In Vivo Efficacy of SYN023, an Anti-Rabies Monoclonal Antibody Cocktail, in Post-Exposure Prophylaxis Animal Models


Abstract
1. Introduction
2. Materials and Methods
2.1. Challenge Virus Isolate
2.2. Biologics
2.3. In Vivo Syrian Hamster Challenge Model
2.4. In Vivo Syrian Hamster Non-Challenge Model
2.5. In Vivo Beagle Challenge Model
2.6. Ethics Statement
3. Results
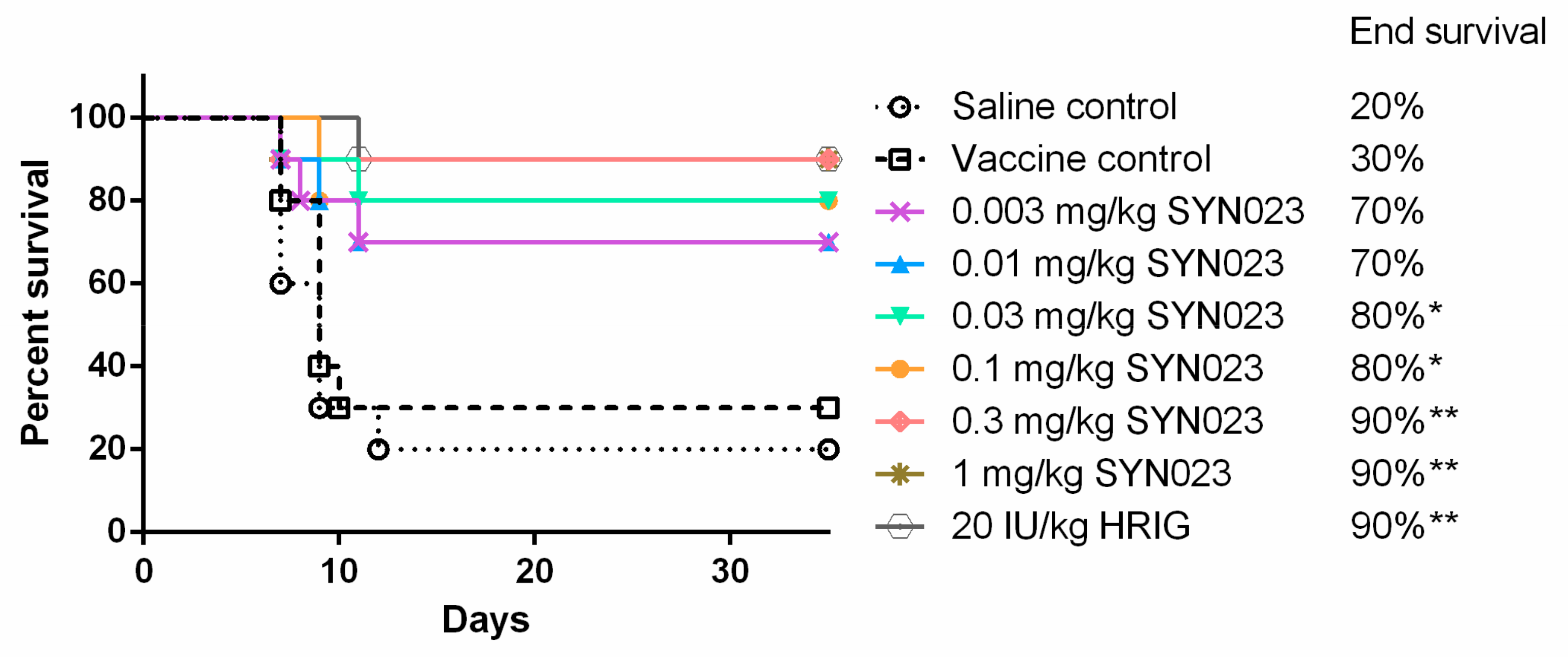
3.1. SYN023 Offers Significant Protection as an In Vivo Model of PEP
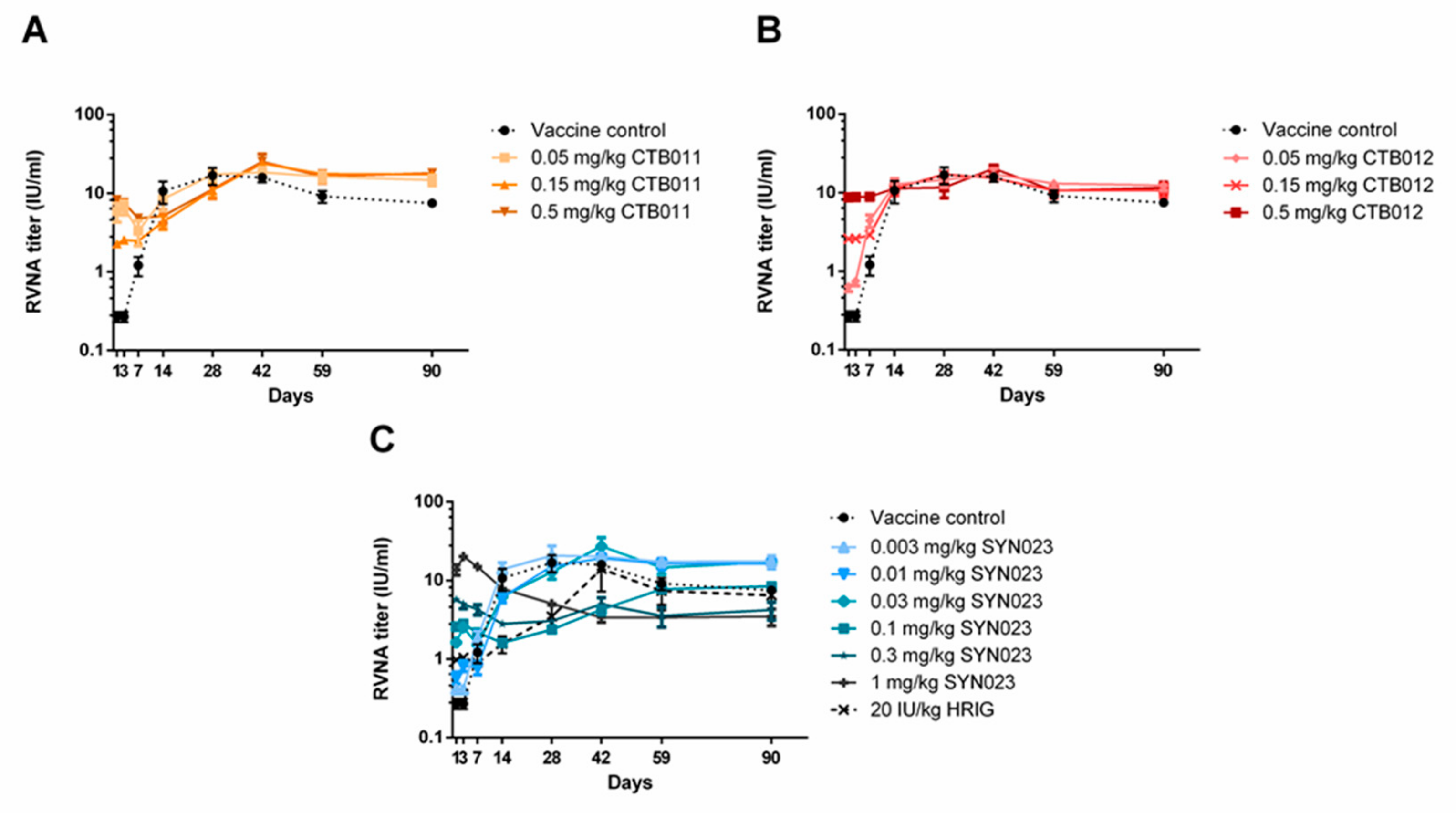
3.2. Effects of CTB011, CTB012, and SYN023 on Serum RVNA Levels in Syrian Hamsters
