
The Chalcogen Exchange: The Replacement of Oxygen with Sulfur and Selenium to Boost the Activity of Natural Products

 ,
,  , , ,
, , ,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
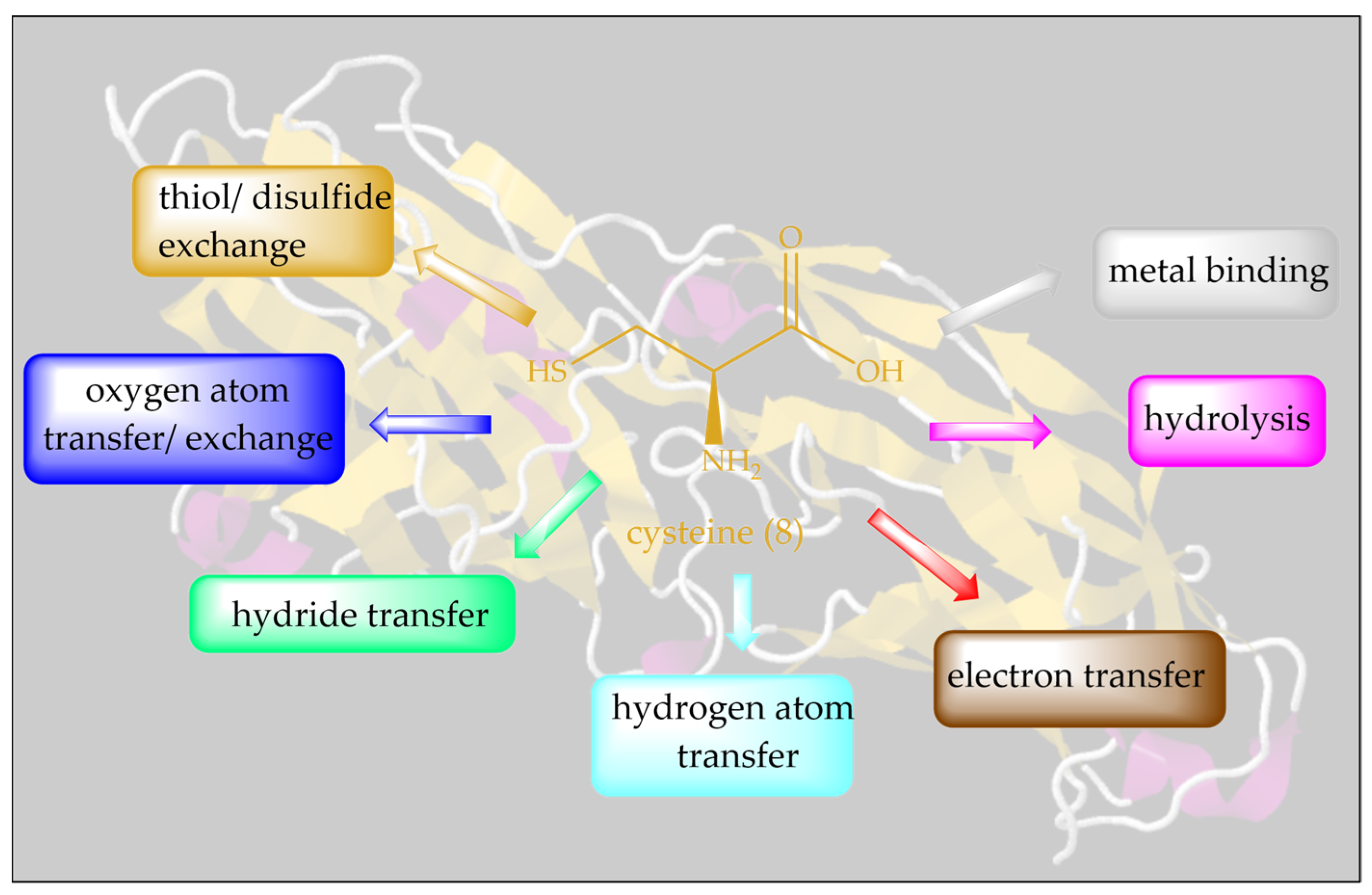
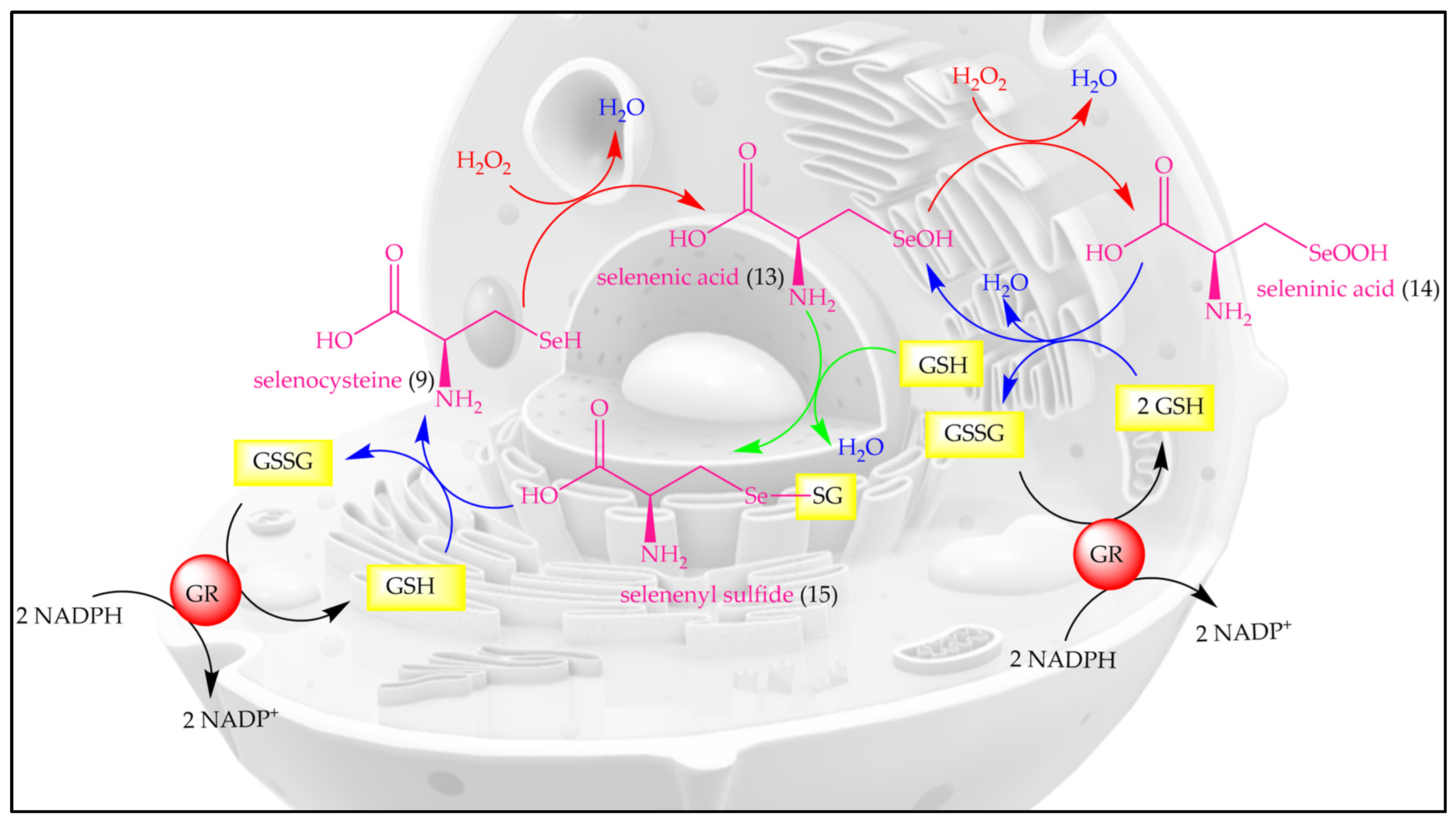
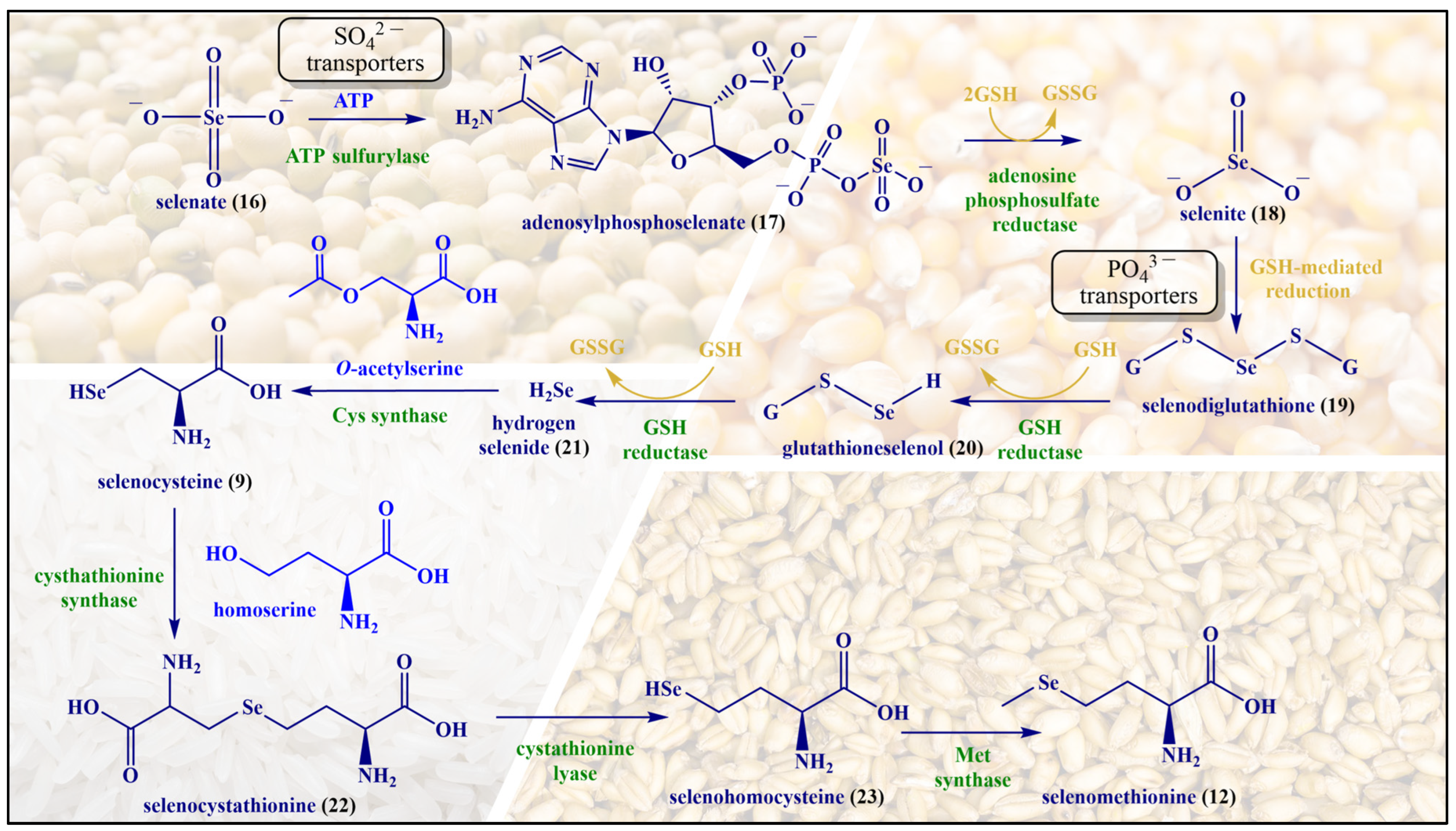
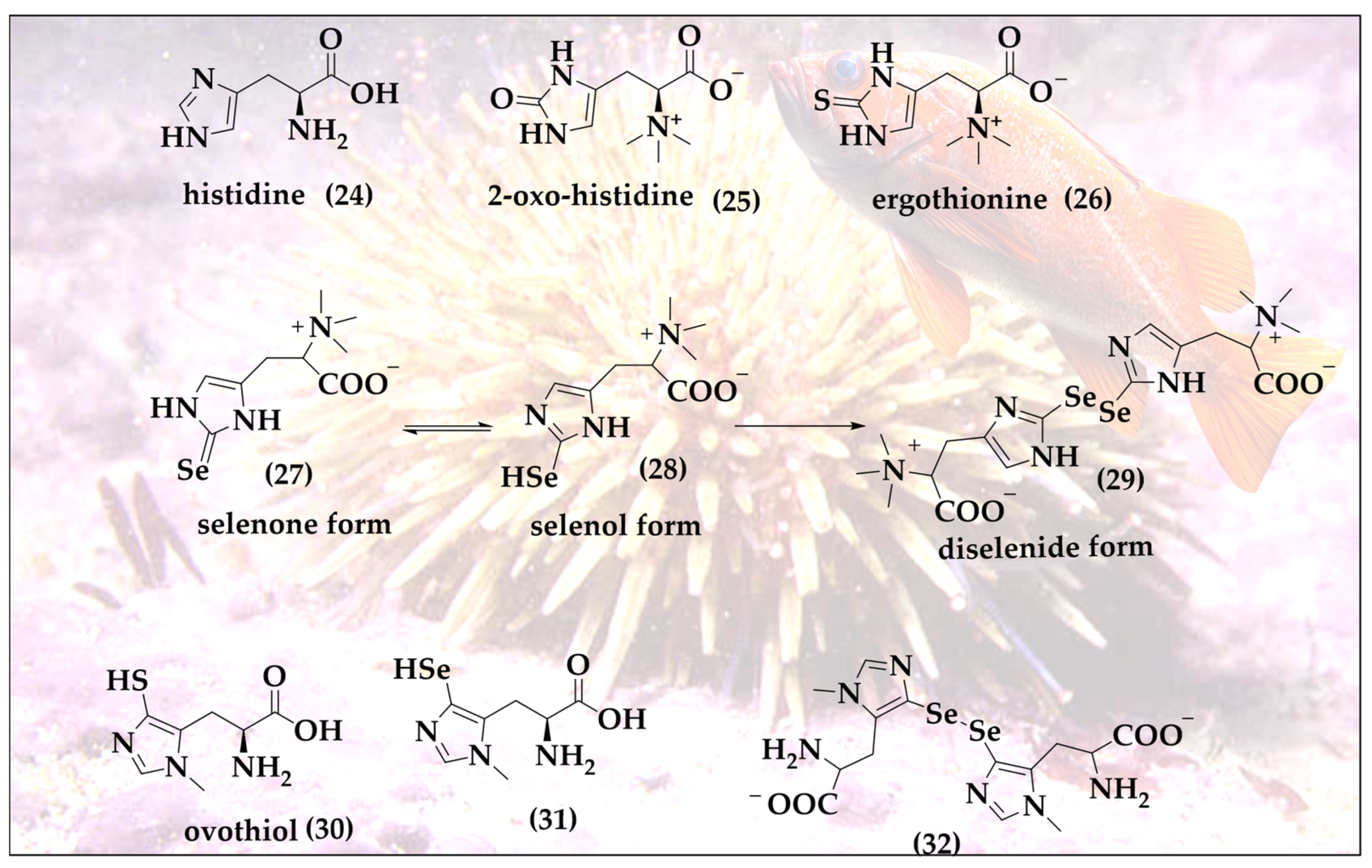
2. Inspired by Nature’s Symphony in Blue, Yellow and Pink
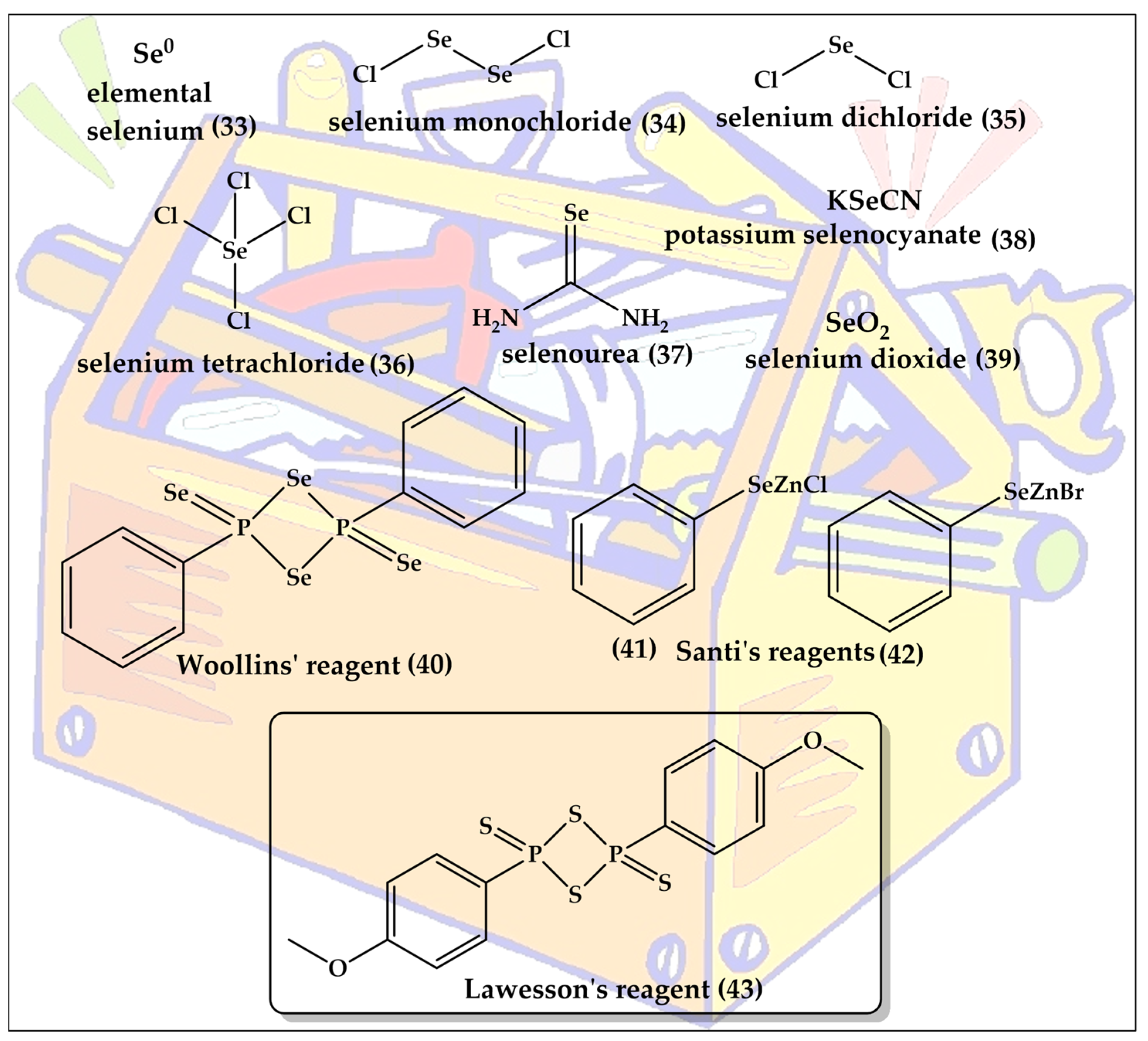
3. Toolkit
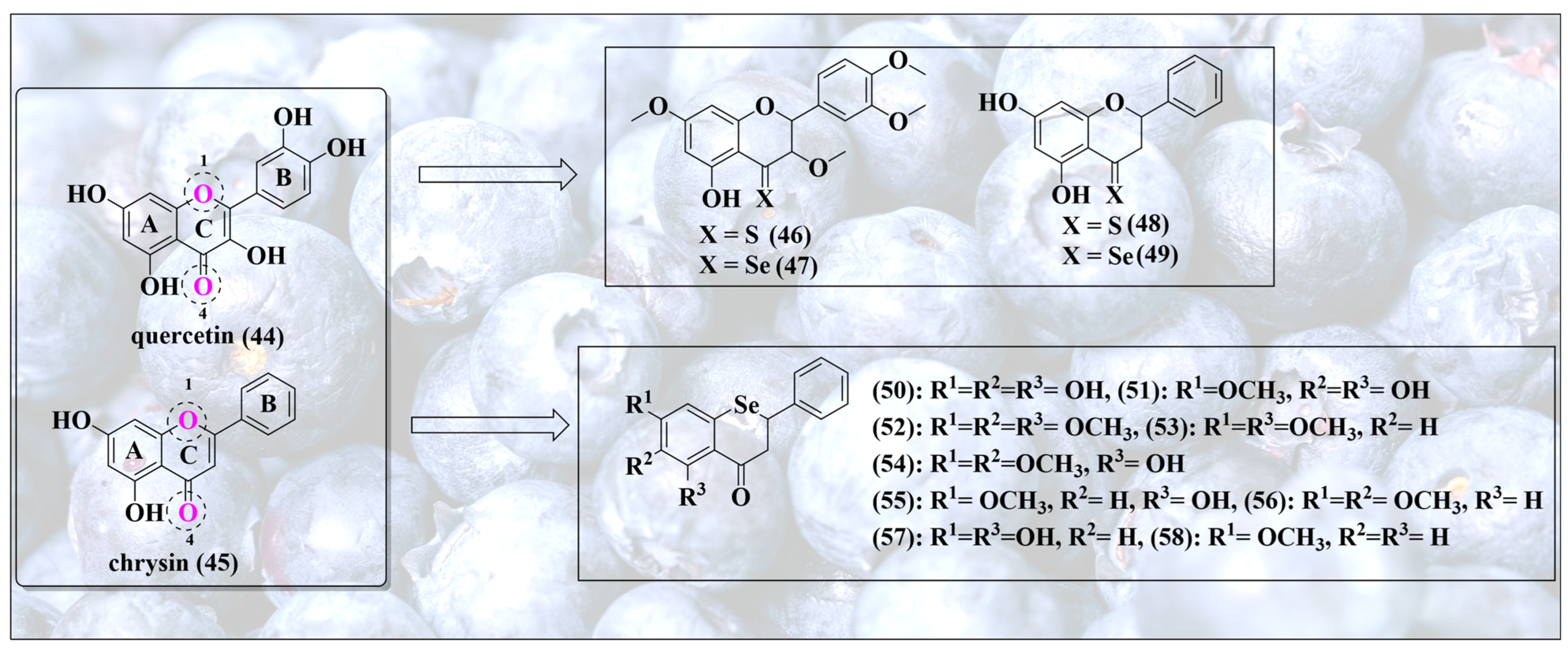
4. Blueprints and Blueberries
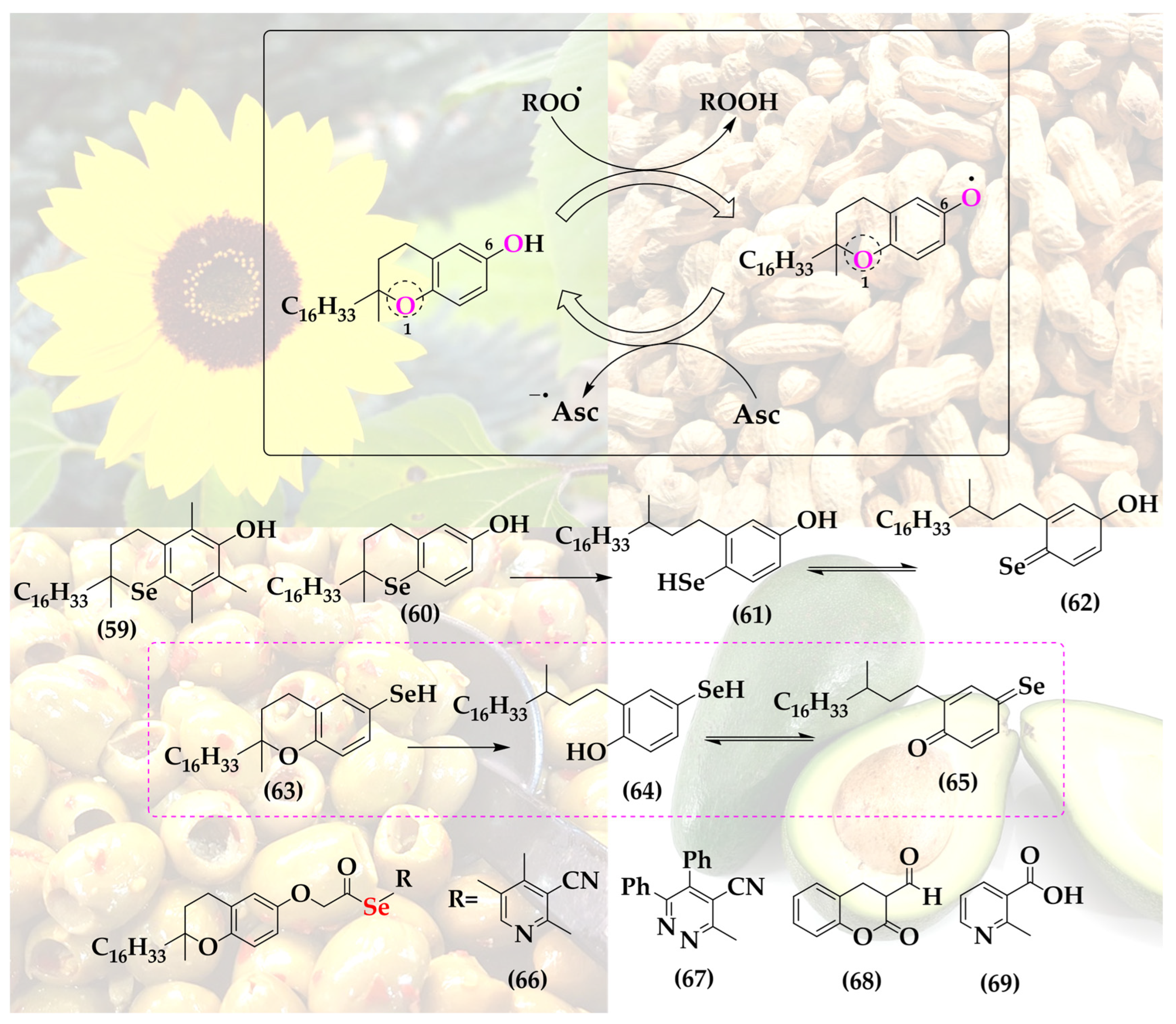
5. Selenium-Substituted Vitamin E
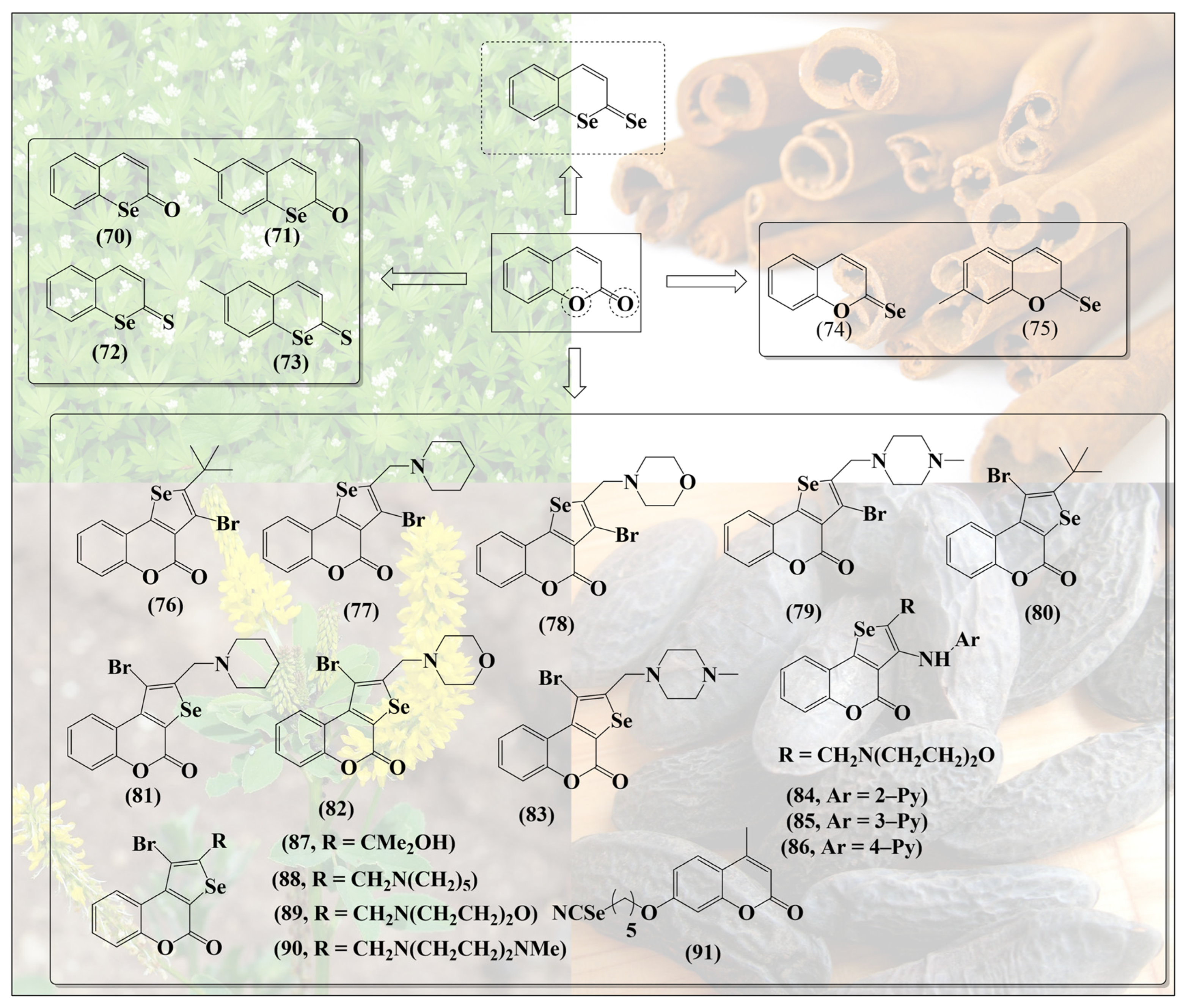
6. Selenium-Containing Coumarins
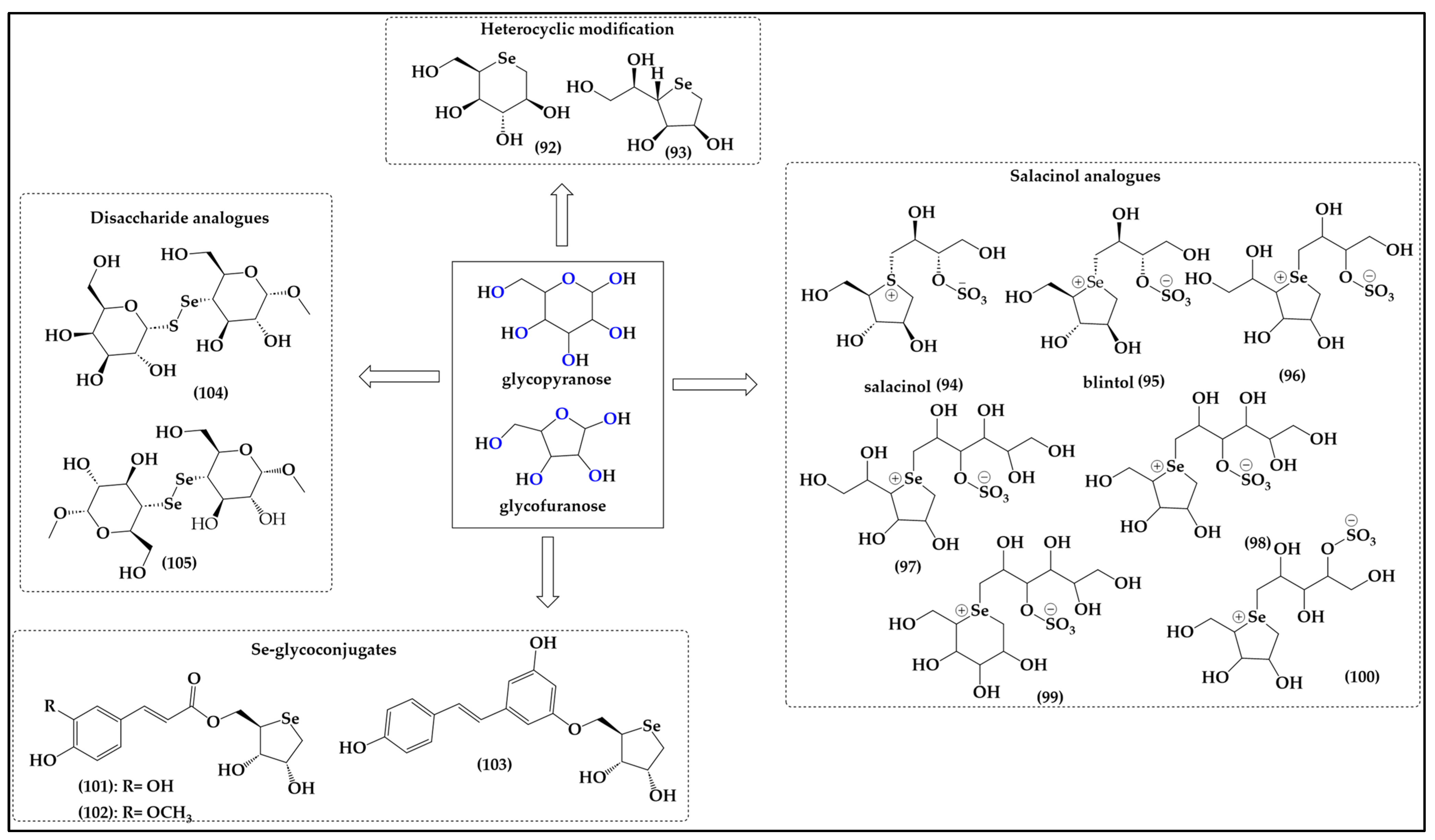
7. Selenium-Substituted Sugars
8. Selenium-Containing Esters of Polyphenolic Acids
9. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Číž, M.; Dvořáková, A.; Skočková, V.; Kubala, L. The Role of Dietary Phenolic Compounds in Epigenetic Modulation Involved in Inflammatory Processes. Antioxidants 2020, 9, 691. [Google Scholar] [CrossRef] [PubMed]
- Zujko, M.E.; Witkowska, A.M. Dietary Antioxidants and Chronic Diseases. Antioxidants 2023, 12, 362. [Google Scholar] [CrossRef] [PubMed]
- González-Manzano, S.; Dueñas, M. Applications of Natural Products in Food. Foods 2021, 10, 300. [Google Scholar] [CrossRef] [PubMed]
- Katidi, A.; Pavlopoulou, A.; Vlassopoulos, A.; Kapsokefalou, M. The Nutritional Composition of Natural and Organic Branded Food Products: A Cross-Sectional Analysis of the Greek Foodscape. Nutrients 2022, 14, 808. [Google Scholar] [CrossRef]
- Puri, V.; Nagpal, M.; Singh, I.; Singh, M.; Dhingra, G.A.; Huanbutta, K.; Dheer, D.; Sharma, A.; Sangnim, T. A Comprehensive Review on Nutraceuticals: Therapy Support and Formulation Challenges. Nutrients 2022, 14, 4637. [Google Scholar] [CrossRef]
- Chandra, S.; Saklani, S.; Kumar, P.; Kim, B.; Coutinho, H.D.M. Nutraceuticals: Pharmacologically Active Potent Dietary Supplements. BioMed Res. Int. 2022, 2022, 2051017. [Google Scholar] [CrossRef]
- Siddiqui, R.A.; Moghadasian, M.H. Nutraceuticals and Nutrition Supplements: Challenges and Opportunities. Nutrients 2020, 12, 1593. [Google Scholar] [CrossRef]
- Lü, J.-M.; Lin, P.H.; Yao, Q.; Chen, C. Chemical and Molecular Mechanisms of Antioxidants: Experimental Approaches and Model Systems. J. Cell. Mol. Med. 2010, 14, 840–860. [Google Scholar] [CrossRef]
- Bertelli, A.; Biagi, M.; Corsini, M.; Baini, G.; Cappellucci, G.; Miraldi, E. Polyphenols: From Theory to Practice. Foods 2021, 10, 2595. [Google Scholar] [CrossRef]
- Huang, J.; Xie, L.; Song, A.; Zhang, C. Selenium Status and Its Antioxidant Role in Metabolic Diseases. Oxid. Med. Cell. Longev. 2022, 2022, 7009863. [Google Scholar] [CrossRef]
- Bjørklund, G.; Shanaida, M.; Lysiuk, R.; Antonyak, H.; Klishch, I.; Shanaida, V.; Peana, M. Selenium: An Antioxidant with a Critical Role in Anti-Aging. Molecules 2022, 27, 6613. [Google Scholar] [CrossRef]
- Shimada, B.K.; Alfulaij, N.; Seale, L.A. The Impact of Selenium Deficiency on Cardiovascular Function. Int. J. Mol. Sci. 2021, 22, 10713. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Li, X.; Wei, Y. Selenium and Selenoproteins in Health. Biomolecules 2023, 13, 799. [Google Scholar] [CrossRef]
- Schomburg, L. Selenium Deficiency Due to Diet, Pregnancy, Severe Illness, or COVID-19—A Preventable Trigger for Autoimmune Disease. Int. J. Mol. Sci. 2021, 22, 8532. [Google Scholar] [CrossRef]
