(+)-Spectaline and Iso-6-Spectaline Induce a Possible Cross-Talk between Autophagy and Apoptosis in Trypanosoma brucei rhodesiense

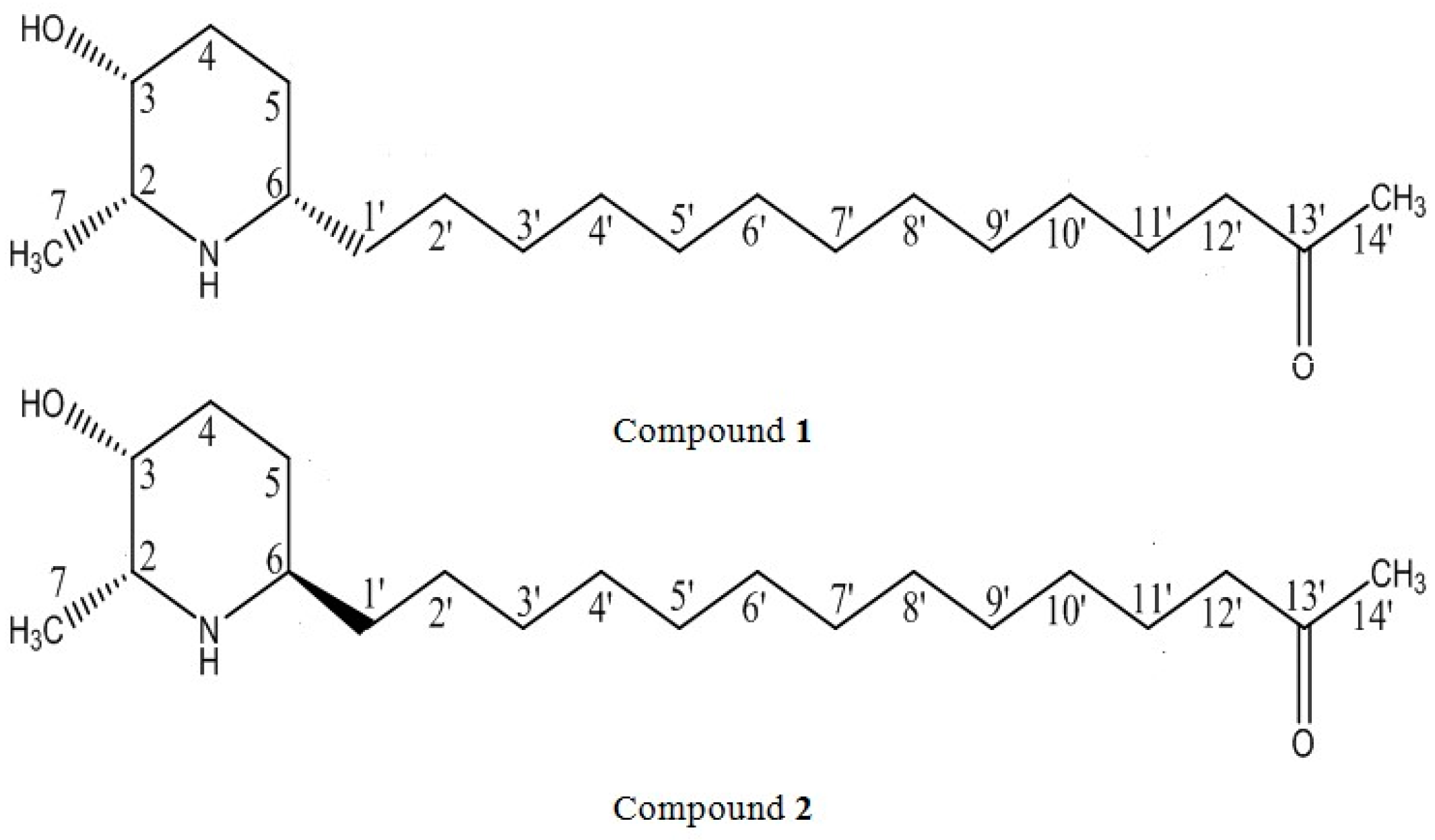{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. General Experimental Procedures
2.2. Measurement of Mitochondrial Membrane Potential
2.3. Detection of PS Exposure
2.4. DNA Fragmentation Analysis
2.5. Determination of Caspase-Like Protease Activity
2.6. Autophagy Assay
3. Results and Discussion
3.1. Measurement of Mitochondrial Membrane Potential
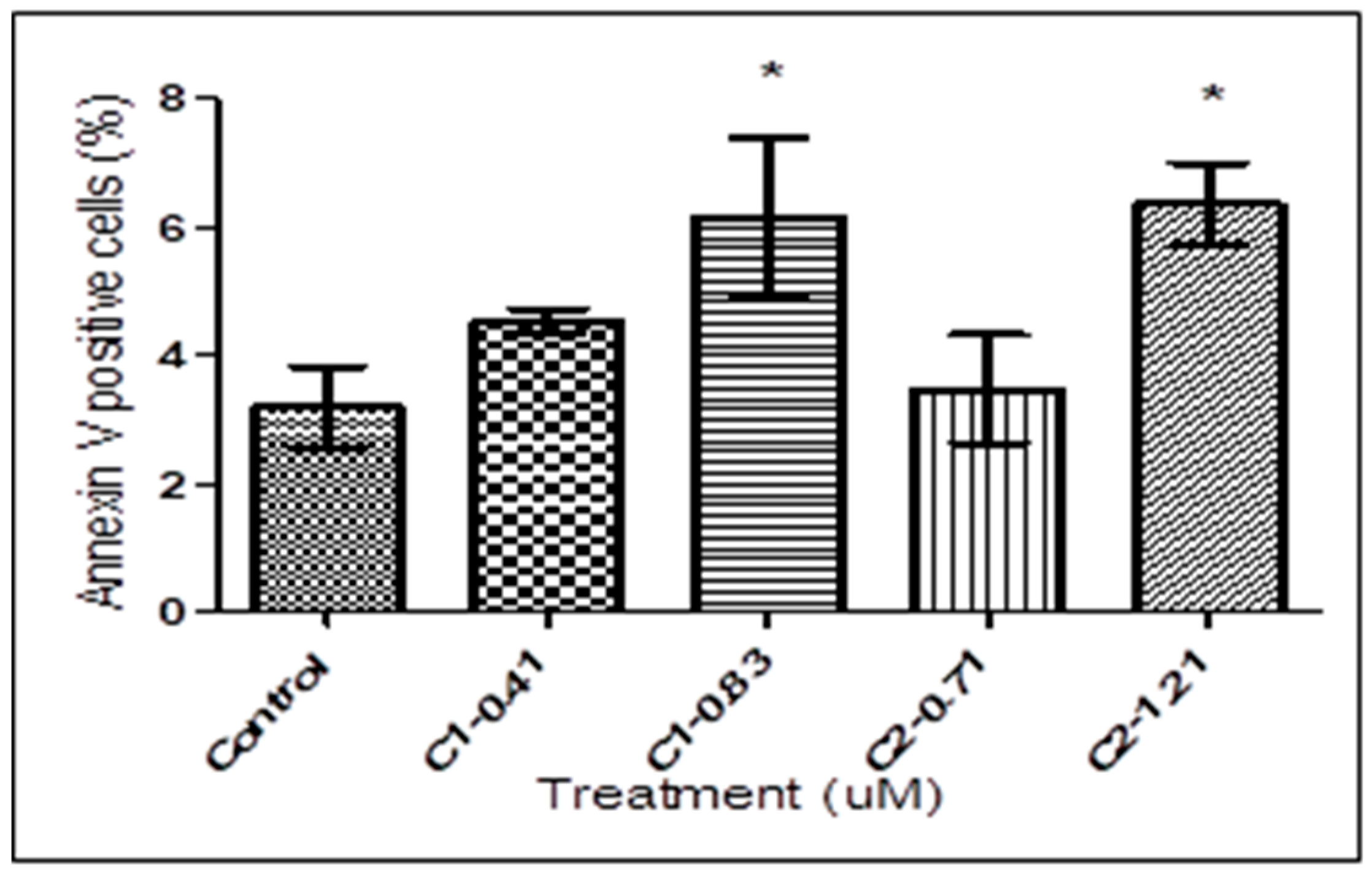
3.2. Detection of PS Exposure
3.3. DNA Fragmentation Analysis
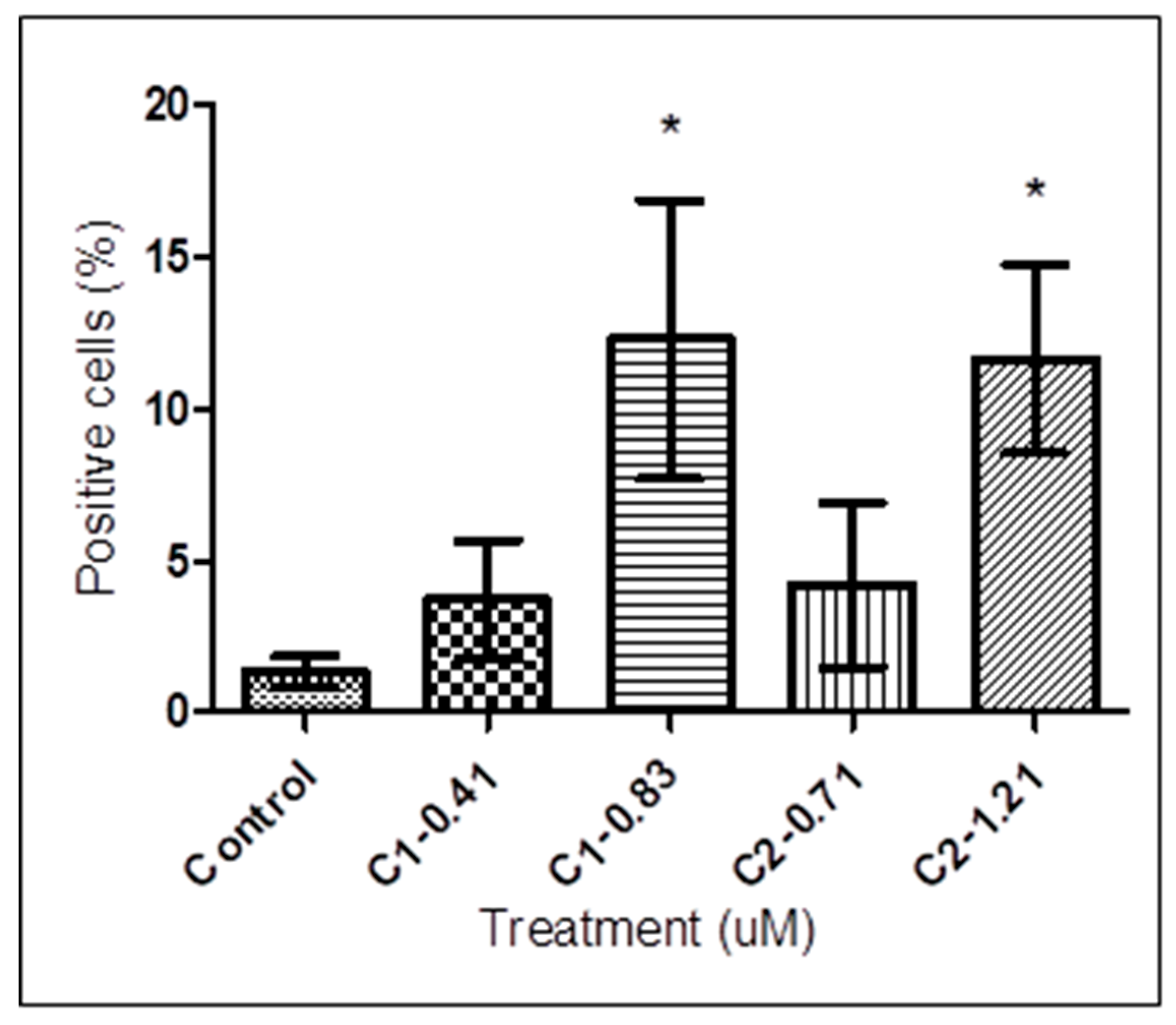
3.4. Determination of Caspase-Like Protease Activity
3.5. Autophagy Assay
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Bowling, T.; Mercer, L.; Don, R.; Jacobs, R.; Nare, B. Application of a resazurin-based high-throughput screening assay for the identification and progression of new treatments for human African trypanosomiasis. Int. J. Parasitol. Drugs Drug Resist. 2012, 2, 262–270. [Google Scholar] [CrossRef] [PubMed]
- Phillips, E.A.; Sexton, D.W.; Steverding, D. Bitter melon extract inhibits proliferation of Trypanosoma brucei bloodstream forms in vitro. Exp. Parasitol. 2013, 133, 353–356. [Google Scholar] [CrossRef] [PubMed]
