Preliminary Characterization of Triatomine Bug Blood Meals on the Island of Trinidad Reveals Opportunistic Feeding Behavior on Both Human and Animal Hosts

 ,
, 
Abstract
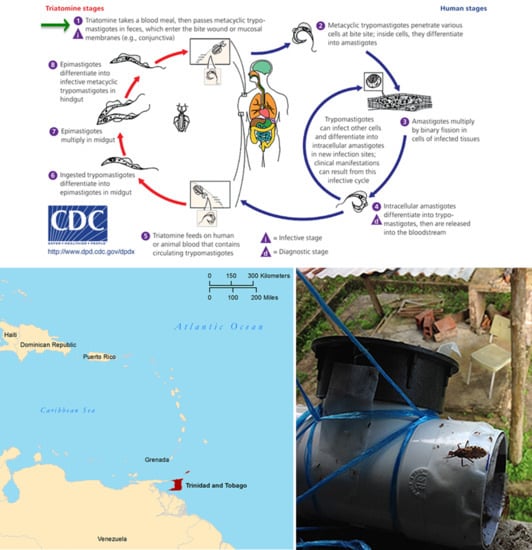
1. Introduction
2. Materials and Methods
2.1. Overview
2.2. Animal Use Ethics Statement
2.3. Study Sites
2.4. Triatomine Bug Collection
2.5. Specimen Processing and Molecular Analyses
2.6. Blood Meal Analyses
3. Results
3.1. T. cruzi Infection
3.2. Blood Meal Analyses
4. Discussion
4.1. P. geniculatus Breeding Habitat Remains Cryptic
4.2. Are Wildlife Population Declines Driving P. geniculatus Host Shifts?
4.3. P. geniculatus: A Cosmopolitan Species of Emerging Importance
4.4. Multiple Paths to T. cruzi Exposure in Trinidad
4.5. Chagas Disease in Trinidad: Official Endemicity Would Bring Critical Resources
4.6. Study Limitations
5. Conclusions and Future Directions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Chagas disease in Latin America: An epidemiological update based on 2010 estimates. Wkly. Epidemiol. Rec. 2015, 33–44. [Google Scholar] [CrossRef]
- Shikanai-Yasuda, M.A.; Carvalho, N.B. Oral transmission of chagas disease. Clin. Infect. Dis. 2012, 54, 845–852. [Google Scholar] [CrossRef] [PubMed]
- Miles, M. Orally acquired Chagas disease: Lessons from an urban school outbreak. J. Infect. Dis. 2010, 201, 1282–1284. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Filigheddu, M.T.; Górgolas, M.; Ramos, J.M. Orally-transmitted Chagas disease. Med. Clín. (Engl. Ed.) 2017, 148, 125–131. [Google Scholar] [CrossRef]
- Nóbrega, A.A.; Garcia, M.H.; Tatto, E.; Obara, M.T.; Costa, E.; Sobel, J.; Araujo, W.N. Oral transmission of chagas disease by consumption of Acai palm fruit, Brazil. Emerg. Infect. Dis. 2009, 15, 653–655. [Google Scholar] [CrossRef] [PubMed]
- Gottdenker, N.L.; Chaves, L.F.; Calzada, J.E.; Saldaña, A.; Carroll, C.R. host life history strategy, species diversity, and habitat influence Trypanosoma cruzi vector infection in changing landscapes. PLoS Negl. Trop. Dis. 2012, 6, e1884. [Google Scholar] [CrossRef] [PubMed]
- Guhl, F.; Pinto, N.; Aguilera, G. Sylvatic triatominae: A new challenge in vector control transmission. Mem. Do Inst. Oswaldo Cruz 2009, 104, 71–75. [Google Scholar] [CrossRef]
- Rabinovich, J.E.; Kitron, U.D.; Obed, Y.; Yoshioka, M.; Gottdenker, N.; Chaves, L.F. Ecological patterns of blood-feeding by kissing-bugs (Hemiptera: Reduviidae: Triatominae). Memórias Do Inst. Oswaldo Cruz 2011, 106, 479–494. [Google Scholar] [CrossRef]
- Petana, W.B. American trypanosomiasis (Chagas’ disease) in the Caribbean. Bull. Panam. Health Organ. 1978, 12, 45–51. [Google Scholar]
- Centers for Disease Control and Prevention. Parasites-American Trypanosomiasis (also known as Chagas Disease). Available online: https://www.cdc.gov/parasites/chagas/gen_info/detailed.html (accessed on 31 May 2020).