- Wang, F.; Sun, N.; Zeng, H.; Gao, Y.; Zhang, N.; Zhang, W. Selenium Deficiency Leads to Inflammation, Autophagy, Endoplasmic Reticulum Stress, Apoptosis and Contraction Abnormalities via Affecting Intestinal Flora in Intestinal Smooth Muscle of Mice. Front. Immunol. 2022, 13, 947655. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Wang, Z.; Gong, P.; Yao, W.; Ba, Q.; Wang, H. Review on the Health-Promoting Effect of Adequate Selenium Status. Front. Nutr. 2023, 10, 1136458. [Google Scholar] [CrossRef] [PubMed]
- Fonseca, S.F.; Lima, D.B.; Alves, D.; Jacob, R.G.; Perin, G.; Lenardão, E.J.; Savegnago, L. Synthesis, Characterization and Antioxidant Activity of Organoselenium and Organotellurium Compound Derivatives of Chrysin. New J. Chem. 2015, 39, 3043–3050. [Google Scholar] [CrossRef]
- Kharma, A.; Jacob, C.; Bozzi, Í.A.O.; Jardim, G.A.M.; Braga, A.L.; Salomão, K.; Gatto, C.C.; Silva, M.F.S.; Pessoa, C.; Stangier, M.; et al. Electrochemical Selenation/Cyclization of Quinones: A Rapid, Green and Efficient Access to Functionalized Trypanocidal and Antitumor Compounds. Eur. J. Org. Chem. 2020, 2020, 4474–4486. [Google Scholar] [CrossRef]
- Gonçalves, A.C.; Capelo, J.L.; Lodeiro, C.; Santos, A.A.D. A Selective Emissive Chromogenic and Fluorogenic Seleno-Coumarin Probe for Cu2+ Detection in Aprotic Media. Photochem. Photobiol. Sci. 2017, 16, 1174–1181. [Google Scholar] [CrossRef]
- Mániková, D.; Medvecová Letavayová, L.; Vlasáková, D.; Košík, P.; Castellucci Estevam, E.; Nasim, M.J.; Gruhlke, M.; Slusarenko, A.; Burkholz, T.; Jacob, C.; et al. Intracellular Diagnostics: Hunting for the Mode of Action of Redox-Modulating Selenium Compounds in Selected Model Systems. Molecules 2014, 19, 12258–12279. [Google Scholar] [CrossRef]
- Ozbek, N.; Baysal, A. Determination of Sulfur in Human Hair Using High Resolution Continuum Source Graphite Furnace Molecular Absorption Spectrometry and Its Correlation with Total Protein and Albumin. Spectrochim. Acta Part B At. Spectrosc. 2017, 130, 17–20. [Google Scholar] [CrossRef]
- Wang, P.; Chen, B.; Huang, Y.; Li, J.; Cao, D.; Chen, Z.; Li, J.; Ran, B.; Yang, J.; Wang, R.; et al. Selenium Intake and Multiple Health-Related Outcomes: An Umbrella Review of Meta-Analyses. Front. Nutr. 2023, 10, 1263853. [Google Scholar] [CrossRef]
- Rotruck, J.T.; Pope, A.L.; Ganther, H.E.; Swanson, A.B.; Hafeman, D.G.; Hoekstra, W.G. Selenium: Biochemical Role as a Component of Glutathione Peroxidase. Science 1973, 179, 588–590. [Google Scholar] [CrossRef]
- Flohe, L.; Günzler, W.; Schock, H.H. Glutathione Peroxidase: A Selenoenzyme. FEBS Lett. 1973, 32, 132–134. [Google Scholar] [CrossRef]
- Rotruck, J.; Swanson, A.; Pope, A.; Hoekstra, W.; Hafeman, D.; Ganther, H. Relationship of Selenium to GSH Peroxidase; Federation of American Societies for Experimental Biology: Bethesda, MD, USA, 1972; Volume 31, p. A691. [Google Scholar]
- Flohé, L.; Toppo, S.; Orian, L. The Glutathione Peroxidase Family: Discoveries and Mechanism. Free Rad. Biol. Med. 2022, 187, 113–122. [Google Scholar] [CrossRef]
- Jacob, C.; Maret, W.; Vallee, B.L. Selenium Redox Biochemistry of Zinc–Sulfur Coordination Sites in Proteins and Enzymes. Proc. Nat. Acad. Sci. USA 1999, 96, 1910–1914. [Google Scholar] [CrossRef] [PubMed]
- Nasim, M.J.; Zuraik, M.M.; Abdin, A.Y.; Ney, Y.; Jacob, C. Selenomethionine: A Pink Trojan Redox Horse with Implications in Aging and Various Age-Related Diseases. Antioxidants 2021, 10, 882. [Google Scholar] [CrossRef] [PubMed]
- Whanger, P.D. Selenocompounds in Plants and Animals and Their Biological Significance. J. Am. Coll. Nutr. 2002, 21, 223–232. [Google Scholar] [CrossRef]
- Weiss, J.F.; Srinivasan, V.; Kumar, K.S.; Landauer, M.R. Radioprotection by Metals: Selenium. Adv. Space Res. 1992, 12, 223–231. [Google Scholar] [CrossRef]
