Abstract
Polymorphisms of the genes encoding proteins involved in immune functions and the binding of malaria parasites to human host cells have been the focus of research in recent years, aiming to understand malaria pathogenesis and case severity and to exploit this knowledge to assert control over malaria. This study investigated the genetic diversity of the human host genes encoding proteins that are involved in immune functions and malaria parasite binding, i.e., MCP1 (−2518), TGFβ1 (−509), TNFα (−308), IL4 (VNTR), IL6 (−174), IL10 (−3575), TLR4 (299), CD36 (−188), and ICAM1 (469) in patients with mono-infection of Plasmodium falciparum and Plasmodium vivax infections in the multidrug-resistant areas along the Thai-Myanmar border. The association between gene polymorphisms and parasite density was also investigated. Genomic DNA (gDNA) of P. falciparum and P. vivax were extracted from whole blood and dried blood spot (DBS). Gene amplification and genotyping were performed by PCR and PCR-RFLP analysis, respectively. Of these samples, 178 and 209 samples were, respectively, mono-infection with P. falciparum and P. vivax. The ratio of P. falciparum: P. vivax was 46%:54%. Results showed marked variation in the frequency distribution and patterns of the genotypes and gene alleles of the nine immune response genes or human host genes. The SNPs of TGFβ1, IL10 and ICAM1, were significantly associated with P. falciparum, but not P. vivax parasite density. TGFβ1, IL10 and ICAM1, may play more significant roles in modulating P. falciparum than P. vivax parasitemia. The prevalence of the genotypes and gene alleles of these genes, including their association with parasite density, may vary depending on patient ethnicity and endemic areas. Information obtained from each endemic area is essential for treatment strategies and the development of vaccines for malaria prophylaxis in specific areas.
Keywords:
Plasmodium falciparum; Plasmodium vivax; MCP1; TGFβ1; TNFα; IL4 (VNTR); IL6; IL10; TLR4; CD36; ICAM1 1. Introduction
Malaria remains one of the major global public health problems despite the declining incidence in recent years. In 2018, 3.2 billion people were at risk of malaria infection, with an estimated 219 million cases and 435,000 malaria-related deaths [1]. Over 90% of death cases were reported from sub-Saharan Africa, most of which were children under five years of age. Five species of malaria parasite infect humans, i.e., Plasmodium falciparum, P. vivax, P. malariae, P. ovale, and P. knowlesi. P. falciparum is the most virulent and widespread infectious species in tropical and subtropical countries due to the parasite’s resistance to the most available antimalarial drugs. The species P. vivax is the next most significant malaria species, which usually causes benign uncomplicated malaria with relapse from dormant hypnozoites in the liver. Its clinical features differ from those of P. falciparum malaria. Both P. falciparum and P. vivax often co-exist in several parts of the world. The signs and symptoms of malaria vary from uncomplicated to complicated or severe malaria. Severe malaria manifests life-threatening clinical signs and symptoms, which are defined according to criteria of the World Health Organization (WHO) as impaired consciousness, respiratory distress, multiple convulsions, circulatory collapse, abnormal bleeding, jaundice, hemoglobinuria, severe anemia, hypoglycemia, acidemia, hyperlactatemia, renal impairment, and hyperparasitemia (>250,000 parasites/µL) [2]. Cerebral malaria is one of the most significant forms of severe malaria involving the two major pathogenesis processes, i.e., cytoadherence and immunogenic reactions. Most cases of cerebral malaria are due to P. falciparum infection, whereas P. vivax infection rarely causes cerebral malaria [3].
Both parasite and human host-related factors play significant roles in malaria susceptibility, pathogenesis, and disease severity [4]. Among the host-related factors, polymorphisms in the genes encoding proteins and involved in either immune functions (MCP1, TGFβ1, TNFα, IL4 VNTR, IL6, and IL10), or parasite binding to human host cells (TLR4, ICAM1, and CD36) have been the focus of research in recent years as researchers try to understand malaria pathogenesis and severity and to exploit the knowledge for malaria control. Results of various reports for the relationship between the host genetic polymorphisms and malaria susceptibility, pathogenesis and disease severity, however, are conflicting, depending on endemic areas and observation periods under investigation. The present study investigated the genetic diversity of the human host genes involved in immune response [monocyte chemoattractant protein 1 (MCP1)-2518, transforming growth factor β1 (TGFβ1)-509, tumor necrosis factor α (TNFα)-308, interleukin 4 (IL4) variable number tandem repeat (VNTR), interleukin 6 (IL6)-174, and interleukin 10 (IL10)-3575], as well as those involved in malaria parasite binding [toll-like receptor 4 (TLR4)-299, cluster of differentiation 36 (CD36)-188, and intercellular adhesion molecule 1 (ICAM1)-469] in patients with mono-infection of P. falciparum or P. vivax, residing in the multidrug-resistant areas along the Thai-Myanmar border [5]. The polymorphisms of the MCP1, TGFβ1, TNFα, IL4 VNTR, IL6, IL10 and CD36 genes involve nucleotide positions located in the promotor region, and the numbers indicate nucleotide positions. The polymorphisms of the TLR4 and ICAM1 genes involve amino acid changes, and the numbers indicate amino acid positions. The association between the polymorphisms (single nucleotide polymorphisms: SNPs) of these genes at specific amino acid/nucleotide positions and parasite density, as a criterion of malaria severity, was also investigated.
2. Materials and Methods
2.1. Samples and Study Site
The study was conducted in 2007 and 2015 at malaria clinics within the two malaria-endemic areas located along the Thai-Myanmar border, i.e., Mae-Sot (Tak province) and Sai Yok (Kanchanaburi province) districts. Approval for the study protocol was obtained from the Ethics Committee for Research in Human Subjects, Ministry of Public Health of Thailand (Number 3/52-293) and the Biosafety Committee of Thammasat University, Thailand (Number 082/2560). The minimum sample size of 385 patients was estimated based on the prevalence of ICAM-1 polymorphism, which was the largest sample size among other gene polymorphisms [6].
