
A Modified Leslie–Gower Model Incorporating Beddington–DeAngelis Functional Response, Double Allee Effect and Memory Effect


Abstract
1. Introduction
2. Model Formulation
3. Preliminaries Results
3.1. Existence and Uniqueness
3.2. Non-Negativity and Boundedness
4. Equilibrium Points and Their Local Stability
- (1)
- The equilibrium points of system (6) for the weak Allee effect are as follows:
- (a)
- Both prey and predator extinction point , which always exists;
- (b)
- The predator extinction point , which always exists;
- (c)
- The prey extinction point where we have the following.Denote as always existing.
- (d)
- (2)
- The equilibrium points of system (6) for the strong Allee effect are as follows:
- (a)
- Both prey and predator extinction point , which always exists;
- (b)
- The predator extinction point , which always exists;
- (c)
- The prey extinction point where is the positive solution of the quadratic equation . The existence of is described as follows:
- (i)
- If , then the prey extinction point does not exist.
- (ii)
- If and , then there exists a unique prey extinction point, .
- (iii)
- If , then there exist two prey extinction points, i.e., the following is the case.
- (d)
- The interior point exists if where and are also all positive roots of the quartic Equation (14).
- (a)
- is always unstable.
- (b)
- is always a saddle point.
- (c)
- is locally asymptotically stable if and .
- (a)
- By substituting to (15), we obtain the Jacobian matrix.
- (b)
- By substituting to (15), we obtain the Jacobian matrix.The Jacobian matrix (17) has eigenvalues and , showing that and . Hence, is always a saddle point.
- (c)
- By evaluating (15) at , we obtain the following.The eigenvalues of (18) are as follows.Thus, and , whenever and .
- (i)
- , , and .
- (ii)
- , and if , or and .
- (a)
- is a saddle point.
- (b)
- is always locally asymptotically stable.
- (c)
- is locally asymptotically stable if and .
- (a)
- By substituting to (15), we obtain the following.It is clear that the eigenvalues of (20) are and , and and . Thus, is a saddle point.
