

Characterization of Beta-Lactamase and Fluoroquinolone Resistance Determinants in Escherichia coli, Klebsiella pneumoniae, and Pseudomonas aeruginosa Isolates from a Tertiary Hospital in Yola, Nigeria


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Isolates
2.2. Bacterial Identification and Antimicrobial Susceptibility Testing
2.3. Phenotypic Detection of Carbapenemase Production
2.4. Whole Genome Sequencing
3. Results
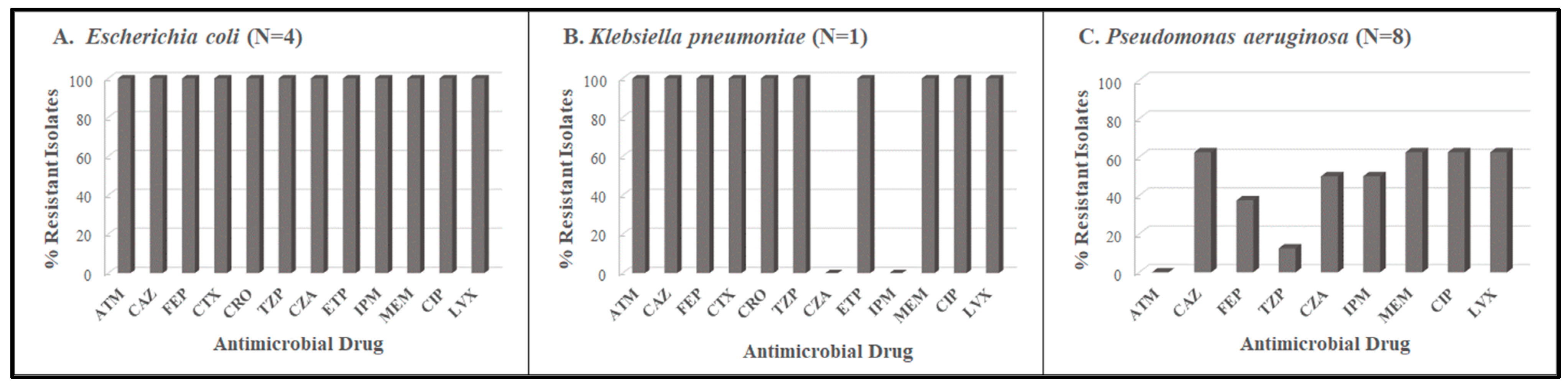
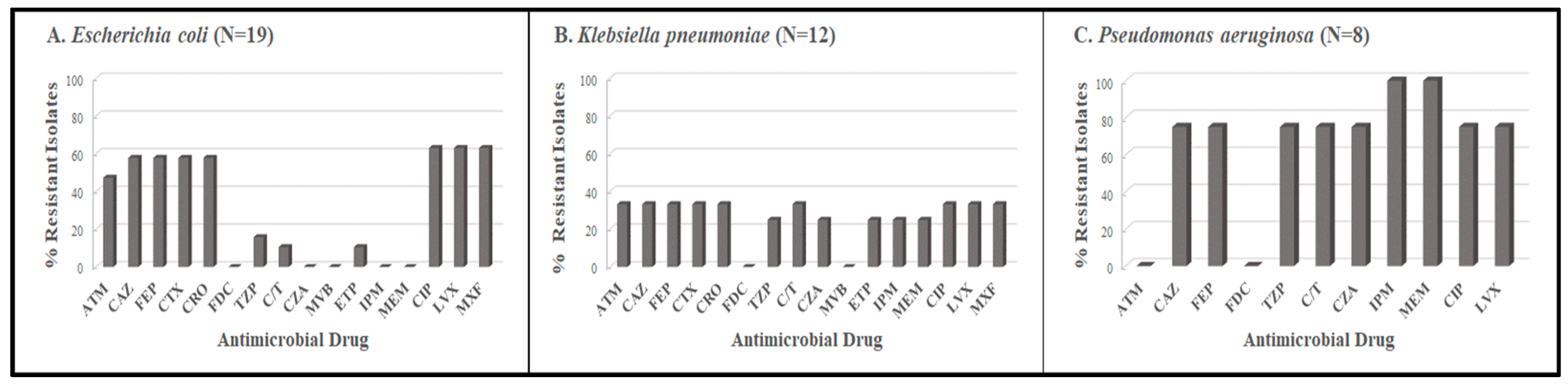
3.1. Antimicrobial Resistance Profiles
3.2. Strain Types and Resistance Determinants among E. coli Isolates
3.3. Strain Types and Resistance Determinants in K. pneumoniae Isolates
3.4. Strain Types and Resistance Determinants in P. aeruginosa Isolates
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Murray, C.J.; Ikuta, K.S.; Sharara, F. Global burden of bacterial antimicrobial resistance in 2019: A systematic analysis. Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef]
- Kariuki, S.; Kering, K.; Warimu, C. Antimicrobial Resistance Rates and Surveillance in Sub-Saharan Africa: Where Are We Now? Infect. Drug Resist. 2022, 15, 3589–3609. [Google Scholar] [CrossRef]
- Godman, B.; Egwuenu, A.; Wesangula, E. Tackling antimicrobial resistance across sub-Saharan Africa: Current challenges and implications for the future. Expert Opin. Drug Saf. 2022, 21, 1089–1111. [Google Scholar] [CrossRef]
- Saka, H.K.; García-Soto, S.; Dabo, N.T. Molecular detection of extended spectrum β-lactamase genes in Escherichia coli clinical isolates from diarrhoeic children in Kano, Nigeria. PLoS ONE 2020, 15, e0243130. [Google Scholar] [CrossRef]
- Kazmierczak, K.M.; Rabine, S.; Hackel, M. Multiyear, multinational survey of the incidence and global distribution of metallo-β-lactamase-producing Enterobacteriaceae and Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2016, 60, 1067–78. [Google Scholar] [CrossRef]
- Ogbolu, D.O.; Webber, M.A. High-level and novel mechanisms of carbapenem resistance in Gram-negative bacteria from tertiary hospitals in Nigeria. Int. J. Antimicrob. Agents. 2014, 43, 412–417. [Google Scholar] [CrossRef]
- Manenzhe, R.I.; Zar, H.J.; Nicol, M.P. The spread of carbapenemase-producing bacteria in Africa: A systematic review. J. Antimicrob. Chemother. 2015, 70, 23–40. [Google Scholar] [CrossRef] [PubMed]