Whole blood and dried blood spot (DBS) samples were collected from patients with confirmed infections with P. falciparum or P. vivax by microscopic examination of Giemsa’s thin blood smears and nested PCR [7]. The parasite density of each sample was determined according to WHO guidelines [8]. Parasite density (per µL blood) in all samples was determined from thick blood smears under a light microscope at 1000× magnification. The number of asexual stage parasites, i.e., rings, trophozoites, and schizonts were counted (three times) against 200 white blood cells (WBC) in the thick blood film. Parasite density (per microliter) was calculated as the number of parasites multiplied by the number of WBCs and divided by 200. Whole blood samples were stored at −20 °C and transferred together with DBS samples to the Center of Excellence in Pharmacology and Molecular Biology of Malaria and Cholangiocarcinoma, Thammasat University, Thailand.
2.2. Preparation of Genomic DNA
Genomic DNA (gDNA) of whole blood and DBS samples infected with P. falciparum or P. vivax (mono-infection) were extracted using QIAamp DNA Blood Mini kit (QIAGEN, Hilden, Germany).
2.3. Gene Amplification by Polymerase Chain Reaction (PCR)
The PCR conditions (total volume of 25 µL) were set as follows: 5 µL of sample gDNA, 1.5 µL of MgCl2, 1.5 µL of 10 × KCl buffer, 0.5 µL of dNTP, 0.25 µL of Taq polymerase enzyme, and 0.5 µL each of forward and reverse primers. The primers used in the reaction are presented in Table 1. Gene amplification, denaturation, and extension steps (35 cycles) were set as 95 °C for 1 min and 72 °C for 1 min. For the annealing step, the temperature gradient specific to each gene was set at 40–60 °C. The amplified gene product was separated by 1.5% agarose gel electrophoresis.

Table 1.
The primers and analysis conditions of nine host genes involved in immune response and malaria parasite binding (amino acid/nucleotide positions with mutation and genetic analysis) under investigations.
2.4. Identification of SNPs and Genotypes by Restriction Fragment Length Polymorphism (RFLP)
Detailed information of the polymorphic points, amino acid changes, genotypes, and analysis conditions of all nine genes investigated are summarized in Table 1. The genetic polymorphism of TLR4 results from an amino acid change from aspartate to glycine at the nucleotide position 896, while that of ICAM-1 results from an amino acid change from lysine to glutamine at the nucleotide position 469. The reaction mixture (20 µL) consisted of 10 µL of PCR amplicon, 2.0 µL of the specific buffer, 7.9 µL of distilled water, and 1 U of specific restriction enzyme of individual SNP. The mixture was incubated at 37 °C and the RFLP products were analyzed by electrophoresis on 3% agarose gel.
2.5. Statistical Analysis
The frequency of genotypes and alleles of each gene are presented as number and percentage (%) values. Quantitative data are summarized as median (range) values. The deviation of gene distribution was determined by Hardy–Weinberg Equilibrium (HWE) (https://dr-petrek.edu>documents>HWE.xls, accessed on 10 January 2021). The distribution patterns of gene types, genotypes, and gene alleles were analyzed using the Chi-square test. The association between genotypes and gene alleles and P. falciparum and P. vivax parasite density was determined using Kruskal–Wallis and Mann–Whitney U tests. The statistical significance level was set at α = 0.05.
3. Results
A total of 387 samples (71 whole blood and 316 DBS samples) were included in the analysis. This number comprised 172 and 215 samples obtained from Tak, and Kanchanaburi province, respectively. Most 96.6% of patients were Burmese, and 3.4% were ethnic minorities. Totals of 178 (46.0%) and 209 (54.0%) samples were mono-infection with either P. falciparum or P. vivax, respectively. Successful amplification of the nine genes under investigation varied between 24.2–100.0% and 61.2–100.0% for P. falciparum and P. vivax-infected samples, respectively. Representative images of the PCR-RFLP products of the nine polymorphic genes under investigation are shown in Figure 1. The distribution of P. falciparum- and P. vivax-infected cases, including the parasite density in the two endemic areas, are summarized in Table 2.
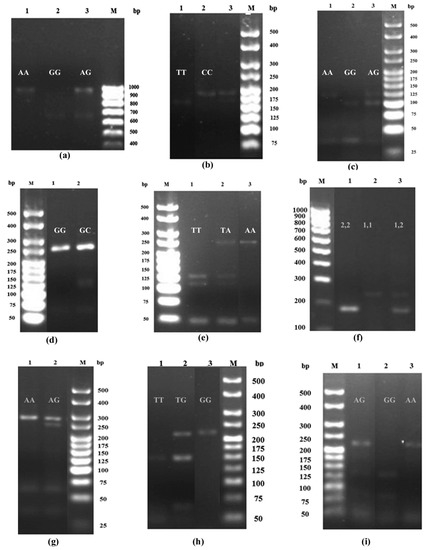
Figure 1.
The PCR-RFLP products for (a) MCP1 polymorphism at position −2518: AA (940 bp), GG (650 bp) and GA (940 and 650 bp) genotypes, (b) TGFβ1 polymorphism at position −509: CC (178 bp), TT (159 bp) and TC (178 and 159 bp) genotypes, (c) TNFα polymorphism at position −308: AA (117 bp), GA (117 and 97) and GG (97 bp) genotypes, (d) IL6 polymorphism at position −174: GG (233 bp) and GC (233 and 122 bp) genotypes, (e) IL10 polymorphism at position −3575: AA (228 bp), TA (228, 121 and 107 bp) and TT (121 and 107 bp) genotypes, (f) IL4 VNTR: 1,1 (253 bp), 2,2 (183 bp) and 1,2 (253 and 183 bp) genotypes, (g) TLR4 polymorphism at position 299: AA (249) and AG (249 and 223 bp) genotypes, (h) CD36 polymorphism at position −188: GG (213 bp), TG (213, 148 bp) and TT (148) genotypes, and (i) ICAM1 polymorphism at position 469: AA (223 bp), GG (136), and AG (223, 136) genotypes.