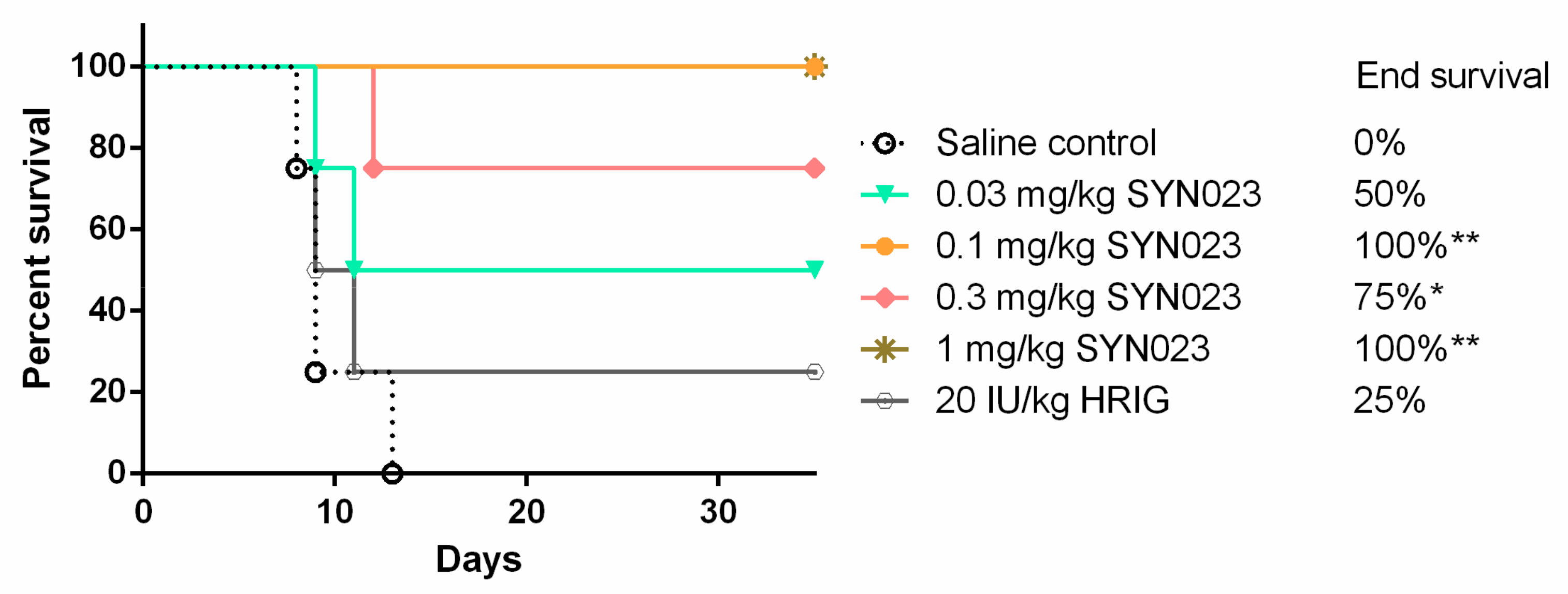
3.3. SYN023 Protects Beagles from a Lethal Challenge of BD06 Street RABV
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rupprecht, C.E.; Hanlon, C.A.; Hemachudha, T. Rabies re-examined. Lancet Infect. Dis. 2002, 2, 327–343. [Google Scholar] [CrossRef]
- Rabies. Available online: https://www.who.int/news-room/fact-sheets/detail/rabies (accessed on 9 December 2019).
- World Health Organization. WHO Expert Consultation on Rabies, Third Report. 2018. Available online: https://apps.who.int/iris/bitstream/10665/85346/1/9789241209823_eng.pdf (accessed on 9 December 2019).
- Dietzschold, B.; Kao, M.; Zheng, Y.M.; Chen, Z.Y.; Maul, G.; Fu, Z.F.; Rupprecht, C.E.; Koprowski, H. Delineation of putative mechanisms involved in antibody-mediated clearance of rabies virus from the central nervous system. Proc. Natl. Acad. Sci. USA 1992, 89, 7252–7256. [Google Scholar] [CrossRef] [PubMed]
- Why a Simple, Lifesaving Rabies Shot Can Cost $10,000 in America. Available online: https://www.vox.com/policy-and-politics/2018/2/7/16851134/rabies-treament-expensive-emergency-room (accessed on 9 December 2019).