- Nordmann, P.; Poirel, L. Epidemiology and diagnostics of carbapenem resistance in Gram-negative bacteria. Clin. Infect Dis. 2019, 69 (Suppl. S7), S521–S528. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, D.M.P.; Forde, B.M. Antimicrobial Resistance in ESKAPE Pathogens Clin. Microbiol. Rev. 2020, 33, e00181-19. [Google Scholar] [CrossRef] [PubMed]
- Chattaway, M.A.; Aboderin, A.O. Fluoroquinolone-resistant enteric bacteria in sub-Saharan Africa: Clones, implications and research needs. Front. Microbiol. 2016, 7, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Salah, F.D.; Soubeiga, S.T.; Ouattara, A.K. Distribution of quinolone resistance gene (qnr) in ESBL-producing Escherichia coli and Klebsiella spp. in Lomé, Togo. Antimicrob Resist. Infect. Control. 2019, 8, 104. [Google Scholar] [CrossRef] [PubMed]
- Kuo, P.Y.; Lo, Y.T.; Chiou, Y.J. Plasmid-mediated quinolone resistance determinants in fluoroquinolone-nonsusceptible Escherichia coli isolated from patients with urinary tract infections in a university hospital, 2009–2010 and 2020. J. Glob. Antimicrob. Resist. 2022, 30, 241–248. [Google Scholar] [CrossRef] [PubMed]
- Monárrez, R.; Braun, M.; Coburn-Flynn, O. A large self-transmissible resistance plasmid from Nigeria contains genes that ameliorate a carrying cost. Sci. Rep. 2019, 9, 19624. [Google Scholar] [CrossRef]
- Eghieye, M.O.; Nkene, I.H.; Abimuku, R.H. Molecular detection of plasmid-mediated quinolone resistance in ciprofloxacin-resistant Escherichia coli from urine of patients attending Garki Hospital, Abuja, Nigeria. Eur. J. Biol. Biotechnol. 2020, 1, 1–7. [Google Scholar] [CrossRef]
- Adekanmbi, A.O.; Usidamen, S.; Akinlabi, O.C. Carriage of plasmid-mediated qnr determinants and quinolone efflux pump (qepA) by ciprofloxacin-resistant bacteria recovered from urinary tract infection (UTI) samples. Bull. Natl. Res. Cent. 2022, 46, 27. [Google Scholar] [CrossRef]
- Raufu, I.A.; Fashae, K.; Ameh, J.A. Persistence of fluoroquinolone-resistant Salmonella enterica serovar Kentucky from poultry and poultry sources in Nigeria. J. Infect. Dev. Ctries. 2014, 8, 384–388. [Google Scholar] [CrossRef]
- Ogbolu, D.O.; Alli, O.A.T.; Webber, M.A. CTX-M-15 is Established in Most Multidrug-Resistant Uropathogenic Enterobacteriaceae and Pseudomonaceae from Hospitals in Nigeria. Eur. J. Microbiol. Immunol. (Bp) 2018, 8, 20–24. [Google Scholar] [CrossRef] [PubMed]
- Shettima, S.A.; Tickler, I.A.; Dela Cruz, C.M.; Tenover, F.C. Characterization of carbapenem-resistant Gram-negative organisms from clinical specimens in Yola. Nigeria. J. Glob. Antimicrob. Resist. 2020, 21, 42–5. [Google Scholar] [CrossRef]
