
Development of an Optimized Process for Functional Recombinant SARS-CoV-2 Spike S1 Receptor-Binding Domain Protein Produced in the Baculovirus Expression Vector System

 , , , ,
, , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Upstream Processing
2.1.1. Construction of the Recombinant pFastBac-S1RBD and Bacmid Generation
2.1.2. Insect Cell Culture
2.1.3. Transfection
2.1.4. Amplification of Baculovirus Stocks
2.1.5. Baculovirus Titration Using a Cell Viability Assay (TCID50)
2.1.6. Production of S1RBD in Shake Flasks
2.1.7. Production of S1RBD in 7L Controlled Bioreactors
2.2. Downstream Processing
2.2.1. Tangential Flow Filtration (TFF)
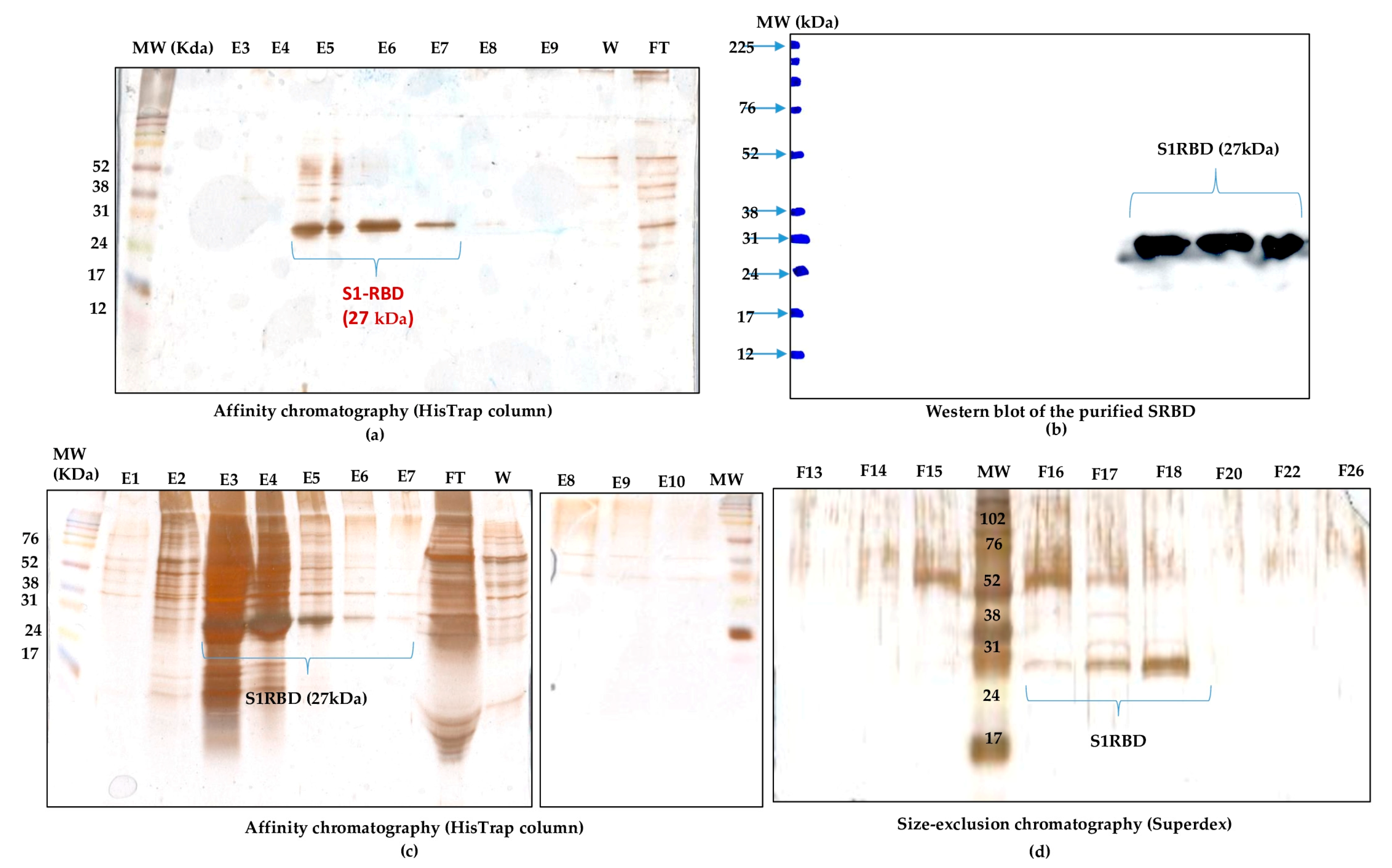
2.2.2. Protein Purification
2.2.3. SDS-PAGE
2.2.4. Western Blot Analysis
2.2.5. In-House Indirect ELISA to Detect IgG Anti-S1RBD
3. Results and Discussion
3.1. Small-Scale Production of the His-Tagged S1RBD Spike Protein in Sf9 Cells
3.1.1. Preliminary Assay
3.1.2. MOI Assay
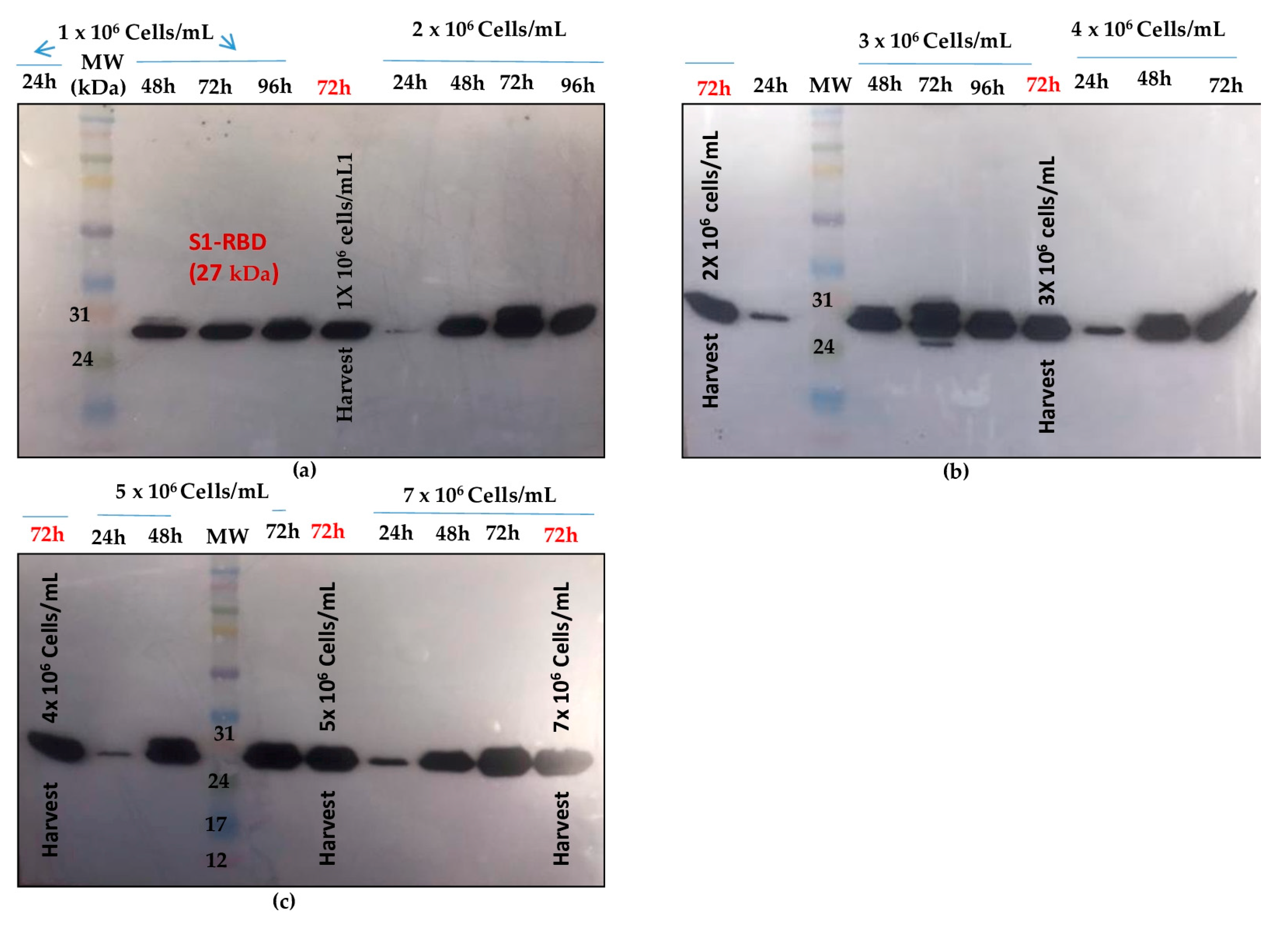
3.1.3. Cell Density at Infection Assay
3.2. Scale-Up of the Optimized Process
3.3. Downstream Process
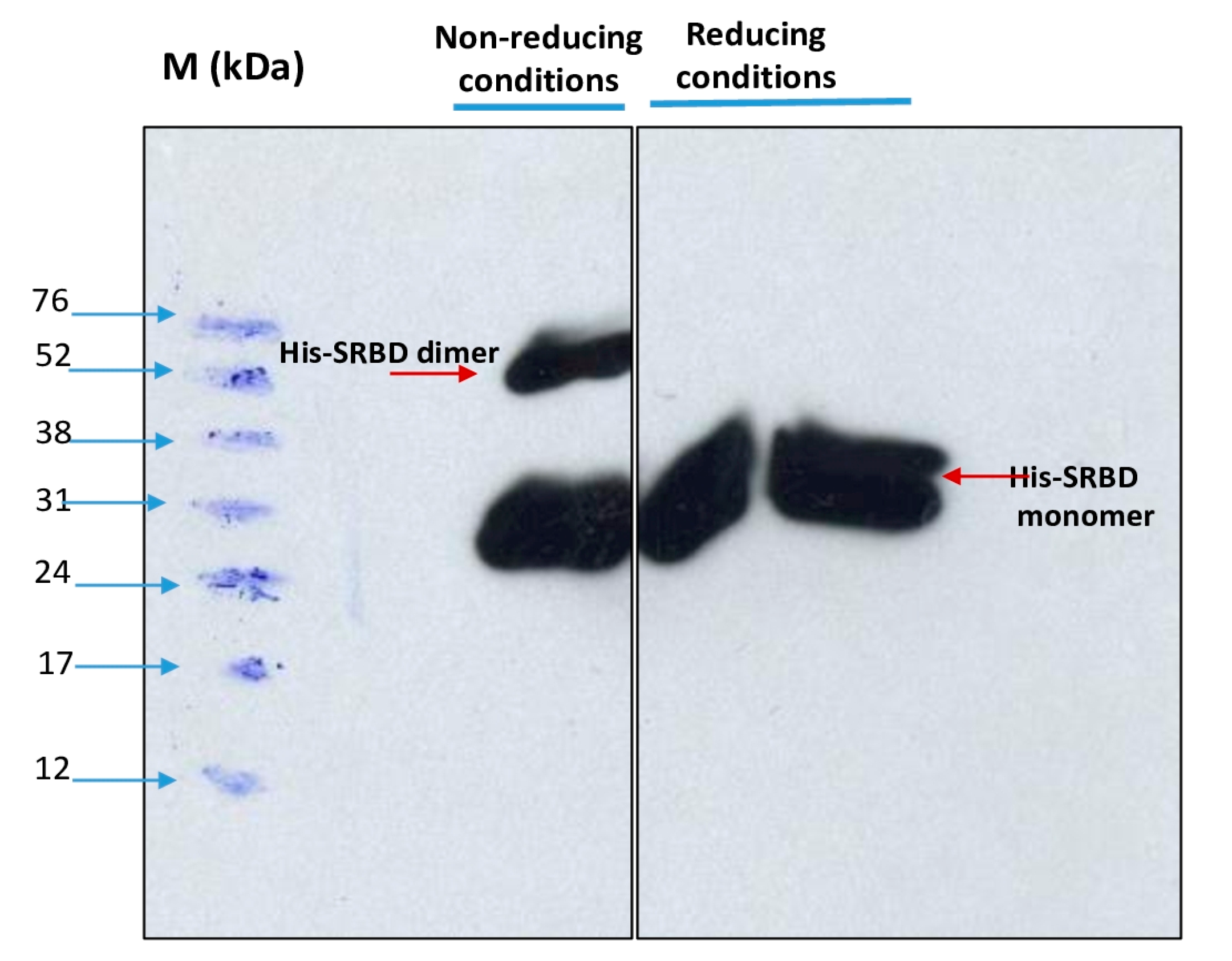
3.4. Immunoassays of the His-Tagged S1RBD Spike Protein
3.5. Development of Indirect ELISA for Anti-S1RBD IgG
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lu, R.; Zhao, X.; Li, J.; Niu, P.; Yang, B.; Wu, H.; Wang, W.; Song, H.; Huang, B.; Zhu, N.; et al. Genomic characterisation and epidemiology of 2019 novel coronavirus: Implications for virus origins and receptor binding. Lancet 2020, 395, 565–574. [Google Scholar] [CrossRef]
- Owen, D.R.; Allerton, C.M.N.; Anderson, A.S.; Aschenbrenner, L.; Avery, M.; Berritt, S.; Boras, B.; Cardin, R.D.; Carlo, A.; Coffman, K.J.; et al. An oral SARS-CoV-2 Mpro inhibitor clinical candidate for the treatment of COVID-19. Science 2021, 24, 1586–1593. [Google Scholar] [CrossRef]