- Sparrow, E.; Torvaldsen, S.; Newall, A.T.; Wood, J.G.; Sheikh, M.; Kieny, M.P.; Abela-Ridder, B. Recent advances in the development of monoclonal antibodies for rabies post exposure prophylaxis: A review of the current status of the clinical development pipeline. Vaccine 2019, 37, A132–A139. [Google Scholar] [CrossRef]
- Müller, T.; Dietzschold, B.; Ertl, H.; Fooks, A.R.; Freuling, C.; Fehlner-Gardiner, C.; Kliemt, J.; Meslin, F.X.; Franka, R.; Rupprecht, C.E.; et al. Development of a mouse monoclonal antibody cocktail for post-exposure rabies prophylaxis in humans. PLoS Negl. Trop. Dis. 2009, 3, e542. [Google Scholar] [CrossRef]
- Nagarajan, T.; Rupprecht, C.E.; Dessain, S.K.; Rangarajan, P.N.; Thiagarajan, D.; Srinivasan, V.A. Human monoclonal antibody and vaccine approaches to prevent human rabies. Curr. Top. Microbiol. Immunol. 2008, 317, 67–101. [Google Scholar] [CrossRef]
- Bakker, A.B.; Marissen, W.E.; Kramer, R.A.; Rice, A.B.; Weldon, W.C.; Niezgoda, M.; Hanlon, C.A.; Thijsse, S.; Backus, H.H.; De Kruif, J.; et al. Novel human monoclonal antibody combination effectively neutralizing natural rabies virus variants and individual in vitro escape mutants. J. Virol. 2005, 79, 9062–9068. [Google Scholar] [CrossRef]
- Sloan, S.E.; Hanlon, C.; Weldon, W.; Niezgoda, M.; Blanton, J.; Self, J.; Rowley, K.J.; Mandell, R.B.; Babcock, G.J.; Thomas, W.D., Jr.; et al. Identification and characterization of a human monoclonal antibody that potently neutralizes a broad panel of rabies virus isolates. Vaccine 2007, 25, 2800–2810. [Google Scholar] [CrossRef]
- Nagarajan, T.; Marissen, W.; Rupprecht, C.E. Monoclonal antibodies for the prevention of rabies: Theory and clinical practice. Antib. Technol. J. 2014, 4, 1–12. [Google Scholar] [CrossRef]
- Shivalingaiah, A.H.; Shankaraiah, R.H.; Hanumanthaiah, A.N.D. Safety of new indigenous human Rabies Monoclonal Antibody (RMAb) for post exposure prophylaxis. Indian J. Community Health 2018, 30, 196–201. [Google Scholar]
- Gogtay, N.J.; Munshi, R.; Narayana, D.H.A.; Mahendra, B.J.; Kshirsagar, V.; Gunale, B.; Moore, S.; Cheslock, P.; Thaker, S.; Deshpande, S.; et al. Comparison of a novel human rabies monoclonal antibody to human rabies immunoglobulin for postexposure prophylaxis: A phase 2/3, randomized, single-blind, noninferiority, controlled study. Clin. Infect. Dis. 2018, 6, 387–395. [Google Scholar] [CrossRef]
- Ilina, E.N.; Larina, M.V.; Aliev, T.K.; Dolgikh, D.A.; Kirpichnikov, M.P. Recombinant monoclonal antibodies for rabies post-exposure prophylaxis. Biochemistry 2018, 83, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Chao, T.Y.; Ren, S.; Shen, E.; Moore, S.; Zhang, S.F.; Chen, L.; Rupprecht, C.E.; Tsao, E. SYN023, a novel humanized monoclonal antibody cocktail, for post-exposure prophylaxis of rabies. PLoS Negl. Trop. Dis. 2017, 11, e0006133. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Ma, J.; Nie, J.; Li, J.; Cao, S.; Wang, L.; Yu, C.; Huang, W.; Li, Y.; Yu, Y.; et al. Antigenic variations of recent street rabies virus. Emerg. Microbes Infect. 2019, 8, 1584–1592. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhang, S.F.; Pan, T.L.; Zhang, F.; Wang, Y.; Lian, H.; Zhang, J.X.; Hu, R.L. Characteristics of rabies street virus strain BD06 for challenge test in dogs. Chin. J. Biol. 2014, 27, 33–35. [Google Scholar]
- Dean, D.J.; Abelseth, M.K.; Atanasiu, P. The Fluorescent Antibody Test. In Laboratory Techniques in Rabies, 4th ed.; Meslin, F.X., Kaplan, M.M., Koprowski, H., Eds.; World Health Organization: Geneva, Switzerland, 1996; pp. 88–95. [Google Scholar]
- Smith, J.S.; Yager, P.A.; Baer, M. A rapid fluorescent focus inhibition test (RFFIT) for determining rabies virus-neutralizing antibody. In Laboratory Techniques in Rabies, 4th ed.; Meslin, F.X., Kaplan, M.M., Koprowski, H., Eds.; World Health Organization: Geneva, Switzerland, 1996; pp. 181–191. [Google Scholar]
- Cabasso, V.J.; Loofbourow, J.C.; Roby, R.E.; Anuskiewicz, W. Rabies immune globulin of human origin: Preparation and dosage determination in non-exposed volunteer subjects. Bull. World Health Organ. 1971, 45, 303–315. [Google Scholar]
- WHO. Rabies Vaccines: WHO Position Paper-April 2018. Wkly. Epidemiol. Rec. 2018, 93, 201–220. Available online: https://www.who.int/rabies/resources/who_wer9316/en/ (accessed on 9 December 2019).
