
Nucleic Acid Prevalence of Zoonotic Babesia in Humans, Animals and Questing Ticks, a Systematic Review and Meta-Analysis



Abstract
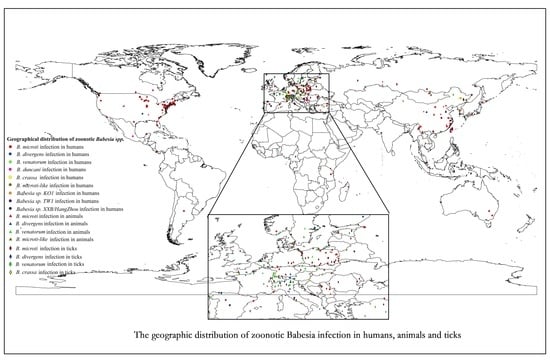
1. Introduction
2. Methods
2.1. Search Strategy and Selection Criteria
2.2. Data Extraction and Quality Assessment
2.3. Statistical Analysis
3. Results
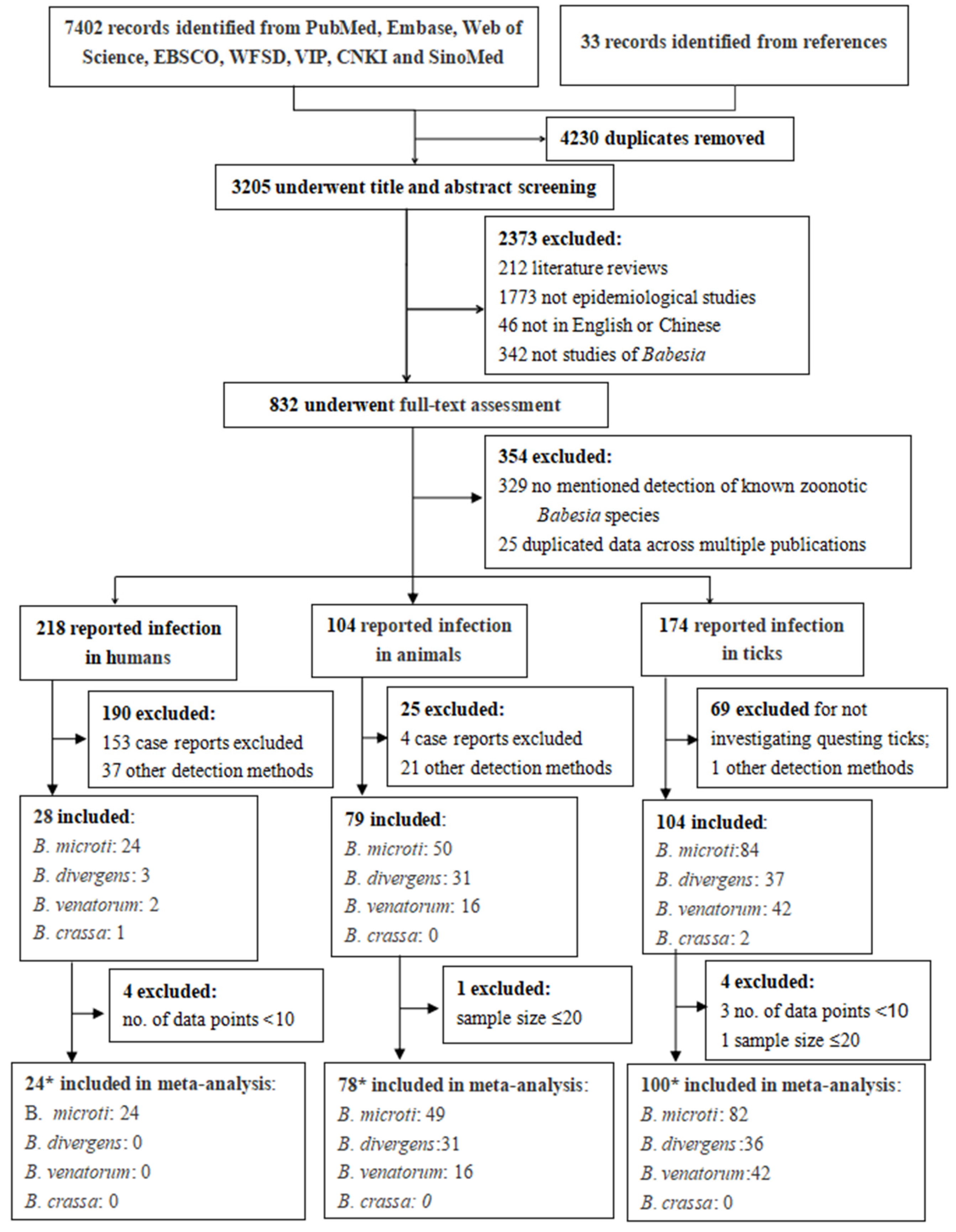
3.1. Literature Search and Selection
3.2. Prevalence of Zoonotic BABESIA in Humans
3.3. Prevalence of Zoonotic Babesia in Animals
3.4. Prevalence of Zoonotic Babesia in Questing Ticks
3.5. Publication Bias and Sensitivity Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| C.I. | Confidence Interval |
| PCR | Polymerase Chain Reaction |
References
- Remesar, S.; Díaz, P.; Prieto, A.; Markina, F.; Díaz Cao, J.M.; López-Lorenzo, G.; Fernández, G.; López, C.M.; Panadero, R.; Díez-Baños, P.; et al. Prevalence and distribution of Babesia and Theileria species in roe deer from Spain. Int. J. Parasitol. Parasites Wildl. 2019, 9, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Antunes, S.; Rosa, C.; Couto, J.; Ferrolho, J.; Domingos, A. Deciphering Babesia-Vector Interactions. Front. Cell. Infect. Microbiol. 2017, 7, 429. [Google Scholar] [CrossRef] [PubMed]
- Elsworth, B.; Duraisingh, M.T. A framework for signaling throughout the life cycle of Babesia species. Mol. Microbiol. 2021, 115, 882–890. [Google Scholar] [CrossRef] [PubMed]
- Krause, P.J. Human babesiosis. Int. J. Parasitol. 2019, 49, 165–174. [Google Scholar] [CrossRef]
- Wang, F.; Jiang, J.F.; Tian, J.; Du, C.H. Clinical characteristics, diagnosis and treatment of human babesiosis: A review. Zhongguo xue xi chong bing fang zhi za zhi = Chin. J. Schistosomiasis Control. 2020, 33, 218–224. [Google Scholar] [CrossRef]
- Laha, R.; Das, M.; Sen, A. Morphology, epidemiology, and phylogeny of Babesia: An overview. Trop. Parasitol. 2015, 5, 94–100. [Google Scholar] [CrossRef]
- Nardini, R.; Bartolomé Del Pino, L.E.; Cersini, A.; Manna, G.; Viola, M.R.; Antognetti, V.; Autorino, G.L.; Scicluna, M.T. Comparison of PCR-based methods for the detection of Babesia caballi and Theileria equi in field samples collected in Central Italy. Parasitol. Res. 2021, 120, 2157–2164. [Google Scholar] [CrossRef]
- Gabrielli, S.; Calderini, P.; Cassini, R.; Galuppi, R.; Tampieri, M.P.; Pietrobelli, M.; Cancrini, G. Human exposure to piroplasms in Central and Northern Italy. Vet. Ital. 2014, 50, 41–47. [Google Scholar] [CrossRef]
- Zanet, S.; Trisciuoglio, A.; Bottero, E.; de Mera, I.G.; Gortazar, C.; Carpignano, M.G.; Ferroglio, E. Piroplasmosis in wildlife: Babesia and Theileria affecting free-ranging ungulates and carnivores in the Italian Alps. Parasites Vectors 2014, 7, 70. [Google Scholar] [CrossRef]
- Karshima, S.N.; Karshima, M.N.; Ahmed, M.I. Global meta-analysis on Babesia infections in human population: Prevalence, distribution and species diversity. Pathog. Glob. Health 2022, 116, 220–235. [Google Scholar] [CrossRef]
- Karshima, S.N.; Karshima, M.N.; Ahmed, M.I. Infection rates, species diversity, and distribution of zoonotic Babesia parasites in ticks: A global systematic review and meta-analysis. Parasitol. Res. 2022, 121, 311–334. [Google Scholar] [CrossRef] [PubMed]
- Karshima, S.N.; Karshima, M.N.; Ahmed, M.I. Animal reservoirs of zoonotic Babesia species: A global systematic review and meta-analysis of their prevalence, distribution and species diversity. Vet. Parasitol. 2021, 298, 109539. [Google Scholar] [CrossRef] [PubMed]
- Onyiche, T.E.; Răileanu, C.; Fischer, S.; Silaghi, C. Global Distribution of Babesia Species in Questing Ticks: A Systematic Review and Meta-Analysis Based on Published Literature. Pathogens 2021, 10, 230. [Google Scholar] [CrossRef]
- Fischer, C.; Jo, W.K.; Haage, V.; Moreira-Soto, A.; de Oliveira Filho, E.F.; Drexler, J.F. Challenges towards serologic diagnostics of emerging arboviruses. Clin. Microbiol. Infect.: Off. Publ. Eur. Soc. Clin. Microbiol. Infect. Dis. 2021, 27, 1221–1229. [Google Scholar] [CrossRef]