- (b)
- The Jacobian matrix (15) evaluated at is the following:where its eigenvalues are and , since , is always locally asymptotically stable.
- (c)
- By evaluating (15) at , we acquire the following.The eigenvalues of (22) are as follows.Therefore is locally asymptotically stable if and because in this case .
- (i)
- , , and .
- (ii)
- , and if , or and ;
5. Global Stability
5.1. System with Weak Allee Effect
- (i)
- and;
- (ii)
- .
5.2. System with Strong Allee Effect
6. Numerical Simulations
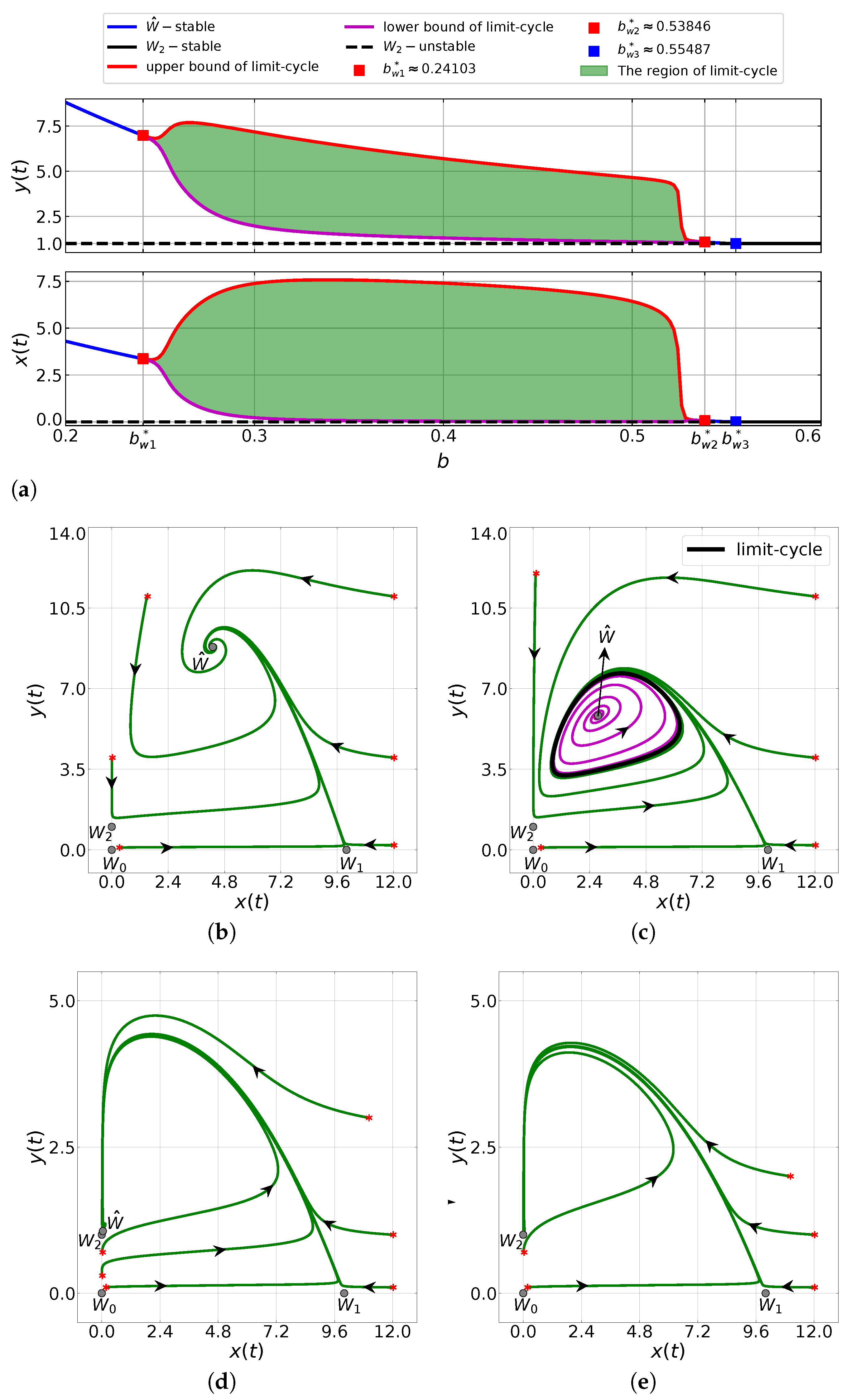
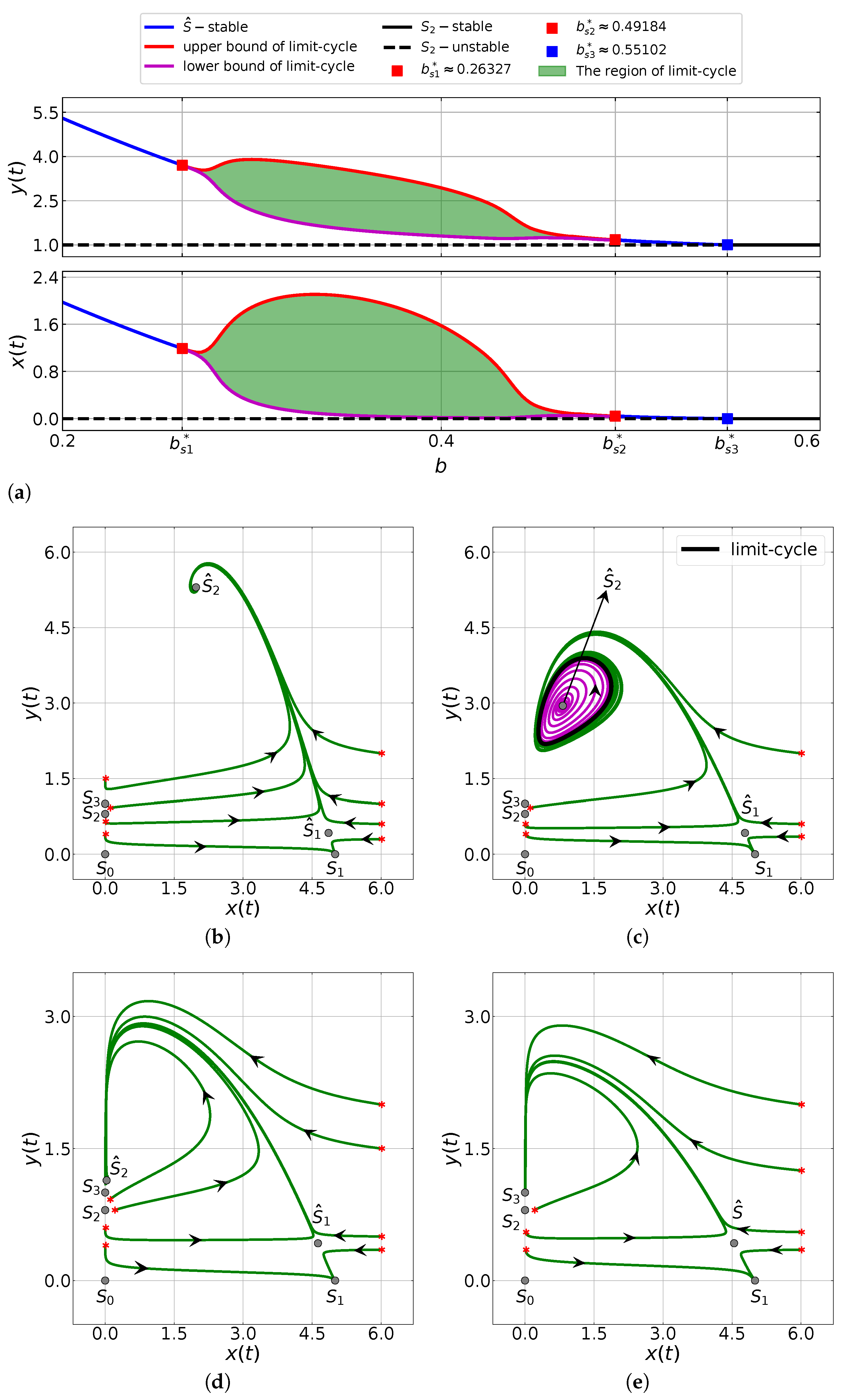
6.1. The Influence of the Capturing Rate
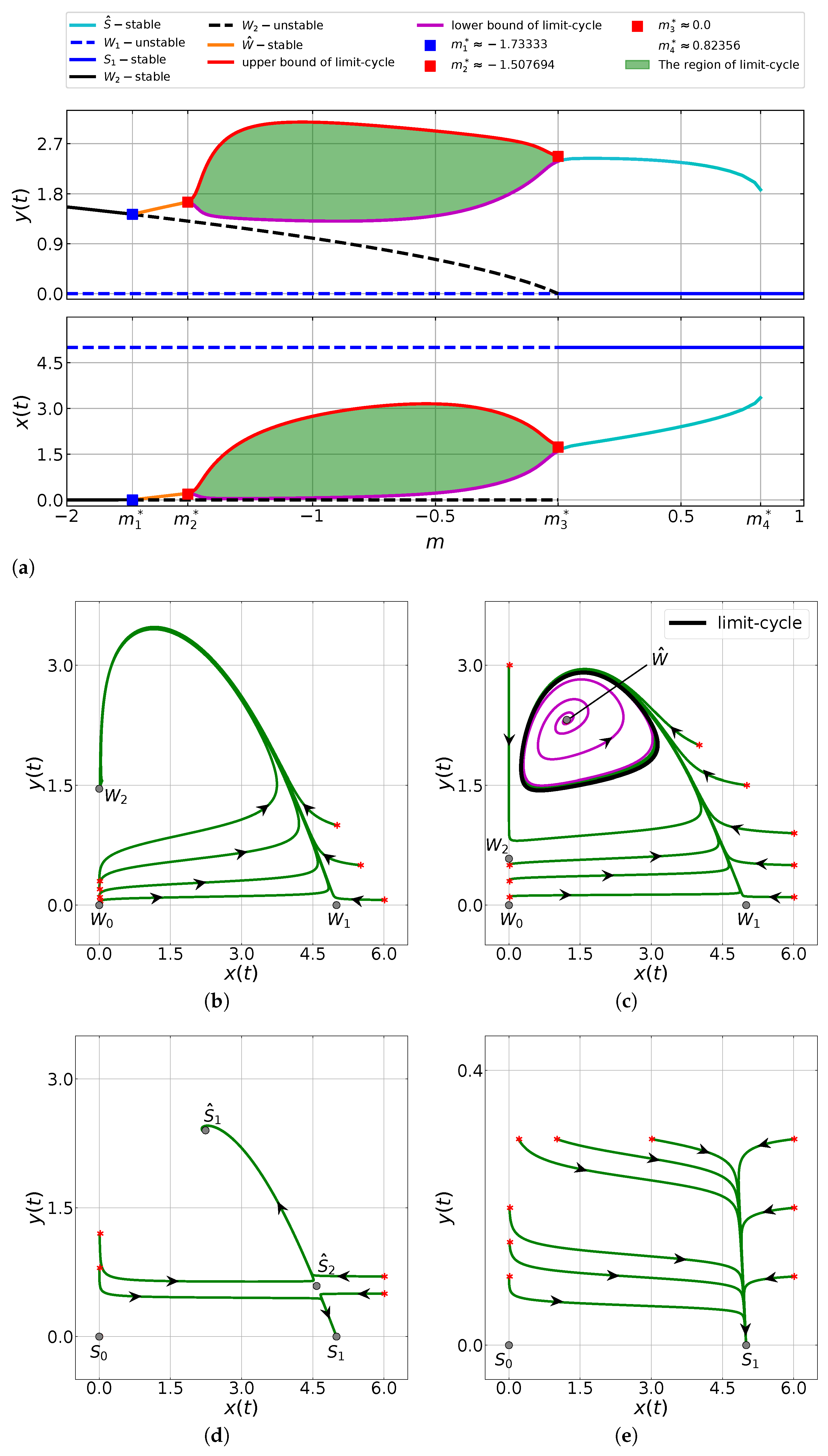
6.2. The Impacts of the Allee Threshold
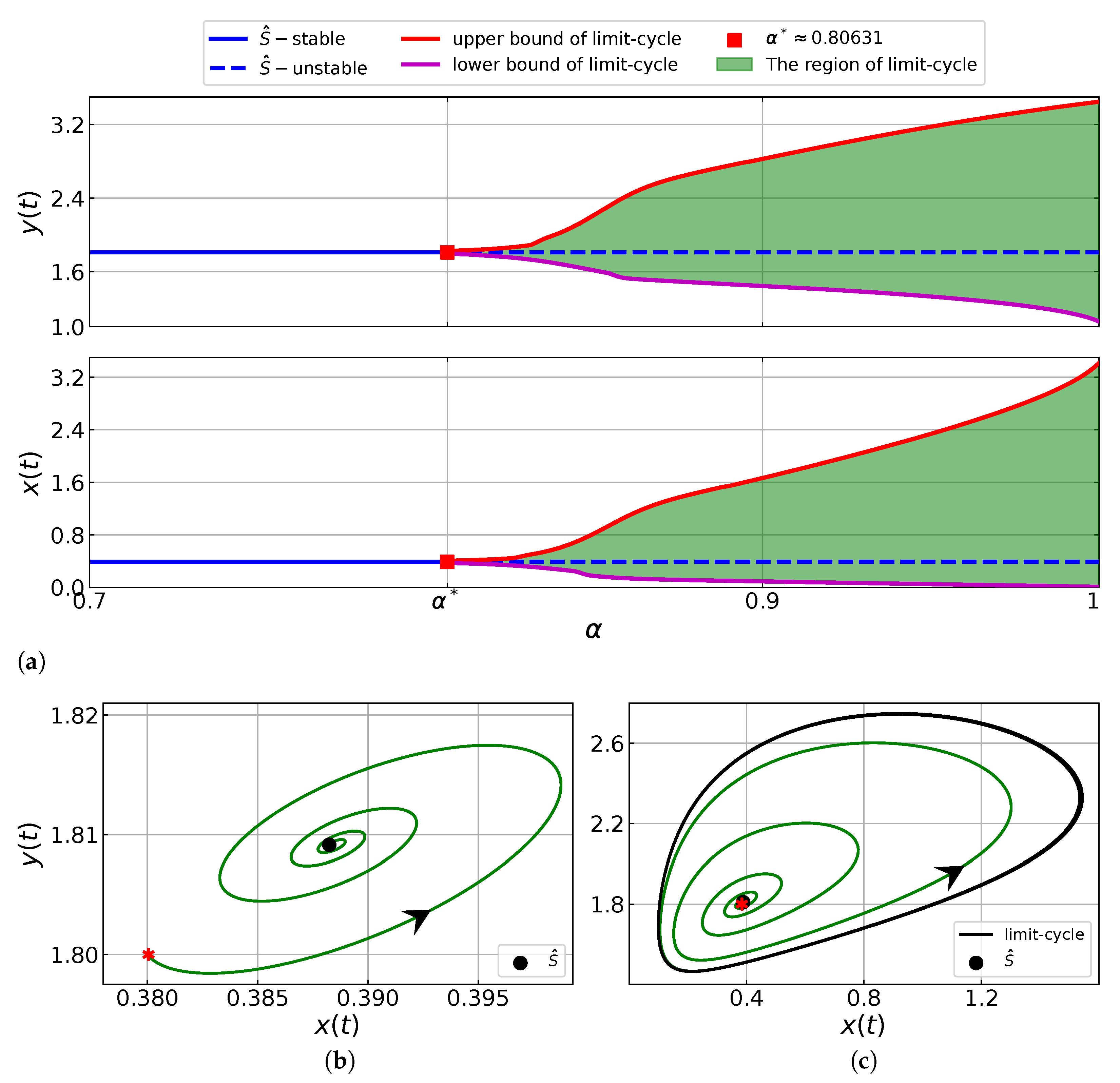
6.3. The Effects of the Order of Fractional System
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cushing, J.M. Backward bifurcations and strong Allee effects in matrix models for the dynamics of structured populations. J. Biol. Dyn. 2014, 8, 57–73. [Google Scholar] [CrossRef]
- Buffoni, G.; Groppi, M.; Soresina, C. Dynamics of predator-prey models with a strong Allee effect on the prey and predator-dependent trophic functions. Nonlinear Anal. Real World Appl. 2016, 30, 143–169. [Google Scholar] [CrossRef]
- Sasmal, S.K.; Chattopadhyay, J. An eco-epidemiological system with infected prey and predator subject to the weak Allee effect. Math. Biosci. 2013, 246, 260–271. [Google Scholar] [CrossRef]
- Saifuddin, M.; Biswas, S.; Samanta, S.; Sarkar, S.; Chattopadhyay, J. Complex dynamics of an eco-epidemiological model with different competition coefficients and weak Allee in the predator. Chaos Solitons Fractals 2016, 91, 270–285. [Google Scholar] [CrossRef]
- Pal, S.; Sasmal, S.K.; Pal, N. Chaos control in a discrete-time predator-prey model with weak Allee effect. Int. J. Biomath. 2018, 11, 1–26. [Google Scholar] [CrossRef]
- Arancibia-Ibarra, C.; Flores, J.; Bode, M.; Pettet, G.; Van Heijster, P. A modified May–Holling–Tanner predator-prey model with multiple Allee effects on the prey and an alternative food source for the predator. Discret. Contin. Dyn. Syst. 2021, 26, 943–962. [Google Scholar] [CrossRef]
