients, with a specific focus on NUE and crop productivity. High-throughput technologies are being utilized to discover potential germplasms. Exciting progress has been made in the field of plant breeding, through the use of innovative plant breeding techniques (NPBTs) and genome editing technologies, with consideration to be made for the biosafety concerns as well as social, ethical, and environmental issues [183].
5.3. Genome-Wide Association Study (GWAS) for Improving NUE
The GWAS (Figure 3) is a widely accepted and powerful genetic tool for understanding the relationship between phenotype and genotype, which facilitates the identification of potential alleles governing specific crop traits [184,185]. Genetic tools such as GWAS genomic selection (GS) are employed for enhancing NUE in cereal crops by improving traits associated with NUE and altering functional adaptive traits. Additionally, the development of phenotypic approaches coupled with the GWAS and GS showed various genes and agronomic traits and their adoptive mechanisms associated with NUE [186]. The GWAS performed on 419 rice varieties using 208,993 SNPs revealed the gene LOC_Os10g33210, which encodes a peptide transporter, and the gene LOC_Os05g51690, which encodes a CCT domain protein function for NUE in rice. These can serve as new genetic resources for breeding and developing rice cultivars with high NUE [187]. The GWAS conducted in 325 rice-variety natural populations using 111,205 SNPs showed that OsOAT and MOC2 genes play a critical role in N use. Similarly, the gene LOC_Os12g41090, which encodes calcium/calmodulin-dependent protein kinases, shows signals near OsCPK12. OsCPK12 is involved in the signal transduction pathway under low-nitrogen (LN) stress response and enhances NUE in rice [188]. The GWAS of agronomic traits associated with NUE in 244 wheat accessions showed four major genes: TraesCS3A01G101900, TraesCS3A01G102200, TraesCS3A01G104100, and TraesCS3A01G105400 expressed in low nitrogen-tolerant wheat cultivars governing N uptake and assimilation. These can be valuable candidate genes for breeding cultivars for low nitrogen conditions [189]. Similarly, the GWAS performed on 282 barley accessions identified the HvNRT2.7 gene, a high-affinity nitrate transporter linked with shoot and root dry weight, playing a key role in enhancing NUE [190].
5.4. Genetic Modifications and Genome Editing for Improving NUE in Crops
The genome editing technology has great potential to transform agriculture production sustainability through enhanced NUE. By utilizing genome editing technology, increased NUE in crops can serve as a means of resilient food production and ensure the food security of present and future generations. The key components of this technology include target gene selection and identification, diverse genome editing tools, delivery methodologies, and the assessment of crops aimed to improve NUE [191]. In recent years, RNA-guided genome editing technology (CRISPR-Cas9), as displayed in Figure 3 has significantly transformed plant genome editing. Engineering of CRISPR/dead (d) Cas9 (dCas9) cytosine, adenine base editing, and prime editing has had great success in achieving desired traits in crops. Moreover, the dCas9-SunTag system is being employed in plants for the activation of target genes and DNA methylation. This genetic improvement will improve crop nutrient stress tolerance and contribute to food sustainability [192]. Epigenetic gene regulation by non-coding RNA is used as a new genome editing approach for the identification and validation of genes governing NUE in crops. Additionally, haplotyping, allele mining, spatial and temporal expression analysis, gene network, and validation can be employed for the development of high-yielding rice varieties with enhanced NUE under optimum nitrogen [193]. Identification of the abnormal cytokinin response1 repressor 1 (HvARE1) gene in barley (Hordeum vulgare) through the GWAS and Cas9 showed that it is expressed predominantly in leaves and shoots with low expression in roots under low nitrogen conditions, demonstrating increases in plant height, tiller numbers, grain protein content, and grain yield [194]. Genetic modifications of crops that target diversified traits governing nitrogen uptake and utilization showed improved NUE. Meta-analysis data revealed that genetic modifications increase crop production by 16.7% on average, which can be justified by the increase in PFPN (9.47%) and NUpE (16.2%) [195]. Overexpression of the high-affinity NRT gene (NpNRT2.1) in Nicotiana plumbaginifolia notably enhanced nitrate uptake in transgenic plants [196].
5.5. Metagenomics and Recent Advancements for Enhancing NUE in Crops
Metagenomics has transformed the study of soil microbial populations by facilitating whole genome analysis without culturing and capturing a broader range of genetic information than traditional methods. It offers deep insights into associated microbiota, highlighting their importance in understanding plant–soil–microbe interactions, improving nutrient use efficiency and sustainable crop production [197]. Plant microbiomes are an important component of biological climate, determining the genetic makeup of microorganisms within soil and plant systems. They are also associated with different mechanisms in plants, such as plant resistance, nutrient uptake, recycling, mobilization, and crop production and productivity. Metagenomics is a modern and advanced tool that facilitates designing the right microbiome for soil and plant well-being and can be targeted to enhance specific crop traits [198]. Optimizing nitrogen (N) application rates and planting densities is crucial for improving rice yields and quality. An integrated metagenomics conducted to investigate the effects of different levels of N fertilizer and plant densities on soil microbial communities and nitrogen uptake revealed that out of high-nitrogen low-density (HNLD), medium-nitrogen medium-density (MNMD), and low-nitrogen high-density (LNHD), the HNLD treatment demonstrated the highest absorption and utilization efficiency. Key bacterial groups, including Actinobacteria, Acidobacteria, Chloroflexi, and Proteobacteria, predominantly play a vital role in nitrogen uptake and assimilation. Moreover, other factors influencing N uptake and efficiency include root nitrite reductase (NIR) activity, soil enzyme activities (sucrose and catalase), and specific bacterial communities [199]. A metagenomic study revealed that fulvic acid increases nitrogen fixation and retention in paddy soils through the combined microbial N and carbon cycle [200]. Adequate N supply reduced the abundance of pathogens in the phyllosphere, and fungi more vitally regulate phyllosphere microbial functions than bacteria [201]. An imported biogeochemical cycle, dissimilatory nitrate reduction to ammonium (DNRA), has recently been reported to have significant potential in improving nitrogen use efficiency, (NUE) while reducing nitrogen loss and N2O emissions in paddy rice [202]. Cyanobacterial strains are involved in increased amounts of nitrate and ammonium in rice. In silico analysis discovered 12 gene loci and 15 OsAMT isoforms in the rice genome [203]. Metagenomic sequencing of rhizospheric soil in comparison to indica and japonica resulted in dominant bacteria at the phylum level as Actinobacteria, Proteobacteria, Chloroflexi, Acidobacteria, Gemmatimonadetes, and Nitrospirae. Indica rice’s rhizosphere was more enriched with microbial communities involved in nitrogen metabolism, contributing to enhanced NUE. Insights into the correlation between deficiency effects contribute to heightened N uptake and utilization in rice production [204]. The metagenomic study identified root oxidation activity (ROA) in bacterial communities related to N management in rice, possessing the potential to increase NUE and yield [205] (Table 3).
Table 3.
Rice cultivars and genotypes demonstrate enhanced NUE and grain yield under varied soil conditions in major rice-growing countries of Asia.
6. Conclusions
Improving NUE is essential for achieving sustainable agriculture amidst an increasing population, food demands, environmental degradation, global warming, and climate change. This review offers deep insights into the benefits of advanced agronomic practices over conventional nitrogen-use approaches and molecular mechanisms of N uptake and assimilation, demonstrating recent molecular and genomic innovations to optimize nitrogen uptake, assimilation, and utilization. The adoption of fertilizer best management practices (BMPs) in farmers’ fields, the integration of efficient N management strategies such as the 4Rs Nutrient Stewardship, enhanced efficiency nitrogen fertilizers (EENFs), nanotechnology, biochar, conservation agriculture (intercropping and green manuring), and sensor-based technologies have all proven to optimize N inputs and increase NUE and crop yields. These approaches are key in reducing yield gaps while promoting sustainable agriculture practices, improving the livelihood of the farmers. The paradigm of molecular mechanisms, including root system architecture (RSA) modulation, nitrogen transporters, signal transduction, and regulatory pathways such as phenylpropanoid biosynthesis, glutathione metabolism, and GOGAT provides a genetic foundation of NUE in rice. Advances and innovations in genomic technologies, including genome editing and CRISPR-Cas9, marker-assisted selection (MAS), genome-wide association studies (GWAS), multi-omics, and metagenomics have revealed key genes and regulatory networks that can be valuable tools for developing nitrogen-efficient rice varieties. Integrating agronomic and genetic strategies creates a robust framework for enhancing NUE, minimizing fertilizer reliance, mitigating environmental pollution, and ensuring global food security.
7. Future Prospects
Future approaches to improve NUE should prioritize the integration of innovative and precise agricultural technologies, such as sensor-based fertilizer management and application, the use of digital soil maps, mobile applications, and real-time decision-making tools, to optimize nitrogen application and NUE. Scaling up the adoption of nano-fertilizers, including nano-urea, can transform nitrogen management by significantly lowering inputs and costs without sacrificing productivity. Insights into multi-omic technologies, such as transcriptomics, proteomics, and metagenomics, will provide a deeper foundation for the molecular and microbial interactions that regulate nitrogen cycling in soils and crops. Genomic approaches, particularly CRISPR-Cas9, can be further utilized to introduce superior alleles and create rice cultivars with enhanced NUE suitable to multiple agroecological zones and soil conditions.
Rice varieties with enhanced NUE lead to higher grain yield. Integrating agronomic and molecular approaches significantly enhance NUE in rice, reduces nutrient losses, and increases crop productivity, leading to sustainable rice production.
Author Contributions
Conceptualization: U.B.K., N.R.P., N.M., and J.W.; methodology: U.B.K., N.R.P., and N.M.; formal analysis: U.B.K., N.R.P., and N.M.; writing—original draft preparation: U.B.K.; writing—review and editing: U.B.K., J.W., N.R.P., and N.M.; supervision: J.W., N.R.P., and N.M. All authors have read and agreed to the published version of the manuscript.
Funding
The major science and technology project in Yunnan Province (202402AE090026-04, 202102AE090017) and Kunming academic free exploration project (KJHZC-2022YS02).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are contained within the article.