- Herrera, N.G.; Morano, N.C.; Celikgil, A.; Georgiev, G.I.; Malonis, R.J.; Lee, J.H.; Tong, K.; Vergnolle, O.; Massimi, A.B.; Yen, L.Y.; et al. Characterization of the SARS-CoV-2 S Protein: Biophysical, Biochemical, Structural, and Antigenic Analysis. ACS Omega 2021, 6, 85–102. [Google Scholar] [CrossRef] [PubMed]
- Hikmet, F.; Méar, L.; Edvinsson, Å.; Micke, P.; Uhlén, M.; Lindskog, C. The protein expression profile of ACE2 in human tissues. Mol. Syst. Biol. 2020, 16, e9610. [Google Scholar] [CrossRef] [PubMed]
- Farnós, O.; Venereo-Sánchez, A.; Xu, X.; Chan, C.; Dash, S.; Chaabane, H.; Sauvageau, J.; Brahimi, F.; Saragovi, U.; Leclerc, D.; et al. Rapid High-Yield Production of Functional SARS-CoV-2 Receptor Binding Domain by Viral and Non-Viral Transient Expression for Pre-Clinical Evaluation. Vaccines 2020, 8, 654. [Google Scholar] [CrossRef]
- Li, F.; Li, W.; Farzan, M.; Harrison, S.C. Structure of SARS coronavirus spike receptor-binding domain complexed with receptor. Science 2005, 309, 1864–1868. [Google Scholar] [CrossRef]
- Walls, A.C.; Tortorici, M.A.; Snijder, J.; Xiong, X.; Bosch, B.-J.; Rey, F.A.; Veesler, D. Tectonic conformational changes of a coronavirus spike glycoprotein promote membrane fusion. Proc. Natl. Acad. Sci. USA 2017, 114, 11157–11162. [Google Scholar] [CrossRef]
- Millet, J.K.; Whittaker, G.R. Physiological and molecular triggers for SARS-CoV membrane fusion and entry into host cells. Virology 2018, 517, 3–8. [Google Scholar] [CrossRef] [PubMed]
- Lan, J.; Ge, J.; Yu, J.; Shan, S.; Zhou, H.; Fan, S.; Zhang, Q.; Shi, X.; Wang, Q.; Zhang, L.; et al. Structure of the SARS-CoV-2 spike receptor-binding domain bound to the ACE2 receptor. Nature 2020, 581, 215–220. [Google Scholar] [CrossRef]