- Wang, X.; Zhang, S.; Sun, C.; Yuan, Z.G.; Wu, X.; Wang, D.; Ding, Z.; Hu, R. Proteomic profiles of mouse neuro N2a cells infected with variant virulence of rabies viruses. J. Microbiol. Biotechnol. 2011, 21, 366–373. [Google Scholar]
- Franka, R.; Carson, W.C.; Ellison, J.A.; Taylor, S.T.; Smith, T.G.; Kuzmina, N.A.; Kuzmin, I.V.; Marissen, W.E.; Rupprecht, C.E. In vivo efficacy of a cocktail of human monoclonal antibodies (CL184) against diverse North American bat rabies virus variants. Trop. Med. Infect. Dis. 2017, 2, 48. [Google Scholar] [CrossRef] [PubMed]
- De Benedictis, P.; Minola, A.; Rota Nodari, E.; Aiello, R.; Zecchin, B.; Salomoni, A.; Foglierini, M.; Agatic, G.; Vanzetta, F.; Lavenir, R.; et al. Development of broad-spectrum human monoclonal antibodies for rabies post-exposure prophylaxis. EMBO Mol. Med. 2016, 8, 407–421. [Google Scholar] [CrossRef]
- Warrell, M.J.; Warrell, D.A.; Suntharasamai, P.; Viravan, C.; Sinhaseni, A.; Udomsakdi, D.; Phanfung, R.; Xueref, C.; Vincent-Falquet, J.C.; Nicholson, K.G.; et al. An economical regimen of human diploid cell strain anti-rabies vaccine for post-exposure prophylaxis. Lancet 1983, 2, 301–304. [Google Scholar] [CrossRef]
- Helmick, C.G.; Johnstone, C.; Sumner, J.; Winkler, W.G.; Fager, S. A clinical study of Merieux human rabies immune globulin. J. Biol. Stand. 1982, 10, 357–367. [Google Scholar] [CrossRef]
- Vodopija, I.; Sureau, P.; Smerdel, S.; Lafon, M.; Baklaić, Z.; Ljubicić, M.; Svjetlicić, M. Interaction of rabies vaccine with human rabies immunoglobulin and reliability of a 2-1-1 schedule application for postexposure treatment. Vaccine 1988, 6, 283–286. [Google Scholar] [CrossRef]
- Lang, J.; Simanjuntak, G.H.; Soerjosembodo, S.; Koesharyono, C. Suppressant effect of human or equine rabies immunoglobulins on the immunogenicity of post-exposure rabies vaccination under the 2-1-1 regimen: A field trial in Indonesia. Bull. World Health Organ. 1998, 76, 491–495. [Google Scholar]
- Suntharasamai, P.; Chaiprasithikul, P.; Wasi, C.; Supanaranond, W.; Auewarakul, P.; Chanthavanich, P.; Supapochana, A.; Areeraksa, S.; Chittamas, S.; Jittapalapongsa, S.; et al. A simplified and economical intradermal regimen of purified chick embryo cell rabies vaccine for postexposure prophylaxis. Vaccine 1994, 12, 508–512. [Google Scholar] [CrossRef]
- Hanna, K.; Cruz, M.C.; Mondou, E.; Corsi, E.; Vandeberg, P. Safety and neutralizing rabies antibody in healthy subjects given a single dose of rabies immune globulin caprylate/chromatography purified. Clin. Pharmacol. 2018, 10, 79–88. [Google Scholar] [CrossRef]