- Berec, L.; Angulo, E.; Courchamp, F. Multiple Allee effects and population management. Trends Ecol. Evol. 2007, 22, 185–191. [Google Scholar] [CrossRef]
- Pavlová, V.; Berec, L.; Boukal, D.S. Caught between two Allee effects: Trade-off between reproduction and predation risk. J. Theor. Biol. 2010, 264, 787–798. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Feng, P.; Kang, Y. Dynamics of a modified Leslie–Gower model with double Allee effects. Nonlinear Dyn. 2015, 80, 1051–1062. [Google Scholar] [CrossRef]
- Lai, L.; Zhu, Z.; Chen, F. Stability and bifurcation in a predator–prey model with the additive Allee effect and the fear effect. Mathematics 2020, 8, 1280. [Google Scholar] [CrossRef]
- Martínez-Jeraldo, N.; Aguirre, P. Allee effect acting on the prey species in a Leslie–Gower predation model. Nonlinear Anal. Real World Appl. 2019, 45, 895–917. [Google Scholar] [CrossRef]
- Zhang, C.; Yang, W. Dynamic behaviors of a predator–prey model with weak additive Allee effect on prey. Nonlinear Anal. Real World Appl. 2020, 55, 103137. [Google Scholar] [CrossRef]
- Xiao, Z.; Xie, X.; Xue, Y. Stability and bifurcation in a Holling type II predator–prey model with Allee effect and time delay. Adv. Differ. Equ. 2018, 2018, 288. [Google Scholar] [CrossRef]
- Teixeira Alves, M.; Hilker, F.M. Hunting cooperation and Allee effects in predators. J. Theor. Biol. 2017, 419, 13–22. [Google Scholar] [CrossRef]
- Guan, X.; Liu, Y.U.; Xie, X. Stability analysis of a Lotka-Volterra type predator-prey system with Allee effect on the predator species. Commun. Math. Biol. Neurosci. 2018, 2018, 9. [Google Scholar] [CrossRef]
- Bodine, E.N.; Yust, A.E. Predator–prey dynamics with intraspecific competition and an Allee effect in the predator population. Lett. Biomath. 2017, 4, 23–38. [Google Scholar] [CrossRef]
- Pal, P.J.; Saha, T. Qualitative analysis of a predator-prey system with double Allee effect in prey. Chaos Solitons Fractals 2015, 73, 36–63. [Google Scholar] [CrossRef]