- Li, Q.; Wu, J.; Nie, J.; Zhang Li Hao, H.; Liu, S.; Zhao, C.; Zhang, Q.; Liu, H.; Nie, L.; Qin, H.; et al. The Impact of Mutations in SARS-CoV-2 Spike on Viral Infectivity and Antigenicity. Cell 2020, 182, 1284–1294.e9. [Google Scholar] [CrossRef] [PubMed]
- Rees-Spear, C.; Muir, L.; Griffith, S.A.; Heaney, J.; Aldon, Y.; Snitselaar, J.L.; Thomas, P.; Graham, C.; Seow, J.; Lee, N.; et al. The impact of Spike mutations on SARS-CoV-2 neutralization. bioRxiv 2021. [Google Scholar] [CrossRef]
- Xie, Y.; Karki, C.B.; Du, D.; Li, H.; Wang, J.; Sobitan, A.; Teng, S.; Tang, Q.; Li, L. Spike Proteins of SARS-CoV and SARS-CoV-2 Utilize Different Mechanisms to Bind with Human ACE2. Front. Mol. Biosci. 2020, 7, 591873. [Google Scholar] [CrossRef]
- Liu, Z.; Xu, W.; Xia, S.; Gu, C.; Wang, X.; Wang, Q.; Zhou, J.; Wu, Y.; Cai, X.; Qu, D.; et al. RBD-Fc-based COVID-19 vaccine candidate induces highly potent SARS-CoV-2 neutralizing antibody response. Signal Transduct. Target. Ther. 2020, 5, 282. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Zheng, Q.; Yu, H.; Wu, D.; Xue, W.; Xiong, H.; Huang, X.; Nie, M.; Yue, M.; Rong, R.; et al. SARS-CoV-2 spike produced in insect cells elicits high neutralization titres in non-human primates. Emerg. Microbes Infect. 2020, 9, 2076–2090. [Google Scholar] [CrossRef]
- Contreras-Gómez, A.; Sánchez-Mirón, A.; García-Camacho, F.; Molina-Grima, E.; Chisti, Y. Protein production using the baculovirus-insect cell expression system. Biotechnol. Prog. 2014, 30, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Perera, R.A.; Mok, C.K.; Tsang, O.T.; Lv, H.; Ko, R.L.; Wu, N.C.; Yuan, M.; Leung, W.S.; Chan, J.M.; Chik, T.S.; et al. Serological assays for severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2), March 2020. Eurosurveillance 2020, 25, 2000421. [Google Scholar] [CrossRef]