Acknowledgments
We acknowledge Yunnan Agricultural University Rice Research Institute supporting this study.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Bandumula, N. Rice Production in Asia: Key to Global Food Security. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 2018, 88, 1323–1328. [Google Scholar] [CrossRef]
- Muthayya, S.; Hall, J.; Bagriansky, J.; Sugimoto, J.; Gundry, D.; Matthias, D.; Prigge, S.; Hindle, P.; Moench-Pfanner, R.; Maberly, G. Rice Fortification: An Emerging Opportunity to Contribute to the Elimination of Vitamin and Mineral Deficiency Worldwide. Food Nutr. Bull. 2012, 33, 296–307. [Google Scholar] [CrossRef] [PubMed]
- Muthayya, S.; Sugimoto, J.D.; Montgomery, S.; Maberly, G.F. An Overview of Global Rice Production, Supply, Trade, and Consumption. Ann. NY Acad. Sci. 2014, 1324, 7–14. [Google Scholar] [CrossRef]
- Prasad, R.; Shivay, Y.S.; Kumar, D. Current Status, Challenges, and Opportunities in Rice Production. In Rice Production Worldwide; Chauhan, B.S., Jabran, K., Mahajan, G., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 1–32. ISBN 978-3-319-47514-1. [Google Scholar]
- Wu, H.; Xiang, J.; Zhang, Y.; Zhang, Y.; Peng, S.; Chen, H.; Zhu, D. Effects of Post-Anthesis Nitrogen Uptake and Translocation on Photosynthetic Production and Rice Yield. Sci. Rep. 2018, 8, 12891. [Google Scholar] [CrossRef] [PubMed]
- Gaihre, Y.K.; Singh, U.; Islam, S.M.M.; Huda, A.; Islam, M.R.; Satter, M.A.; Sanabria, J.; Islam, M.R.; Shah, A.L. Impacts of Urea Deep Placement on Nitrous Oxide and Nitric Oxide Emissions from Rice Fields in Bangladesh. Geoderma 2015, 259–260, 370–379. [Google Scholar] [CrossRef]
- Snyder, C.S.; Bruulsema, T.W.; Jensen, T.L.; Fixen, P.E. Review of Greenhouse Gas Emissions from Crop Production Systems and Fertilizer Management Effects. Agric. Ecosyst. Environ. 2009, 133, 247–266. [Google Scholar] [CrossRef]
- Kumar, N.; Kumar, V. Production Potential and Nitrogen Fractionation of Sugarcane-Based Cropping System as Influenced by Planting Materials and Nitrogen Nutrition. Sugar Tech 2020, 22, 622–629. [Google Scholar] [CrossRef]
- Omara, P.; Aula, L.; Oyebiyi, F.; Raun, W.R. World Cereal Nitrogen Use Efficiency Trends: Review and Current Knowledge. Agrosystems Geosci. Environ. 2019, 2, 1–8. [Google Scholar] [CrossRef]
- Hu, B.; Wang, W.; Chen, J.; Liu, Y.; Chu, C. Genetic Improvement toward Nitrogen-Use Efficiency in Rice: Lessons and Perspectives. Mol. Plant 2023, 16, 64–74. [Google Scholar] [CrossRef]
- Kumar, N.; Kumar, V.; Kishor, K.; Singh, A.K. Optimizing Nutrient Application and Nitrogen Transformation to Maximize the Growth and Productivity of Bud Chip Transplanted Sugarcane. J. Plant Nutr. 2024, 47, 2583–2596. [Google Scholar] [CrossRef]
- Kunwar, U.B.; Wen, J.; Subedi, R.; Bist, N.S.; Pandit, N.R. Adaptations of Rice Seed Germination to Drought and Hypoxic Conditions: Molecular and Physiological Insights. Seeds 2024, 3, 656–676. [Google Scholar] [CrossRef]
- United Nations Department of Economic and Social Affairs, P.D. World Population Prospects 2024: Summary of Results; UN DESA/POP/2024/TR/NO. 9; United Nations: New York City, NY, USA, 2024. [Google Scholar]
- Yulong, Y.; Qingfeng, M.; Hao, Y.; Qingsong, Z.; Ye, L.; Cui, Z. Climate Change Increases Nitrogen Concentration in Rice with Low Nitrogen Use Efficiency. Earth Future 2021, 9, e2020EF001878. [Google Scholar] [CrossRef]
- Han, M.; Okamoto, M.; Beatty, P.H.; Rothstein, S.J.; Good, A.G. The Genetics of Nitrogen Use Efficiency in Crop Plants. Annu. Rev. Genet. 2015, 49, 269–289. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, Y.; Han, W.; Tang, A.; Shen, J.; Cui, Z.; Vitousek, P.; Erisman, J.W.; Goulding, K.; Christie, P.; et al. Enhanced Nitrogen Deposition over China. Nature 2013, 494, 459–462. [Google Scholar] [CrossRef]
- Sapkota, T.B.; Bijay-Singh; Takele, R. Improving Nitrogen Use Efficiency and Reducing Nitrogen Surplus through Best Fertilizer Nitrogen Management in Cereal Production: The Case of India and China. In Advances in Agronomy; Elsevier: Amsterdam, The Netherlands, 2023; Volume 178, pp. 233–294. ISBN 978-0-443-19260-9. [Google Scholar]
- Kumar, N.; Rana, L.; Singh, A.K.; Pramanick, B.; Gaber, A.; Alsuhaibani, A.M.; Skalicky, M.; Hossain, A. Precise Macronutrient Application Can Improve Cane Yield and Nutrient Uptake in Widely Spaced Plant-Ratoon Cycles in the Indo-Gangetic Plains of India. Front. Sustain. Food Syst. 2023, 7, 1223881. [Google Scholar] [CrossRef]
- Hou, M.; Yu, M.; Li, Z.; Ai, Z.; Chen, J. Molecular Regulatory Networks for Improving Nitrogen Use Efficiency in Rice. Int. J. Mol. Sci. 2021, 22, 9040. [Google Scholar] [CrossRef] [PubMed]
- Salama, E.A.A.; Kambale, R.; Gnanapanditha Mohan, S.V.; Premnath, A.; Fathy Yousef, A.; Moursy, A.R.A.; Abdelsalam, N.R.; Abd El Moneim, D.; Muthurajan, R.; Manikanda Boopathi, N. Empowering Rice Breeding with NextGen Genomics Tools for Rapid Enhancement Nitrogen Use Efficiency. Gene 2024, 927, 148715. [Google Scholar] [CrossRef] [PubMed]
- Chivenge, P.; Sharma, S.; Bunquin, M.A.; Hellin, J. Improving Nitrogen Use Efficiency—A Key for Sustainable Rice Production Systems. Front. Sustain. Food Syst. 2021, 5, 737412. [Google Scholar] [CrossRef]
- De Datta, S.K. Improving Nitrogen Fertilizer Efficiency in Lowland Rice in Tropical Asia. In Nitrogen Economy of Flooded Rice Soils; De Datta, S.K., Patrick, W.H., Eds.; Springer: Dordrecht, The Netherlands, 1986; pp. 171–186. ISBN 978-94-010-8471-0. [Google Scholar]
- Baral, B.R.; Pande, K.R.; Gaihre, Y.K.; Baral, K.R.; Sah, S.K.; Thapa, Y.B.; Singh, U. Increasing Nitrogen Use Efficiency in Rice through Fertilizer Application Method Under Rainfed Drought Conditions in Nepal. Nutr. Cycl. Agroecosyst. 2020, 118, 103–114. [Google Scholar] [CrossRef]
- Dobermann, A.R. Nitrogen Use Efficiency—State of the Art; International Fertilizer Industry Association: Paris, France, 2005; p. 31. [Google Scholar]
- Ohnishi, M.; Horie, T.; Homma, K.; Supapoj, N.; Takano, H.; Yamamoto, S. Nitrogen Management and Cultivar Effects on Rice Yield and Nitrogen Use Efficiency in Northeast Thailand. Field Crops Res. 1999, 64, 109–120. [Google Scholar] [CrossRef]
- Zhang, F.S.; Wang, J.Q.; Zhang, W.F.; Cui, Z.L.; Ma, W.Q.; Chen, X.P.; Jiang, R.F. Nutrient Use Efficiencies of Major Cereal Crops in China and Measures for Improvement. Acta Pedol. Sin. 2008, 45, 915–924. [Google Scholar]
- Huda, A.; Gaihre, Y.; Islam, M.; Singh, U.; Islam, M.; Sanabria, J.; Satter, M.; Afroz, H.; Halder, A.; Jahiruddin, M. Floodwater Ammonium, Nitrogen Use Efficiency and Rice Yields with Fertilizer Deep Placement and Alternate Wetting and Drying Under Triple Rice Cropping Systems. Nutr. Cycl. Agroecosyst. 2016, 104, 53–66. [Google Scholar] [CrossRef]
- Liang, X.Q.; Li, H.; Wang, S.X.; Ye, Y.S.; Ji, Y.J.; Tian, G.M.; van Kessel, C.; Linquist, B.A. Nitrogen Management to Reduce Yield-Scaled Global Warming Potential in Rice. Field Crops Res. 2013, 146, 66–74. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, J.; Yang, J. Improving Nitrogen Use Efficiency of Rice Crop through an Optimized Root System and Agronomic Practices. Crop Environ. 2023, 2, 192–201. [Google Scholar] [CrossRef]
- Balasubramanian, V.; Morales, A.; Cruz, R.; Thiyagarajan, T.; Nagarajan, R.; Babu, M.; Abdulrachman, S.; Hai, L. Adaptation of the Chlorophyll Meter (SPAD) Technology for Real-Time N Management in Rice: A Review. Int. Rice Res. Notes 2000, 25, 4–8. [Google Scholar]
- Sairam, M. Hand-Held Optical Sensors for Optimizing Nitrogen Application and Improving Nutrient Use Efficiency. Int. J. Bioresour. Sci. 2023, 10, 9–18. [Google Scholar] [CrossRef]
- Argento, F.; Anken, T.; Abt, F.; Vogelsanger, E.; Walter, A.; Liebisch, F. Site-Specific Nitrogen Management in Winter Wheat Supported by Low-Altitude Remote Sensing and Soil Data. Precis. Agric. 2021, 22, 364–386. [Google Scholar] [CrossRef]
- Dobermann, A.; Witt, C.; Dawe, D.; Abdulrachman, S.; Gines, H.C.; Nagarajan, R.; Satawathananont, S.; Son, T.T.; Tan, P.S.; Wang, G.H.; et al. Site-Specific Nutrient Management for Intensive Rice Cropping Systems in Asia. Field Crops Res. 2002, 74, 37–66. [Google Scholar] [CrossRef]