- Steverding, D. The development of drugs for treatment of sleeping sickness: A historical review. Parasit. Vectors 2010, 3, 15. [Google Scholar] [CrossRef] [PubMed]
- Bacchi, C.J. Chemotherapy of human African trypanosomiasis. Interdiscip. Perspect. Infect. Dis. 2009, 2009, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Gehrig, S.; Efferth, T. Development of drug resistance in Trypanosoma brucei rhodesiense and Trypanosoma brucei gambiense. Treatment of human African trypanosomiasis with natural products (Review). Int. J. Mol. Med. 2008, 22, 411–419. [Google Scholar] [PubMed]
- Martyn, D.C.; Jones, D.C.; Fairlamb, A.H.; Clardy, J. High-throughput screening affords novel and selective trypanothione reductase inhibitors with anti-trypanosomal activity. Bioorg. Med. Chem. Lett. 2007, 17, 1280–1283. [Google Scholar] [CrossRef] [PubMed]
- Lim, K.T.; Amanah, A.; Chear, N.J.; Zahari, Z.; Zainuddin, Z.; Adenan, M.I. Inhibitory effects of (+)-spectaline and iso-6-spectaline from Senna spectabilis on the growth and ultrastructure of human-infective species Trypanosoma brucei rhodesiense bloodstream form. Exp. Parasitol. 