- Hariharan, S.; Dharmaraj, S. Selenium and Selenoproteins: It’s Role in Regulation of Inflammation. Inflammopharmacology 2020, 28, 667–695. [Google Scholar] [CrossRef]
- Hussein, R.A.; Ahmed, M.; Heinemann, S.H. Selenomethionine Mis-Incorporation and Redox-Dependent Voltage-Gated Sodium Channel Gain of Function. J. Neurochem. 2023, 167, 262–276. [Google Scholar] [CrossRef]
- Zhao, G.; Zhang, Y.; Tian, Y.; Huang, J.; Gao, P.; Zhao, Q.; Yang, Z. Antioxidant and Anti-Inflammatory Effects of Selenomethionine Promote Osteogenesis via Wnt/β-Catenin Pathway. Biochem. Biophys. Rep. 2023, 36, 101559. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, M.; Yamashita, Y.; Suzuki, T.; Kani, Y.; Mizusawa, N.; Imamura, S.; Takemoto, K.; Hara, T.; Hossain, M.A.; Yabu, T.; et al. Selenoneine, a Novel Selenium-Containing Compound, Mediates Detoxification Mechanisms against Methylmercury Accumulation and Toxicity in Zebrafish Embryo. Mar. Biotechnol. 2013, 15, 559–570. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, Y.; Yabu, T.; Yamashita, M. Discovery of the Strong Antioxidant Selenoneine in Tuna and Selenium Redox Metabolism. World J. Biol. Chem. 2010, 1, 144–150. [Google Scholar] [CrossRef]
- Alhasan, R.; Nasim, M.J.; Jacob, C.; Gaucher, C. Selenoneine: A Unique Reactive Selenium Species From the Blood of Tuna with Implications for Human Diseases. Curr. Pharmacol. Rep. 2019, 5, 163–173. [Google Scholar] [CrossRef]
- Yamashita, Y.; Yamashita, M. Identification of a Novel Selenium-Containing Compound, Selenoneine, as the Predominant Chemical Form of Organic Selenium in the Blood of Bluefin Tuna. J. Biol. Chem. 2010, 285, 18134–18138. [Google Scholar] [CrossRef]
- Nachimuthu, S.; Kandasamy, R.; Ponnusamy, R.; Deruiter, J.; Dhanasekaran, M.; Thilagar, S. L-Ergothioneine: A Potential Bioactive Compound from Edible Mushrooms. In Medicinal Mushrooms: Recent Progress in Research and Development; Agrawal, D.C., Dhanasekaran, M., Eds.; Springer: Singapore, 2019; pp. 391–407. ISBN 9789811363825. [Google Scholar] [CrossRef]
- Fahey, R.C. Novel Thiols of Prokaryotes. Annu. Rev. Microbiol. 2001, 55, 333–356. [Google Scholar] [CrossRef]
- El Hanafi, K.; Pedrero, Z.; Ouerdane, L.; Marchán Moreno, C.; Queipo-Abad, S.; Bueno, M.; Pannier, F.; Corns, W.T.; Cherel, Y.; Bustamante, P.; et al. First Time Identification of Selenoneine in Seabirds and Its Potential Role in Mercury Detoxification. Environ. Sci. Technol. 2022, 56, 3288–3298. [Google Scholar] [CrossRef]
- Kroepfl, N.; Francesconi, K.A.; Schwerdtle, T.; Kuehnelt, D. Selenoneine and Ergothioneine in Human Blood Cells Determined Simultaneously by HPLC/ICP-QQQ-MS. J. Anal. At. Spectrom. 2019, 34, 127–134. [Google Scholar] [CrossRef]
- Yamashita, M.; Yamashita, Y.; Ando, T.; Wakamiya, J.; Akiba, S. Identification and Determination of Selenoneine, 2-Selenyl-Nα,Nα,Nα -Trimethyl-l-Histidine, as the Major Organic Selenium in Blood Cells in a Fish-Eating Population on Remote Japanese Islands. Biol. Trace Elem. Res. 2013, 156, 36–44. [Google Scholar] [CrossRef]
- Little, M.; Achouba, A.; Dumas, P.; Ouellet, N.; Ayotte, P.; Lemire, M. Determinants of Selenoneine Concentration in Red Blood Cells of Inuit from Nunavik (Northern Québec, Canada). Environ. Int. 2019, 127, 243–252. [Google Scholar] [CrossRef] [PubMed]
- Milito, A.; Cocurullo, M.; Columbro, A.; Nonnis, S.; Tedeschi, G.; Castellano, I.; Arnone, M.I.; Palumbo, A. Ovothiol Ensures the Correct Developmental Programme of the Sea Urchin Paracentrotus Lividus Embryo. Open Biol. 2022, 12, 210262. [Google Scholar] [CrossRef] [PubMed]