- Muschietti-Piana, M.D.P.; Cipriotti, P.A.; Urricariet, S.; Peralta, N.R.; Niborski, M. Using Site-Specific Nitrogen Management in Rainfed Corn to Reduce the Risk of Nitrate Leaching. Agric. Water Manag. 2018, 199, 61–70. [Google Scholar] [CrossRef]
- Pandit, N.R.; Adhikari, S.; Vista, S.P.; Choudhary, D. Nitrogen Management Utilizing 4R Nutrient Stewardship: A Sustainable Strategy for Enhancing NUE, Reducing Maize Yield Gap and Increasing Farm Profitability. Nitrogen 2025, 6, 7. [Google Scholar] [CrossRef]
- Fixen, P.E. A Brief Account of the Genesis of 4R Nutrient Stewardship. Agron. J. 2020, 112, 4511–4518. [Google Scholar] [CrossRef]
- Jat, M.L.; Satyanarayana, T.; Majumdar, K.; Parihar, C.M.; Jat, S.L.; Tetarwal, J.P.; Jat, R.K.; Saharawat, Y.S. Fertiliser Best Management Practices for Maize Systems. Indian J. Fertil. 2013, 9, 80–94. [Google Scholar]
- Pandit, N.R.; Gaihre, Y.K.; Gautam, S.; Maharjan, S.; Vista, S.P.; Choudhary, D. Enhanced-Efficiency Nitrogen Fertilizer Boosts Cauliflower Productivity and Farmers’ Income: Multi-Location and Multi-Year Field Trials across Nepal. Exp. Agric. 2022, 58, e14. [Google Scholar] [CrossRef]
- Ishfaq, M.; Akbar, N.; Zulfiqar, U.; Ali, N.; Jabran, K.; Nawaz, M.; Farooq, M. Influence of Nitrogen Fertilization Pattern on Productivity, Nitrogen Use Efficiencies, and Profitability in Different Rice Production Systems. J. Soil Sci. Plant Nutr. 2021, 21, 145–161. [Google Scholar] [CrossRef]
- Redda, A.; Redae, W.; Hailegebriel, K.; Weldegerima, G.; Tsegay, G.; Yirgalem, T.; Eyasu, A.; Hussien, S. Nitrogen Fertilizer Management Factors Affecting Nitrogen Use Efficiency and Yield of Rice (Oryza sativa L.): A REVIEW. Asian J. Plant Soil. Sci. 2022, 7, 1–22. [Google Scholar] [CrossRef]
- Pandit, N.R.; Choudhary, D.; Maharjan, S.; Dhakal, K.; Vista, S.P.; Gaihre, Y.K. Optimum Rate and Deep Placement of Nitrogen Fertilizer Improves Nitrogen Use Efficiency and Tomato Yield in Nepal. Soil Syst. 2022, 6, 72. [Google Scholar] [CrossRef]
- Thapa, G.; Choudhary, D.; Pandit, N.R.; Dongol, P. Fertilizer Demonstration, Agricultural Performance, and Food Security of Smallholder Farmers: Empirical Evidence from Nepal. World Dev. Sustain. 2025, 6, 100196. [Google Scholar] [CrossRef]
- Bruulsema, T. Managing Nutrients to Mitigate Soil Pollution. Environ. Pollut. 2018, 243, 1602–1605. [Google Scholar] [CrossRef]
- Feng, J.; Li, F.; Deng, A.; Feng, X.; Fang, F.; Zhang, W. Integrated Assessment of the Impact of Enhanced-Efficiency Nitrogen Fertilizer on N2O Emission and Crop Yield. Agric. Ecosyst. Environ. 2016, 231, 218–228. [Google Scholar] [CrossRef]
- Halvorson, A.D.; Snyder, C.S.; Blaylock, A.D.; Del Grosso, S.J. Enhanced-Efficiency Nitrogen Fertilizers: Potential Role in Nitrous Oxide Emission Mitigation. Agron. J. 2014, 106, 715–722. [Google Scholar] [CrossRef]
- Chen, Z.; Wang, Q.; Ma, J.; Zou, P.; Jiang, L. Impact of Controlled-Release Urea on Rice Yield, Nitrogen Use Efficiency and Soil Fertility in a Single Rice Cropping System. Sci. Rep. 2020, 10, 10432. [Google Scholar] [CrossRef] [PubMed]
- Dimkpa, C.O.; Fugice, J.; Singh, U.; Lewis, T.D. Development of Fertilizers for Enhanced Nitrogen Use Efficiency—Trends and Perspectives. Sci. Total Environ. 2020, 731, 139113. [Google Scholar] [CrossRef]
- Linquist, B.A.; Liu, L.; Van Kessel, C.; Van Groenigen, K.J. Enhanced Efficiency Nitrogen Fertilizers for Rice Systems: Meta-Analysis of Yield and Nitrogen Uptake. Field Crops Res. 2013, 154, 246–254. [Google Scholar] [CrossRef]
- Pandit, N.R.; Gaihre, Y.K.; Choudhary, D.; Subedi, R.; Thapa, S.B.; Maharjan, S.; Khadka, D.; Vista, S.P.; Rusinamhodzi, L. Slow but Sure: The Potential of Slow-Release Nitrogen Fertilizers to Increase Crop Productivity and Farm Profit in Nepal. J. Plant Nutr. 2022, 45, 2986–3003. [Google Scholar] [CrossRef]
- Trenkel, M.E. Slow- and Controlled-Release and Stabilized Fertilizers: An Option for Enhancing Nutrient Use Efficiency in Agriculture, 2nd ed.; International Fertilizer Industry Association (IFA): Paris, France, 2010; ISBN 978-2-9523139-7-1. [Google Scholar]
- Zheng, W.; Zhang, M.; Liu, Z.; Zhou, H.; Lu, H.; Zhang, W.; Yang, Y.; Li, C.; Chen, B. Combining Controlled-Release Urea and Normal Urea to Improve the Nitrogen Use Efficiency and Yield under Wheat-Maize Double Cropping System. Field Crops Res. 2016, 197, 52–62. [Google Scholar] [CrossRef]
- Xu, R.; Chen, S.; Xu, C.M.; Liu, Y.H.; Zhang, X.F.; Wang, D.Y.; Chu, G. Polymer-Coated Urea Application Can Increase Both Grain Yield and Nitrogen Use Efficiency in Japonica-Indica Hybrid Rice. J. Agric. Sci. 2023, 161, 51–59. [Google Scholar] [CrossRef]
- Tang, C.; Han, M.; Yang, X.; Shen, T.; Gao, Y.; Wang, Y.; Zhang, S.; Chen, D.; He, D.; Li, Y.C. Gene Expression, Enzyme Activity, Nitrogen Use Efficiency, and Yield of Rice Affected by Controlled-Release Nitrogen. ACS Omega 2023, 8, 23772–23781. [Google Scholar] [CrossRef]
- Agyin-Birikorang, S.; Winings, J.H.; Yin, X.; Singh, U.; Sanabria, J. Field Evaluation of Agronomic Effectiveness of Multi-Nutrient Fertilizer Briquettes for Upland Crop Production. Nutr. Cycl. Agroecosyst. 2018, 110, 395–406. [Google Scholar] [CrossRef]
- Dhakal, K.; Baral, B.R.; Pokhrel, K.R.; Pandit, N.R.; Thapa, S.B.; Gaihre, Y.K.; Vista, S.P. Deep Placement of Briquette Urea Increases Agronomic and Economic Efficiency of Maize in Sandy Loam Soil. Agrivita J. Agric. Sci. 2020, 42, 499–508. [Google Scholar] [CrossRef]
- Eric Derrick, B.; Etienne, I.; Mathusalem, K. Comparative Study of Urea in Prilled and Briquette Forms on Rice Production in Marshlands of Rwanda. J. Fertil. Pestic. 2017, 8, 178. [Google Scholar] [CrossRef]
- Zhu, C.; Xiang, J.; Zhang, Y.; Zhang, Y.; Zhu, D.; Chen, H. Mechanized Transplanting with Side Deep Fertilization Increases Yield and Nitrogen Use Efficiency of Rice in Eastern China. Sci. Rep. 2019, 9, 5653. [Google Scholar] [CrossRef] [PubMed]
- Lyu, Y.; Yang, X.; Pan, H.; Zhang, X.; Cao, H.; Ulgiati, S.; Wu, J.; Zhang, Y.; Wang, G.; Xiao, Y. Impact of Fertilization Schemes with Different Ratios of Urea to Controlled Release Nitrogen Fertilizer on Environmental Sustainability, Nitrogen Use Efficiency and Economic Benefit of Rice Production: A Study Case from Southwest China. J. Clean. Prod. 2021, 293, 126198. [Google Scholar] [CrossRef]
- Lan, C.; Zou, J.; Li, J.; Xu, H.; Lin, W.; Weng, P.; Fang, C.; Zhang, Z.; Chen, H.; Lin, W. Slow-Release Fertilizer Deep Placement Increased Rice Yield and Reduced the Ecological and Environmental Impact in Southeast China: A Life-Cycle Perspective. Field Crops Res. 2024, 306, 109224. [Google Scholar] [CrossRef]
- Iqbal, M.A. Nano-Fertilizers for Sustainable Crop Production Under Changing Climate: A Global Perspective; IntechOpen: London, UK, 2019; ISBN 978-1-78985-317-9. [Google Scholar]
- Rana, L.; Kumar, M.; Rajput, J.; Kumar, N.; Sow, S.; Kumar, S.; Kumar, A.; Singh, S.N.; Jha, C.K.; Singh, A.K.; et al. Nexus between Nanotechnology and Agricultural Production Systems: Challenges and Future Prospects. Discov. Appl. Sci. 2024, 6, 555. [Google Scholar] [CrossRef]
- Kumar, A.; Sheoran, P.; Kumar, N.; Devi, S.; Kumar, A.; Malik, K.; Rani, M.; Bhardwaj, A.K.; Mann, A. Elucidating Morphogenic and Physiological Traits of Rice with Nitrogen Substitution through Nano-Nitrogen under Salt Stress Conditions. BMC Plant Biol. 2024, 24, 908. [Google Scholar] [CrossRef]
- Periakaruppan, R.; Romanovski, V.; Thirumalaisamy, S.K.; Palanimuthu, V.; Sampath, M.P.; Anilkumar, A.; Sivaraj, D.K.; Ahamed, N.A.N.; Murugesan, S.; Chandrasekar, D.; et al. Innovations in Modern Nanotechnology for the Sustainable Production of Agriculture. ChemEngineering 2023, 7, 61. [Google Scholar] [CrossRef]
- Rahale, C.S. Nutrient Release Pattern of Nanofertilizer Formulation. Ph.D. Thesis, Tamilnadu Agricultural University, Coimbatore, Tamil Nadu, India, 2011. [Google Scholar]