- Singh, M.K.; Bhadauria, B.S.; Singh, B.K. Bifurcation analysis of modified Leslie-Gower predator-prey model with double Allee effect. Ain Shams Eng. J. 2018, 9, 1263–1277. [Google Scholar] [CrossRef]
- Courchamp, F.; Clutton-Brock, T.; Grenfell, B. Multipack dynamics and the Allee effect in the African wild dog, Lycaon pictus. Anim. Conserv. 2000, 3, 277–285. [Google Scholar] [CrossRef]
- El-Shahed, M.; Ahmed, A.M.; Abdelstar, I.M.E. Dynamics of a plant-herbivore model with fractional order. Progr. Fract. Differ. Appl. 2017, 3, 59–67. [Google Scholar] [CrossRef]
- Suryanto, A.; Darti, I.; Panigoro, H.S.; Kilicman, A. A fractional-order predator–prey model with ratio-dependent functional response and linear harvesting. Mathematics 2019, 7, 1100. [Google Scholar] [CrossRef]
- Khoshsiar Ghaziani, R.; Alidousti, J.; Eshkaftaki, A.B. Stability and dynamics of a fractional order Leslie–Gower prey–predator model. Appl. Math. Model. 2016, 40, 2075–2086. [Google Scholar] [CrossRef]
- Panigoro, H.S.; Suryanto, A.; Kusumawinahyu, W.M.; Darti, I. Continuous threshold harvesting in a Gause-type predator-prey model with fractional-order. AIP Conf. Proc. 2020, 2264, 040001. [Google Scholar] [CrossRef]
- Panigoro, H.S.; Suryanto, A.; Kusumawinahyu, W.M.; Darti, I. Dynamics of an eco-epidemic predator–prey model involving fractional derivatives with power-law and Mittag–Leffler kernel. Symmetry 2021, 13, 785. [Google Scholar] [CrossRef]
- Petras, I. Fractional-Order Nonlinear Systems; Springer: London, UK, 2011. [Google Scholar]
- Kumar, S.; Kumar, R.; Cattani, C.; Samet, B. Chaotic behaviour of fractional predator-prey dynamical system. Chaos Solitons Fractals 2020, 135, 109811. [Google Scholar] [CrossRef]
- Singh, J.; Ganbari, B.; Kumar, D.; Baleanu, D. Analysis of fractional model of guava for biological pest control with memory effect. J. Adv. Res. 2021, in press. [Google Scholar] [CrossRef]
- El-Ajou, A.; Al-Smadi, M.; Oqielat, M.N.; Momani, S.; Hadid, S. Smooth expansion to solve high-order linear conformable fractional PDEs via residual power series method: Applications to physical and engineering equations. Ain Shams Eng. J. 2020, 11, 1243–1254. [Google Scholar] [CrossRef]