- Tickler, I.A.; Shettima, S.A.; Dela Cruz, C.M. Characterization of Carbapenem-Resistant Gram-Negative Bacterial Isolates From Nigeria by Whole Genome Sequencing. Diagn Microbiol. Infect. Dis. 2021, 101, 115422. [Google Scholar] [CrossRef]
- Jorgensen, J.H.; Pfaller, M.A. Manual of Clinical Microbiology, 11th ed.; ASM Press: Washington, DC, USA, 2015; pp. 685–790. [Google Scholar]
- Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Disk Susceptibility Tests; Approved Standard-Thirteenth Edition M2-A13; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018. [Google Scholar]
- Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing; Thirty-second Edition; CLSI supplement M100-S32; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2022. [Google Scholar]
- Castanheira, M.; Simner, P.J.; Bradford, P.A. Extended-spectrum β-lactamases: An update on their characteristics, epidemiology and detection. JAC Antimicrob. Resist. 2021, 3, dlab092. [Google Scholar] [CrossRef]
- Medugu, N.; Aworh, M.K.; Iregbu, K. Molecular characterization of multi drug resistant Escherichia coli isolates at a tertiary hospital in Abuja, Nigeria. Sci. Rep. 2022, 12, 14822. [Google Scholar] [CrossRef]
- Tickler, I.A.; Kawa, D.; Obradovich, A.E. The Healthcare Associated Infections Consortium. Characterization of Carbapenemase- and ESBL-Producing Gram-Negative Bacilli Isolated from Patients with Urinary Tract and Bloodstream Infections. Antibiotics 2023, 12, 1386. [Google Scholar] [CrossRef]
- Medugu, N.; Tickler, I.A.; Duru, C. Phenotypic and molecular characterization of beta-lactam resistant Multidrug-resistant Enterobacterales isolated from patients attending six hospitals in Northern Nigeria. Sci. Rep. 2023, 13, 10306. [Google Scholar] [CrossRef] [PubMed]
- Nsofor, C.M.; Tattfeng, M.Y.; Nsofor, C.A. High prevalence of qnrA and qnrB genes among fluoroquinolone-resistant Escherichia coli isolates from a tertiary hospital in Southern Nigeria. Bull. Natl. Res. Cent. 2021, 45, 26. [Google Scholar] [CrossRef]
- Essack, S.Y.; Desta, A.T.; Abotsi, R.E. Antimicrobial resistance in the WHO African region: Current status and roadmap for action. J. Public. Health. 2017, 39, 8–13. [Google Scholar] [CrossRef]
- Sekyere, J.O.; Amoako, D.G. Genomic and phenotypic characterisation of fluoroquinolone resistance mechanisms in Enterobacteriaceae in Durban, South Africa. PLoS ONE 2017, 12, e0178888. [Google Scholar] [CrossRef]