- Yu, A.C.; Vatcher, G.; Yue, X.; Dong, Y.; Li, M.H.; Tam, P.H.; Tsang, P.Y.; Wong, A.K.; Hui, M.H.; Yang, B.; et al. Nucleic acid-based diagnostics for infectious diseases in public health affairs. Front. Med. 2012, 6, 173–186. [Google Scholar] [CrossRef] [PubMed]
- Overzier, E.; Pfister, K.; Herb, I.; Mahling, M.; Böck, G.J.; Silaghi, C. Detection of tick-borne pathogens in roe deer (Capreolus capreolus), in questing ticks (Ixodes ricinus), and in ticks infesting roe deer in southern Germany. Ticks Tick-Borne Dis. 2013, 4, 320–328. [Google Scholar] [CrossRef] [PubMed]
- Lai, M.C.; Kassee, C.; Besney, R.; Bonato, S.; Hull, L.; Mandy, W.; Szatmari, P.; Ameis, S.H. Prevalence of co-occurring mental health diagnoses in the autism population: A systematic review and meta-analysis. Lancet Psychiatry 2019, 6, 819–829. [Google Scholar] [CrossRef]
- Farkas, R.; Takács, N.; Hornyák, Á.; Nachum-Biala, Y.; Hornok, S.; Baneth, G. First report on Babesia cf. microti infection of red foxes (Vulpes vulpes) from Hungary. Parasites Vectors 2015, 8, 55. [Google Scholar] [CrossRef]
- Lee, S.; Hong, Y.; Chung, D.I.; Jang, H.K.; Goo, Y.K.; Xuan, X. Evolutionary analysis of Babesia vulpes and Babesia microti-like parasites. Parasites Vectors 2022, 15, 404. [Google Scholar] [CrossRef]
- Sun, X.; Zhen, X.; Hu, X.; Li, Y.; Gu, S.; Gu, Y.; Dong, H. Osteoarthritis in the Middle-Aged and Elderly in China: Prevalence and Influencing Factors. Int. J. Environ. Res. Public Health 2019, 16, 4701. [Google Scholar] [CrossRef]
- Hoy, D.; Brooks, P.; Woolf, A.; Blyth, F.; March, L.; Bain, C.; Baker, P.; Smith, E.; Buchbinder, R. Assessing risk of bias in prevalence studies: Modification of an existing tool and evidence of interrater agreement. J. Clin. Epidemiol. 2012, 65, 934–939. [Google Scholar] [CrossRef]
- Barendregt, J.J.; Doi, S.A.; Lee, Y.Y.; Norman, R.E.; Vos, T. Meta-analysis of prevalence. J. Epidemiol. Community Health 2013, 67, 974–978. [Google Scholar] [CrossRef] [PubMed]
- Crisafulli, S.; Sultana, J.; Fontana, A.; Salvo, F.; Messina, S.; Trifirò, G. Global epidemiology of Duchenne muscular dystrophy: An updated systematic review and meta-analysis. Orphanet J. Rare Dis. 2020, 15, 141. [Google Scholar] [CrossRef] [PubMed]
- Melsen, W.G.; Bootsma, M.C.; Rovers, M.M.; Bonten, M.J. The effects of clinical and statistical heterogeneity on the predictive values of results from meta-analyses. Clin. Microbiol. Infect.: Off. Publ. Eur. Soc. Clin. Microbiol. Infect. Dis. 2014, 20, 123–129. [Google Scholar] [CrossRef]
- Kanters, S. Fixed- and Random-Effects Models. In Methods in Molecular Biology; Humana: Clifton, NJ, USA, 2022; Volume 2345, pp. 41–65. [Google Scholar] [CrossRef]
- Aubin, J.M.; Rekman, J.; Vandenbroucke-Menu, F.; Lapointe, R.; Fairfull-Smith, R.J.; Mimeault, R.; Balaa, F.K.; Martel, G. Systematic review and meta-analysis of liver resection for metastatic melanoma. Br. J. Surg. 2013, 100, 1138–1147. [Google Scholar] [CrossRef]
- Yong, P.J.; Matwani, S.; Brace, C.; Quaiattini, A.; Bedaiwy, M.A.; Albert, A.; Allaire, C. Endometriosis and Ectopic Pregnancy: A Meta-analysis. J. Minim. Invasive Gynecol. 2020, 27, 352–361. [Google Scholar] [CrossRef] [PubMed]
- Sha, C.M.; Lehrer, E.J.; Hwang, C.; Trifiletti, D.M.; Mackley, H.B.; Drabick, J.J.; Zaorsky, N.G. Toxicity in combination immune checkpoint inhibitor and radiation therapy: A systematic review and meta-analysis. Radiother. Oncol.: J. Eur. Soc. Ther. Radiol. Oncol. 2020, 151, 141–148. [Google Scholar] [CrossRef]
- Viechtbauer, W.; Cheung, M.W. Outlier and influence diagnostics for meta-analysis. Res. Synth. Methods 2010, 1, 112–125. [Google Scholar] [CrossRef]
- Baujat, B.; Mahé, C.; Pignon, J.P.; Hill, C. A graphical method for exploring heterogeneity in meta-analyses: Application to a meta-analysis of 65 trials. Stat. Med. 2002, 21, 2641–2652. [Google Scholar] [CrossRef]
- Liu, H.B.; Wei, R.; Ni, X.B.; Zheng, Y.C.; Huo, Q.B.; Jiang, B.G.; Ma, L.; Jiang, R.R.; Lv, J.; Liu, Y.X.; et al. The prevalence and clinical characteristics of tick-borne diseases at One Sentinel Hospital in Northeastern China. Parasitology 2019, 146, 161–167. [Google Scholar] [CrossRef]