- Alhasan, R.; Martins, G.M.; de Castro, P.P.; Saleem, R.S.Z.; Zaiter, A.; Fries-Raeth, I.; Kleinclauss, A.; Perrin-Sarrado, C.; Chaimbault, P.; da Silva Júnior, E.N.; et al. Selenoneine-Inspired Selenohydantoins with Glutathione Peroxidase-like Activity. Bioorganic Med. Chem. 2023, 94, 117479. [Google Scholar] [CrossRef]
- Abdulnabi, Z.A.; Al-doghachi, F.A.J.; Abdulsahib, H.T. Synthesis, Characterization and Thermogravimetric Study of Some Metal Complexes of Selenazone Ligand Nanoparticles Analogue of Dithizone. Indones. J. Chem. 2021, 21, 1231–1243. [Google Scholar] [CrossRef]
- Kaya, B.; Gholam Azad, M.; Suleymanoglu, M.; Harmer, J.R.; Wijesinghe, T.P.; Richardson, V.; Zhao, X.; Bernhardt, P.V.; Dharmasivam, M.; Richardson, D.R. Isosteric Replacement of Sulfur to Selenium in a Thiosemicarbazone: Promotion of Zn(II) Complex Dissociation and Transmetalation to Augment Anticancer Efficacy. J. Med. Chem. 2024, 67, 12155–12183. [Google Scholar] [CrossRef] [PubMed]
- Khatoon, H.; Abdulmalek, E. A Focused Review of Synthetic Applications of Lawesson’s Reagent in Organic Synthesis. Molecules 2021, 26, 6937. [Google Scholar] [CrossRef]
- Ascherl, L.; Nordheider, A.; Arachchige, K.S.A.; Cordes, D.B.; Karaghiosoff, K.; Bühl, M.; Slawin, A.M.Z.; Woollins, J.D. The Activation of Woollins’ Reagent. Isolation of Pyridine Stabilised PhPSe2. Chem. Commun. 2014, 50, 6214–6216. [Google Scholar] [CrossRef]
- Domínguez-Álvarez, E.; Rácz, B.; Marć, M.A.; Nasim, M.J.; Szemerédi, N.; Viktorová, J.; Jacob, C.; Spengler, G. Selenium and Tellurium in the Development of Novel Small Molecules and Nanoparticles as Cancer Multidrug Resistance Reversal Agents. Drug Resist. Updat. 2022, 63, 100844. [Google Scholar] [CrossRef]
- Spengler, G.; Gajdács, M.; Marć, M.A.; Domínguez-Álvarez, E.; Sanmartín, C. Organoselenium Compounds as Novel Adjuvants of Chemotherapy Drugs-A Promising Approach to Fight Cancer Drug Resistance. Molecules 2019, 24, 336. [Google Scholar] [CrossRef]
- Sonego, J.M.; de Diego, S.I.; Szajnman, S.H.; Gallo-Rodriguez, C.; Rodriguez, J.B. Organoselenium Compounds: Chemistry and Applications in Organic Synthesis. Chem. Eur. J. 2023, 29, e202300030. [Google Scholar] [CrossRef]
- Hou, W.; Dong, H.; Zhang, X.; Wang, Y.; Su, L.; Xu, H. Selenium as an Emerging Versatile Player in Heterocycles and Natural Products Modification. Drug Discov. Today 2022, 27, 2268–2277. [Google Scholar] [CrossRef] [PubMed]
- Santi, C.; Santoro, S.; Battistelli, B.; Testaferri, L.; Tiecco, M. Preparation of the First Bench-Stable Phenyl Selenolate: An Interesting “On Water” Nucleophilic Reagent. Eur. J. Org. Chem. 2008, 2008, 5387–5390. [Google Scholar] [CrossRef]
- Lai, S.; Liang, X.; Zeng, Q. Recent Progress in Synthesis and Application of Chiral Organoselenium Compounds. Chem. Eur. J. 2024, 30, e202304067. [Google Scholar] [CrossRef]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An Overview. J. Nutr. Sci. 2016, 5, e47. [Google Scholar] [CrossRef]
- Chen, S.; Wang, X.; Cheng, Y.; Gao, H.; Chen, X. A Review of Classification, Biosynthesis, Biological Activities and Potential Applications of Flavonoids. Molecules 2023, 28, 4982. [Google Scholar] [CrossRef] [PubMed]
- Martins, I.L.; Charneira, C.; Gandin, V.; Ferreira da Silva, J.L.; Justino, G.C.; Telo, J.P.; Vieira, A.J.S.C.; Marzano, C.; Antunes, A.M.M. Selenium-Containing Chrysin and Quercetin Derivatives: Attractive Scaffolds for Cancer Therapy. J. Med. Chem. 2015, 58, 4250–4265. [Google Scholar] [CrossRef]