- Armed Forces Pest Management Board. Regional Disease Vector Ecology Profile: Caribbean; Walter Reed Army Medical Center: Washington, DC, USA, 2002. [Google Scholar]
- Fistein, B. A review of Chagas disease in Trinidad. Caribb. Med. J. 1981, 42, 17–28. [Google Scholar]
- Fistein, B.; Sutton, R.N.P. Chagas disease in the West Indies. Lancet 1963, 281, 330–331. [Google Scholar] [CrossRef]
- Fistein, B. Aetiology of Jamaican cardiomyopathy. Lancet 1966, 288, 386. [Google Scholar] [CrossRef]
- World Health Organization. Working to Overcome the Global Impact of Neglected Tropical Diseases First WHO Report on Neglected Tropical Diseases; No. WHO/HTM/NTD/2010.1.; World Health Organization: Geneva, Switzerland, 2010. [Google Scholar]
- Erlich, R.N.; Barrett, S.F. Cenozoic Plate tectonic history of the northern Venezuela-Trinidad Area. Tectonics 1990, 9, 161–184. [Google Scholar] [CrossRef]
- Omah-Maharaj, I. Studies on vectors of Trypanosoma cruzi in Trinidad. Med. Vet. Entomol. 1992, 6, 115–120. [Google Scholar] [CrossRef]
- Ryckman, R.E. The Triatominae of South America: A checklist with synonymy (Hemiptera: Reduviidae: Triatominae). Bull. Soc. Vector Ecol. 1986, 11, 199–208. [Google Scholar]
- Cazorla-Perfetti, D. Revisión de los vectores de la enfermedad de Chagas en Venezuela (Hemiptera-Heteroptera, Reduviidae, Triatominae). Saber Univ. De Orient Venez. 2016, 28, 387–470. [Google Scholar]
- Reyes-Lugo, M.; Rodriguez-Acosta, A. Domiciliation of the sylvatic Chagas disease vector Panstrongylus geniculatus Latreille, 1811 (Triatominae: Reduviidae) in Venezuela. Trans. R. Soc. Trop. Med. Hyg. 2000, 94, 508. [Google Scholar] [CrossRef]
- Feliciangeli, M.D.; Carrasco, H.; Patterson, J.S.; Suarez, B.; Martínez, C.; Medina, M. Mixed domestic infestation by Rhodnius prolixus StäL, 1859 and Panstrongylus geniculatus Latreille, 1811, vector incrimination, and seroprevalence for Trypanosoma cruzi among inhabitants in El Guamito, Lara State, Venezuela. Am. J. Trop. Med. Hyg. 2004, 71, 501–505. [Google Scholar] [CrossRef]
- Carrasco, H.J.; Torrellas, A.; García, C.; Segovia, M.; Feliciangeli, M.D. Risk of Trypanosoma cruzi I (Kinetoplastida: Trypanosomatidae) transmission by Panstrongylus geniculatus (Hemiptera: Reduviidae) in Caracas (Metropolitan District) and neighboring States, Venezuela. Int. J. Parasitol. 2005, 35, 1379–1384. [Google Scholar] [CrossRef]
- Carrasco, H.J.; Segovia, M.; Londoño, J.C.; Ortegoza, J.; Rodríguez, M.; Martínez, C.E. Panstrongylus geniculatus and four other species of triatomine bug involved in the Trypanosoma cruzi enzootic cycle: High risk factors for Chagas’ disease transmission in the Metropolitan District of Caracas, Venezuela. Parasites Vectors 2014, 7, 1003–1028. [Google Scholar] [CrossRef]