- Ali, H.M.; Habib, M.A.; Miah, M.M.; Akbar, M.A. Solitary wave solutions to some nonlinear fractional evolution equations in mathematical physics. Heliyon 2020, 6, e03727. [Google Scholar] [CrossRef] [PubMed]
- Shi, G.; Gao, J. European option pricing problems with fractional uncertain processes. Chaos Solitons Fractals 2021, 143, 110606. [Google Scholar] [CrossRef]
- Suryanto, A.; Darti, I.; Anam, S. Stability analysis of a fractional order modified Leslie-Gower model with additive Allee effect. Int. J. Math. Math. Sci. 2017, 2017, 8273430. [Google Scholar] [CrossRef]
- Baisad, K.; Moonchai, S. Analysis of stability and Hopf bifurcation in a fractional Gauss-type predator–prey model with Allee effect and Holling type-III functional response. Adv. Differ. Equ. 2018, 2018, 82. [Google Scholar] [CrossRef]
- Indrajaya, D.; Suryanto, A.; Alghofari, A.R. Dynamics of modified Leslie-Gower predator-prey model with Beddington-DeAngelis functional response and additive Allee effect. Int. J. Ecol. Dev. 2016, 31, 60–71. [Google Scholar]
- Ginzburg, L.R. Assuming reproduction to be a function of consumption raises doubts about some popular predator-prey models. J. Anim. Ecol. 1998, 67, 325–327. [Google Scholar] [CrossRef]
- Leslie, P.H.; Gower, J.C. The properties of a stochastic model for the predator-prey type of interaction between two species. Biometrika 1960, 47, 219–234. [Google Scholar] [CrossRef]
- Mallet, J. The struggle for existence: How the notion of carrying capacity, K, obscures the links between demography, Darwinian evolution, and speciation. Evol. Ecol. Res 2012, 14, 627–665. [Google Scholar]
- Gabriel, J.P.; Saucy, F.; Bersier, L.-F. Paradoxes in the logistic equation? Ecol. Model. 2005, 185, 147–151. [Google Scholar] [CrossRef]
- Arditi, R.; Bersier, L.-F.; Rohr, R.P. The perfect mixing paradox and the logistic equation: Verhulst vs. Lotka. Ecosphere 2016, 7, e01599. [Google Scholar] [CrossRef]