- Merino-Montiel, P.; Maza, S.; Martos, S.; López, Ó.; Maya, I.; Fernández-Bolaños, J.G. Synthesis and Antioxidant Activity of O-Alkyl Selenocarbamates, Selenoureas and Selenohydantoins. Eur. J. Pharm. Sci. 2013, 48, 582–592. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.-S.; Kim, D.-M.; Kim, Y.-J.; Yang, S.; Lee, K.-T.; Ryu, J.H.; Jeong, J.-H. Synthesis and Evaluation of Neuroprotective Selenoflavanones. Int. J. Mol. Sci. 2015, 16, 29574–29582. [Google Scholar] [CrossRef]
- Choi, Y.S.; Kim, Y.J.; Lee, J.Y.; Lee, J.; Jeong, J.H. Synthesis and Evaluation of Selenoflavones That Have Potential Neuroprotective Effects. Heterocycles 2014, 89, 2794–2805. [Google Scholar] [CrossRef]
- Niki, E. Evidence for Beneficial Effects of Vitamin E. Korean J. Intern. Med. 2015, 30, 571–579. [Google Scholar] [CrossRef]
- Gamna, F.; Spriano, S. Vitamin E: A Review of Its Application and Methods of Detection When Combined with Implant Biomaterials. Materials 2021, 14, 3691. [Google Scholar] [CrossRef] [PubMed]
- Al-Maharik, N.; Engman, L.; Malmström, J.; Schiesser, C.H. Intramolecular Homolytic Substitution at Selenium: Synthesis of Novel Selenium-Containing Vitamin E Analogues. J. Org. Chem. 2001, 66, 6286–6290. [Google Scholar] [CrossRef] [PubMed]
- Shanks, D.; Amorati, R.; Fumo, M.G.; Pedulli, G.F.; Valgimigli, L.; Engman, L. Synthesis and Antioxidant Profile of All-Rac-α-Selenotocopherol. J. Org. Chem. 2006, 71, 1033–1038. [Google Scholar] [CrossRef]
- Design, Synthesis and Cytotoxic Activity of Vitamin E Bearing Selenium Compounds against Human Breast Cancer Cell Line (MCF-7). Phosphorus Sulfur. Silicon Relat. Elem. 2017, 192, 1114–1118. [CrossRef]
- Reagan-Shaw, S.; Nihal, M.; Ahsan, H.; Mukhtar, H.; Ahmad, N. Combination of Vitamin E and Selenium Causes an Induction of Apoptosis of Human Prostate Cancer Cells by Enhancing Bax/Bcl-2 Ratio. Prostate 2008, 68, 1624–1634. [Google Scholar] [CrossRef]
- Angeli, A.; Trallori, E.; Carta, F.; Di Cesare Mannelli, L.; Ghelardini, C.; Supuran, C.T. Heterocoumarins Are Selective Carbonic Anhydrase IX and XII Inhibitors with Cytotoxic Effects against Cancer Cells Lines. ACS Med. Chem. Lett. 2018, 9, 947–951. [Google Scholar] [CrossRef] [PubMed]
- Murai, T.; Yoshida, A.; Mizutani, T.; Kubuki, H.; Yamaguchi, K.; Maruyama, T.; Shibahara, F. The First Selenium Isologues of 2-Pyrones and Coumarins: Synthesis, Structures, and Reactions. Chem. Lett. 2017, 46, 1017–1019. [Google Scholar] [CrossRef]
- Domracheva, I.; Kanepe-Lapsa, I.; Jackevica, L.; Vasiljeva, J.; Arsenyan, P. Selenopheno Quinolinones and Coumarins Promote Cancer Cell Apoptosis by ROS Depletion and Caspase-7 Activation. Life Sci. 2017, 186, 92–101. [Google Scholar] [CrossRef]
- Arsenyan, P.; Vasiljeva, J.; Shestakova, I.; Domracheva, I.; Jaschenko, E.; Romanchikova, N.; Leonchiks, A.; Rudevica, Z.; Belyakov, S. Selenopheno[3,2-c]- and [2,3-c]Coumarins: Synthesis, Cytotoxicity, Angiogenesis Inhibition, and Antioxidant Properties. Comptes Rendus Chim. 2015, 18, 399–409. [Google Scholar] [CrossRef]
- Patra, A.R.; Roy, S.S.; Basu, A.; Bhuniya, A.; Bhattacharjee, A.; Hajra, S.; Hossain Sk, U.; Baral, R.; Bhattacharya, S. Design and synthesis of coumarin-based organoselenium as a new hit for myeloprotection and synergistic therapeutic efficacy in adjuvant therapy. Sci. Rep. 2018, 8, 2194. [Google Scholar] [CrossRef]
- Davies, M.J.; Schiesser, C.H. 1,4-Anhydro-4-Seleno-D-Talitol (SeTal): A Remarkable Selenium-Containing Therapeutic Molecule. New J. Chem. 2019, 43, 9759–9765. [Google Scholar] [CrossRef]
- Storkey, C.; Pattison, D.I.; White, J.M.; Schiesser, C.H.; Davies, M.J. Preventing Protein Oxidation with Sugars: Scavenging of Hypohalous Acids by 5-Selenopyranose and 4-Selenofuranose Derivatives. Chem. Res. Toxicol. 2012, 25, 2589–2599. [Google Scholar] [CrossRef]