Table 2.
Demographic data of samples collected for genetic analysis. Data are presented as number (%) or median (range) value.
3.1. Genotype and Allele Distribution
The distribution patterns of genotypes and gene alleles of the nine genes in patients infected with P. falciparum and P. vivax are summarized in Table 3, respectively. The predominant genotypes of most genes in patients with P. falciparum infection were homozygous wild type/mutant genotypes, with the exception of MCP1 and CD36, of which the predominant genotypes were heterozygous. The dominant alleles of most genes were wild type alleles (with a frequency of up to 100.0%), with the exception of IL4VNTR, of which the predominant allele was two gene repeats. The genotypes and gene alleles of most genes showed a significant difference in distribution patterns. For P. vivax, similarly to P. falciparum isolates, the predominant genotypes for most genes were homozygous wild type, with the exception of MCP1 and TNFα1, of which the predominant genotypes were heterozygous. The dominant alleles of most genes were wild type alleles (with a frequency of up to 99.0%), with the exception of IL4VNTR, of which the predominant allele was two gene copies. The genotypes but not gene alleles of patients infected with P. vivax showed a significant difference in distribution patterns. The distribution patterns of genotypes of the samples obtained from the malaria-infected patients significantly deviated from Hardy–Weinberg Equilibrium (HWE), i.e., IL10 (p = 0.006), ICAM1 (p = 0.015), TGFβ1 (p = 0.027), and CD36 (p < 0.001) in P. falciparum-infected patients and TNFα1 (p < 0.001), IL10 (p < 0.001), and ICAM1 (p = 0.003) in P. vivax-infected patients.
Table 3.
Distribution of genotypes and gene alleles of host genes related to malaria severity. Data are presented as number (%) or median (range) value.
3.2. Association between Genotypes/Gene Alleles and Parasite Density
The association between the genotypes of the nine genes in patients with P. falciparum and P. vivax infections are summarized in Figure 2a,b. Significant associations were observed for TGFβ1, IL10, and ICAM1 genotypes in patients with P. falciparum infection. The parasite density in patients with the heterozygous genotype for TGFβ1 was significantly higher than those with the wild type genotypes [median (range): 4568 (118–22,750) vs. 706 (40–258,535) /μL)]. Conversely, the parasite density in the isolates with the heterozygous or homozygous mutant genotype for IL10 was significantly lower than the wild type genotype [median (range): 588 (40–258,535) vs. 694 (40–109,874) vs. 4,970 (236–48,475) /μL)]. The parasite density in patients carrying the heterozygous genotype of ICAM1 was significantly lower than the wild type genotype [median (range): 620 (40–109,874) vs. 3780 (79–258,535) /μL)]. No association between genotypes and P. vivax parasite density was found.
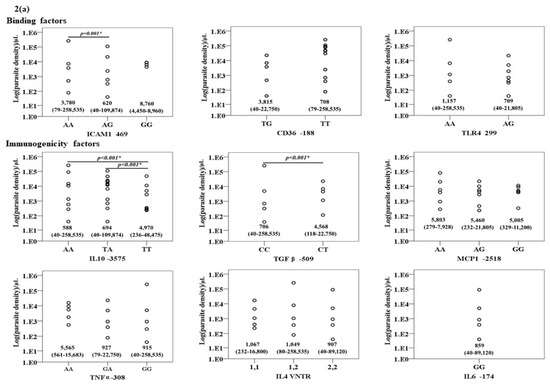
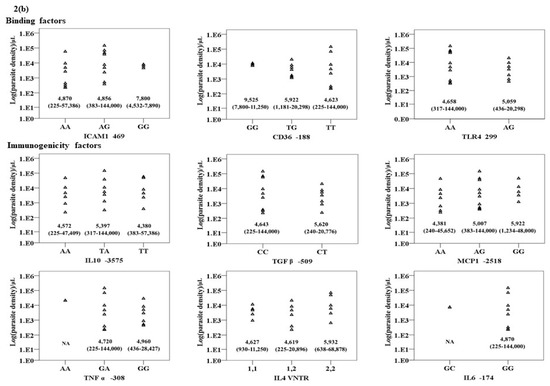
Figure 2.
The relationship between the genotypes of the nine genes under investigation and Log (parasite density) (/μL) in (a) P. falciparum and (b) P. vivax infections.
For the gene alleles, similarly to those observed with genotypes, significant associations were observed between the gene alleles of TGFβ1, IL10 and ICAM1, and parasite density in patients with P. falciparum infection. The parasite density in the isolates carrying the mutant allele of TGFβ1 was significantly higher than those with the wild type allele [median (range): 4568 (118–22,750) vs. 1109 (40–258,535) /µL)]. Conversely, the parasite density in patients carrying heterozygous alleles of IL10 and ICAM1 was significantly lower than those with the wild type allele [median (range): 623 (40–258,535) vs. 2300 (40–109,874) /µL and 627 (40–109,874) vs. 1109 (40–258,535) vs. /µL, respectively]. No association between genotypes and P. vivax parasite density was found.
4. Discussion
The frequencies and distribution patterns of genotypes and gene alleles of most genes under investigation were markedly different, depending on the species of the malaria parasite. It is noteworthy that results may suggest a more significant contribution of these genes in patients with P. falciparum compared with P. vivax infection for malaria parasite density, one of the criteria for malaria severity. The HWE deviation observed with TGFβ1, TNFα, IL10, CD36 and ICAM1 could be due to gene mutation, population migration, and/or selection of genotypes/alleles of the study populations. In the present study, the SNPs of the two host genes involved in immune functions, i.e., TGFβ1 and IL10, and the one involved in malaria parasite binding (ICAM1) were significantly associated with P. falciparum parasitemia. The SNPs of TGFβ1 (CT genotype) was significantly associated with high parasite density, whereas that of IL10 (A allele and TA and AA genotypes) and ICAM1 (G allele and AG genotype) were significantly associated with low parasite density compared with their corresponding wild type alleles/genotypes. However, this association was not observed in patients with P. vivax infection. This may suggest the significant contribution of these genes in host susceptibility to infection and disease severity in P. falciparum cases but not P. vivax.