- Jahfari, S.; Hofhuis, A.; Fonville, M.; van der Giessen, J.; van Pelt, W.; Sprong, H. Molecular Detection of Tick-Borne Pathogens in Humans with Tick Bites and Erythema Migrans, in the Netherlands. PLoS Negl. Trop. Dis. 2016, 10, e0005042. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Li, H.; Gao, X.; Bian, A.; Yan, H.; Kong, D.; Liu, X. Human Babesiosis in China: A systematic review. Parasitol. Res. 2019, 118, 1103–1112. [Google Scholar] [CrossRef]
- O’Brien, S.F.; Delage, G.; Scalia, V.; Lindsay, R.; Bernier, F.; Dubuc, S.; Germain, M.; Pilot, G.; Yi, Q.L.; Fearon, M.A. Seroprevalence of Babesia microti infection in Canadian blood donors. Transfusion 2016, 56, 237–243. [Google Scholar] [CrossRef] [PubMed]
- Burkot, T.R.; Schneider, B.S.; Pieniazek, N.J.; Happ, C.M.; Rutherford, J.S.; Slemenda, S.B.; Hoffmeister, E.; Maupin, G.O.; Zeidner, N.S. Babesia microti and Borrelia bissettii transmission by Ixodes spinipalpis ticks among prairie voles, Microtus ochrogaster, in Colorado. Parasitology 2000, 121 Pt 6, 595–599. [Google Scholar] [CrossRef] [PubMed]
- Hamšíková, Z.; Kazimírová, M.; Haruštiaková, D.; Mahríková, L.; Slovák, M.; Berthová, L.; Kocianová, E.; Schnittger, L. Babesia spp. in ticks and wildlife in different habitat types of Slovakia. Parasites Vectors 2016, 9, 292. [Google Scholar] [CrossRef]
- Azagi, T.; Jaarsma, R.I.; Docters van Leeuwen, A.; Fonville, M.; Maas, M.; Franssen, F.F.J.; Kik, M.; Rijks, J.M.; Montizaan, M.G.; Groenevelt, M.; et al. Circulation of Babesia Species and Their Exposure to Humans through Ixodes ricinus. Pathogens 2021, 10, 386. [Google Scholar] [CrossRef]
- Wang, H.; Yang, J.; Mukhtar, M.U.; Liu, Z.; Zhang, M.; Wang, X. Molecular detection and identification of tick-borne bacteria and protozoans in goats and wild Siberian roe deer (Capreolus pygargus) from Heilongjiang Province, northeastern China. Parasites Vectors 2019, 12, 296. [Google Scholar] [CrossRef]
- Rar, V.A.; Epikhina, T.I.; Livanova, N.N.; Panov, V.V. Genetic diversity of Babesia in Ixodes persulcatus and small mammals from North Ural and West Siberia, Russia. Parasitology 2011, 138, 175–182. [Google Scholar] [CrossRef]
- Rar, V.; Yakimenko, V.; Makenov, M.; Tikunov, A.; Epikhina, T.; Tancev, A.; Bobrova, O.; Tikunova, N. High prevalence of Babesia microti ‘Munich’ type in small mammals from an Ixodes persulcatus/Ixodes trianguliceps sympatric area in the Omsk region, Russia. Parasitol. Res. 2016, 115, 3619–3629. [Google Scholar] [CrossRef]
- Chen, Z.; Liu, Q.; Jiao, F.C.; Xu, B.L.; Zhou, X.N. Detection of piroplasms infection in sheep, dogs and hedgehogs in Central China. Infect. Dis. Poverty 2014, 3, 18. [Google Scholar] [CrossRef]
- Jiang, J.; An, H.; Lee, J.S.; O’Guinn, M.L.; Kim, H.C.; Chong, S.T.; Zhang, Y.; Song, D.; Burrus, R.G.; Bao, Y.; et al. Molecular characterization of Haemaphysalis longicornis-borne rickettsiae, Republic of Korea and China. Ticks Tick-Borne Dis. 2018, 9, 1606–1613. [Google Scholar] [CrossRef] [PubMed]
- Jouglin, M.; Perez, G.; Butet, A.; Malandrin, L.; Bastian, S. Low prevalence of zoonotic Babesia in small mammals and Ixodes ricinus in Brittany, France. Vet. Parasitol. 2017, 238, 58–60. [Google Scholar] [CrossRef] [PubMed]
- Obiegala, A.; Pfeffer, M.; Pfister, K.; Karnath, C.; Silaghi, C. Molecular examinations of Babesia microti in rodents and rodent-attached ticks from urban and sylvatic habitats in Germany. Ticks Tick-Borne Dis. 2015, 6, 445–449. [Google Scholar] [CrossRef] [PubMed]
- Blaňarová, L.; Stanko, M.; Miklisová, D.; Víchová, B.; Mošanský, L.; Kraljik, J.; Bona, M.; Derdáková, M. Presence of Candidatus Neoehrlichia mikurensis and Babesia microti in rodents and two tick species (Ixodes ricinus and Ixodes trianguliceps) in Slovakia. Ticks Tick-Borne Dis. 2016, 7, 319–326. [Google Scholar] [CrossRef] [PubMed]
- Li, L.H.; Zhu, D.; Zhang, C.C.; Zhang, Y.; Zhou, X.N. Experimental transmission of Babesia microti by Rhipicephalus haemaphysaloides. Parasites Vectors 2016, 9, 231. [Google Scholar] [CrossRef]