- Aziz-Alaoui, M.A.; Okiye, M.D. Boundedness and Global Stability for a Predator-Prey Model with Modified and Holling-Type II Schemes. Appl. Math. Lett. 2002, 16, 1069–1075. [Google Scholar] [CrossRef]
- Yu, S. Global stability of a modified Leslie-Gower model with Beddington-DeAngelis functional response. Adv. Differ. Equ. 2014, 2014, 84. [Google Scholar] [CrossRef]
- Vera-Damián, Y.; Vidal, C.; González-Olivares, E. Dynamics and bifurcations of a modified Leslie-Gower–type model considering a Beddington-DeAngelis functional response. Math. Methods Appl. Sci. 2019, 42, 3179–3210. [Google Scholar] [CrossRef]
- Melese, D.; Feyissa, S. Stability and bifurcation analysis of a diffusive modified Leslie-Gower prey-predator model with prey infection and Beddington DeAngelis functional response. Heliyon 2021, 7, e06193. [Google Scholar] [CrossRef]
- Arancibia-Ibarra, C.; Flores, J. Dynamics of a Leslie–Gower predator–prey model with Holling type II functional response, Allee effect and a generalist predator. Math. Comput. Simul. 2021, 188, 1–22. [Google Scholar] [CrossRef]
- Tiwari, B.; Raw, S.N. Dynamics of Leslie–Gower model with double Allee effect on prey and mutual interference among predators. Nonlinear Dyn. 2021, 1229–1257, 1229–1257. [Google Scholar] [CrossRef]
- Zhou, S.R.; Liu, Y.F.; Wang, G. The stability of predator–prey systems subject to the Allee effects. Theor. Popul. Biol. 2005, 67, 23–31. [Google Scholar] [CrossRef]
- Diethelm, K. A fractional calculus based model for the simulation of an outbreak of dengue fever. Nonlinear Dyn. 2013, 71, 613–619. [Google Scholar] [CrossRef]
- Moustafa, M.; Mohd, M.H.; Ismail, A.I.; Abdullah, F.A. Dynamical analysis of a fractional-order eco-epidemiological model with disease in prey population. Adv. Differ. Equ. 2020, 2020, 48. [Google Scholar] [CrossRef]
- Panigoro, H.S.; Suryanto, A.; Kusumawinahyu, W.M.; Darti, I. A Rosenzweig–MacArthur model with continuous threshold harvesting in predator involving fractional derivatives with power law and mittag–leffler kernel. Axioms 2020, 9, 122. [Google Scholar] [CrossRef]