- Upadhyay, P.K.; Dey, A.; Singh, V.K.; Dwivedi, B.S.; Singh, T.; Rajanna, G.A.; Babu, S.; Rathore, S.S.; Singh, R.K.; Shekhawat, K.; et al. Conjoint Application of Nano-Urea with Conventional Fertilizers: An Energy Efficient and Environmentally Robust Approach for Sustainable Crop Production. PLoS ONE 2023, 18, e0284009. [Google Scholar] [CrossRef]
- Cowling, S.A.; Field, C.B. Environmental Control of Leaf Area Production: Implications for Vegetation and Land-surface Modeling. Glob. Biogeochem. Cycles 2003, 17, 7-1–7-14. [Google Scholar] [CrossRef]
- Kumar, Y.; Singh, T.; Raliya, R.; Tiwari, K. Nano Fertilizers for Sustainable Crop Production, Higher Nutrient Use Efficiency and Enhanced Profitability. Indian J. Fertil. 2021, 17, 1206–1214. [Google Scholar]
- Rai, M.; Ingle, A.; Pandit, R. Nanotechnology in Agro-Food: From Field to Plate. Food Nutr. J. 2015, 6, 40–47. [Google Scholar]
- Singh, B.V.; Rana, N.S.; Sharma, K.; Verma, A.; Rai, A.K.; Singh, N.K.; Pandey, S.K. Impact of Nano-Fertilizers on Productivity and Profitability of Wheat (Triticum aestivum L.). J. Soil Sci. Plant Nutr. 2024, 25, 69–76. [Google Scholar] [CrossRef]
- Bashir, S.D.; Bhat, T.A.; Jamsheed, B.; Nazir, A.; Jan, B.; Kanth, R.H.; Saxena, A.; Dar, K.A.; Khan, I.M.; Wani, F.J.; et al. Effect of Nano-Urea Based Nitrogen Application on the Growth, Phenology and Yield of Direct Seeded Rice (Oryza sativa L.). Arch. Curr. Res. Int. 2024, 24, 385–395. [Google Scholar] [CrossRef]
- Prem, B. Nano Urea the Philosophy of Future. Res. Gate 2021. [Google Scholar] [CrossRef]
- Kumar, Y.; Tiwari, K.N.; Singh, T.; Sain, N.K.; Laxmi, S.; Verma, R.; Sharma, G.C.; Raliya, R. Nanofertilizers for Enhancing Nutrient Use Efficiency, Crop Productivity and Economic Returns in Winter Season Crops of Rajasthan. Res. Gate 2020. [Google Scholar] [CrossRef]
- Ravikumar, S.; Vellingiri, G.; Sellaperumal, P.; Pandian, K.; Sivasankar, A.; Sangchul, H. Real-Time Nitrogen Monitoring and Management to Augment N Use Efficiency and Ecosystem Sustainability—A Review. J. Hazard. Mater. Adv. 2024, 16, 100466. [Google Scholar] [CrossRef]
- Gautam, S.; Tiwari, U.; Sapkota, B.; Sharma, B.; Parajuli, S.; Pandit, N.R.; Gaihre, Y.K.; Dhakal, K. Field Evaluation of Slow-Release Nitrogen Fertilizers and Real-Time Nitrogen Management Tools to Improve Grain Yield and Nitrogen Use Efficiency of Spring Maize in Nepal. Heliyon 2022, 8, e09566. [Google Scholar] [CrossRef]
- Gorai, T.; Yadav, P.K.; Choudhary, G.L.; Kumar, A. Site-Specific Crop Nutrient Management for Precision Agriculture—A Review. Curr. J. Appl. Sci. Technol. 2021, 37–52. [Google Scholar] [CrossRef]
- Takoutsing, B. Digital Soil Mapping Using Uncertain Soil Observations to Support Agricultural Intensification in West and Central Africa. Ph.D. Thesis, Wageningen University, Wageningen, The Netherlands, 2024. [Google Scholar]
- Mohan, S.S. GPS and Sensor Based Technologies in Variable Rate Fertilizer Application. Int. J. Agric. Environ. Biotechnol. 2021, 14, 21–27. [Google Scholar] [CrossRef]
- Soussi, A.; Zero, E.; Sacile, R.; Trinchero, D.; Fossa, M. Smart Sensors and Smart Data for Precision Agriculture: A Review. Sensors 2024, 24, 2647. [Google Scholar] [CrossRef]
- Lehmann, J.; Gaunt, J.; Rondon, M. Bio-Char Sequestration in Terrestrial Ecosystems—A Review. Mitig. Adapt. Strat. Glob. Change 2006, 11, 403–427. [Google Scholar] [CrossRef]
- Kang, S.-W.; Cheong, Y.H.; Yun, J.-J.; Park, J.-H.; Park, J.-H.; Seo, D.-C.; Cho, J.-S. Effect of Biochar Application on Nitrogen Use Efficiency for Sustainable and Productive Agriculture under Different Field Crops. J. Plant Nutr. 2021, 44, 2849–2862. [Google Scholar] [CrossRef]
- Kimani, S.M.; Bimantara, P.O.; Kautsar, V.; Tawaraya, K.; Cheng, W. Poultry Litter Biochar Application in Combination with Chemical Fertilizer and Azolla Green Manure Improves Rice Grain Yield and Nitrogen Use Efficiency in Paddy Soil. Biochar 2021, 3, 591–602. [Google Scholar] [CrossRef]
- Oladele, S.; Adeyemo, A.; Awodun, M.; Ajayi, A.; Fasina, A. Effects of Biochar and Nitrogen Fertilizer on Soil Physicochemical Properties, Nitrogen Use Efficiency and Upland Rice (Oryza sativa) Yield Grown on an Alfisol in Southwestern Nigeria. Int. J. Recycl. Org. Waste Agric. 2019, 8, 295–308. [Google Scholar] [CrossRef]
- Zheng, Y.; Han, X.; Li, Y.; Liu, S.; Ji, J.; Tong, Y. Effects of Mixed Controlled Release Nitrogen Fertilizer with Rice Straw Biochar on Rice Yield and Nitrogen Balance in Northeast China. Sci. Rep. 2020, 10, 9452. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Zhao, C.; Zhao, X.; Wang, Y.; Lv, X.; Zhu, X.; Song, X. Beneficial Effects of Biochar Application with Nitrogen Fertilizer on Soil Nitrogen Retention, Absorption and Utilization in Maize Production. Agronomy 2022, 13, 113. [Google Scholar] [CrossRef]
- Selvarajh, G.; Ch’ng, H.Y. Enhancing Soil Nitrogen Availability and Rice Growth by Using Urea Fertilizer Amended with Rice Straw Biochar. Agronomy 2021, 11, 1352. [Google Scholar] [CrossRef]
- Pandit, N.R.; Sipkhan, P.; Sharma, S.S.; Dawadi, D.; Vista, S.P.; Raut, P. Cattle-Urine-Enriched Biochar Enhances Soil Fertility, Nutrient Uptake, and Yield of Maize in a Low-Productive Soil. Nitrogen 2024, 5, 16–27. [Google Scholar] [CrossRef]
- Schmidt, H.; Pandit, B.; Martinsen, V.; Cornelissen, G.; Conte, P.; Kammann, C. Fourfold Increase in Pumpkin Yield in Response to Low-Dosage Root Zone Application of Urine-Enhanced Biochar to a Fertile Tropical Soil. Agriculture 2015, 5, 723–741. [Google Scholar] [CrossRef]
- Selvarajh, G.; Ch’ng, H.Y.; Md Zain, N.; Seong Wei, L.; Liew, J.Y.; Mohammad Azmin, S.N.H.; Naher, L.; Abdullah, P.S.; Ahmed, O.H.; Jalloh, M.B.; et al. Enriched Rice Husk Biochar Superior to Commercial Biochar in Ameliorating Ammonia Loss from Urea Fertilizer and Improving Plant Uptake. Heliyon 2024, 10, e32080. [Google Scholar] [CrossRef]
- Utomo, W.; Islami, T.; Wisnubroto, E.; Soelistyari, H.T. Biochar as a Carrier for Nitrogen Plant Nutrition: 3. Effect of Enriched Biochar on Rice (Oryza sativa L.) Yield and Soil Qualities. Int. J. Appl. Eng. Res. 2017, 12, 10426–10432. [Google Scholar]
- Hagemann, N.; Joseph, S.; Conte, P.; Albu, M.; Obst, M.; Borch, T.; Orsetti, S.; Subdiaga, E.; Behrens, S.; Kappler, A. Composting-Derived Organic Coating on Biochar Enhances Its Affinity to Nitrate. In Proceedings of the EGU General Assembly Conference Abstracts, Vienna, Austria, 23–28 April 2017; p. 10775. [Google Scholar]
- Patel, M.R.; Panwar, N.L. Evaluating the Agronomic and Economic Viability of Biochar in Sustainable Crop Production. Biomass Bioenergy 2024, 188, 107328. [Google Scholar] [CrossRef]
- Collino, D.J.; Salvagiotti, F.; Perticari, A.; Piccinetti, C.; Ovando, G.; Urquiaga, S.; Racca, R.W. Biological Nitrogen Fixation in Soybean in Argentina: Relationships with Crop, Soil, and Meteorological Factors. Plant Soil 2015, 392, 239–252. [Google Scholar] [CrossRef]
- Zhang, Z.; Masuda, Y.; Xu, Z.; Shiratori, Y.; Ohba, H.; Senoo, K. Active Nitrogen Fixation by Iron-Reducing Bacteria in Rice Paddy Soil and Its Further Enhancement by Iron Application. Appl. Sci. 2023, 13, 8156. [Google Scholar] [CrossRef]
- Song, X.; Zhang, J.; Li, D.; Peng, C. Nitrogen-Fixing Cyanobacteria Have the Potential to Improve Nitrogen Use Efficiency through the Reduction of Ammonia Volatilization in Red Soil Paddy Fields. Soil Tillage Res. 2022, 217, 105274. [Google Scholar] [CrossRef]
- Choudhury, A.T.M.A.; Kennedy, I.R. Prospects and Potentials for Systems of Biological Nitrogen Fixation in Sustainable Rice Production. Biol. Fertil. Soils 2004, 39, 219–227. [Google Scholar] [CrossRef]
- Das, K.; Biswakarma, N.; Zhiipao, R.; Kumar, A.; Ghasal, P.C.; Pooniya, V. Significance and Management of Green Manures. In Soil Health; Giri, B., Varma, A., Eds.; Soil Biology; Springer International Publishing: Cham, Switzerland, 2020; Volume 59, pp. 197–217. ISBN 978-3-030-44363-4. [Google Scholar]
- Meelu, O.P.; Morris, R.A. Green Manure Management in Rice Based Cropping Systems. In Sustainable Agriculture: Green Manure in Rice Farming; International Rice Research Institute: Manila, Philippines, 1988; pp. 209–222. [Google Scholar]