- Da Rold, G.; Ravagnan, S.; Soppelsa, F.; Porcellato, E.; Soppelsa, M.; Obber, F.; Citterio, C.V.; Carlin, S.; Danesi, P.; Montarsi, F.; et al. Ticks are more suitable than red foxes for monitoring zoonotic tick-borne pathogens in northeastern Italy. Parasites Vectors 2018, 11, 137. [Google Scholar] [CrossRef]
- Georges, K.; Loria, G.R.; Riili, S.; Greco, A.; Caracappa, S.; Jongejan, F.; Sparagano, O. Detection of haemoparasites in cattle by reverse line blot hybridisation with a note on the distribution of ticks in Sicily. Vet. Parasitol. 2001, 99, 273–286. [Google Scholar] [CrossRef]
- Qi, C.; Zhou, D.; Liu, J.; Cheng, Z.; Zhang, L.; Wang, L.; Wang, Z.; Yang, D.; Wang, S.; Chai, T. Detection of Babesia divergens using molecular methods in anemic patients in Shandong Province, China. Parasitol. Res. 2011, 109, 241–245. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, S.; Yang, J.; Liu, J.; Zhang, D.; Li, Y.; Luo, J.; Guan, G.; Yin, H. Babesia divergens in human in Gansu province, China. Emerg. Microbes Infect. 2019, 8, 959–961. [Google Scholar] [CrossRef]
- Prince, H.E.; Lapé-Nixon, M.; Patel, H.; Yeh, C. Comparison of the Babesia duncani (WA1) IgG detection rates among clinical sera submitted to a reference laboratory for WA1 IgG testing and blood donor specimens from diverse geographic areas of the United States. Clin. Vaccine Immunol.: CVI 2010, 17, 1729–1733. [Google Scholar] [CrossRef]
- Kim, J.Y.; Cho, S.H.; Joo, H.N.; Tsuji, M.; Cho, S.R.; Park, I.J.; Chung, G.T.; Ju, J.W.; Cheun, H.I.; Lee, H.W.; et al. First case of human babesiosis in Korea: Detection and characterization of a novel type of Babesia sp. (KO1) similar to ovine Babesia. J. Clin. Microbiol. 2007, 45, 2084–2087. [Google Scholar] [CrossRef] [PubMed]
- Shih, C.M.; Liu, L.P.; Chung, W.C.; Ong, S.J.; Wang, C.C. Human babesiosis in Taiwan: Asymptomatic infection with a Babesia microti-like organism in a Taiwanese woman. J. Clin. Microbiol. 1997, 35, 450–454. [Google Scholar] [CrossRef] [PubMed]
- Man, S.Q.; Qiao, K.; Cui, J.; Feng, M.; Fu, Y.F.; Cheng, X.J. A case of human infection with a novel Babesia species in China. Infect. Dis. Poverty 2016, 5, 28. [Google Scholar] [CrossRef]
- Strasek-Smrdel, K.; Korva, M.; Pal, E.; Rajter, M.; Skvarc, M.; Avsic-Zupanc, T. Case of Babesia crassa-Like Infection, Slovenia, 2014. Emerg. Infect. Dis. 2020, 26, 1038–1040. [Google Scholar] [CrossRef] [PubMed]
- Doderer-Lang, C.; Filisetti, D.; Badin, J.; Delale, C.; Clavier, V.; Brunet, J.; Gommenginger, C.; Abou-Bacar, A.; Pfaff, A.W. Babesia crassa-Like Human Infection Indicating Need for Adapted PCR Diagnosis of Babesiosis, France. Emerg. Infect. Dis. 2022, 28, 449–452. [Google Scholar] [CrossRef]
- Jia, N.; Zheng, Y.C.; Jiang, J.F.; Jiang, R.R.; Jiang, B.G.; Wei, R.; Liu, H.B.; Huo, Q.B.; Sun, Y.; Chu, Y.L.; et al. Human Babesiosis Caused by a Babesia crassa-Like Pathogen: A Case Series. Clin. Infect. Dis.: Off. Publ. Infect. Dis. Soc. Am. 2018, 67, 1110–1119. [Google Scholar] [CrossRef]
- Skotarczak, B.; Wodecka, B.; Cichocka, A. Coexistence DNA of Borrelia burgdorferi sensu lato and Babesia microti in Ixodes ricinus ticks from north-western Poland. Ann. Agric. Environ. Med.: AAEM 2002, 9, 25–28. [Google Scholar]
- Skotarczak, B.; Cichocka, A. Isolation and amplification by polymerase chain reaction DNA of Babesia microti and Babesia divergens in ticks in Poland. Ann. Agric. Environ. Med.: AAEM 2001, 8, 187–189. [Google Scholar]
- Schorn, S.; Pfister, K.; Reulen, H.; Mahling, M.; Silaghi, C. Occurrence of Babesia spp., Rickettsia spp. and Bartonella spp. in Ixodes ricinus in Bavarian public parks, Germany. Parasites Vectors 2011, 4, 135. [Google Scholar] [CrossRef]
- Orkun, Ö.; Çakmak, A.; Nalbantoğlu, S.; Karaer, Z. Turkey tick news: A molecular investigation into the presence of tick-borne pathogens in host-seeking ticks in Anatolia; Initial evidence of putative vectors and pathogens, and footsteps of a secretly rising vector tick, Haemaphysalis parva. Ticks Tick-Borne Dis. 2020, 11, 101373. [Google Scholar] [CrossRef]
- Hunfeld, K.P.; Brade, V. Zoonotic Babesia: Possibly emerging pathogens to be considered for tick-infested humans in Central Europe. Int. J. Med. Microbiol.: IJMM 2004, 293 (Suppl. S37), 93–103. [Google Scholar] [CrossRef]