- Mena, J.A.; Ramírez, O.T.; Palomares, L.A. Titration of non-occluded baculovirus using a cell viability assay. Biotechniques 2003, 34, 260–262, 264. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Rourou, S.; Van Der Ark, A.; Majoul, S.; Trabelsi, K.; Van der Velden, T.; Kallel, H. A novel animal-component-free medium for rabies virus production in Vero cells grown on Cytodex 1 microcarriers in a stirred bioreactor. Appl. Microbiol. Biotechnol. 2009, 85, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Liang, F.-Y.; Lin, L.-C.; Ying, T.-H.; Yao, C.-W.; Tang, T.-K.; Chen, Y.-W.; Hou, M.-H. Immunoreactivity characterisation of the three structural regions of the human coronavirus OC43 nucleocapsid protein by Western blot: Implications for the diagnosis of coronavirus infection. J. Virol. Methods 2013, 187, 413–420. [Google Scholar] [CrossRef]
- Faburay, B.; Wilson, W.; McVey, D.S.; Drolet, B.S.; Weingartl, H.; Madden, D.; Young, A.; Ma, W.; Richt, J.A. Rift Valley Fever Virus Structural and Nonstructural Proteins: Recombinant Protein Expression and Immunoreactivity against Antisera from Sheep. Vector Borne Zoonot. Dis. 2013, 13, 619–629. [Google Scholar] [CrossRef] [PubMed]
- Carinhas, N.; Bernal, V.; Yokomizo, A.Y.; Carrondo, M.J.T.; Oliveira, R.; Alves, P.M. Baculovirus production for gene therapy: The role of cell density, multiplicity of infection and medium exchange. Appl. Microbiol. Biotechnol. 2009, 81, 1041–1049. [Google Scholar] [CrossRef]