2018, 184, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Jimenez, V.; Paredes, R.; Sosa, M.A.; Galanti, N. Natural programmed cell death in T. cruzi epimastigotes maintained in axenic cultures. J. Cell Biochem. 2008, 105, 688–698. [Google Scholar] [CrossRef] [PubMed]
- Rosenkranz, V.; Wink, M. Alkaloids Induce Programmed Cell Death in Bloodstream Forms of Trypanosomes (Trypanosoma b. brucei). Molecules 2008, 13, 2462–2473. [Google Scholar] [CrossRef] [PubMed]
- Helms, M.J.; Ambit, A.; Appleton, P.; Tetley, L.; Coombs, G.H.; Mottram, J.C. Bloodstream form Trypanosoma brucei depend upon multiple metacaspases associated with RAB11-positive endosomes. J. Cell Sci. 2006, 119, 1105–1117. [Google Scholar] [CrossRef][Green Version]
- Casanova, M.; Gonzalez, I.J.; Sprissler, C.; Zalila, H.; Dacher, M.; Basmaciyan, L.; Späth, G.F.; Azas, N.; Fasel, N. Implication of different domains of the Leishmania major metacaspase in cell death and autophagy. Cell Death Dis. 2015, 6, e1933. [Google Scholar] [CrossRef]
- Menna-Barreto, R.F.S.; Salomão, K.; Dantas, A.P.; Santa-Rita, R.M.; Soares, M.J.; Barbosa, H.S.; de Castro, S.L. Different cell death pathways induced by drugs in Trypanosoma cruzi: An ultrastructural study. Micron 2009, 40, 157–168. [Google