- Rahmi, E.; Darti, I.; Suryanto, A.; Trisilowati; Panigoro, H.S. Stability analysis of a fractional-order Leslie-Gower model with Allee effect in predator. J. Phys. Conf. Ser. 2021, 1821, 012051. [Google Scholar] [CrossRef]
- Li, Y.; Chen, Y.Q.; Podlubny, I. Stability of fractional-order nonlinear dynamic systems: Lyapunov direct method and generalized Mittag-Leffler stability. Comput. Math. Appl. 2010, 59, 1810–1821. [Google Scholar] [CrossRef]
- Li, H.L.; Zhang, L.; Hu, C.; Jiang, Y.L.; Teng, Z. Dynamical analysis of a fractional-order predator-prey model incorporating a prey refuge. J. Appl. Math. Comput. 2017, 54, 435–449. [Google Scholar] [CrossRef]
- Ahmed, E.; El-Sayed, A.; El-Saka, H.A. On some Routh–Hurwitz conditions for fractional order differential equations and their applications in Lorenz, Rössler, Chua and Chen systems. Phys. Lett. A 2006, 358, 1–4. [Google Scholar] [CrossRef]
- Li, X.; Wu, R. Hopf bifurcation analysis of a new commensurate fractional-order hyperchaotic system. Nonlinear Dyn. 2014, 78, 279–288. [Google Scholar] [CrossRef]
- Vargas-De-León, C. Volterra-type Lyapunov functions for fractional-order epidemic systems. Commun. Nonlinear. Sci. Numer. Simul. 2015, 24, 75–85. [Google Scholar] [CrossRef]
- Huo, J.; Zhao, H.; Zhu, L. The effect of vaccines on backward bifurcation in a fractional order HIV model. Nonlinear Anal. Real World Appl. 2015, 26, 289–305. [Google Scholar] [CrossRef]
- Arenas, A.J.; González-Parra, G.; Chen-Charpentier, B.M. Construction of nonstandard finite difference schemes for the SI and SIR epidemic models of fractional order. Math. Comput. Simul 2016, 121, 48–63. [Google Scholar] [CrossRef]
- Barman, D.; Roy, J.; Alrabaiah, H.; Panja, P.; Mondal, S.P.; Alam, S. Impact of predator incited fear and prey refuge in a fractional order prey predator model. Chaos Solitons Fractals 2020, 142, 110420. [Google Scholar] [CrossRef]
- Das, M.; Samanta, G.P. A prey-predator fractional order model with fear effect and group defense. Int. J. Dyn. Control 2020, 9, 334–349. [Google Scholar] [CrossRef]