- Radford, K.M.; Cavegn, C.; Bertrand, M.; Bernard, A.R.; Reid, S.; Greenfield, P.F. The indirect effects of multiplicity of infection on baculovirus expressed proteins in insect cells: Secreted and non-secreted products. Cytotechnology 1997, 24, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Irons, S.L.; Chambers, A.C.; Lissina, O.; King, L.A.; Possee, R.D. Protein Production Using the Baculovirus Expression System. Curr. Protoc. Protein Sci. 2018, 91, 5.5.1–5.5.22. [Google Scholar] [CrossRef]
- Wong, K.T.; Peter, C.H.; Greenfield, P.F.; Reid, S.; Nielsen, L.K. Low multiplicity infection of insect cells with a recombinant baculovirus: The cell yield concept. Biotechnol. Bioeng. 1996, 49, 659–666. [Google Scholar] [CrossRef]
- Zhang, Y.H.; Enden, G.; Merchuk, J.C. Insect cells-Baculovirus system: Factors affecting growth and low MOI infection. Biochem. Eng. J. 2005, 27, 8–16. [Google Scholar] [CrossRef]
- Mehalko, J.; Drew, M.; Snead, K.; Denson, J.-P.; Wall, V.; Taylor, T.; Sadtler, K.; Messing, S.; Gillette, W.; Esposito, D. Improved production of SARS-CoV-2 spike receptor-binding domain (RBD) for serology assays. Protein Expr. Purif. 2021, 179, 105802. [Google Scholar] [CrossRef]
- Thompson, C.M.; Montes, J.; Aucoin, M.G.; Kamen, A.A. Recombinant Protein Production in Large-Scale Agitated Bioreactors Using the Baculovirus Expression Vector System. Methods Mol. Biol. 2016, 1350, 241–261. [Google Scholar] [CrossRef]
- Castro, R.; Nobre, L.S.; Eleutério, R.P.; Thomaz, M.; Pires, A.; Monteiro, S.M.; Mendes, S.; Gomes, R.A.; Clemente, J.J.; Sousa, M.F.Q.; et al. Production of high-quality SARS-CoV-2 antigens: Impact of bioprocess and storage on glycosylation, biophysical attributes, and ELISA serologic tests performance. Biotechnol. Bioeng. 2021, 118, 2202–2219. [Google Scholar] [CrossRef]