- Kumar, A.; O’Bryan, J.; Krause, P.J. The Global Emergence of Human Babesiosis. Pathogens 2021, 10, 1447. [Google Scholar] [CrossRef] [PubMed]
- Au, W.Y.; Cheung, P.P.H. Diagnostic performances of common nucleic acid tests for SARS-CoV-2 in hospitals and clinics: A systematic review and meta-analysis. Lancet Microbe 2021, 2, e704–e714. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.J.; Bauer, C.; Grevelding, C.G.; Quack, T. Improved PCR/nested PCR approaches with increased sensitivity and specificity for the detection of pathogens in hard ticks. Ticks Tick-Borne Dis. 2013, 4, 409–416. [Google Scholar] [CrossRef]
- Mardosaitė-Busaitienė, D.; Radzijevskaja, J.; Balčiauskas, L.; Paulauskas, A. Babesia microti in Rodents from Different Habitats of Lithuania. Animals 2021, 11, 1707. [Google Scholar] [CrossRef]
- Usluca, S.; Celebi, B.; Karasartova, D.; Gureser, A.S.; Matur, F.; Oktem, M.A.; Sozen, M.; Karatas, A.; Babur, C.; Mumcuoglu, K.Y.; et al. Molecular Survey of Babesia microti (Aconoidasida: Piroplasmida) in Wild Rodents in Turkey. J. Med. Entomol. 2019, 56, 1605–1609. [Google Scholar] [CrossRef]
- Akram, I.N.; Parveen, T.; Abrar, A.; Mehmood, A.K.; Iqbal, F. Molecular detection of Babesia microti in dogs and cat blood samples collected from Punjab (Pakistan). Trop. Biomed. 2019, 36, 304–309. [Google Scholar]
- Moustafa, M.A.M.; Sasaki, A.; Shimozuru, M.; Nakao, R.; Sashika, M.; Yamazaki, K.; Koike, S.; Tanaka, J.; Tamatani, H.; Yamanaka, M.; et al. Molecular detection of apicomplexan protozoa in Hokkaido brown bears (Ursus arctos yesoensis) and Japanese black bears (Ursus thibetanus japonicus). Parasitol. Res. 2020, 119, 3739–3753. [Google Scholar] [CrossRef]
- Spada, E.; Proverbio, D.; Galluzzo, P.; Perego, R.; Bagnagatti De Giorgi, G.; Roggero, N.; Caracappa, S. Frequency of piroplasms Babesia microti and Cytauxzoon felis in stray cats from northern Italy. BioMed Res. Int. 2014, 2014, 943754. [Google Scholar] [CrossRef]
- Camacho, A.T.; Pallas, E.; Gestal, J.J.; Guitián, F.J.; Olmeda, A.S.; Telford, S.R.; Spielman, A. Ixodes hexagonus is the main candidate as vector of Theileria annae in northwest Spain. Vet. Parasitol. 2003, 112, 157–163. [Google Scholar] [CrossRef]
- Duh, D.; Petrovec, M.; Bidovec, A.; Avsic-Zupanc, T. Cervids as Babesiae hosts, Slovenia. Emerg. Infect. Dis. 2005, 11, 1121–1123. [Google Scholar] [CrossRef]
- Mathis, A.; Hilpertshauser, H.; Deplazes, P. Piroplasms of ruminants in Switzerland and zoonotic significance of Babesia. Schweizer Archiv fur Tierheilkunde 2006, 148, 151–159. [Google Scholar] [CrossRef]
- Garafutdinov, R.R.; Galimova, A.A.; Sakhabutdinova, A.R. The influence of quality of primers on the formation of primer dimers in PCR. Nucleosides Nucleotides Nucleic Acids 2020, 39, 1251–1269. [Google Scholar] [CrossRef]
- Fischhoff, I.R.; Bowden, S.E.; Keesing, F.; Ostfeld, R.S. Correction to: Systematic review and meta-analysis of tick-borne disease risk factors in residential yards, neighborhoods, and beyond. BMC Infect. Dis. 2019, 19, 1035. [Google Scholar] [CrossRef]
- Jacob, S.S.; Sengupta, P.P.; Paramanandham, K.; Suresh, K.P.; Chamuah, J.K.; Rudramurthy, G.R.; Roy, P. Bovine babesiosis: An insight into the global perspective on the disease distribution by systematic review and meta-analysis. Vet. Parasitol. 2020, 283, 109136. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| No. of Data Points | Sample Size | No. of Positives | Pooled Prevalence, % (95% CI) | I2, % (95% CI) | R2, % | QM (p) | QE (p) | |
|---|---|---|---|---|---|---|---|---|
| B. microti | 24 | 852,344 | 933 | 1.93 (0.32; 4.69) | 98.4 (98.2; 98.7) | |||
| Type of population | 34.34 | 12.24 (0.0005) | 1215.40 (<0.0001) | |||||
| General population | 15 | 849,480 | 776 | 0.30 (0.00; 1.82) | 98.4 | |||
| High-risk population | 9 | 2864 | 157 | 7.92 (3.14; 14.49) | 97.6 | |||
| Continent | 29.06 | 12.93 (0.0116) | 1041.26 (<0.0001) | |||||
| Asia | 10 | 4790 | 203 | 7.43 (2.89; 13.72) | 97.3 | |||
| North America | 10 | 845,911 | 713 | 0.06 (0.00; 1.59) | 98.7 | |||