- Rojas-Cortez, M.; Pinazo, M.J.; Garcia, L.; Arteaga, M.; Uriona, L.; Gamboa, S.; Mejía, C.; Lozano, D.; Gascon, J.; Torrico, F.; et al. Trypanosoma cruzi-infected Panstrongylus geniculatus and Rhodnius robustus adults invade households in the Tropics of Cochabamba region of Bolivia. Parasites Vectors 2016, 9, 1–6. [Google Scholar] [CrossRef]
- Fistein, B. Trypanosoma cruzi infection in blood-sucking Reduviid bugs in Trinidad. Trans. R. Soc. Trop. Med. Hyg. 1966, 60, 536–538. [Google Scholar] [CrossRef]
- Aziz, E.S.; Parsad, K.R. Trypanosoma cruzi infection associated with Chagas’ heart disease in humans in South Trinidad. West. Indian Med. J. 1993, 42, 18. [Google Scholar]
- Downs, W.G. The presence of Trypanosoma cruzi in the island of Trinidad, W.I. J. Parasitol. 1963, 49, 50. [Google Scholar] [CrossRef]
- Abad-Franch, F.; Noireau, F.; Paucar, A.C.; Aguilar, H.M.V.; Carpio, C.C.; Racines, J.V. The use of live-bait traps for the study of sylvatic Rhodnius populations (Hemiptera: Reduviidae) in palm trees. Trans. R. Soc. Trop. Med. Hyg. 2000, 94, 629–630. [Google Scholar] [CrossRef]
- Curtis-Robles, R.; Wozniak, E.J.; Auckland, L.D.; Hamer, G.L.; Hamer, S.A. Combining public health education and disease ecology research: Using citizen science to assess Chagas disease entomological risk in Texas. PLoS Negl. Trop. Dis. 2015, 9, e0004235. [Google Scholar] [CrossRef]
- Lent, H.; Wygodzinsky, Z. Revision of the Triatominae (Hemiptera, Reduviidae), and their significance as vectors of Chagas’ disease. Bull. Am. Mus. Nat. Hist. 1979, 163, 1–520. [Google Scholar]
- Carcavallo, R.U.; Galindez Giron, I.; Jurberg, J.; Lent, H. Atlas of Chagas Disease Vectors in the Americas; Editora Fiocruz: Rio de Janeiro, Brazil, 1998. [Google Scholar]
- Patterson, J.S.; Barbosa, S.E.; Feliciangeli, M.D. On the genus Panstrongylus Berg 1879: Evolution, ecology and epidemiological significance. Acta Trop. 2009, 110, 187–199. [Google Scholar] [CrossRef]
- Peterson, J.K.; Graham, A.L.; Dobson, A.P.; Chavez, O.T. Rhodnius prolixus life history outcomes differ when Infected with different Trypanosoma cruzi I strains. Am. J. Trop. Med. Hyg. 2015, 93, 564–572. [Google Scholar] [CrossRef]
- Combs, T.P.; Nagajyothi; Mukherjee, S.; de Almeida, C.J.G.; Jelicks, L.A.; Schubert, W.; Lin, Y.; Jayabalan, D.S.; Zhao, D.; Braunstein, V.L.; et al. The adipocyte as an important target cell for Trypanosoma cruzi infection. J. Biol. Chem. 2005, 280, 24085–24094. [Google Scholar] [CrossRef]
- Boakye, D.A.; Tang, J.; Truc, P.; Merriweather, A.; Unnasch, T.R. Identification of bloodmeals in haematophagous Diptera by cytochrome B heteroduplex analysis. Med. Vet. Entomol. 1999, 13, 282–287. [Google Scholar] [CrossRef]
- Ngo, K.A.; Kramer, L.D. Identification of mosquito bloodmeals using polymerase chain reaction (PCR) with order-specific primers. J. Med. Entomol. 2003, 40, 215–222. [Google Scholar] [CrossRef]