- Tee, K.L.; Jackson, P.J.; Scarrott, J.M.; Jaffe, S.R.P.; Johnson, A.O.; Johari, Y.; Pohle, T.H.; Mozzanino, T.; Price, J.; Grinham, J.; et al. Purification of recombinant SARS-CoV-2 spike, its receptor binding domain, and CR3022 mAb for serological assay. bioRxiv 2020. [Google Scholar] [CrossRef]
- An, Y.; Li, S.; Jin, X.; Han, J.B.; Xu, K.; Xu, S.; Han, Y.; Liu, C.; Zheng, T.; Liu, M.; et al. A tandem-repeat dimeric RBD protein-based COVID-19 vaccine zf2001 protects mice and nonhuman primates. Emerg. Microbes Infect. 2022, 11, 1058–1071. [Google Scholar] [CrossRef]
- Benabdessalem, C.; Ben Hamouda, W.; Marzouki, S.; Faye, R.; Mbow, A.-A.; Diouf, B.; Ndiaye Ndongo Dia, O.; Faye, O.; Sall, A.-A.; Diagne, C.-T.; et al. Development and comparative evaluation of SARS-CoV-2 S-RBD and N based ELISA tests in various African endemic settings. Diagn. Microbiol. Infect. Dis. 2023, 105, 115903. [Google Scholar] [CrossRef]
- Cherif, I.; Kharroubi, G.; Chaabane, S.; Yazidi, R.; Dellagi, M.; Snoussi, M.A.; Salem, S.; Marzouki, S.; Kammoun Rebai, W.; Rourou, S.; et al. COVID-19 in Tunisia (North Africa): Seroprevalence of SARS-CoV-2 in the General Population of the Capital City Tunis. Diagnostics 2022, 12, 971. [Google Scholar] [CrossRef] [PubMed]
- Kharroubi, G.; Cherif, I.; Ghawar, W.; Dhaouadi, N.; Yazidi, R.; Chaabane, S.; Snoussi, M.-A.; Salem, S.; Ben Hammouda, W.; Ben Hammouda, S.; et al. Incidence and risk factors of SARS-CoV-2 infection among workers in a public health laboratory in Tunisia. Arch. Virol. 2023, 168, 69. [Google Scholar] [CrossRef]