- Qaswar, M.; Huang, J.; Ahmed, W.; Liu, S.; Li, D.; Zhang, L.; Liu, L.; Xu, Y.; Han, T.; Du, J.; et al. Substitution of Inorganic Nitrogen Fertilizer with Green Manure (GM) Increased Yield Stability by Improving C Input and Nitrogen Recovery Efficiency in Rice Based Cropping System. Agronomy 2019, 9, 609. [Google Scholar] [CrossRef]
- Gebru, H. A Review on the Comparative Advantages of Intercropping to Mono-Cropping System. J. Biol. Agric. Healthc. 2015, 5, 1–13. [Google Scholar]
- Rusinamhodzi, L.; Corbeels, M.; Nyamangara, J.; Giller, K.E. Maize–Grain Legume Intercropping Is an Attractive Option for Ecological Intensification That Reduces Climatic Risk for Smallholder Farmers in Central Mozambique. Field Crops Res. 2012, 136, 12–22. [Google Scholar] [CrossRef]
- Hei, Z.; Xiang, H.; Zhang, J.; Liang, K.; Zhong, J.; Li, M.; Lu, Y. Rice Intercropping with Water Mimosa (Neptunia oleracea Lour.) Can Facilitate Soil N Utilization and Alleviate Apparent N Loss. Agric. Ecosyst. Environ. 2021, 313, 107378. [Google Scholar] [CrossRef]
- Peoples, M.B.; Herridge, D.F.; Ladha, J.K. Biological Nitrogen Fixation: An Efficient Source of Nitrogen for Sustainable Agricultural Production? Plant Soil 1995, 174, 3–28. [Google Scholar] [CrossRef]
- Kamuruzzaman, M.; Rees, R.M.; Islam, M.T.; Drewer, J.; Sutton, M.; Bhatia, A.; Bealey, W.J.; Hasan, M.M. Improving Nitrogen Fertilizer Management for Yield and N Use Efficiency in Wetland Rice Cultivation in Bangladesh. Agronomy 2024, 14, 2758. [Google Scholar] [CrossRef]
- Liu, C.; Sun, Y.; Wu, G.; Wang, X.; Yuan, M.; Wang, J.; He, W.; Chen, F.; LeCocq, K.; Wang, L.; et al. Amendment with Controlled Release Urea Increases Leaf Morpho-physiological Traits, Grain Yield and NUE in a Double-cropping Rice System in Southern China. J. Sci. Food Agric. 2023, 103, 1692–1703. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Guo, X.; Zhou, D.; Zhang, Q.; Ren, X.; Wang, R.; Wang, X.; Chen, X.; Li, J. Quantify the Effect of Manure Fertilizer Addition and Optimal Nitrogen Input on Rainfed Wheat Yield and Nitrogen Requirement Using Nitrogen Nutrition Index. Agric. Ecosyst. Environ. 2023, 345, 108319. [Google Scholar] [CrossRef]
- Asif, M.A.A.; Mahjabin, F.; Singha, S.K.; Rahman Jahangir, M.M.; Hoque, S.M. Application of Nano-Urea in Conventional Flood-Irrigated Boro Rice in Bangladesh and Nitrogen Losses Investigation. Heliyon 2024, 10, e37150. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.; Tripathi, S.C.; Yadav, D.B.; Samota, S.R.; Venkatesh, K.; Sareen, S.; Singh, G. Boosting Wheat Yield, Profitability and NUE with Prilled and Nano Urea in Conservation Tillage. Sci. Rep. 2023, 13, 18073. [Google Scholar] [CrossRef]
- Adams, A.; Gore, J.; Musser, F.; Cook, D.; Catchot, A.; Walker, T.; Awuni, G.A. Impact of Water Management on Efficacy of Insecticide Seed Treatments Against Rice Water Weevil (Coleoptera: Curculionidae) in Mississippi Rice. J. Econ. Entomol. 2015, 108, 1079–1085. [Google Scholar] [CrossRef][Green Version]
- Huang, Y.; Ji, Z.; Tao, Y.; Wei, S.; Jiao, W.; Fang, Y.; Jian, P.; Shen, C.; Qin, Y.; Zhang, S.; et al. Improving Rice Nitrogen-Use Efficiency by Modulating a Novel Monouniquitination Machinery for Optimal Root Plasticity Response to Nitrogen. Nat. Plants 2023, 9, 1902–1914. [Google Scholar] [CrossRef]
- Mandal, V.K.; Sharma, N.; Raghuram, N. Molecular Targets for Improvement of Crop Nitrogen Use Efficiency: Current and Emerging Options. In Engineering Nitrogen Utilization in Crop Plants; Shrawat, A., Zayed, A., Lightfoot, D.A., Eds.; Springer International Publishing: Cham, Switzerland, 2018; pp. 77–93. ISBN 978-3-319-92957-6. [Google Scholar]
- Yan, J.; Wu, Q.; Qi, D.; Zhu, J. Rice Yield, Water Productivity, and Nitrogen Use Efficiency Responses to Nitrogen Management Strategies under Supplementary Irrigation for Rain-Fed Rice Cultivation. Agric. Water Manag. 2022, 263, 107486. [Google Scholar] [CrossRef]
- Nieder, R.; Benbi, D.K.; Scherer, H.W. Fixation and Defixation of Ammonium in Soils: A Review. Biol. Fertil. Soils 2011, 47, 1–14. [Google Scholar] [CrossRef]
- Foulkes, M.J.; Hawkesford, M.J.; Barraclough, P.B.; Holdsworth, M.J.; Kerr, S.; Kightley, S.; Shewry, P.R. Identifying Traits to Improve the Nitrogen Economy of Wheat: Recent Advances and Future Prospects. Field Crops Res. 2009, 114, 329–342. [Google Scholar] [CrossRef]
- Liu, L.; Cui, K.; Qi, X.; Wu, Y.; Huang, J.; Peng, S. Varietal Responses of Root Characteristics to Low Nitrogen Application Explain the Differing Nitrogen Uptake and Grain Yield in Two Rice Varieties. Front. Plant Sci. 2023, 14, 1244281. [Google Scholar] [CrossRef] [PubMed]
- Anis, G.B.; Zhang, Y.; Islam, A.; Zhang, Y.; Cao, Y.; Wu, W.; Cao, L.; Cheng, S. RDWN6XB, a Major Quantitative Trait Locus Positively Enhances Root System Architecture under Nitrogen Deficiency in Rice. BMC Plant Biol. 2019, 19, 12. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Liang, Z.; Chen, S.; Sun, H.; Fan, X.; Wang, C.; Xu, G.; Zhang, Y. A Transcription Factor, OsMADS57, Regulates Long-Distance Nitrate Transport and Root Elongation. Plant Physiol. 2019, 180, 882–895. [Google Scholar] [CrossRef]
- Li, G.; Zhang, Y.; Xu, J.; Zhu, C.; Hu, Q.; Xu, K. Nitrogen Uptake and Carbon-Nitrogen Synergistic Translocation Improve Yield and Nitrogen Use Efficiency in the Dep1 Rice Line. J. Integr. Agric. 2025. [Google Scholar] [CrossRef]
- Gao, J.; Ge, S.; Wang, H.; Fang, Y.; Sun, L.; He, T.; Cheng, X.; Wang, D.; Zhou, X.; Cai, H.; et al. Biochar-Extracted Liquor Stimulates Nitrogen Related Gene Expression on Improving Nitrogen Utilization in Rice Seedling. Front. Plant Sci. 2023, 14, 1131937. [Google Scholar] [CrossRef] [PubMed]
- Al Aasmi, A.; Li, J.; Hamoud, Y.A.; Lan, Y.; Alordzinu, K.E.; Appiah, S.A.; Shaghaleh, H.; Sheteiwy, M.; Wang, H.; Qiao, S.; et al. Impacts of Slow-Release Nitrogen Fertilizer Rates on the Morpho-Physiological Traits, Yield, and Nitrogen Use Efficiency of Rice under Different Water Regimes. Agriculture 2022, 12, 86. [Google Scholar] [CrossRef]
- Ji, Y.; Huang, W.; Wu, B.; Fang, Z.; Wang, X. The Amino Acid Transporter AAP1 Mediates Growth and Grain Yield by Regulating Neutral Amino Acid Uptake and Reallocation in Oryza sativa. J. Exp. Bot. 2020, 71, 4763–4777. [Google Scholar] [CrossRef]
- Zhang, S.; Zhu, L.; Shen, C.; Ji, Z.; Zhang, H.; Zhang, T.; Li, Y.; Yu, J.; Yang, N.; He, Y.; et al. Natural Allelic Variation in a Modulator of Auxin Homeostasis Improves Grain Yield and Nitrogen Use Efficiency in Rice. Plant Cell 2021, 33, 566–580. [Google Scholar] [CrossRef]
- Jiang, N.; Zou, T.; Huang, H.; Li, C.; Xia, Y.; Yang, L. Auxin Synthesis Promotes N Metabolism and Optimizes Root Structure Enhancing N Acquirement in Maize (Zea mays L.). Planta 2024, 259, 46. [Google Scholar] [CrossRef]
- Tang, C.; Zhang, Y.; Liu, X.; Zhang, B.; Si, J.; Xia, H.; Fan, S.; Kong, L. Nitrate Starvation Induces Lateral Root Organogenesis in Triticum aestivum via Auxin Signaling. Int. J. Mol. Sci. 2024, 25, 9566. [Google Scholar] [CrossRef]
- Yousuf, P.Y.; Shabir, P.A.; Hakeem, K.R. Advances in Plant Nitrogen Metabolism, 1st ed.; CRC Press: New York City, NY, USA, 2022; ISBN 978-1-00-324836-1. [Google Scholar]
- Li, Z.; Chen, H.; Guan, Q.; Li, L.; Xuan, Y.H. Gibberellic Acid Signaling Promotes Resistance to Saline-Alkaline Stress by Increasing the Uptake of Ammonium in Rice. Plant Physiol. Biochem. 2024, 207, 108424. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yao, Q.; Zhang, Y.; Zhang, Y.; Xing, J.; Yang, B.; Mi, G.; Li, Z.; Zhang, M. The Role of Gibberellins in Regulation of Nitrogen Uptake and Physiological Traits in Maize Responding to Nitrogen Availability. Int. J. Mol. Sci. 2020, 21, 1824. [Google Scholar] [CrossRef]
- Kiba, T.; Kudo, T.; Kojima, M.; Sakakibara, H. Hormonal Control of Nitrogen Acquisition: Roles of Auxin, Abscisic Acid, and Cytokinin. J. Exp. Bot. 2011, 62, 1399–1409. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Jia, L.; Duan, X.; Lv, Y.; Ye, C.; Ding, C.; Zhang, Y.; Qi, W.; Motte, H.; Beeckman, T.; et al. A Nitrogen-responsive Cytokinin Oxidase/Dehydrogenase Regulates Root Response to High Ammonium in Rice. New Phytol. 2024, 244, 1391–1407. [Google Scholar] [CrossRef]