- Wilson, P.J.; Oertli, E.H.; Hunt, P.R.; Sidwa, T.J. Evaluation of a postexposure rabies prophylaxis protocol for domestic animals in Texas: 2000–2009. J. Am. Vet. Med. Assoc. 2010, 237, 1395–1401. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Group No. | Treatment | Survival | Serum RVNA Levels Determined by RFFIT (IU/mL) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| #1 | #2 | #3 | #4 | #5 | #6 | #7 | #8 | #9 | Mean ± SD 1 | |||
| 1 | Saline | 2 | 0 | 0 | − | − | − | − | − | − | − | 0 |
| 2 | Vaccine | 3 | 14 | 4.0 | 7.8 | − | − | − | − | − | − | 8.7 ± 5.2 |
| 3 | 0.003 mg/kg SYN023 | 7 | 8.3 | 12 | 12 | 5.7 | 5.6 | 19 | 2.9 | − | − | 9.4 ± 5.5 |
| 4 | 0.01 mg/kg SYN023 | 7 | 9.2 | 9.2 | 11 | 7.9 | 3.1 | 11 | 9.7 | − | − | 8.7 ± 2.7 |
| 5 | 0.03 mg/kg SYN023 | 8 | 8.7 | 5.5 | 10 | 8.8 | 12 | 6.7 | 11 | 9.6 | − | 9.1 ± 2.2 |
| 6 | 0.1 mg/kg SYN023 | 8 | 9.7 | 5.2 | 9.0 | 8.3 | 4.4 | 11 | 15 | 7.9 | − | 8.9 ± 3.4 |
| 7 | 0.3 mg/kg SYN023 | 9 | 4.7 | 11 | 4.2 | 7.5 | 9.4 | 19 | 5.3 | 2.8 | 3.8 | 7.5 ± 3.3 |
| 8 | 1 mg/kg SYN023 | 9 | 3.7 | 9.4 | 5.0 | 7.8 | 10. | 14 | 5.4 | 5.2 | 4.3 | 7.2 ± 3.3 |
| 9 | 20 IU/mL HRIG | 9 | 13 | 9.1 | 3.8 | 13 | 6.0 | 3.5 | 7.8 | 7.7 | 2.8 | 7.4 ± 3.9 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chao, T.-Y.; Zhang, S.-f.; Chen, L.; Tsao, E.; Rupprecht, C.E. In Vivo Efficacy of SYN023, an Anti-Rabies Monoclonal Antibody Cocktail, in Post-Exposure Prophylaxis Animal Models. Trop. Med. Infect. Dis. 2020, 5, 31. https://doi.org/10.3390/tropicalmed5010031
Chao T-Y, Zhang S-f, Chen L, Tsao E, Rupprecht CE. In Vivo Efficacy of SYN023, an Anti-Rabies Monoclonal Antibody Cocktail, in Post-Exposure Prophylaxis Animal Models. Tropical Medicine and Infectious Disease. 2020; 5(1):31. https://doi.org/10.3390/tropicalmed5010031
Chicago/Turabian StyleChao, Tzu-Yuan, Shou-feng Zhang, Li Chen, Eric Tsao, and Charles E. Rupprecht. 2020. "In Vivo Efficacy of SYN023, an Anti-Rabies Monoclonal Antibody Cocktail, in Post-Exposure Prophylaxis Animal Models" Tropical Medicine and Infectious Disease 5, no. 1: 31. https://doi.org/10.3390/tropicalmed5010031
APA StyleChao, T.-Y., Zhang, S.-f., Chen, L., Tsao, E., & Rupprecht, C. E. (2020). In Vivo Efficacy of SYN023, an Anti-Rabies Monoclonal Antibody Cocktail, in Post-Exposure Prophylaxis Animal Models. Tropical Medicine and Infectious Disease, 5(1), 31. https://doi.org/10.3390/tropicalmed5010031