- Yuan, M.; Wu, N.C.; Zhu, X.; Lee, C.-C.D.; So, R.T.Y.; Lv, H.; Mok, C.K.P.; Wilson, I.A. A highly conserved cryptic epitope in the receptor binding domains of SARS-CoV-2 and SARS-CoV. Science 2020, 368, 630–633. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boumaiza, M.; Chaabene, A.; Akrouti, I.; Ben Zakour, M.; Askri, H.; Salhi, S.; Ben Hamouda, W.; Marzouki, S.; Benabdessalem, C.; Ben Ahmed, M.; et al. Development of an Optimized Process for Functional Recombinant SARS-CoV-2 Spike S1 Receptor-Binding Domain Protein Produced in the Baculovirus Expression Vector System. Trop. Med. Infect. Dis. 2023, 8, 501. https://doi.org/10.3390/tropicalmed8110501
Boumaiza M, Chaabene A, Akrouti I, Ben Zakour M, Askri H, Salhi S, Ben Hamouda W, Marzouki S, Benabdessalem C, Ben Ahmed M, et al. Development of an Optimized Process for Functional Recombinant SARS-CoV-2 Spike S1 Receptor-Binding Domain Protein Produced in the Baculovirus Expression Vector System. Tropical Medicine and Infectious Disease. 2023; 8(11):501. https://doi.org/10.3390/tropicalmed8110501
Chicago/Turabian StyleBoumaiza, Mohamed, Ameni Chaabene, Ines Akrouti, Meriem Ben Zakour, Hana Askri, Said Salhi, Wafa Ben Hamouda, Soumaya Marzouki, Chaouki Benabdessalem, Melika Ben Ahmed, and et al. 2023. "Development of an Optimized Process for Functional Recombinant SARS-CoV-2 Spike S1 Receptor-Binding Domain Protein Produced in the Baculovirus Expression Vector System" Tropical Medicine and Infectious Disease 8, no. 11: 501. https://doi.org/10.3390/tropicalmed8110501
APA StyleBoumaiza, M., Chaabene, A., Akrouti, I., Ben Zakour, M., Askri, H., Salhi, S., Ben Hamouda, W., Marzouki, S., Benabdessalem, C., Ben Ahmed, M., Trabelsi, K., & Rourou, S. (2023). Development of an Optimized Process for Functional Recombinant SARS-CoV-2 Spike S1 Receptor-Binding Domain Protein Produced in the Baculovirus Expression Vector System. Tropical Medicine and Infectious Disease, 8(11), 501. https://doi.org/10.3390/tropicalmed8110501