- Tadesse, G.; Tessema, T.S.; Beyene, G. Molecular epidemiology of fluoroquinolone resistant Salmonella in Africa: A systematic review and meta-analysis. PLoS ONE 2018, 13, e0192575. [Google Scholar] [CrossRef]
- Kariuki, K.; Diakhate, M.M.; Musembi, S. Plasmid-mediated quinolone resistance genes detected in Ciprofloxacin non-susceptible Escherichia coli and Klebsiella isolated from children under five years at hospital discharge, Kenya. BMC Microbiol. 2023, 23, 129. [Google Scholar] [CrossRef]
- Piekarska, K.; Wołkowicz, T.; Zacharczuk, K. Co-existence of plasmid-mediated quinolone resistance determinants and mutations in gyrA and parC among fluoroquinolone-resistant clinical Enterobacteriaceae isolated in a tertiary hospital in Warsaw, Poland. Int. J. Antimicrob. Agents. 2015, 45, 238–43. [Google Scholar] [CrossRef]
- Hamed, S.M.; Elkhatib, W.F.; El-Mahallawy, H.A. Multiple mechanisms contributing to ciprofloxacin resistance among Gram negative bacteria causing infections to cancer patients. Sci. Rep. 2018, 8, 12268. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Collection Period | Organism | Blood Culture | Ear Swab | Endocervical Swab | Eye Swab | Stool | Vaginal Swab | Sputum | Urethral Swab | Urine | Wound | Total |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Period 1 November 2017–February 2018 | Escherichia coli | 1 | 3 | 4 | ||||||||
| Klebsiella pneumoniae | 1 | 1 | ||||||||||
| Pseudomonas aeruginosa | 1 | 2 | 3 | 2 | 8 | |||||||
| Period 2 October 2021–January 2022 | Escherichia coli | 3 | 12 | 4 | 19 | |||||||
| Klebsiella pneumoniae | 1 | 1 | 4 | 1 | 3 | 2 | 12 | |||||
| Pseudomonas aeruginosa | 2 | 1 | 1 | 3 | 1 | 8 | ||||||
| Total | 1 | 3 | 1 | 1 | 1 | 3 | 7 | 1 | 25 | 9 | 52 |
| ID | Organism | Source | MLST | Beta-Lactamase Genes | Fluoroquinolone PMQR Genes | Fluoroquinolone QRDR Mutations | ||
|---|---|---|---|---|---|---|---|---|
| gyrA | parC | parE | ||||||
| 15949 | Escherichia coli | ear swab | ST692 | blaCMY-59, blaCTX-M-15, blaNDM-7, blaOXA-1, blaTEM-1, ampH | aac(6′)Ib-cr | S83L, D87N | S80I | S458A |
| 16020 | Escherichia coli | urine | ST692 | blaCMY-59, blaCTX-M-15, blaNDM-7, blaOXA-10, blaOXA-140, blaTEM-1, ampH, ampC | aac(6′)Ib-cr | WT | S80I | S458A |
| 16029 | Escherichia coli | urine | ST692 | blaCMY-59, blaCTX-M-15, blaNDM-7, blaTEM-1, ampH, ampC | aac(6′)Ib-cr | S83L, D87N | S80I | S458A |
| 16032 | Escherichia coli | urine | ST692 | blaCTX-M-15, blaNDM-7, blaOXA-1, blaCMY-59, blaTEM-1, ampH, ampC | aac(6′)Ib-cr | S83L, D87N | S80I | S458A |