| Europe | 2 | 1174 | 6 | 0.33 (0.00; 7.91) | 89.2 | |||
| South America | 1 | 271 | 9 | 3.32 (0.00; 23.29) | NE | |||
| Africa | 1 | 198 | 2 | 1.01 (0.00; 17.27) | NE | |||
| Testing method | 25.34 | 9.02 (0.0110) | 1033.13 (<0.0001) | |||||
| PCR | 9 | 226,805 | 145 | 0.85 (0.00; 3.92) | 91.2 | |||
| Nested PCR | 8 | 3742 | 189 | 7.65 (2.58; 14.96) | 97.0 | |||
| Real-time PCR | 7 | 621,797 | 599 | 0.10 (0.00; 2.71) | 99.3 |
| No. of Data Points | Sample Size | No. of Positives | Pooled Prevalence, % (95% CI) | I2, % (95% CI) | R2, % | QM (p) | QE (p) | |
|---|---|---|---|---|---|---|---|---|
| B. microti | 66 | 32958 | 2815 | 7.80 (5.25; 10.77) | 98.4 (98.3; 98.6) | |||
| Taxonomy of animal host | 15.61 | 18.36 (0.0105) | 3126.73 (<0.0001) | |||||
| Rodent | 39 | 24,466 | 2344 | 10.13 (6.76; 14.07) | 98.70 | |||
| Carnivora | 5 | 1673 | 33 | 1.98 (0.00; 9.50) | 68.74 | |||
| Soricomorpha | 8 | 1132 | 259 | 14.81 (6.14; 26.16) | 95.40 | |||
| Primate | 4 | 2354 | 150 | 8.05 (0.92; 20.97) | 93.50 | |||
| Artiodactyla | 5 | 2723 | 2 | 0.02 (0.00; 4.11) | 36.10 | |||
| Chiroptera | 1 | 203 | 9 | 4.43 (0.00; 31.17) | NE | |||
| Passeriformes | 2 | 121 | 18 | 9.10 (0.00; 30.97) | 96.70 | |||
| Lagomorpha | 2 | 286 | 0 | 0.00 (0.00; 9.59) | NE | |||
| Continent | 7.92 | 8.32 (0.0490) | 3835.07 (<0.0001) | |||||
| North America | 9 | 1820 | 315 | 16.84 (7.60; 28.65) | 98.10 | |||
| Europe | 26 | 14,275 | 1342 | 8.06 (3.97; 13.36) | 99.20 | |||
| Asia | 29 | 16,535 | 1122 | 6.40 (2.99; 10.91) | 96.70 | |||
| Africa | 2 | 328 | 36 | 11.55 (0.10; 35.75) | 95.50 | |||
| Sample type | 16.25 | 13.86 (0.0010) | 2710.05 (<0.0001) | |||||
| Blood | 38 | 15,608 | 2266 | 11.75 (8.07; 15.99) | 98.40 | |||
| Tissue | 20 | 10,248 | 429 | 5.24 (1.96; 9.83) | 94.10 | |||
| Blood + Tissue | 8 | 7102 | 120 | 0.79 (0.00; 4.94) | 96.50 | |||
| Nucleic acid testing method | 3.64 | 4.46 (0.1078) | 4104.60 (<0.0001) | |||||
| PCR | 29 | 10,938 | 730 | 4.83 (2.02; 8.65) | 96.50 | |||
| Nested PCR | 26 | 15,000 | 1343 | 11.16 (6.64; 16.64) | 99.50 | |||
| Real-time PCR | 11 | 7020 | 742 | 9.01 (6.64; 16.64) | 98.20 | |||
| B. divergens | 35 | 15,974 | 381 | 2.12 (0.73; 4.08) | 95.4 (94.4; 96.3) | |||
| Taxonomy of animal host | 0.52 | 4.20 (0.3790) | 556.88 (<0.0001) | |||||
| Artiodactyla | 27 | 12,219 | 379 | 3.16 (1.26; 5.77) | 95.30 | |||
| Rodent | 5 | 2647 | 2 | 0.07 (0.00; 3.91) | 56.50 | |||
| Carnivora | 1 | 812 | 0 | 0.00 (0.00; 10.20) | NE | |||
| Lagomorpha | 1 | 238 | 0 | 0.00 (0.00; 11.30) | NE | |||
| Passeriformes | 1 | 58 | 0 | 0.00 (0.00; 14.39) | NE | |||
| Continent | 0.00 | 0.22 (0.8962) | 635.06 (<0.0001) | |||||
| Europe | 27 | 13,014 | 228 | 2.16 (0.61; 4.47) | 94.70 | |||
| North America | 1 | 33 | 0 | 0.00 (0.00; 17.57) | NE | |||
| Asia | 7 | 2927 | 153 | 2.44 (0.00; 8.21) | 95.70 | |||
| Nucleic acid testing method | 0.00 | 0.59 (0.7434) | 668.94 (<0.0001) | |||||
| PCR | 22 | 9898 | 286 | 1.94 (0.33; 4.52) | 94.70 | |||
| Nested PCR | 5 | 900 | 26 | 3.92 (0.05; 11.92) | 91.10 | |||
| Real-time PCR | 8 | 5176 | 69 | 1.76 (0.00; 6.15) | 96.90 | |||
| Sample type | 4.99 | 3.64 (0.1621) | 633.72 (<0.0001) | |||||
| Blood | 24 | 9250 | 319 | 3.17 (1.19; 5.91) | 95.80 | |||
| Tissue | 5 | 1683 | 31 | 1.32 (0.00; 7.08) | 81.90 | |||
| Blood + Tissue | 6 | 5041 | 31 | 0.17 (0.00; 3.06) | 92.00 | |||
| B. venatorum | 22 | 9186 | 120 | 1.42 (0.30; 3.16) | 92.9 (90.6; 94.7) | |||
| Taxonomy of animal host | 24.60 | 9.97 (0.0410) | 198.97 (<0.0001) | |||||
| Artiodactyla | 14 | 5205 | 120 | 3.26 (1.32; 5.92) | 93.40 | |||
| Carnivora | 3 | 1121 | 0 | 0.00 (0.00; 2.39) | NE | |||
| Lagomorpha | 1 | 238 | 0 | 0.00 (0.00; 6.21) | NE | |||
| Rodent | 3 | 2564 | 0 | 0.00 (0.00; 2.01) | NE | |||
| Passeriformes | 1 | 58 | 0 | 0.00 (0.00; 8.90) | NE | |||
| Continent | 0.00 | 0.73 (0.3929) | 292.72 (<0.0001) | |||||
| Asia | 1 | 408 | 0 | 0.00 (0.00; 7.44) | 0.00 | |||
| Europe | 21 | 8778 | 120 | 1.56 (0.34; 3.43) | 93.20 | |||
| Exact testing method | 0.00 | 0.89 (0.6423) | 278.08 (<0.0001) | |||||
| PCR | 11 | 3566 | 74 | 2.20 (0.32; 5.36) | 93.60 | |||