The CC genotype of TGFβ has been reported to link with low parasite density in P. falciparum, while the CT genotype is linked with high parasite density. It is possible that the homozygous genotype (CC) results in the decrease in TGFβ cytokine production for malaria eradication [20], whereas the heterozygous genotype (CT) results in the increase in TGFβ cytokine production and high parasite density, thereby, perhaps, enhancing the risk of severe malaria [21]. The T allele may play a more significant contribution to treatment outcomes than the C allele, and anchoring T allele may suppress the function of C allele for TGFβ level production [22,23,24] through enhancing promoter transcription at the Yin-Yang1 binding site [25]. Nevertheless, some studies reported the association between low TGFβ1 level and severe clinical outcomes in both P. falciparum and P. vivax infections [26,27,28].
For IL10 gene, the A allele and TA and AA genotypes were associated with low parasite density in patients with P. falciparum infection compared with the wild type genotype/allele. It is hypothesized that the T allele and TT genotype may increase the production of IL10 and severe malaria [29,30]. This, in turn, suppresses the production of IL10 by IL10 producing cells [31,32], which alleviates the action of this cytokine on reducing malaria parasitemia, and thus the risk of hyperparasitemia. An opposite effect may occur with the A allele and AA and TA genotypes. Our results support the results from a previous study showing a more significant association between IL10 -3575A and low plasma IL10 level compared with the -3575 T allele [33]. In addition, a significant association was found between IL10 -1082A and decreased IL10 production in severe malaria [34]. It is likely that these allele/genotypes result in the reduction of IL10 production [16,32], thereby triggering the production of this cytokine by the target cells [35]. For P. vivax infection, in spite of the high plasma levels of IL10 and TGFβ during the infection phase [36], minimum parasite density over the threshold level is required for pathogenesis induction [37]. Plasmodium vivax has a hypnozoite stage, mono-invasion and long incubation period of the asexual stage. These factors result in low parasitemia and pathogenicity [38]. Since P. vivax infection produces non-severe symptoms, patients usually do not seek treatment with antimalarial drugs. This may lead to chronic morbidity and high plasma cytokine levels. The association between human genetics and plasma levels of these cytokines remains conflicting. The result of the current study in the Thai-Myanmar area, however, showed no association between the genetic polymorphisms of the human immune cytokines and P. vivax parasite density.
The adhesion of infected red blood cells to the microvasculature of the brain is a key feature of cerebral malaria [39,40]. The host binding molecule ICAM1 or CD54 is located on the surface of endothelial cells. It acts as a binding receptor of the parasite virulence protein PfEMP1 (Plasmodium falciparum erythrocyte membrane protein 1), leading to an enhanced risk of severe clinical outcomes [40]. In the present study, the G allele and AG genotype were associated with low P. falciparum density. The mutation of the ICAM1 gene at this position may result in an impaired binding property of the ICAM1 molecule on the infected erythrocytes, which was supported by the results of an in vivo study [41]. This leads to a low risk of parasite cytoadherence, thereby, decreasing the severity of malaria pathogenesis [42,43]. However, some studies reported conflicting results [44]. For P. vivax, the parasite exports vir protein encoded by vir gene (variant interspersed repeats gene), which binds to ICAM1 receptor molecule. Unlike PfEMP1, vir protein is not a potent cytoadherence and sequestering ligand [45,46]; therefore, severe disease manifestation is unlikely. Our result supports no association between the polymorphisms of these parasite binding genes and susceptibility to high P. vivax parasitemia in the Thai-Myanmar border area.
There was no significant association between P. falciparum or P. vivax parasite density and SNPs of the MCP1, TNFα, IL4, IL6, IL10, TLR4, or CD36 genes. Current reports on the association between malaria parasite density and severity and the SNPs of these genes are controversial. High MCP1 levels during P. falciparum and P. vivax infection were associated with severe malaria and mortality [47,48,49,50,51,52]. The IL4 VNTR 1R allele was associated with severe malaria with increased plasma IL4 level [53] and reduced risk of severe malaria, while the IL4 VNTR 2R allele was associated with decreased plasma IL4 level [16]. The SNP of IL6 (-174GC) was associated with hyperparasitemia, malaria severity and IL6 level [14,54]. The TLR4 SNP (299AG) was associated with severe anemia and hyperparasitemia in Ghanaian children [55,56]. A heterozygous genotype (TG) of CD36 was protective against severe malaria in India [57,58].
It was noted that hyperparasitemia is only one criterion for malaria severity. Malaria symptoms and disease severity can be influenced by many factors such as parasite mixed strain infection, endemic malaria area, host genetic background, time of infection (the duration from infection and sample collection), medication history, nutrition, prior infections, and co-infection with other pathogens (viruses and bacteria). These factors could be confounding factors for conflicting results reported in this as well as other studies in other malaria-endemic areas. This should be considered when analyzing and interpreting the results. An additional limitation of the current study is that the number of samples used in each association analysis is small, particularly for the TNFα and MCP1 genes. Information obtained from each endemic area, therefore, is essential for treatment strategies and the development of vaccines for malaria prophylaxis in those specific areas. Furthermore, other potential genes were identified in the large-scale genome-wide association studies, e.g., gene expression profiles related to neutrophil and erythroid activity, sickle cell hemoglobin S variant, interleukin receptors and kelch-like protein KLHL3 [59,60,61], should be included in the analysis.