Scholar] [CrossRef] [PubMed]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lim, K.T.; Yeoh, C.Y.; Zainuddin, Z.; Ilham Adenan, M. (+)-Spectaline and Iso-6-Spectaline Induce a Possible Cross-Talk between Autophagy and Apoptosis in Trypanosoma brucei rhodesiense. Trop. Med. Infect. Dis. 2019, 4, 98. https://doi.org/10.3390/tropicalmed4030098
Lim KT, Yeoh CY, Zainuddin Z, Ilham Adenan M. (+)-Spectaline and Iso-6-Spectaline Induce a Possible Cross-Talk between Autophagy and Apoptosis in Trypanosoma brucei rhodesiense. Tropical Medicine and Infectious Disease. 2019; 4(3):98. https://doi.org/10.3390/tropicalmed4030098
Chicago/Turabian StyleLim, Kah Tee, Chiann Ying Yeoh, Zafarina Zainuddin, and Mohd. Ilham Adenan. 2019. "(+)-Spectaline and Iso-6-Spectaline Induce a Possible Cross-Talk between Autophagy and Apoptosis in Trypanosoma brucei rhodesiense" Tropical Medicine and Infectious Disease 4, no. 3: 98. https://doi.org/10.3390/tropicalmed4030098
APA StyleLim, K. T., Yeoh, C. Y., Zainuddin, Z., & Ilham Adenan, M. (2019). (+)-Spectaline and Iso-6-Spectaline Induce a Possible Cross-Talk between Autophagy and Apoptosis in Trypanosoma brucei rhodesiense. Tropical Medicine and Infectious Disease, 4(3), 98. https://doi.org/10.3390/tropicalmed4030098