| 16047 | Klebsiella pneumoniae | urine | ST147 | blaCTX-M-15, blaOXA-1, blaSHV-187, K. pneumoniae OmpK37, E. coli ampH | aac(6′)Ib-cr oqxA, oqxB qnrA1, qnrB1 | S83Y, D87A | WT | WT |
| 15958 | Pseudomonas aeruginosa | sputum | ST2935 | blaOXA-50, blaPDC-10 | None | S912del, E913del | F254V, S331T, A346Q | ND |
| 15964 | Pseudomonas aeruginosa | wound | ST1203 | blaGES-9, blaOXA-21, blaOXA-50, blaPDC-1, blaVIM-5 | aac(6′)Ib-cr, qnrVC1 | T83I, S912del, E913del | S87L, F254V, A346Q | ND |
| 15965 | Pseudomonas aeruginosa | urine | ST773 | blaNDM-1, blaOXA-50, blaPDC-1 | qnrVC1 | T83I | S87L, F254V, A346Q | ND |
| 15966 | Pseudomonas aeruginosa | urine | ST1203 | blaGES-9, blaOXA-21-like, blaOXA-50, blaPDC-1, blaVIM-5 | aac(6′)Ib-cr, qnrVC1 | T83I, S912del, E913del | S87L, F254V, A346Q | ND |
| 15986 | Pseudomonas aeruginosa | wound | ST654 | blaOXA-10, blaOXA-50, blaPDC-3, blaVIM-5 | aac(6′)Ib-cr, qnrVC1 | T83I, S912del, E913del | S87L, F254V, A346Q | ND |
| 16014 | Pseudomonas aeruginosa | stool | ST244 | blaOXA-486, blaPDC-1 | None | WT | F254V, A346Q | ND |
| 16018 | Pseudomonas aeruginosa | urine | ST654 | blaOXA-10, blaOXA-50, blaPDC-3, blaTEM-1, blaVIM-5, blaSCO-1 | aac(6′)Ib-cr, qnrVC1 | T83I, S912del, E913del | S87L, F254V, A346Q | WT |
| 16048 | Pseudomonas aeruginosa | sputum | ST1555 | blaOXA-50, blaPDC-10 | None | S912del, E913del | F254V, S331T, A346Q | ND |
| ID | Organism | Source | MLST | Beta-Lactamase Genes | Fluoroquinolone PMQR Genes | Fluoroquinolone QRDR Mutations | ||
|---|---|---|---|---|---|---|---|---|
| gyrA | parC | parE | ||||||
| 17757 | Escherichia coli | urine | Ambiguous (ST506, ST566) | blaCTX-M-14, blaTEM-1B | None | S83L, D87G | S80I | S458A, I529L |
| 17758 | Escherichia coli | VS | Ambiguous (ST27, ST129) | blaTEM-1B | None | S83L | WT | WT |
| 17762 | Escherichia coli | urine | Ambiguous (ST27, ST129) | blaTEM-1B | None | S83L | WT | WT |
| 17771 | Escherichia coli | urine | ST2 | blaTEM-1B | None | WT | WT | WT |
| 17772 | Escherichia coli | urine | ST2 | blaCTX-M-15, blaOXA-1, blaTEM-1B | aac(6′)-Ib-cr | S83L, D87N | S80I | S458A |
| 17773 | Escherichia coli | VS | ST83 | None | qnrB7 | WT | WT | WT |
| 17775 | Escherichia coli | urine | Inconclusive (ST721, ST662, ST472) | blaCTX-M-15, blaOXA-1, blaTEM-1B | aac(6′)-Ib-cr | S83L, D87N | S80I | S458A |
| 17776 | Escherichia coli | urine | Inconclusive (ST466, ST210, ST132) | blaTEM-1B | None | S83L, D87N | S80I | S458A |
| 17781 | Escherichia coli | urine | ST692 | blaCMY-2, blaCTX-M-15, blaOXA-181, blaTEM-1B | qnrS1 | S83L, D87N | S80I | S458A |