5. Conclusions
The genes involved in host immune response and binding of the malaria parasite, particularly TGFβ1, IL10 and ICAM1 may play a more significant role in malaria parasitemia in P. falciparum than in P. vivax infection. The SNPs of these genes were associated with P. falciparum parasite density. The prevalence of the genotypes and alleles of these genes reported in various studies, including their association with malaria parasite density and disease severity, is conflicting due to confounding factors. Interpretation and conclusion on the association of the polymorphisms of genes involved in host immune response and binding of the malaria parasite and P. falciparum and P. vivax parasitemia should be based on studies conducted in each endemic area with adequate sample size and minimized confounding factors.
Author Contributions
Conceptualization, K.N.-B., W.C.; methodology, W.C., K.S.; validation, W.C., K.S.; sample collection, K.N.-B., W.C.; data analysis, W.C., K.N.-B.; draft preparation, K.S.; manuscript finalization, K.N.-B., funding acquisition, K.N.-B. All authors have read and agreed to the published version of the manuscript.
Funding
The study was supported by Thammasat University (Center of Excellence in Pharmacology and Molecular Biology of Malaria and Cholangiocarcinoma) and the National Research Council of Thailand. Kesara Na-Bangchang is supported by the National Research Council of Thailand under the Research Team Promotion grant (grant number NRCT 820/2563).
Institutional Review Board Statement
Approval of the study protocol was obtained from the Ethics Committee for Research in Human Subjects, Ministry of Public Health of Thailand (Number 3/52-293).
Informed Consent Statement
All research participants gave written informed consent for study participation.
Data Availability Statement
Data will be made available upon request.
Acknowledgments
We are grateful to the staff of the malaria clinics in Mae-Sot (Tak province) and Sai Yok (Kanchanaburi province) for their kind assistance during data collection. We thank Ethan Vindvamara for English editing of the manuscript.
Conflicts of Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
- World Health Organization. World Malaria Report 2018; World Health Organisation: Geneva, Switzerland, 2018. [Google Scholar]
- World Health Organization. Severe malaria. Trop. Med. Int. Health 2014, 19 (Suppl. 1), 7–131. [Google Scholar] [CrossRef]
- Mathews, S.E.; Bhagwati, M.M.; Agnihotri, V. Clinical spectrum of Plasmodium vivax infection, from benign to severe malaria: A tertiary care prospective study in adults from Delhi, India. Trop. Parasitol. 2019, 9, 88–92. [Google Scholar] [CrossRef] [PubMed]
- Mackinnon, M.J.; Mwangi, T.W.; Snow, R.W.; Marsh, K.; Williams, T.N. Heritability of malaria in Africa. PLoS Med. 2005, 2, e340. [Google Scholar] [CrossRef]
- Na-Bangchang, K.; Muhamad, P.; Ruaengweerayut, R.; Chaijaroenkul, W.; Karbwang, J. Identification of resistance of Plasmodium falciparum to artesunate-mefloquine combination in an area along the Thai-Myanmar border: Integration of clinico-parasitological response, systemic drug exposure, and in vitro parasite sensitivity. Malar. J. 2013, 12, 263. [Google Scholar] [CrossRef]
- Mishra, P.; Prasad, K.N.; Singh, K.; Sahu, R.N.; Ojha, B.K. Association of ICAM-1 (K469E) and MCP-1-2518 A > G gene polymorphism with brain abscess. J. Neuroimmunol. 2016, 292, 102–107. [Google Scholar] [CrossRef]
- Snounou, G.; Singh, B. Nested PCR analysis of Plasmodium parasites. Methods Mol. Med. 2002, 72, 189–203. [Google Scholar] [PubMed]
- World Health Organization. Basic Malaria Microscopy: Tutor’s Guide, 2nd ed.; WHO Press: Geneva, Switzerland, 2010; pp. 25–27. [Google Scholar]
- Vechvongvan, V.; Suthamdireklap, C.; Prathumthong, A.; Limtrakul, A.; Pumeechockchai, W.; Tangteerawatana, P. Distribution of TGF-β1 C-509T polymorphism in Thai population. J. Med. Health Sci. 2017, 24, 28–36. [Google Scholar]
- Ogawa, E.; Ruan, J.; Connett, J.E.; Anthonisen, N.R.; Paré, P.D.; Sandford, A.J. Transforming growth factor-beta1 polymorphisms, airway responsiveness and lung function decline in smokers. Respir. Med. 2007, 101, 938–943. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Mombo, L.E.; Ntoumi, F.; Bisseye, C.; Ossari, S.; Lu, C.Y.; Nagel, R.L.; Krishnamoorthy, R. Human genetic polymorphisms and asymptomatic Plasmodium falciparum malaria in Gabonese schoolchildren. Am. J. Trop. Med. Hyg. 2003, 68, 186–190. [Google Scholar] [CrossRef]
- Cabantous, S.; Poudiougou, B.; Oumar, A.A.; Traore, A.; Barry, A.; Vitte, J.; Bongrand, P.; Marquet, S.; Doumbo, O.; Dessein, A.J. Genetic Evidence for the Aggravation ofPlasmodium falciparumMalaria by Interleukin 4. J. Infect. Dis. 2009, 200, 1530–1539. [Google Scholar] [CrossRef]