- Gu, J.; Li, Z.; Mao, Y.; Struik, P.C.; Zhang, H.; Liu, L.; Wang, Z.; Yang, J. Roles of Nitrogen and Cytokinin Signals in Root and Shoot Communications in Maximizing of Plant Productivity and Their Agronomic Applications. Plant Sci. 2018, 274, 320–331. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Wang, G.; Gao, Y.; Lu, G.; Habben, J.E.; Mao, G.; Chen, G.; Wang, J.; Yang, F.; Zhao, X.; et al. A Cytokinin-Activation Enzyme-like Gene Improves Grain Yield under Various Field Conditions in Rice. Plant Mol. Biol. 2020, 102, 373–388. [Google Scholar] [CrossRef]
- Xing, J.; Wang, Y.; Yao, Q.; Zhang, Y.; Zhang, M.; Li, Z. Brassinosteroids Modulate Nitrogen Physiological Response and Promote Nitrogen Uptake in Maize (Zea mays L.). Crop J. 2022, 10, 166–176. [Google Scholar] [CrossRef]
- Yang, W.; Wan, G.-F.; Zhou, J.-Q.; Song, G.-C.; Zhao, J.; Huang, F.-L.; Meng, S. The Effects of Brassinosteroids on Nitrogen Utilization in Rice. Agronomy 2024, 14, 604. [Google Scholar] [CrossRef]
- Hang, J.; Wu, B.; Qiu, D.; Yang, G.; Fang, Z.; Zhang, M. OsNPF3.1, a Nitrate, Abscisic Acid and Gibberellin Transporter Gene, Is Essential for Rice Tillering and Nitrogen Utilization Efficiency. J. Integr. Agric. 2024, 23, 1087–1104. [Google Scholar] [CrossRef]
- Aluko, O.O.; Kant, S.; Adedire, O.M.; Li, C.; Yuan, G.; Liu, H.; Wang, Q. Unlocking the Potentials of Nitrate Transporters at Improving Plant Nitrogen Use Efficiency. Front. Plant Sci. 2023, 14, 1074839. [Google Scholar] [CrossRef]
- Fan, X.; Naz, M.; Fan, X.; Xuan, W.; Miller, A.J.; Xu, G. Plant Nitrate Transporters: From Gene Function to Application. J. Exp. Bot. 2017, 68, 2463–2475. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.-H.; Tsay, Y.-F. Switching between the Two Action Modes of the Dual-Affinity Nitrate Transporter CHL1 by Phosphorylation. EMBO J. 2003, 22, 1005–1013. [Google Scholar] [CrossRef]
- Miller, A.J.; Fan, X.; Orsel, M.; Smith, S.J.; Wells, D.M. Nitrate Transport and Signalling. J. Exp. Bot. 2007, 58, 2297–2306. [Google Scholar] [CrossRef]
- O’Brien, J.A.; Vega, A.; Bouguyon, E.; Krouk, G.; Gojon, A.; Coruzzi, G.; Gutiérrez, R.A. Nitrate Transport, Sensing, and Responses in Plants. Mol. Plant 2016, 9, 837–856. [Google Scholar] [CrossRef]
- Von Wittgenstein, N.J.; Le, C.H.; Hawkins, B.J.; Ehlting, J. Evolutionary Classification of Ammonium, Nitrate, and Peptide Transporters in Land Plants. BMC Evol. Biol. 2014, 14, 11. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.; Fan, X.; Li, Q.; Feng, H.; Miller, A.J.; Shen, Q.; Xu, G. Knockdown of a Rice Stelar Nitrate Transporter Alters Long-Distance Translocation But Not Root Influx. Plant Physiol. 2012, 160, 2052–2063. [Google Scholar] [CrossRef]
- Xu, G.; Fan, X.; Miller, A.J. Plant Nitrogen Assimilation and Use Efficiency. Annu. Rev. Plant Biol. 2012, 63, 153–182. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhang, Q.; Gao, N.; Liu, M.; Zhang, C.; Luo, J.; Sun, Y.; Feng, Y. Nitrate Transporters and Mechanisms of Nitrate Signal Transduction in Arabidopsis and Rice. Physiol. Plant. 2024, 176, e14486. [Google Scholar] [CrossRef]
- Sandhu, N.; Sethi, M.; Kumar, A.; Dang, D.; Singh, J.; Chhuneja, P. Biochemical and Genetic Approaches Improving Nitrogen Use Efficiency in Cereal Crops: A Review. Front. Plant Sci. 2021, 12, 657629. [Google Scholar] [CrossRef]
- Ye, J.Y.; Tian, W.H.; Jin, C.W. Nitrogen in Plants: From Nutrition to the Modulation of Abiotic Stress Adaptation. Stress Biol. 2022, 2, 4. [Google Scholar] [CrossRef]
- Yuan, L.; Loqué, D.; Kojima, S.; Rauch, S.; Ishiyama, K.; Inoue, E.; Takahashi, H.; Von Wirén, N. The Organization of High-Affinity Ammonium Uptake in Arabidopsis Roots Depends on the Spatial Arrangement and Biochemical Properties of AMT1-Type Transporters. Plant Cell 2007, 19, 2636–2652. [Google Scholar] [CrossRef]
- Gazzarrini, S.; Lejay, L.; Gojon, A.; Ninnemann, O.; Frommer, W.B.; Von Wirén, N. Three Functional Transporters for Constitutive, Diurnally Regulated, and Starvation-Induced Uptake of Ammonium into Arabidopsis Roots. Plant Cell 1999, 11, 937–947. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Hu, B.; Chu, C. Nitrogen Use Efficiency in Crops: Lessons from Arabidopsis and Rice. J. Exp. Bot. 2017, 68, 2477–2488. [Google Scholar] [CrossRef]
- Li, C.; Tang, Z.; Wei, J.; Qu, H.; Xie, Y.; Xu, G. The OsAMT1.1 Gene Functions in Ammonium Uptake and Ammonium–Potassium Homeostasis over Low and High Ammonium Concentration Ranges. J. Genet. Genom. 2016, 43, 639–649. [Google Scholar] [CrossRef]
- Sonoda, Y.; Ikeda, A.; Saiki, S.; Yamaya, T.; Yamaguchi, J. Feedback Regulation of the Ammonium Transporter Gene Family AMT1 by Glutamine in Rice. Plant Cell Physiol. 2003, 44, 1396–1402. [Google Scholar] [CrossRef] [PubMed]
- Thomsen, H.C.; Eriksson, D.; Møller, I.S.; Schjoerring, J.K. Cytosolic Glutamine Synthetase: A Target for Improvement of Crop Nitrogen Use Efficiency? Trends Plant Sci. 2014, 19, 656–663. [Google Scholar] [CrossRef] [PubMed]
- Schauser, L.; Wieloch, W.; Stougaard, J. Evolution of NIN-Like Proteins in Arabidopsis, Rice, and Lotus japonicus. J. Mol. Evol. 2005, 60, 229–237. [Google Scholar] [CrossRef]
- Boisson, M.; Mondon, K.; Torney, V.; Nicot, N.; Laine, A.-L.; Bahrman, N.; Gouy, A.; Daniel-Vedele, F.; Hirel, B.; Sourdille, P.; et al. Partial Sequences of Nitrogen Metabolism Genes in Hexaploid Wheat. Theor. Appl. Genet. 2005, 110, 932–940. [Google Scholar] [CrossRef]
- Campbell, W.H. Molecular Control of Nitrate Reductase and Other Enzymes Involved in Nitrate Assimilation. In Photosynthetic Nitrogen Assimilation and Associated Carbon and Respiratory Metabolism; Foyer, C.H., Noctor, G., Eds.; Advances in Photosynthesis and Respiration; Springer: Dordrecht, The Netherlands, 2002; Volume 12, pp. 35–48. ISBN 978-0-7923-6336-1. [Google Scholar]
- Sétif, P.; Hirasawa, M.; Cassan, N.; Lagoutte, B.; Tripathy, J.N.; Knaff, D.B. New Insights into the Catalytic Cycle of Plant Nitrite Reductase. Electron Transfer Kinetics and Charge Storage. Biochemistry 2009, 48, 2828–2838. [Google Scholar] [CrossRef]
- Fortunato, S.; Nigro, D.; Lasorella, C.; Marcotuli, I.; Gadaleta, A.; De Pinto, M.C. The Role of Glutamine Synthetase (GS) and Glutamate Synthase (GOGAT) in the Improvement of Nitrogen Use Efficiency in Cereals. Biomolecules 2023, 13, 1771. [Google Scholar] [CrossRef]
- Brauer, E.K.; Rochon, A.; Bi, Y.; Bozzo, G.G.; Rothstein, S.J.; Shelp, B.J. Reappraisal of Nitrogen Use Efficiency in Rice Overexpressing Glutamine synthetase1. Physiol. Plant. 2011, 141, 361–372. [Google Scholar] [CrossRef] [PubMed]
- Chichkova, S.; Arellano, J.; Vance, C.P.; Hernández, G. Transgenic Tobacco Plants That Overexpress Alfalfa NADH-Glutamate Synthase Have Higher Carbon and Nitrogen Content. J. Exp. Bot. 2001, 52, 2079–2087. [Google Scholar] [CrossRef] [PubMed]
- Han, R.; Xu, Z.; Li, C.; Rasheed, A.; Pan, X.; Shi, Q.; Wu, Z. The Removal of Nitrate Reductase Phosphorylation Enhances Tolerance to Ammonium Nitrogen Deficiency in Rice. J. Integr. Agric. 2022, 21, 631–643. [Google Scholar] [CrossRef]
- Wang, M.; Hasegawa, T.; Beier, M.; Hayashi, M.; Ohmori, Y.; Yano, K.; Teramoto, S.; Kamiya, T.; Fujiwara, T. Growth and Nitrate Reductase Activity Are Impaired in Rice Osnlp4 Mutants Supplied with Nitrate. Plant Cell Physiol. 2021, 62, 1156–1167. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Zhang, Z.; Xia, J.; Alfatih, A.; Song, Y.; Huang, Y.; Wan, G.; Sun, L.; Tang, H.; Liu, Y.; et al. Rice NIN-LIKE PROTEIN 4 Plays a Pivotal Role in Nitrogen Use Efficiency. Plant Biotechnol. J. 2021, 19, 448–461. [Google Scholar] [CrossRef]
- Sun, H.; Li, J.; Song, W.; Tao, J.; Huang, S.; Chen, S.; Hou, M.; Xu, G.; Zhang, Y. Nitric Oxide Generated by Nitrate Reductase Increases Nitrogen Uptake Capacity by Inducing Lateral Root Formation and Inorganic Nitrogen Uptake Under Partial Nitrate Nutrition in Rice. J. Exp. Bot. 2015, 66, 2449–2459. [Google Scholar] [CrossRef]
- Yang, X.; Nong, B.; Chen, C.; Wang, J.; Xia, X.; Zhang, Z.; Wei, Y.; Zeng, Y.; Feng, R.; Wu, Y.; et al. OsNPF3.1, a Member of the NRT1/PTR Family, Increases Nitrogen Use Efficiency and Biomass Production in Rice. Crop J. 2023, 11, 108–118. [Google Scholar] [CrossRef]