- Zacharias, T.; Flouda, K.; Jepps, T.A.; Gammelgaard, B.; Schiesser, C.H.; Davies, M.J. Effects of a Novel Selenium Substituted-Sugar (1,4-Anhydro-4-Seleno-d-Talitol, SeTal) on Human Coronary Artery Cell Lines and Mouse Aortic Rings. Biochem. Pharmacol. 2020, 173, 113631. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Pinto, B.M. Efficient Synthesis of the Glucosidase Inhibitor Blintol, the Selenium Analogue of the Naturally Occurring Glycosidase Inhibitor Salacinol. J. Org. Chem. 2005, 70, 753–755. [Google Scholar] [CrossRef]
- Liu, H.; Pinto, B.M. Design and Synthesis of Selenonium and Sulfonium Ions Related to the Naturally Occurring Glucosidase Inhibitor Salacinol. Can. J. Chem. 2006, 84, 1351–1362. [Google Scholar] [CrossRef]
- Liu, H.; Pinto, B.M. Synthesis of Zwitterionic Selenonium and Sulfonium Sulfates from D-Mannose as Potential Glycosidase Inhibitors. Can. J. Chem. 2006, 84, 497–505. [Google Scholar] [CrossRef]
- Morikawa, T.; Ninomiya, K.; Tanabe, G.; Matsuda, H.; Yoshikawa, M.; Muraoka, O. A Review of Antidiabetic Active Thiosugar Sulfoniums, Salacinol and Neokotalanol, from Plants of the Genus Salacia. J. Nat. Med. 2021, 75, 449–466. [Google Scholar] [CrossRef]
- Morikawa, T.; Akaki, J.; Ninomiya, K.; Kinouchi, E.; Tanabe, G.; Pongpiriyadacha, Y.; Yoshikawa, M.; Muraoka, O. Salacinol and Related Analogs: New Leads for Type 2 Diabetes Therapeutic Candidates from the Thai Traditional Natural Medicine Salacia Chinensis. Nutrients 2015, 7, 1480–1493. [Google Scholar] [CrossRef]
- Nasi, R.; Sim, L.; Rose, D.R.; Pinto, B.M. New Chain-Extended Analogues of Salacinol and Blintol and Their Glycosidase Inhibitory Activities. Mapping the Active-Site Requirements of Human Maltase Glucoamylase. J. Org. Chem. 2007, 72, 180–186. [Google Scholar] [CrossRef]
- Liu, H.; Nasi, R.; Jayakanthan, K.; Sim, L.; Heipel, H.; Rose, D.R.; Pinto, B.M. New Synthetic Routes to Chain-Extended Selenium, Sulfur, and Nitrogen Analogues of the Naturally Occurring Glucosidase Inhibitor Salacinol and Their Inhibitory Activities against Recombinant Human Maltase Glucoamylase. J. Org. Chem. 2007, 72, 6562–6572. [Google Scholar] [CrossRef]
- Nasi, R.; Sim, L.; Rose, D.R.; Pinto, B.M. Synthesis and Glycosidase Inhibitory Activities of Chain-Modified Analogues of the Glycosidase Inhibitors Salacinol and Blintol. Carb. Res. 2007, 342, 1888–1894. [Google Scholar] [CrossRef] [PubMed]
- Serpico, L.; De Nisco, M.; Cermola, F.; Manfra, M.; Pedatella, S. Stereoselective Synthesis of Selenium-Containing Glycoconjugates via the Mitsunobu Reaction. Molecules 2021, 26, 2541. [Google Scholar] [CrossRef]
- Cimmino, G.; De Nisco, M.; Alonso, C.; Gravina, C.; Piscopo, V.; Lemos, R.; Coderch, L.; Piccolella, S.; Pacifico, S.; Pedatella, S. Novel Synthesized Seleno-Glycoconjugates as Cosmeceutical Ingredients: Antioxidant Activity and in Vitro Skin Permeation. Eur. J. Med. Chem. Rep. 2024, 12, 100240. [Google Scholar] [CrossRef]
- Serpico, L.; Dello Iacono, S.; De Stefano, L.; De Martino, S.; Battisti, M.; Dardano, P.; Pedatella, S.; De Nisco, M. PH-Sensitive Release of Antioxidant Se-Glycoconjugates through a Flexible Polymeric Patch. Eur. Polym. J. 2022, 178, 111486. [Google Scholar] [CrossRef]
- Chakka, N.; Johnston, B.D.; Pinto, B.M. Synthesis and Conformational Analysis of Disaccharide Analogues Containing Disulfide and Selenosulfide Functionalities in the Interglycosidic Linkages. Can. J. Chem. 2005, 83, 929–936. [Google Scholar] [CrossRef]
- Marchiosi, R.; dos Santos, W.D.; Constantin, R.P.; de Lima, R.B.; Soares, A.R.; Finger-Teixeira, A.; Mota, T.R.; de Oliveira, D.M.; Foletto-Felipe, M.d.P.; Abrahão, J.; et al. Biosynthesis and Metabolic Actions of Simple Phenolic Acids in Plants. Phytochem. Rev. 2020, 19, 865–906. [Google Scholar] [CrossRef]