| Real-time PCR | 8 | 5070 | 42 | 0.92 (0.00; 3.71) | 93.70 | |||
| Nested PCR | 3 | 550 | 4 | 0.53 (0.00; 6.18) | 82.20 | |||
| Sample | 7.11 | 3.30 (0.1923) | 213.88 (<0.0001) | |||||
| Blood | 10 | 2370 | 72 | 2.71 (0.52; 6.18) | 92.90 | |||
| Tissue | 6 | 1775 | 37 | 1.50 (0.00; 5.15) | 91.90 | |||
| Blood + Tissue | 6 | 5041 | 11 | 0.23 (0.00; 2.39) | 80.50 | |||
| B. microti-like | 10 | 6487 | 471 | 42.14 (20.65; 65.26) | 99.5 (99.4; 99.6) | |||
| Taxonomy of animal host | 0.00 | 0.01 (0.9349) | 1629.65 (<0.0001) | |||||
| Carnivora | 8 | 6437 | 449 | 41.70 (17.13; 68.62) | 99.60 | |||
| Rodent | 2 | 50 | 22 | 44.14 (1.59; 93.26) | 0.00 | |||
| Continent | 0.00 | 0.33 (0.8467) | 707.96 (<0.0001) | |||||
| Asia | 3 | 74 | 24 | 30.46 (0.20; 77.29) | 81.80 | |||
| North America | 3 | 5294 | 88 | 48.00 (8.35; 89.23) | 99.40 | |||
| Europe | 4 | 1119 | 359 | 46.32 (11.44; 83.40) | 99.10 | |||
| Exact testing method | 0.00 | 0.58 (0.4481) | 936.22 (<0.0001) | |||||
| PCR | 4 | 5385 | 151 | 53.38 (18.37; 86.62) | 99.50 | |||
| Nested PCR | 6 | 1102 | 320 | 34.64 (9.42; 65.40) | 98.20 | |||
| Sample | 0.00 | 0.15 (0.7006) | 957.17 (<0.0001) | |||||
| Blood | 7 | 6187 | 275 | 39.07 (13.77; 67.87) | 99.40 | |||
| Tissue | 3 | 300 | 194 | 49.48 (10.38; 88.96) | 92.70 |
| No. of Data Points | Sample Size | No. of Positives | Pooled Prevalence, % (95% CI) | I2, % (95% CI) | R2, % | QM (p) | QE (p) | |
|---|---|---|---|---|---|---|---|---|
| B. microti | 130 | 117134 | 2311 | 2.30 (1.59; 3.13) | 97.1 (96.8; 97.4) | |||
| Tick species | 0.38 | 11.13 (0.4328) | 3511.71 (<0.0001) | |||||
| Ixodes ricinus | 63 | 66,405 | 914 | 2.45 (1.43; 3.71) | 97.20 | |||
| Ixodes scapulans | 27 | 26,846 | 1005 | 4.22 (2.29; 6.67) | 96.10 | |||
| Ixodes persulcatus | 17 | 15,373 | 189 | 1.29 (0.16; 3.29) | 95.90 | |||
| Ixodes ovatus | 1 | 789 | 38 | 4.82 (0.00; 22.47) | NE | |||
| Haemaphysalis longicornis | 4 | 2678 | 113 | 3.21 (0.11; 9.75) | 98.80 | |||
| Haemaphysalis concinna | 6 | 2624 | 3 | 0.07 (0.00; 2.33) | 46.80 | |||
| Haemaphys alisqinghaiensis | 1 | 34 | 0 | 0.00 (0.00;14.30) | NE | |||
| Dermacentor reticulatus | 4 | 1339 | 38 | 2.12 (0.00; 8.10) | 82.40 | |||
| Dermacentor nuttalli | 3 | 219 | 0 | 0.00 (0.00; 4.93) | 0.00 | |||
| Rhipicephalus turanicus | 1 | 85 | 1 | 1.18 (0.00; 16.25) | NE | |||
| Rhipicephalus microplus | 2 | 372 | 6 | 1.43 (0.00; 10.11) | 59.10 | |||
| Rhipicephalus sanguineus | 1 | 370 | 4 | 1.08 (0.00; 13.97) | NE | |||
| Genus of ticks | 0.02 | 3.38 (0.3367) | 4440.02 (<0.0001) | |||||
| Ixodes spp. | 108 | 109,413 | 2146 | 2.65 (1.82; 3.61) | 97.40 | |||
| Haemaphysalis spp. | 11 | 5336 | 116 | 0.73 (0.00; 3.04) | 97.00 | |||
| Dermacentor spp. | 7 | 1558 | 38 | 0.88 (0.00; 4.66) | 77.50 | |||
| Rhipicephalus spp. | 4 | 827 | 11 | 1.26 (0.00; 6.60) | 0.00 | |||
| Instar | 0.56 | 2.77 (0.2503) | 4328.21 (<0.0001) | |||||
| adult | 65 | 41,781 | 1019 | 2.04 (1.12; 3.18) | 95.80 | |||
| nymph | 43 | 52,223 | 820 | 2.07 (1.00; 3.48) | 97.40 | |||
| not distinguished | 22 | 23,130 | 472 | 3.83 (1.75; 6.58) | 98.20 | |||
| Continent | 1.91 | 4.14 (0.1265) | 3713.68 (<0.0001) | |||||
| North America | 28 | 27,211 | 1005 | 3.95 (2.13; 6.27) | 96.20 | |||
| Europe | 78 | 73,157 | 991 | 2.08 (1.23; 3.12) | 96.60 | |||
| Asia | 24 | 16,766 | 315 | 1.40 (0.28; 3.15) | 96.90 | |||
| Exact detecting method | 0.00 | 0.55 (0.7593) | 4412.19 (<0.0001) | |||||
| Nested PCR | 44 | 26,621 | 555 | 2.73 (1.47; 4.34) | 96.50 | |||
| Real-time PCR | 25 | 34,091 | 581 | 2.00 (0.60; 4.03) | 98.10 | |||
| PCR | 61 | 56,422 | 1175 | 2.14 (1.18; 3.34) | 96.90 | |||
| B. divergens | 59 | 87,015 | 212 | 0.16 (0.05; 0.32) | 88.2 (85.6; 90.4) | |||
| Tick species | 0.00 | 3.18 (0.8683) | 464.76 (<0.0001) | |||||
| Ixodes ricinus | 37 | 66,338 | 151 | 0.16 (0.02; 0.38) | 91.00 | |||
| Ixodes persulcatus | 11 | 13,585 | 55 | 0.36 (0.05; 0.86) | 89.80 | |||
| Haemaphysalis concinna | 6 | 2624 | 3 | 0.08 (0.00; 0.64) | NE | |||
| Haemaphysalis longicornis | 1 | 390 | 3 | 0.77 (0.00; 3.87) | NE | |||
| Ixodes ovatus | 1 | 595 | 0 | 0.00 (0.00; 1.42) | NE | |||