- Mout, R.; Willemze, R.; Landegent, J.E. Repeat polymorphisms in the interleukin-4 gene (IL4). Nucleic Acids Res. 1991, 19, 3763. [Google Scholar] [CrossRef]
- Mendonça, V.R.R.; Souza, L.C.L.; Garcia, G.C.; Magalhães, B.M.L.; Lacerda, M.V.G.; Andrade, B.B.; Gonçalves, M.S.; Barral-Netto, M. DDX39B (BAT1), TNF and IL6 gene polymorphisms and association with clinical outcomes of patients with Plasmodium vivax malaria. Malar. J. 2014, 13, 278. [Google Scholar] [CrossRef]
- Yalçın, S.; Kayaaltı, Z.; Söylemezoğlu, T. Role of interleukin-6-174 G/C promoter polymorphism in trace metal levels of autopsy kidney and liver tissues. Int. J. Hyg. Environ. Health 2011, 214, 219–224. [Google Scholar] [CrossRef] [PubMed]
- Domingues, W.; Kanunfre, K.A.; Rodrigues, J.C.; Teixeira, L.E.; Yamamoto, L.; Okay, T.S. Preliminary report on the putative association of IL10 -3575 T/A genetic polymorphism with malaria symptoms. Rev. Inst. Med. Trop. Sao. Paulo. 2016, 58, 30. [Google Scholar] [CrossRef]
- Iwalokun, B.A.; Oluwadun, A.; Iwalokun, S.O.; Agomo, P. Toll-like receptor (TLR4) Asp299Gly and Thr399Ile polymorphisms in relation to clinical falciparum malaria among Nigerian children: A multisite cross-sectional immunogenetic study in Lagos. Genes Environ. 2015, 37, 3. [Google Scholar] [CrossRef] [PubMed]
- Aitman, T.J.; Cooper, L.D.; Norsworthy, P.; Wahid, F.N.; Gray, J.K.; Curtis, B.R.; McKeigue, P.M.; Kwiatkowski, D.; Greenwood, B.M.; Snow, R.; et al. Malaria susceptibility and CD36 mutation. Nat. Cell Biol. 2000, 405, 1015–1016. [Google Scholar] [CrossRef]
- Babiker, M.Y.; Mergani, A.; Elwali, N.-E.M. A Cd36 polymorphism associated with eight-times increased susceptibility to cerebral malaria in Central Sudan. Int. J. Med. Med. Sci. 2014, 6, 116–120. [Google Scholar]
- Peng, Z.; Zhan, L.; Chen, S.; Xu, E. Association of transforming growth factor-β1 gene C-509T and T869C polymorphisms with atherosclerotic cerebral infarction in the Chinese: A case-control study. Lipids Health Dis. 2011, 10, 100. [Google Scholar] [CrossRef] [PubMed]
- Hanisch, B.R.; Bangirana, P.; Opoka, R.O.; Park, G.S.; John, C.C. Thrombocytopenia May Mediate Disease Severity in Plasmodium falciparum Malaria Through Reduced Transforming Growth Factor Beta-1 Regulation of Proinflammatory and Anti- inflammatory Cytokines. Pediatr. Infect. Dis. J. 2015, 34, 783–788. [Google Scholar] [CrossRef] [PubMed]
- Grainger, D.J.; Heathcote, K.; Chiano, M.; Snieder, H.; Kemp, P.R.; Metcalfe, J.C.; Carter, N.D.; Spector, T.D. Genetic control of the circulating concentration of transforming growth factor type beta1. Hum. Mol. Genet. 1999, 8, 93–97. [Google Scholar] [CrossRef] [PubMed]
- Hobbs, K.; Negri, J.; Klinnert, M.; Rosenwasser, L.J.; Borish, L. Interleukin-10 and transforming growth factor-beta promoter polymorphisms in allergies and asthma. Am. J. Respir. Crit. Care Med. 1998, 158, 1958–1962. [Google Scholar] [CrossRef] [PubMed]
- Shah, R.; Hurley, C.K.; Posch, P.E. A molecular mechanism for the differential regulation of TGF-beta1 expression due to the common SNP-509C-T (c.-1347C > T). Hum. Genet. 2006, 120, 461–469. [Google Scholar] [CrossRef]
- Greve, B.; Kremsner, P.G.; Lell, B.; Luckner, D.; Schmid, D. Malarial anaemia in African children associated with high oxygen-radical production. Lancet 2000, 355, 40–41. [Google Scholar]
- Mahanta, A.; Kakati, S.; Baruah, S. The association of IL-8-251T/A polymorphism with complicated malaria in Karbi Anglong district of Assam. Cytokine 2014, 65, 210–216. [Google Scholar] [CrossRef] [PubMed]
- Perkins, D.J.; Weinberg, J.B.; Kremsner, P.G. Reduced interleukin-12 and transforming growth factor-beta1 in severe childhood malaria: Relationship of cytokine balance with disease severity. J. Infect. Dis. 2000, 182, 988–992. [Google Scholar] [CrossRef] [PubMed]
- Kaur, H.; Sehgal, R.; Kumar, A.; Sehgal, A.; Bansal, D.; Sultan, A.A. Screening and identification of potential novel biomarker for diagnosis of complicated Plasmodium vivax malaria. J. Transl. Med. 2018, 16, 272. [Google Scholar] [CrossRef]
- Medina, T.S.; Costa, S.P.T.; Oliveira, M.D.; Ventura, A.M.; Souza, J.M.; Gomes, T.F.; Vallinoto, A.C.R.; Póvoa, M.M.; Silva, J.S.; Cunha, M.G.; et al. Increased interleukin-10 and interferon-γ levels in Plasmodium vivax malaria suggest a reciprocal regulation which is not altered by IL-10 gene promoter polymorphism. Malar. J. 2011, 10, 264. [Google Scholar] [CrossRef] [PubMed]