- Liu, Y.; Deng, J.; Zhao, T.; Yang, X.; Zhang, J.; Yang, H. Bioavailability and Mechanisms of Dietary Polyphenols Affected by Non-Thermal Processing Technology in Fruits and Vegetables. Curr. Res. Food Sci. 2024, 8, 100715. [Google Scholar] [CrossRef]
- Wang, M.; Zhu, X.; Wang, K.; Lu, C.; Luo, M.; Shan, T.; Zhang, Z. A Wheat Caffeic Acid 3-O-Methyltransferase TaCOMT-3D Positively Contributes to Both Resistance to Sharp Eyespot Disease and Stem Mechanical Strength. Sci. Rep. 2018, 8, 6543. [Google Scholar] [CrossRef]
- Mughal, A.; Jabeen, N.; Ashraf, K.; Sultan, K.; Farhan, M.; Hussain, M.I.; Deng, G.; Alsudays, I.M.; Saleh, M.A.; Tariq, S.; et al. Exploring the Role of Caffeic Acid in Mitigating Abiotic Stresses in Plants: A Review. Plant Stress. 2024, 12, 100487. [Google Scholar] [CrossRef]
- Narnoliya, L.K.; Sangwan, N.; Jadaun, J.S.; Bansal, S.; Sangwan, R.S. Defining the Role of a Caffeic Acid 3-O-Methyltransferase from Azadirachta Indica Fruits in the Biosynthesis of Ferulic Acid through Heterologous over-Expression in Ocimum Species and Withania Somnifera. Planta 2021, 253, 20. [Google Scholar] [CrossRef]
- Zafar-ul-Hye, M.; Akbar, M.N.; Iftikhar, Y.; Abbas, M.; Zahid, A.; Fahad, S.; Datta, R.; Ali, M.; Elgorban, A.M.; Ansari, M.J.; et al. Rhizobacteria Inoculation and Caffeic Acid Alleviated Drought Stress in Lentil Plants. Sustainability 2021, 13, 9603. [Google Scholar] [CrossRef]
- Bešlo, D.; Golubić, N.; Rastija, V.; Agić, D.; Karnaš, M.; Šubarić, D.; Lučić, B. Antioxidant Activity, Metabolism, and Bioavailability of Polyphenols in the Diet of Animals. Antioxidants 2023, 12, 1141. [Google Scholar] [CrossRef] [PubMed]
- Zduńska, K.; Dana, A.; Kolodziejczak, A.; Rotsztejn, H. Antioxidant Properties of Ferulic Acid and Its Possible Application. Ski. Pharmacol. Physiol. 2018, 31, 332–336. [Google Scholar] [CrossRef]
- Magnani, C.; Isaac, V.L.B.; Correa, M.A.; Salgado, H.R.N. Caffeic Acid: A Review of Its Potential Use in Medications and Cosmetics. Anal. Methods 2014, 6, 3203–3210. [Google Scholar] [CrossRef]
- Liu, Y.-M.; Shen, J.-D.; Xu, L.-P.; Li, H.-B.; Li, Y.-C.; Yi, L.-T. Ferulic Acid Inhibits Neuro-Inflammation in Mice Exposed to Chronic Unpredictable Mild Stress. Int. Immunopharmacol. 2017, 45, 128–134. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Xue, F.; Han, C.; Yang, H.; Han, L.; Li, K.; Li, J.; Xu, Q.; Li, Z.; Yuan, B.; et al. Ferulic Acid Ameliorated Placental Inflammation and Apoptosis in Rat with Preeclampsia. Clin. Exp. Hypertens. 2019, 41, 524–530. [Google Scholar] [CrossRef]
- Huang, D.-W.; Shen, S.-C.; Wu, J.S.-B. Effects of Caffeic Acid and Cinnamic Acid on Glucose Uptake in Insulin-Resistant Mouse Hepatocytes. J. Agric. Food Chem. 2009, 57, 7687–7692. [Google Scholar] [CrossRef]
- Azhar, M.K.; Anwar, S.; Hasan, G.M.; Shamsi, A.; Islam, A.; Parvez, S.; Hassan, M.I. Comprehensive Insights into Biological Roles of Rosmarinic Acid: Implications in Diabetes, Cancer and Neurodegenerative Diseases. Nutrients 2023, 15, 4297. [Google Scholar] [CrossRef]
- Alam, M.; Ashraf, G.M.; Sheikh, K.; Khan, A.; Ali, S.; Ansari, M.M.; Adnan, M.; Pasupuleti, V.R.; Hassan, M.I. Potential Therapeutic Implications of Caffeic Acid in Cancer Signaling: Past, Present, and Future. Front. Pharmacol. 2022, 13, 845871. [Google Scholar] [CrossRef]
- Chrrng, J.-M.; Shieh, D.-E.; Chiang, W.; Chang, M.-Y.; Chiang, L.-C. Chemopreventive Effects of Minor Dietary Constituents in Common Foods on Human Cancer Cells. Biosci. Biotech. Biochem. 2007, 71, 1500–1504. [Google Scholar] [CrossRef]