| 17782 | Escherichia coli | urine | ST471 | blaCTX-M-15, blaOXA-1 | None | S83L, D87N | S80I | S458A |
| 17786 | Escherichia coli | wound | ST132 | blaTEM-1B | None | S83L, D87N | S80I | S458A |
| 17789 | Escherichia coli | urine | Inconclusive (ST500, ST437) | blaTEM-1B | qnrS1 | WT | WT | WT |
| 17792 | Escherichia coli | wound | ST2632 | blaCTX-M-15, blaOXA-1, blaTEM-1B | aac(6′)-Ib-cr | S83L, D87N | S80I | S458A |
| 17795 | Escherichia coli | urine | ST86 | blaCTX-M-15, blaTEM-1B | qnrS1 | S83L, D87N | S80I | |
| 17796 | Escherichia coli | urine | Ambiguous (ST566, ST506) | blaCTX-M-14, blaTEM-1B | None | S83L, D87G | S80I | S458A, I529L |
| 17803 | Escherichia coli | wound | ST2 | blaCTX-M-15, blaOXA-1, blaSHV-187, blaTEM-1B | aac(6′)-Ib-cr, oqxA, oqxB | S83L, D87N | S80I | S458A |
| 17804 | Escherichia coli | wound | ST471 | blaCTX-M-15 | aac(6′)-Ib-cr | WT | WT | WT |
| 17862 | Escherichia coli | VS | Inconclusive (ST500, ST437) | blaTEM-1B | qnrS1 | WT | WT | WT |
| 17864 | Escherichia coli | urine | ST58 | blaCTX-M-15, blaOXA-1, blaOXA-320/534, blaTEM-1B | None | S83L, D87N | S80I | S458T |
| 17759 | Klebsiella pneumoniae | urine | ST340 | blaCTX-M-27, blaNDM-7, blaSHV-187 | aac(6′)-Ib-cr, oqxA, oqxB, qnrB6 | WT | WT | WT |
| 17765 | Klebsiella pneumoniae | sputum | ST86 | blaOXA-1, blaSHV-187 | aac(6′)-Ib-cr, oqxA, oqxB | WT | WT | WT |
| 17766 | Klebsiella pneumoniae | sputum | ST20 | blaSHV-187 | oqxA, oqxB | WT | WT | WT |
| 17767 | Klebsiella pneumoniae | US | ST392 | blaSHV-11 | aac(6′)-Ib-cr, oqxA, oqxB qnrB6 | WT | WT | WT |
| 17768 | Klebsiella pneumoniae | urine | ST340 | blaCTX-M-27, blaNDM-7, blaSHV-187 | aac(6′)-Ib-cr, oqxA, oqxB, qnrB6 | WT | WT | WT |
| 17770 | Klebsiella pneumoniae | BC | ST86 | blaSHV-187 | oqxA, oqxB | WT | WT | WT |
| 17785 | Klebsiella pneumoniae | wound | ST45 | blaCTX-M-15, blaSHV-187, blaTEM-1B | oqxA, oqxB | WT | WT | WT |
| 17787 | Klebsiella pneumoniae | sputum | ST2632 | blaSHV-93 | oqxA, oqxB | WT | WT | WT |
| 17790 | Klebsiella pneumoniae | wound | ST86 | blaSHV-187 | oqxA, oqxB | WT | WT | WT |
| 17793 | Klebsiella pneumoniae | sputum | ST661 | blaSHV-187 | oqxA, oqxB | WT | WT | WT |
| 17861 | Klebsiella pneumoniae | urine | ST86 | blaSHV-187 | oqxA, oqxB | WT | WT | WT |
| 17863 | Klebsiella pneumoniae | ES | ST340 | blaCTX-M-27, blaNDM-7, blaSHV-187 | aac(6′)-Ib-cr, oqxA, oqxB, qnrB18, qnrB6 | WT | WT | WT |