- Andrade, B.B.; Reis-Filho, A.; Souza-Neto, S.M.; Clarêncio, J.; Camargo, L.M.; Barral, A.; Barral-Netto, M. Severe Plasmodium vivax malaria exhibits marked inflammatory imbalance. Malar. J. 2010, 9, 13. [Google Scholar] [CrossRef]
- Banerjee, N.; Nandy, S.; Kearns, J.K.; Bandyopadhyay, A.K.; Das, J.K.; Majumder, P.; Basu, S.; Banerjee, S.; Sau, T.J.; States, J.C.; et al. Polymorphisms in the TNF-α and IL10 gene promoters and risk of arsenic-induced skin lesions and other nondermatological health effects. J. Toxicol. Sci. 2011, 121, 132–139. [Google Scholar] [CrossRef]
- Gibson, A.W.; Edberg, J.C.; Wu, J.; Westendorp, R.G.; Huizinga, T.W.; Kimberly, R.P. Novel single nucleotide polymorphisms in the distal IL-10 promoter affect IL-10 production and enhance the risk of systemic lupus erythematosus. J. Immunol. 2001, 166, 3915–3922. [Google Scholar] [CrossRef]
- Pattaradilokrat, S.; Li, J.; Wu, J.; Qi, Y.; Eastman, R.T.; Zilversmit, M.; Nair, S.C.; Huaman, M.C.; Quinones, M.; Jiang, H.; et al. Plasmodium genetic loci linked to host cytokine and chemokine responses. Genes Immun. 2014, 15, 145–152. [Google Scholar] [CrossRef]
- Ouma, C.; Davenport, G.C.; Were, T.; Otieno, M.F.; Hittner, J.B.; Vulule, J.M.; Martinson, J.; Ong’echa, J.M.; Ferrell, R.E.; Perkins, D.J. Haplotypes of IL-10 promoter variants are associated with susceptibility to severe malarial anemia and functional changes in IL-10 production. Hum. Genet. 2008, 124, 515–524. [Google Scholar] [CrossRef][Green Version]
- Ouyang, W.; Rutz, S.; Crellin, N.K.; Valdez, P.A.; Hymowitz, S.G. Regulation and functions of the IL-10 family of cytokines in inflammation and disease. Annu. Rev. Immunol. 2011, 29, 71–109. [Google Scholar] [CrossRef]
- Hojo-Souza, N.S.; Pereira, D.B.; de Souza, F.S.H.; Mendes, T.A.d.; Cardoso, M.S.; Tada, M.S.; Zanini, G.M.; Bartholomeu, D.C.; Fujiwara, R.T.; Bueno, L.L. On the cytokine/chemokine network during Plasmodium vivax malaria: New insights to understand the disease. Malar. J. 2017, 16, 42. [Google Scholar] [CrossRef]
- Olliaro, P.L.; Barnwell, J.W.; Barry, A.; Mendis, K.; Mueller, I.; Reeder, J.C.; Shanks, G.D.; Snounou, G.; Wongsrichanalai, C. Implications of Plasmodium vivax Biology for Control, Elimination, and Research. Am. J. Trop. Med. Hyg. 2016, 95, 4–14. [Google Scholar] [CrossRef]
- Ohashi, J.; Naka, I.; Patarapotikul, J.; Hananantachai, H.; Looareesuwan, S.; Tokunaga, K. Absence of association between the allele coding methionine at position 29 in the N-terminal domain of ICAM-1 (ICAM-1(Kilifi)) and severe malaria in the northwest of Thailand. Jpn. J. Infect. Dis. 2001, 54, 114–116. [Google Scholar]
- Newbold, C.; Warn, P.; Black, G.; Berendt, A.; Craig, A.; Snow, B.; Msobo, M.; Peshu, N.; Marsh, K. Receptor-specific adhesion and clinical disease in Plasmodium falciparum. Am. J. Trop. Med. Hyg. 1997, 57, 389–398. [Google Scholar] [CrossRef] [PubMed]
- Heddini, A.; Chen, Q.; Obiero, J.; Kai, O.; Fernandez, V.; Marsh, K.; Muller, W.A.; Wahlgren, M. Binding of Plasmodium falciparum-infected erythrocytes to soluble platelet endothelial cell adhesion molecule-1 (PECAM-1/CD31): Frequent recognition by clinical isolates. Am. J. Trop. Med. Hyg. 2001, 65, 47–51. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, D.A.; Lin, J.W.; Brugat, T.; Jarra, W.; Tumwine, I.; Kushinga, G.; Ramesar, J.; Franke-Fayard, B.; Langhorne, J. ICAM-1 is a key receptor mediating cytoadherence and pathology in the Plasmodium chabaudi malaria model. Malar. J. 2017, 16, 185. [Google Scholar] [CrossRef]
- Fry, A.E.; Auburn, S.; Diakite, M.; Green, A.; Richardson, A.; Wilson, J.; Jallow, M.; Sisay-Joof, F.; Pinder, M.; Griffiths, M.J.; et al. Variation in the ICAM1 gene is not associated with severe malaria phenotypes. Genes Immun. 2008, 9, 462–469. [Google Scholar] [CrossRef] [PubMed]
- Vinita, K.; Sripriya, S.; Prathiba, K.; Vaitheeswaran, K.; Sathyabaarathi, R.; Rajesh, M.; Amali, J.; Umashankar, V.; Kumaramanickavel, G.; Pal, S.S.; et al. ICAM-1 K469E polymorphism is a genetic determinant for the clinical risk factors of T2D subjects with retinopathy in Indians: A population-based case-control study. BMJ Open 2012, 2, e001036. [Google Scholar] [CrossRef]
- Fernandez-Reyes, D.; Craig, A.G.; Kyes, S.A.; Peshu, N.; Snow, R.W.; Berendt, A.R.; Marsh, K.; Newbold, C.I. A high frequency African coding polymorphism in the N-terminal domain of ICAM-1 predisposing to cerebral malaria in Kenya. Hum. Mol. Genet. 1997, 6, 1357–1360. [Google Scholar] [CrossRef]
- Carvalho, B.O.; Lopes, S.; Nogueira, P.A.; Orlandi, P.P.; Bargieri, D.; Blanco, Y.C.; Mamoni, R.; Leite, J.A.; Rodrigues, M.M.; Soares, I.D.S.; et al. On the Cytoadhesion ofPlasmodium vivax–Infected Erythrocytes. J. Infect. Dis. 2010, 202, 638–647. [Google Scholar] [CrossRef]