- Alidousti, J.; Ghafari, E. Dynamic behavior of a fractional order prey-predator model with group defense. Chaos Solitons Fractals 2020, 134, 109688. [Google Scholar] [CrossRef]
- Jena, R.M.; Chakraverty, S.; Baleanu, D.; Jena, S.K. Analysis of time-fractional dynamical model of romantic and interpersonal relationships with non-singular kernels: A comparative study. Math. Meth. Appl. Sci. 2020, 44, 2183–2199. [Google Scholar] [CrossRef]
- Ullah, I.; Ahmad, S.; Rahman, M.; Arfan, M. Investigation of fractional order tuberculosis (TB) model via Caputo derivative. Chaos Solitons Fractals 2020, 142, 110479. [Google Scholar] [CrossRef]
- Mahmood, S.; Shah, R.; Khan, H.; Arif, M. Laplace adomian decomposition method for multi dimensional time fractional model of Navier-Stokes equation. Symmetry 2019, 11, 149. [Google Scholar] [CrossRef]
- Diethelm, K.; Ford, N.J.; Freed, A.D. A predictor-corrector approach for the numerical solution of fractional differential equations. Nonlinear Dyn. 2002, 29, 3–22. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rahmi, E.; Darti, I.; Suryanto, A.; Trisilowati. A Modified Leslie–Gower Model Incorporating Beddington–DeAngelis Functional Response, Double Allee Effect and Memory Effect. Fractal Fract. 2021, 5, 84. https://doi.org/10.3390/fractalfract5030084
Rahmi E, Darti I, Suryanto A, Trisilowati. A Modified Leslie–Gower Model Incorporating Beddington–DeAngelis Functional Response, Double Allee Effect and Memory Effect. Fractal and Fractional. 2021; 5(3):84. https://doi.org/10.3390/fractalfract5030084
Chicago/Turabian StyleRahmi, Emli, Isnani Darti, Agus Suryanto, and Trisilowati. 2021. "A Modified Leslie–Gower Model Incorporating Beddington–DeAngelis Functional Response, Double Allee Effect and Memory Effect" Fractal and Fractional 5, no. 3: 84. https://doi.org/10.3390/fractalfract5030084
APA StyleRahmi, E., Darti, I., Suryanto, A., & Trisilowati. (2021). A Modified Leslie–Gower Model Incorporating Beddington–DeAngelis Functional Response, Double Allee Effect and Memory Effect. Fractal and Fractional, 5(3), 84. https://doi.org/10.3390/fractalfract5030084