- Pifano, F. El potencial enzootico silvestre del complejo ecológico Schizotrypanum cruzi—Didelphis marsupialis—Panstrongylus geniculatus y sus incursiones a la vivienda humana del valle de Caracas. Bol Acad Cien Fis Mat. Nat. (Caracas) 1986, XLVI, 9–37. [Google Scholar]
- Peterson, J.K.; Bartsch, S.M.; Lee, B.Y.; Dobson, A.P. Broad patterns in domestic vector-borne Trypanosoma cruzi transmission dynamics: Synanthropic animals and vector control. Parasites Vectors 2015, 8, 537. [Google Scholar] [CrossRef]
- Martínez-Ibarra, J.A.; Martínez-Grant, J.A.; Verdugo-Cervantes, M.R.; Bustos-Saldaña, R.; Nogueda-Torres, B. Monitoring triatomid bug (Hemiptera: Reduviidae) presence by sentinel chicken coops in Southern Jalisco State, México. Biomed. Rev. Del Inst. Nac. De Salud 2010, 30, 140–145. [Google Scholar]
- Valente, V.D.C. Potential for domestication of Panstrongylus geniculatus (Latreille 1811) (Hemiptera, Reduviidae, Triatominae) in the municipality of Muaná, Marajó Island, Pará State, Brazil. Mem. Do Inst. Oswaldo Cruz 1999, 94, 399–400. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Valente, V.C.; Valente, S.A.S.; Noireau, F.; Carrasco, H.J.; Miles, M.A. Chagas disease in the Amazon basin: Associaion of Panstrongylus geniculatus (Hemiptera: Reduviidae) with domestic pigs. J. Med. Entomol. 1998, 35, 5. [Google Scholar] [CrossRef]
- Wolff, M.; Castillo, D. Evidencias de domesticación y aspectos biológicos de panstrongylus de Panstrongylus geniculatus (Latreille 1811) (Hemiptera: Reduviidae). Acta Entomol. Chil. 2000, 24, 77–83. [Google Scholar]
- Rabinovich, J.E.; Feliciangeli, M.D. Vital statistics of Triatominae (Hemiptera: Reduviidae) under laboratory conditions: IV. Panstrongylus Geniculatus. J. Med. Entomol. 2015, 52, 797–805. [Google Scholar] [CrossRef]
- Ceretti-Junior, W.; Vendrami, D.P.; de Matos-Junior, M.O.; Rimoldi-Ribeiro, A.; Alvarez, J.V.; Marques, S.; Duarte, A.N.; da Silva, R.A.; da Rosa, J.A.; Marrelli, M.T. Occurrences of triatomines (Hemiptera: Reduviidae) and first reports of Panstrongylus geniculatus in urban environments in the city of Sao Paulo, Brazil. Rev. Inst. Med. Trop. S. Paulo 2018, 60. [Google Scholar] [CrossRef] [PubMed]
- 45. Peixoto, S.R.; da Silva Rocha, D.; Dale, C.; Galvão, C. Panstrongylus geniculatus (Latreille, 1811) (Hemiptera, Reduviidae, Triatominae): First record on Ilha Grande, Rio de Janeiro, Brazil. Check List 2020, 16, 391–394. [Google Scholar] [CrossRef]
- Segovia, M.; Carrasco, H.J.; Martínez, C.E.; Messenger, L.A.; Nessi, A.; Londoño, J.C.; Espinosa, R.; Martínez, C.; Alfredo, M.; Bonfante-Cabarcas, R.; et al. Molecular epidemiologic source tracking of orally transmitted chagas disease, Venezuela. Emerg. Infect. Dis. 2013, 19, 1098–1101. [Google Scholar] [CrossRef]
- Alarcón de Noya, B.; Díaz-Bello, Z.; Colmenares, C.; Ruiz-Guevara, R.; Mauriello, L.; Zavala-Jaspe, R.; Suarez, J.A.; Abate, T.; Naranjo, L.; Paiva, M.; et al. Large urban outbreak of orally acquired acute Chagas disease at a school in Caracas, Venezuela. J. Infect. Dis. 2010, 201, 1308–1315. [Google Scholar] [CrossRef]