| 17760 | Pseudomonas aeruginosa | sputum | ST274 | blaOXA-486, blaPDC-24 | None | Silent | F254V, A346Q | ND |
| 17761 | Pseudomonas aeruginosa | ear swab | Inconclusive (nearest ST244) | blaOXA-847, blaPDC-423 | None | Silent | F254V, A346Q | ND |
| 17763 | Pseudomonas aeruginosa | urine | ST773 | blaNDM-1, blaOXA-395, blaPDC-385 | qnrVC1 | T83I | S87L, F254V, A346Q | ND |
| 17777 | Pseudomonas aeruginosa | urine | ST773 | blaNDM-1, blaOXA-395, blaPDC-385 | qnrVC1 | T83I | S87L, F254V, A346Q | ND |
| 17778 | Pseudomonas aeruginosa | ear swab | ST773 | blaNDM-1, blaOXA-395, blaPDC-385 | qnrVC1 | T83I | S87L, F254V, A346Q | ND |
| 17779 | Pseudomonas aeruginosa | urine | ST773 | blaNDM-1, blaOXA-395, blaPDC-385 | qnrVC1 | T83I | S87L, F254V, A346Q | ND |
| 17791 | Pseudomonas aeruginosa | eye swab | ST773 | blaNDM-1, blaOXA-395, blaPDC-385 | qnrVC1 | T83I | S87L, F254V, A346Q | ND |
| 17794 | Pseudomonas aeruginosa | wound | ST773 | blaNDM-1, blaOXA-395, blaPDC-385 | qnrVC1 | T83I | S87L, F254V, A346Q | ND |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kawa, D.E.; Tickler, I.A.; Tenover, F.C.; Shettima, S.A. Characterization of Beta-Lactamase and Fluoroquinolone Resistance Determinants in Escherichia coli, Klebsiella pneumoniae, and Pseudomonas aeruginosa Isolates from a Tertiary Hospital in Yola, Nigeria. Trop. Med. Infect. Dis. 2023, 8, 500. https://doi.org/10.3390/tropicalmed8110500
Kawa DE, Tickler IA, Tenover FC, Shettima SA. Characterization of Beta-Lactamase and Fluoroquinolone Resistance Determinants in Escherichia coli, Klebsiella pneumoniae, and Pseudomonas aeruginosa Isolates from a Tertiary Hospital in Yola, Nigeria. Tropical Medicine and Infectious Disease. 2023; 8(11):500. https://doi.org/10.3390/tropicalmed8110500
Chicago/Turabian StyleKawa, Diane E., Isabella A. Tickler, Fred C. Tenover, and Shuwaram A. Shettima. 2023. "Characterization of Beta-Lactamase and Fluoroquinolone Resistance Determinants in Escherichia coli, Klebsiella pneumoniae, and Pseudomonas aeruginosa Isolates from a Tertiary Hospital in Yola, Nigeria" Tropical Medicine and Infectious Disease 8, no. 11: 500. https://doi.org/10.3390/tropicalmed8110500
APA StyleKawa, D. E., Tickler, I. A., Tenover, F. C., & Shettima, S. A. (2023). Characterization of Beta-Lactamase and Fluoroquinolone Resistance Determinants in Escherichia coli, Klebsiella pneumoniae, and Pseudomonas aeruginosa Isolates from a Tertiary Hospital in Yola, Nigeria. Tropical Medicine and Infectious Disease, 8(11), 500. https://doi.org/10.3390/tropicalmed8110500