| Ixodes scapularis | 1 | 2858 | 0 | 0.00 (0.00; 1.09) | NE | |||
| Dermacentor reticulatus | 1 | 582 | 0 | 0.00 (0.00; 1.43) | NE | |||
| Dermacentor nuttalli | 1 | 43 | 0 | 0.00 (0.00; 5.30) | NE | |||
| Genus of ticks | 0.00 | 0.41 (0.8149) | 491.71 (<0.0001) | |||||
| Ixodes spp. | 50 | 83,376 | 206 | 0.18 (0.05; 0.37) | 89.80 | |||
| Haemaphysalis spp. | 7 | 3014 | 6 | 0.14 (0.00; 0.68) | 34.40 | |||
| Dermacentor spp. | 2 | 625 | 0 | 0.00 (0.00; 0.94) | NE | |||
| Continent | 0.00 | 1.13 (0.5688) | 476.79 (<0.0001) | |||||
| Asia | 13 | 13,437 | 51 | 0.30 (0.04; 0.72) | 88.30 | |||
| Europe | 45 | 70,720 | 161 | 0.14 (0.02; 0.32) | 88.30 | |||
| North America | 1 | 2858 | 0 | 0.00 (0.00; 1.05) | NE | |||
| Instar | 18.68 | 10.69 (0.0048) | 484.91 (<0.0001) | |||||
| adult | 30 | 26,087 | 21 | 0.07 (0.00; 0.25) | 78.10 | |||
| nymph | 16 | 24,826 | 18 | 0.04 (0.00; 0.24) | 8.20 | |||
| not distinguished | 13 | 36,102 | 133 | 0.73 (0.33; 1.25) | 96.40 | |||
| Exact detecting method | 6.73 | 5.88 (0.0528) | 426.92 (<0.0001) | |||||
| Real-time PCR | 8 | 33,746 | 12 | 0.03 (0.00; 0.34) | 48.90 | |||
| PCR | 35 | 41,751 | 180 | 0.28 (0.10; 0.54) | 91.40 | |||
| Nested PCR | 16 | 11,518 | 20 | 0.06 (0.00; 0.33) | 4.60 | |||
| B. venatorum | 71 | 91,727 | 601 | 0.39 (0.26; 0.54) | 77.0 (71.2;81.6) | |||
| Tick species | 20.78 | 18.18 (0.0058) | 256.50 (<0.0001) | |||||
| Ixodes ricinus | 47 | 73,656 | 500 | 0.50 (0.34; 0.68) | 76.50 | |||
| Ixodes persulcatus | 13 | 13,712 | 100 | 0.53 (0.26; 0.88) | 79.70 | |||
| Haemaphysalis concinna | 5 | 2460 | 1 | 0.01 (0.00; 0.28) | 0.00 | |||
| Dermacentor nuttalli | 2 | 168 | 0 | 0.00 (0.00; 1.29) | 0.00 | |||
| Dermacentor reticulatus | 2 | 746 | 0 | 0.00 (0.00; 0.49) | 0.00 | |||
| Haemaphysalis longicornis | 1 | 390 | 0 | 0.00 (0.00; 0.85) | NE | |||
| Ixodes ovatus | 1 | 595 | 0 | 0.00 (0.00; 0.69) | NE | |||
| Genus of ticks | 21.91 | 14.86 (0.0006) | 265.51 (<0.0001) | |||||
| Ixodes spp. | 61 | 87,963 | 600 | 0.49 (0.35; 0.64) | 77.20 | |||
| Haemaphysalis spp. | 6 | 2850 | 1 | 0.01 (0.00; 0.23) | 0.00 | |||
| Dermacentor spp. | 4 | 914 | 0 | 0.00 (0.00; 0.27) | 0.00 | |||
| Continent | 2.16 | 2.17 (0.1404) | 299.68 (<0.0001) | |||||
| Asia | 14 | 13,164 | 75 | 0.25 (0.06; 0.54) | 80.00 | |||
| Europe | 57 | 78,563 | 526 | 0.43 (0.28; 0.60) | 76.10 | |||
| Instar | 0.00 | 2.45 (0.2936) | 280.71 (<0.0001) | |||||
| adult | 37 | 27,082 | 169 | 0.35 (0.19; 0.55) | 71.00 | |||
| nymph | 23 | 26,096 | 133 | 0.34 (0.14; 0.61) | 76.10 | |||
| not distinguished | 11 | 38,549 | 299 | 0.63 (0.28; 1.09) | 84.50 | |||
| Exact detecting method | 0.00 | 0.34 (0.8440) | 302.28 (<0.0001) | |||||
| Nested PCR | 21 | 16,672 | 124 | 0.38 (0.16; 0.66) | 75.30 | |||
| Real-time PCR | 14 | 35,652 | 228 | 0.48 (0.19; 0.86) | 85.70 | |||
| PCR | 36 | 39,403 | 249 | 0.37 (0.19; 0.58) | 73.10 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yao, X.-Y.; Yu, S.-Q.; Tian, N.; Wang, F.; Li, S.-Z.; Li, L.-H. Nucleic Acid Prevalence of Zoonotic Babesia in Humans, Animals and Questing Ticks, a Systematic Review and Meta-Analysis. Trop. Med. Infect. Dis. 2023, 8, 132. https://doi.org/10.3390/tropicalmed8030132
Yao X-Y, Yu S-Q, Tian N, Wang F, Li S-Z, Li L-H. Nucleic Acid Prevalence of Zoonotic Babesia in Humans, Animals and Questing Ticks, a Systematic Review and Meta-Analysis. Tropical Medicine and Infectious Disease. 2023; 8(3):132. https://doi.org/10.3390/tropicalmed8030132
Chicago/Turabian StyleYao, Xiao-Yan, Shao-Qi Yu, Na Tian, Fei Wang, Shi-Zhu Li, and Lan-Hua Li. 2023. "Nucleic Acid Prevalence of Zoonotic Babesia in Humans, Animals and Questing Ticks, a Systematic Review and Meta-Analysis" Tropical Medicine and Infectious Disease 8, no. 3: 132. https://doi.org/10.3390/tropicalmed8030132
APA StyleYao, X.-Y., Yu, S.-Q., Tian, N., Wang, F., Li, S.-Z., & Li, L.-H. (2023). Nucleic Acid Prevalence of Zoonotic Babesia in Humans, Animals and Questing Ticks, a Systematic Review and Meta-Analysis. Tropical Medicine and Infectious Disease, 8(3), 132. https://doi.org/10.3390/tropicalmed8030132