- Brenière, S.F.; Waleckx, E.; Aznar, C. Other forms of transmission. In American Trypanosomiasis Chagas Disease; Elsevier: Amsterdam, The Netherlands, 2017; pp. 561–578. ISBN 978-0-12-801029-7. [Google Scholar]
- de Noya, B.A.; Díaz-Bello, Z.; Colmenares, C.; Ruiz-Guevara, R.; Mauriello, L.; Muñoz-Calderón, A.; Noya, O. Update on oral Chagas disease outbreaks in Venezuela: Epidemiological, clinical and diagnostic approaches. Mem. Inst. Oswaldo Cruz 2015, 110, 377–386. [Google Scholar] [CrossRef]
- Pan American Health Organization. Chagas Disease; Pan American Health Organization: Washington, DC, USA, 2017. [Google Scholar]
- Manne, J.; Snively, C.S.; Levy, M.Z.; Reich, M.R. Supply chain problems for Chagas disease treatment. Lancet Infect. Dis. 2012, 12, 173–175. [Google Scholar] [CrossRef]
- Pan American Health Organization. Supply of Blood for Transfusion in Latin American and Caribbean Countries, 2014 and 2015; Pan American Health Organization: Washington, DC, USA, 2017. [Google Scholar]
- Peterson, J.K.; Yoshioka, K.; Hashimoto, K.; Caranci, A.; Gottdenker, N.; Monroy, C.; Saldaña, A.; Rodriguez, S.; Dorn, P.; Zúniga, C. Chagas disease epidemiology in Central America: An update. Curr. Trop. Med. Rep. 2019, 6, 92–105. [Google Scholar] [CrossRef]
- Pan American Health Organization. EMTCT Plus. In Framework for Elimination of Mother-to-Child Transmission of HIV, Syphilis, Hepatitis B, and Chagas; Pan American Health Organization: Washington, DC, USA, 2017. [Google Scholar]
- Schofield, C.J.; Dias, J.C.P. The southern cone initiative against Chagas disease. Adv. Parasitol. 1999, 42, 1–27. [Google Scholar]
- Iniciativa de los Países de América Central, Para la Interrupción de la Transmisión Vectorial y Transfusional de la Enfermedad de Chagas (IPCA). Historia de 12 años de una Iniciativa Subregional 1998-2010. Representación de la OPS/OMS en Honduras; 2011; 89p, ISBN 978-99926-742-5-3. [Google Scholar]
- Pan American Health Organization. 2017–2021 PAHO/WHO Country Cooperation Strategy Report Trinidad and Tobago; Pan American Health Organization: Washington, DC, USA, 2018. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site Name | Location | Site Description | Bugs Collected 1,2 | Human Blood Meals | ||||
|---|---|---|---|---|---|---|---|---|
| Infected | Uninfected | Total | Infected 3 | Uninfected 3 | Total | |||
| Blanchisseuse | Northern coast | Coastal human home adjacent to secondary forest | 1 (100%) | 0 | 1 | 1 (100%) | 0 | 1 |
| Coal Mine | Central range 4 | Scattered human homes adjacent to secondary forest | 40 (87%) | 6 (13%) | 46 | 22 (92%) | 2 (8%) | 24 |
| Matura | Northeast coast | Rural human home surrounded by secondary forest | 1 (50%) | 1 (50%) | 2 | 0 | 1 (100%) | 1 |