- Zhang, Z.; Chu, C. Nitrogen-Use Divergence Between Indica and Japonica Rice: Variation at Nitrate Assimilation. Mol. Plant 2020, 13, 6–7. [Google Scholar] [CrossRef]
- Hu, Z.; Guo, Y.; Ying, S.; Tang, Y.; Niu, J.; Wang, T.; Huang, R.; Xie, H.; Wang, W.; Peng, X. OsCBL1 Modulates Rice Nitrogen Use Efficiency via Negative Regulation of OsNRT2.2 by OsCCA1. BMC Plant Biol. 2023, 23, 502. [Google Scholar] [CrossRef]
- Wang, D.; Xu, T.; Yin, Z.; Wu, W.; Geng, H.; Li, L.; Yang, M.; Cai, H.; Lian, X. Overexpression of OsMYB305 in Rice Enhances the Nitrogen Uptake Under Low-Nitrogen Condition. Front. Plant Sci. 2020, 11, 369. [Google Scholar] [CrossRef]
- Araus, V.; Vidal, E.A.; Puelma, T.; Alamos, S.; Mieulet, D.; Guiderdoni, E.; Gutiérrez, R.A. Members of BTB Gene Family Regulate Negatively Nitrate Uptake and Nitrogen Use Efficiency in Arabidopsis thaliana and Oryza sativa. Plant Physiol. 2016, 171, 1523–1532. [Google Scholar] [CrossRef]
- Wu, Y.; Yang, W.; Wei, J.; Yoon, H.; An, G. Transcription Factor OsDOF18 Controls Ammonium Uptake by Inducing Ammonium Transporters in Rice Roots. Mol. Cells 2017, 40, 178–185. [Google Scholar] [CrossRef] [PubMed]
- Ishiyama, K.; Inoue, E.; Watanabe-Takahashi, A.; Obara, M.; Yamaya, T.; Takahashi, H. Kinetic Properties and Ammonium-Dependent Regulation of Cytosolic Isoenzymes of Glutamine Synthetase in Arabidopsis. J. Biol. Chem. 2004, 279, 16598–16605. [Google Scholar] [CrossRef] [PubMed]
- Kusano, M.; Fukushima, A.; Tabuchi-Kobayashi, M.; Funayama, K.; Kojima, S.; Maruyama, K.; Yamamoto, Y.Y.; Nishizawa, T.; Kobayashi, M.; Wakazaki, M.; et al. Cytosolic GLUTAMINE SYNTHETASE1;1 Modulates Metabolism and Chloroplast Development in Roots. Plant Physiol. 2020, 182, 1894–1909. [Google Scholar] [CrossRef] [PubMed]
- Tamura, W.; Hidaka, Y.; Tabuchi, M.; Kojima, S.; Hayakawa, T.; Sato, T.; Obara, M.; Kojima, M.; Sakakibara, H.; Yamaya, T. Reverse Genetics Approach to Characterize a Function of NADH-Glutamate Synthase1 in Rice Plants. Amino Acids 2010, 39, 1003–1012. [Google Scholar] [CrossRef]
- Potel, F.; Valadier, M.; Ferrario-Méry, S.; Grandjean, O.; Morin, H.; Gaufichon, L.; Boutet-Mercey, S.; Lothier, J.; Rothstein, S.J.; Hirose, N.; et al. Assimilation of Excess Ammonium into Amino Acids and Nitrogen Translocation in Arabidopsis thaliana—Roles of Glutamate Synthases and Carbamoylphosphate Synthetase in Leaves. FEBS J. 2009, 276, 4061–4076. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, Y.; Qiao, H.; Zheng, Y.; Hou, X.; Shi, L. An Integrated Transcriptome and Physiological Analysis of Nitrogen Use Efficiency in Rice (Oryza sativa L. Ssp. Indica) under Drought Stress. Front. Genet. 2024, 15, 1483113. [Google Scholar] [CrossRef]
- Wang, Y.; Jiang, J.; Qian, Y.; Miao, S.; Wang, W.; Xu, J.; Fu, B.; Zhang, F.; Zhao, X. Multi-Omics Analysis Reveals the Regulatory and Metabolic Mechanisms Underlying Low-Nitrogen Tolerance at the Flowering Stage in Rice. Agronomy 2023, 13, 578. [Google Scholar] [CrossRef]
- Liang, T.; Yuan, Z.; Fu, L.; Zhu, M.; Luo, X.; Xu, W.; Yuan, H.; Zhu, R.; Hu, Z.; Wu, X. Integrative Transcriptomic and Proteomic Analysis Reveals an Alternative Molecular Network of Glutamine Synthetase 2 Corresponding to Nitrogen Deficiency in Rice (Oryza sativa L.). Int. J. Mol. Sci. 2021, 22, 7674. [Google Scholar] [CrossRef]
- Nan, Y.; Xie, Y.; He, H.; Wu, H.; Gao, L.; Atif, A.; Zhang, Y.; Tian, H.; Hui, J.; Gao, Y. Integrated BSA-Seq and RNA-Seq Analysis to Identify Candidate Genes Associated with Nitrogen Utilization Efficiency (NUtE) in Rapeseed (Brassica napus L.). Int. J. Biol. Macromol. 2024, 254, 127771. [Google Scholar] [CrossRef]
- Guo, Y.; Kong, F.; Xu, Y.; Zhao, Y.; Liang, X.; Wang, Y.; An, D.; Li, S. QTL Mapping for Seedling Traits in Wheat Grown Under Varying Concentrations of N, P and K Nutrients. Theor. Appl. Genet. 2012, 124, 851–865. [Google Scholar] [CrossRef] [PubMed]
- Mahajan, R.; Kapoor, N. Molecular Breeding Strategies for Genetic Improvement in Rice (Oryza sativa L.). In Advances in Plant Breeding Strategies: Cereals; Al-Khayri, J.M., Jain, S.M., Johnson, D.V., Eds.; Springer International Publishing: Cham, Switzerland, 2019; pp. 317–341. ISBN 978-3-030-23107-1. [Google Scholar]
- Kumar, R.; Vinod, K.K.; Krishnan, S.G.; Kumar, D.; Mehrotra, S.; Sathee, L.; Ellur, R.K.; Singh, A.K.; Bhowmick, P.K.; Bollinedi, H.; et al. Meta-QTLs Linked to Nitrogen Use Efficiency Are Randomly Distributed in Indian Rice Germplasm. Indian J. Genet. Plant Breed. 2022, 82, 7–15. [Google Scholar] [CrossRef]
- Ogawa, S.; Valencia, M.O.; Lorieux, M.; Arbelaez, J.D.; McCouch, S.; Ishitani, M.; Selvaraj, M.G. Identification of QTLs Associated with Agronomic Performance under Nitrogen-Deficient Conditions Using Chromosome Segment Substitution Lines of a Wild Rice Relative, Oryza Rufipogon. Acta Physiol. Plant 2016, 38, 103. [Google Scholar] [CrossRef]
- Neeraja, C.N.; Subramanyam, D.; Surekha, K.; Rao, P.R.; Rao, L.V.S.; Babu, M.B.B.P.; Voleti, S.R. Advances in Genetic Basis of Nitrogen Use Efficiency of Rice. Indian J. Plant Physiol. 2016, 21, 504–513. [Google Scholar] [CrossRef]
- Li, J.; Zhang, X.; Sun, Y.; Zhang, J.; Du, W.; Guo, X.; Li, S.; Zhao, Y.; Xia, L. Efficient Allelic Replacement in Rice by Gene Editing: A Case Study of the NRT1.1B Gene. J. Integr. Plant Biol. 2018, 60, 536–540. [Google Scholar] [CrossRef]
- Yadav, M.R.; Kumar, S.; Lal, M.K.; Kumar, D.; Kumar, R.; Yadav, R.K.; Kumar, S.; Nanda, G.; Singh, J.; Udawat, P.; et al. Mechanistic Understanding of Leakage and Consequences and Recent Technological Advances in Improving Nitrogen Use Efficiency in Cereals. Agronomy 2023, 13, 527. [Google Scholar] [CrossRef]
- Fiaz, S.; Wang, X.; Khan, S.A.; Ahmar, S.; Noor, M.A.; Riaz, A.; Ali, K.; Abbas, F.; Mora-Poblete, F.; Figueroa, C.R.; et al. Novel Plant Breeding Techniques to Advance Nitrogen Use Efficiency in Rice: A Review. GM Crops Food 2021, 12, 627–646. [Google Scholar] [CrossRef]
- Panahabadi, R.; Ahmadikhah, A.; Farrokhi, N.; Bagheri, N. Genome-Wide Association Study (GWAS) of Germination and Post-Germination Related Seedling Traits in Rice. Euphytica 2022, 218, 112. [Google Scholar] [CrossRef]
- Zhu, C.; Gore, M.; Buckler, E.S.; Yu, J. Status and Prospects of Association Mapping in Plants. Plant Genome 2008, 1, 5–20. [Google Scholar] [CrossRef]
- Mallikarjuna, B.P.; Shettigar, N.; Radhika, D.H.; Devi, E.L.; Bhat, J.S.; Patil, B.S.; Lohithaswa, H.C.; Mallikarjuna, M.G. Genome-Wide Association Studies and Genomic Selection for Nutrient Use Efficiency in Cereals. In Next-Generation Plant Breeding Approaches for Stress Resilience in Cereal Crops; Gowdra Mallikarjuna, M., Nayaka, S.C., Kaul, T., Eds.; Springer Nature Singapore: Singapore, 2022; pp. 161–197. ISBN 978-981-19144-4-7. [Google Scholar]
- Liao, Z.; Xia, X.; Zhang, Z.; Nong, B.; Guo, H.; Feng, R.; Chen, C.; Xiong, F.; Qiu, Y.; Li, D.; et al. Genome-Wide Association Study Using Specific-Locus Amplified Fragment Sequencing Identifies New Genes Influencing Nitrogen Use Efficiency in Rice Landraces. Front. Plant Sci. 2023, 14, 1126254. [Google Scholar] [CrossRef]
- Lv, Y.; Ma, J.; Wang, Y.; Wang, Q.; Lu, X.; Hu, H.; Qian, Q.; Guo, L.; Shang, L. Loci and Natural Alleles for Low-Nitrogen-Induced Growth Response Revealed by the Genome-Wide Association Study Analysis in Rice (Oryza sativa L.). Front. Plant Sci. 2021, 12, 770736. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Peng, C.; Xu, W.; Li, Y.; Qi, X.; Zhao, M. Genome-Wide Association Study of Agronomic Traits Related to Nitrogen Use Efficiency in Henan Wheat. BMC Genom. 2024, 25, 7. [Google Scholar] [CrossRef] [PubMed]
- Karunarathne, S.D.; Han, Y.; Zhang, X.-Q.; Zhou, G.; Hill, C.B.; Chen, K.; Angessa, T.; Li, C. Genome-Wide Association Study and Identification of Candidate Genes for Nitrogen Use Efficiency in Barley (Hordeum vulgare L.). Front. Plant Sci. 2020, 11, 571912. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Subbaiah, M.; Roy, J.; Phogat, S.; Kaushik, M.; Saini, M.R.; Madhavan, J.; Sevanthi, A.M.; Mandal, P.K. Strategies to Utilize Genome Editing for Increasing Nitrogen Use Efficiency in Crops. Nucleus 2024, 67, 205–225. [Google Scholar] [CrossRef]