- Costa, F.T.M.; Lopes, S.C.P.; Ferrer, M.; Leite, J.A.; Martin-Jaular, L.; Bernabeu, M.; Nogueira, P.A.; Mourão, M.P.G.; Fernandez-Becerra, C.; Lacerda, M.V.G.; et al. On cytoadhesion of Plasmodium vivax: Raison d’être? Mem. Inst. Oswaldo Cruz 2011, 106, 79–84. [Google Scholar] [CrossRef]
- A Pereira, V.; Sánchez-Arcila, J.C.; Teva, A.; Perce-Da-Silva, D.S.; Vasconcelos, M.P.; Lima, C.A.; Aprígio, C.J.; Rodrigues-Da-Silva, R.N.; O Santos, D.; Banic, D.M.; et al. IL10A genotypic association with decreased IL-10 circulating levels in malaria infected individuals from endemic area of the Brazilian Amazon. Malar. J. 2015, 14, 30. [Google Scholar] [CrossRef] [PubMed]
- Alvarado-Arnez, L.E.; Amaral, E.P.; Sales-Marques, C.; Durães, S.M.; Cardoso, C.C.; Nunes Sarno, E.; Pacheco, A.G.; Lana, F.C.; Moraes, M.O. Association of IL10 Polymorphisms and Leprosy: A Meta-Analysis. PLoS ONE 2015, 10, e0136282. [Google Scholar]
- Abrams, E.T.; Brown, H.; Chensue, S.W.; Turner, G.D.H.; Tadesse, E.; Lema, V.M.; Molyneux, M.E.; Rochford, R.; Meshnick, S.R.; Rogerson, S.J. Host response to malaria during pregnancy: Placental monocyte recruitment is associated with elevated beta chemokine expression. J. Immunol. 2003, 170, 2759–2764. [Google Scholar] [CrossRef]
- Fernandes, A.A.; Carvalho, L.J.; Zanini, G.M.; Ventura, A.M.; Souza, J.M.; Cotias, P.M.; Silva-Filho, I.L.; Daniel-Ribeiro, C.T. Similar cytokine responses and degrees of anemia in patients with Plasmodium falciparum and Plasmodium vivax infections in the Brazilian Amazon region. Clin. Vac. Immunol. 2008, 15, 650–658. [Google Scholar] [CrossRef]
- Deroost, K.; Tyberghein, A.; Lays, N.; Noppen, S.; Schwarzer, E.; Vanstreels, E.; Komuta, M.; Prato, M.; Lin, J.-W.; Pamplona, A.; et al. Hemozoin Induces Lung Inflammation and Correlates with Malaria-Associated Acute Respiratory Distress Syndrome. Am. J. Respir. Cell Mol. Biol. 2013, 48, 589–600. [Google Scholar] [CrossRef]
- Ayimba, E.; Hegewald, J.; Ségbéna, A.Y.; Gantin, R.G.; Lechner, C.J.; Agosssoum, A.; Banla, M.; Soboslay, P.T. Proinflammatory and regulatory cytokines and chemokines in infants with uncomplicated and severe Plasmodium falciparum malaria. Clin. Exp. Immunol. 2011, 166, 218–226. [Google Scholar] [CrossRef]
- Fang, G.F.; Fan, X.Y.; Shen, F.H. The Relationship between Polymorphisms of Interleukin-4 Gene and Silicosis. Biomed. Environ. Sci. 2011, 24, 678–682. [Google Scholar] [PubMed]
- Sortica, V.A.; Cunha, M.G.; Ohnishi, M.D.O.; Souza, J.M.; Ribeiro-dos-Santos, Â.K.C.; Santos, S.E.B.; Hutz, M.H. Role of IL6, IL12B and VDR gene polymorphisms in Plasmodium vivax malaria severity, parasitemia and gametocytemia levels in an Amazonian Brazilian population. Cytokine 2014, 65, 42–47. [Google Scholar] [CrossRef]
- Mockenhaupt, F.; Cramer, J.P.; Hamann, L.; Stegemann, M.S.; Eckert, J.; Oh, N.-R.; Otchwemah, R.N.; Dietz, E.; Ehrhardt, S.; Schroder, N.W.J.; et al. Toll-like receptor (TLR) polymorphisms in African children: Common TLR-4 variants predispose to severe malaria. Proc. Natl. Acad. Sci. USA 2006, 103, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Leoratti, F.M.; Farias, L.; Alves, F.P.; Suarez-Mútis, M.C.; Coura, J.R.; Kali, J.; Camargo, E.P.; Moraes, S.L.; Ramasawmy, R. Variants in the toll-like receptor signaling pathway and clinical outcomes of malaria. J. Infect. Dis. 2008, 198, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Das, A.; Das, T.K.; Sahu, U.; Das, B.P.; Kar, S.K.; Ranjit, M.R. CD36 T188G gene polymorphism and severe falciparum malaria in India. Trans. R. Soc. Trop. Med. Hyg. 2009, 103, 687–690. [Google Scholar] [CrossRef]
- Omi, K.; Ohashi, J.; Patarapotikul, J.; Hananantachai, H.; Naka, I.; Looareesuwan, S.; Tokunaga, K. CD36 polymorphism is associated with protection from cerebral malaria. Am. J. Hum. Genet. 2003, 72, 364–374. [Google Scholar] [CrossRef]
- Damena, D.; Denis, A.; Golassa, L.; Chimusa, E.R. Genome-wide association studies of severe P. falciparum malaria susceptibility: Progress, pitfalls and prospects. BMC Med. Genom. 2019, 12, 120. [Google Scholar] [CrossRef]
- Malaria Genomic Epidemiology Network. Insights into malaria susceptibility using genome-wide data on 17,000 individuals from Africa, Asia and Oceania. Nat. Commun. 2019, 10, 1. [Google Scholar]
- Ravenhall, M.; Campino, S.; Sepúlveda, N.; Manjurano, A.; Nadjm, B.; Mtove, G.; Wangai, H.; Maxwell, C.; Olomi, R.; Reyburn, H.; et al. Novel genetic polymorphisms associated with severe malaria and under selective pressure in North-eastern Tanzania. PLoS Gen. 2018, 30, 1. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).