| Mt Harris | Central range 4 | Rural neighborhood in close proximity to secondary forest | 3 (60%) | 2 (40%) | 5 | 2 (67%) | 1 (33%) | 3 |
| Santa Cruz | Northern range 4 | Laundry area of peri-urban human home | 1 (100%) | 0 | 1 | 1 (100%) | 0 | 1 |
| Blood Meal Hosts 1 | Triatomine Bugs 2 | Collection Sites 5 | BLAST Results 3 | ||||
|---|---|---|---|---|---|---|---|
| Class | Genus/Species | Common Name | Blood Meals 1 | T. cruzi-Positive Bugs 4 | % Identity Match | E-Value | |
| Mammalia | Homo sapiens | Human | 30 | 26 (87%) | All | 99% | 1 × 10−58 |
| Alouatta seniculus | Red howler monkey | 2 | 2 (100%) | CM, MH | 98% | 5 × 10−134 | |
| Dasyprocta leporina | Red-rumped agouti | 2 | 2 (100%) | CM, MT | 100% | 2 × 10−128 | |
| Coendou prehensilis | Brazilian porcupine | 1 | 1 (100%) | CM | 100% | 1 × 10−144 | |
| Herpestes javanicus | Javan mongoose | 1 | 1 (100%) | CM | 100% | 8 × 10−146 | |
| Metachirus nudicaudatus | Brown four-eyed opossum | 1 | 0 (0%) | CM | 99% | 8 × 10−136 | |
| Nectomys6 | Water rat | 1 | 1 (100%) | CM | 98% | 9 × 10−138 | |
| Proechimys trinitatis | Trinidad spiny rat | 1 | 0 (0%) | MH | 100% | 3 × 10−64 | |
| Aves | Gallus gallus | Chicken | 10 | 9 (10%) | CM | 99% | 3 × 10−106 |
| Pionus menstruus | Blue-headed parrot | 2 | 1 (50%) | CM | 90% | 2.5 × 10−77 | |
| Dryocopus lineatus | Lineated woodpecker | 1 | 1 (100%) | CM | 99% | 1 × 10−92 | |
| Pulsatrix perspicillata | Spectacled owl | 1 | 0 (0%) | CM | 98% | 0 * | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hylton, A.; Fitzpatrick, D.M.; Suepaul, R.; Dobson, A.P.; Charles, R.A.; Peterson, J.K. Preliminary Characterization of Triatomine Bug Blood Meals on the Island of Trinidad Reveals Opportunistic Feeding Behavior on Both Human and Animal Hosts. Trop. Med. Infect. Dis. 2020, 5, 166. https://doi.org/10.3390/tropicalmed5040166
Hylton A, Fitzpatrick DM, Suepaul R, Dobson AP, Charles RA, Peterson JK. Preliminary Characterization of Triatomine Bug Blood Meals on the Island of Trinidad Reveals Opportunistic Feeding Behavior on Both Human and Animal Hosts. Tropical Medicine and Infectious Disease. 2020; 5(4):166. https://doi.org/10.3390/tropicalmed5040166
Chicago/Turabian StyleHylton, Alexandra, Daniel M. Fitzpatrick, Rod Suepaul, Andrew P. Dobson, Roxanne A. Charles, and Jennifer K. Peterson. 2020. "Preliminary Characterization of Triatomine Bug Blood Meals on the Island of Trinidad Reveals Opportunistic Feeding Behavior on Both Human and Animal Hosts" Tropical Medicine and Infectious Disease 5, no. 4: 166. https://doi.org/10.3390/tropicalmed5040166
APA StyleHylton, A., Fitzpatrick, D. M., Suepaul, R., Dobson, A. P., Charles, R. A., & Peterson, J. K. (2020). Preliminary Characterization of Triatomine Bug Blood Meals on the Island of Trinidad Reveals Opportunistic Feeding Behavior on Both Human and Animal Hosts. Tropical Medicine and Infectious Disease, 5(4), 166. https://doi.org/10.3390/tropicalmed5040166