- Sathee, L.; Jagadhesan, B.; Pandesha, P.H.; Barman, D.; Adavi, B.S.; Nagar, S.; Krishna, G.K.; Tripathi, S.; Jha, S.K.; Chinnusamy, V. Genome Editing Targets for Improving Nutrient Use Efficiency and Nutrient Stress Adaptation. Front. Genet. 2022, 13, 900897. [Google Scholar] [CrossRef]
- Neeraja, C.N.; Barbadikar, K.M.; Mangrauthia, S.K.; Rao, P.R.; Subrahmanayam, D.; Sundaram, R.M. Genes for NUE in Rice: A Way Forward for Molecular Breeding and Genome Editing. Plant Physiol. Rep. 2021, 26, 587–599. [Google Scholar] [CrossRef]
- Karunarathne, S.D.; Han, Y.; Zhang, X.; Li, C. CRISPR/Cas9 Gene Editing and Natural Variation Analysis Demonstrate the Potential for HvARE1 in Improvement of Nitrogen Use Efficiency in Barley. J. Integr. Plant Biol. 2022, 64, 756–770. [Google Scholar] [CrossRef]
- Li, M.; Xu, J.; Gao, Z.; Tian, H.; Gao, Y.; Kariman, K. Genetically Modified Crops Are Superior in Their Nitrogen Use Efficiency-A Meta-Analysis of Three Major Cereals. Sci. Rep. 2020, 10, 8568. [Google Scholar] [CrossRef]
- Fraisier, V.; Gojon, A.; Tillard, P.; Daniel-Vedele, F. Constitutive Expression of a Putative High-Affinity Nitrate Transporter in Nicotiana Plumbaginifolia: Evidence for Post-Transcriptional Regulation by a Reduced Nitrogen Source. Plant J. 2000, 23, 489–496. [Google Scholar] [CrossRef]
- Nwachukwu, B.C.; Babalola, O.O. Metagenomics: A Tool for Exploring Key Microbiome With the Potentials for Improving Sustainable Agriculture. Front. Sustain. Food Syst. 2022, 6, 886987. [Google Scholar] [CrossRef]
- Chandrashekharaiah, P.S.; Kodgire, S.; Sanyal, D.; Dasgupta, S. Battling Climate Change: Improving Crop Productivity and Quality by Increasing Photosynthetic Efficiency, Deploying Microbiome Metagenomics, and Effectively Utilizing Digital Technology. In Climate Change and the Microbiome: Sustenance of the Ecosphere; Choudhary, D.K., Mishra, A., Varma, A., Eds.; Springer International Publishing: Cham, Switzerland, 2021; pp. 637–660. ISBN 978-3-030-76863-8. [Google Scholar]
- Sun, C.; Lu, Y.; Tang, G.; Wang, R.; Wu, H.; Zhang, J.; Cai, S.; Zhu, J.; Xiong, Q. Integrated Metagenomics and 15N Isotope Tracing Reveal the Mechanisms Through Which the Nitrogen-Planting Density Interaction Impacts Rice Root Nitrogen Uptake Efficiency. J. Soil Sci. Plant Nutr. 2024, 24, 2665–2678. [Google Scholar] [CrossRef]
- Xue, S.; Yi, X.; Peng, J.; Bak, F.; Zhang, L.; Duan, G.; Liesack, W.; Zhu, Y. Fulvic Acid Enhances Nitrogen Fixation and Retention in Paddy Soils through Microbial-Coupled Carbon and Nitrogen Cycling. Environ. Sci. Technol. 2024, 58, 18777–18787. [Google Scholar] [CrossRef]
- Wu, W.-F.; Li, X.-Y.; Chen, S.-C.; Jin, B.-J.; Wu, C.-Y.; Li, G.; Sun, C.; Zhu, Y.-G.; Lin, X.-Y. Nitrogen Fertilization Modulates Rice Phyllosphere Functional Genes and Pathogens through Fungal Communities. Sci. Total Environ. 2024, 929, 172622. [Google Scholar] [CrossRef]
- Kaviraj, M.; Kumar, U.; Chatterjee, S.; Parija, S.; Padbhushan, R.; Nayak, A.K.; Gupta, V.V.S.R. Dissimilatory Nitrate Reduction to Ammonium (DNRA): A Unique Biogeochemical Cycle to Improve Nitrogen (N) Use Efficiency and Reduce N-Loss in Rice Paddy. Rhizosphere 2024, 30, 100875. [Google Scholar] [CrossRef]
- Babajanpour Bora, A.; Ranjbar, G.; Hashemi-Petroudi, S.H.; Nematzadeh, G.; Najafi Zarini, H. The Growth Promoting Effect of Native Cyanobacterial Strains on Rice Germination, as Well as a In Silico Analysis of Ammonium Transporter (OsAMT) Gene Family. Crop Biotechnol. 2024, 14, 45–65. [Google Scholar] [CrossRef]
- Wang, R.; Tang, G.; Lu, Y.; Zhang, D.; Cai, S.; He, H.; Zhang, H.; Xiong, Q. Root Physiological and Soil Microbial Mechanisms Underlying Responses to Nitrogen Deficiency and Compensation in Indica and Japonica Rice. Physiol. Plant. 2024, 176, e14549. [Google Scholar] [CrossRef]
- Zhu, K.; Fu, J.; Zhang, Y.; Ren, W.; Zhang, W.; Gu, J.; Xu, Y.; Zhang, H.; Wang, Z.; Liu, L.; et al. Root Activity and Rhizospheric Bacteria in Response to Nitrogen Management in Rice (Oryza sativa L.). Eur. J. Agron. 2024, 159, 127294. [Google Scholar] [CrossRef]
- Ju, C.; Buresh, R.J.; Wang, Z.; Zhang, H.; Liu, L.; Yang, J.; Zhang, J. Root and Shoot Traits for Rice Varieties with Higher Grain Yield and Higher Nitrogen Use Efficiency at Lower Nitrogen Rates Application. Field Crops Res. 2015, 175, 47–55. [Google Scholar] [CrossRef]
- Singh, U.; Ladha, J.K.; Castillo, E.G.; Punzalan, G.; Tirol-Padre, A.; Duqueza, M. Genotypic Variation in Nitrogen Use Efficiency in Medium- and Long-Duration Rice. Field Crops Res. 1998, 58, 35–53. [Google Scholar] [CrossRef]
- Chen, G.; Chen, Y.; Zhao, G.; Cheng, W.; Guo, S.; Zhang, H.; Shi, W. Do High Nitrogen Use Efficiency Rice Cultivars Reduce Nitrogen Losses from Paddy Fields? Agric. Ecosyst. Environ. 2015, 209, 26–33. [Google Scholar] [CrossRef]
- Wu, L.; Yuan, S.; Huang, L.; Sun, F.; Zhu, G.; Li, G.; Fahad, S.; Peng, S.; Wang, F. Physiological Mechanisms Underlying the High-Grain Yield and High-Nitrogen Use Efficiency of Elite Rice Varieties Under a Low Rate of Nitrogen Application in China. Front. Plant Sci. 2016, 7, 1024. [Google Scholar] [CrossRef]
- Natarajan, S.; Karuppasamy, K.S.; Ramasamy, A.; Chellamuthu, T.; Sadasivam, N.; Kovilpillai, B.; Govindan, S.K.; Veerasamy, R.; Muthurajan, R. Exploring Rice Genotypes for Nitrogen Use Efficiency under Different Nitrogen Regimes. Plant Physiol. Rep. 2024, 30, 175–190. [Google Scholar] [CrossRef]
- Tyagi, A.; Chakraborty, N.; Raghuram, N. Morpho-Physiological Evaluation of Indica Rice Genotypes with Contrasting Crop Duration for Nitrogen Use Efficiency Under Graded Urea Doses. J. Plant Growth Regul. 2024, 43, 4826–4847. [Google Scholar] [CrossRef]
- Srikanth, B.; Subrahmanyam, D.; Sanjeeva Rao, D.; Narender Reddy, S.; Supriya, K.; Raghuveer Rao, P.; Surekha, K.; Sundaram, R.M.; Neeraja, C.N. Promising Physiological Traits Associated with Nitrogen Use Efficiency in Rice under Reduced N Application. Front. Plant Sci. 2023, 14, 1268739. [Google Scholar] [CrossRef]
- Gawdiya, S.; Kumar, D.; Shivay, Y.S.; Bhatia, A.; Mehrotra, S.; Chandra, M.S.; Kumawat, A.; Kumar, R.; Price, A.H.; Raghuram, N.; et al. Field-Based Evaluation of Rice Genotypes for Enhanced Growth, Yield Attributes, Yield and Grain Yield Efficiency Index in Irrigated Lowlands of the Indo-Gangetic Plains. Sustainability 2023, 15, 8793. [Google Scholar] [CrossRef]
- Padhan, B.K.; Sathee, L.; Kumar, S.; Chinnusamy, V.; Kumar, A. Variation in Nitrogen Partitioning and Reproductive Stage Nitrogen Remobilization Determines Nitrogen Grain Production Efficiency (NUEg) in Diverse Rice Genotypes Under Varying Nitrogen Supply. Front. Plant Sci. 2023, 14, 1093581. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Xiang, H.; Fu, Y.; Zhou, C.; Wang, X.; Yuan, S.; Yu, X.; Peng, S. Optimal Nitrogen Management Increases Nitrogen Use Efficiency of Direct-Seeded Double-Season Rice Using Ultrashort-Duration Cultivars. Field Crops Res. 2024, 316, 109495. [Google Scholar] [CrossRef]
- Hussain, T.; Hussain, N.; Ahmed, M.; Nualsri, C.; Duangpan, S. Impact of Nitrogen Application Rates on Upland Rice Performance, Planted under Varying Sowing Times. Sustainability 2022, 14, 1997. [Google Scholar] [CrossRef]
- Jagadhesan, B.; Meena, H.S.; Jha, S.K.; Krishna, K.G.; Kumar, S.; Elangovan, A.; Chinnusamy, V.; Kumar, A.; Sathee, L. Association of Nitrogen Utilisation Efficiency with Sustenance of Reproductive Stage Nitrogen Assimilation, Transcript Abundance and Sequence Variation of Nitrogen Metabolism Genes in Rice (Oryza sativa L.) Sub-Species. Plant Physiol. Rep. 2024, 29, 931–947. [Google Scholar] [CrossRef]
- Haefele, S.M.; Jabbar, S.M.A.; Siopongco, J.D.L.C.; Tirol-Padre, A.; Amarante, S.T.; Sta Cruz, P.C.; Cosico, W.C. Nitrogen Use Efficiency in Selected Rice (Oryza sativa L.) Genotypes under Different Water Regimes and Nitrogen Levels. Field Crops Res. 2008, 107, 137–146. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).