- Jaganathan, S.K. Growth Inhibition by Caffeic Acid, One of the Phenolic Constituents of Honey, in HCT 15 Colon Cancer Cells. Sci. World J. 2012, 2012, 372345. [Google Scholar] [CrossRef] [PubMed]
- Tsai, Y.-L.; Chiou, S.-Y.; Chan, K.-C.; Sung, J.-M.; Lin, S.-D. Caffeic Acid Derivatives, Total Phenols, Antioxidant and Antimutagenic Activities of Echinacea purpurea Flower Extracts. LWT-Food Sci. Technol. 2012, 46, 169–176. [Google Scholar] [CrossRef]
- Wang, W.; Sun, W.; Jin, L. Caffeic Acid Alleviates Inflammatory Response in Rheumatoid Arthritis Fibroblast-like Synoviocytes by Inhibiting Phosphorylation of IκB Kinase α/β and IκBα. Int. Immunopharmacol. 2017, 48, 61–66. [Google Scholar] [CrossRef] [PubMed]
- Shifa ul Haq, H.M.; Ashfaq, R.; Mehmood, A.; Shahid, W.; Azam, H.G.; Azam, M.; Tasneem, S.; Akram, S.J.; Malik, K.; Riazuddin, S. Priming with Caffeic Acid Enhances the Potential and Survival Ability of Human Adipose-Derived Stem Cells to Counteract Hypoxia. Regen. Ther. 2023, 22, 115–127. [Google Scholar] [CrossRef]
- Cui, Z.; Zhang, J.; Wang, J.; Liu, J.; Sun, P.; Li, J.; Li, G.; Sun, Y.; Ying, J.; Li, K.; et al. Caffeic Acid Phenethyl Ester: An Effective Antiviral Agent against Porcine Reproductive and Respiratory Syndrome Virus. Antivir. Res. 2024, 225, 105868. [Google Scholar] [CrossRef]
- Saivish, M.V.; Pacca, C.C.; da Costa, V.G.; de Lima Menezes, G.; da Silva, R.A.; Nebo, L.; da Silva, G.C.D.; de Aguiar Milhim, B.H.G.; da Silva Teixeira, I.; Henrique, T.; et al. Caffeic Acid Has Antiviral Activity against Ilhéus Virus In Vitro. Viruses 2023, 15, 494. [Google Scholar] [CrossRef] [PubMed]
- Adem, Ş.; Eyupoglu, V.; Sarfraz, I.; Rasul, A.; Zahoor, A.F.; Ali, M.; Abdalla, M.; Ibrahim, I.M.; Elfiky, A.A. Caffeic Acid Derivatives (CAFDs) as Inhibitors of SARS-CoV-2: CAFDs-Based Functional Foods as a Potential Alternative Approach to Combat COVID-19. Phytomedicine 2021, 85, 153310. [Google Scholar] [CrossRef]
- Alam, M.; Ahmed, S.; Elasbali, A.M.; Adnan, M.; Alam, S.; Hassan, M.I.; Pasupuleti, V.R. Therapeutic Implications of Caffeic Acid in Cancer and Neurological Diseases. Front. Oncol. 2022, 12, 860508. [Google Scholar] [CrossRef]
- Hsu, L.-Y.; Lin, C.-F.; Hsu, W.-C.; Hsu, W.-L.; Chang, T.-C. Evaluation of Polyphenolic Acid Esters as Potential Antioxidants. Biol. Pharm. Bull. 2005, 28, 1211–1215. [Google Scholar] [CrossRef]
- Kougan, G.B.; Tabopda, T.; Kuete, V.; Verpoorte, R. 6—Simple Phenols, Phenolic Acids, and Related Esters from the Medicinal Plants of Africa. In Medicinal Plant Research in Africa; Kuete, V., Ed.; Elsevier: Oxford, UK, 2013; pp. 225–249. ISBN 978-0-12-405927-6. [Google Scholar] [CrossRef]
- Lin, C.-F.; Chang, T.-C.; Chiang, C.-C.; Tsai, H.-J.; Hsu, L.-Y. Synthesis of Selenium-Containing Polyphenolic Acid Esters and Evaluation of Their Effects on Antioxidation and 5-Lipoxygenase Inhibition. Chem. Pharm. Bull. 2005, 53, 1402–1407. [Google Scholar] [CrossRef]
- Sentkowska, A.; Pyrzyńska, K. Investigation of Antioxidant Activity of Selenium Compounds and Their Mixtures with Tea Polyphenols. Mol. Biol. Rep. 2019, 46, 3019–3024. [Google Scholar] [CrossRef] [PubMed]
- Alexander, J.; Olsen, A.-K. Selenium—A Scoping Review for Nordic Nutrition Recommendations 2023. Food Nutr. Res. 2023, 67, 10320. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Sagada, G.; Wang, R.; Li, P.; Xu, B.; Zhang, C.; Qiao, J.; Yan, Y. Different Forms of Selenium Supplementation in Fish Feed: The Bioavailability, Nutritional Functions, and Potential Toxicity. Aquaculture 2022, 549, 737819. [Google Scholar] [CrossRef]
- Ivory, K.; Prieto, E.; Spinks, C.; Armah, C.N.; Goldson, A.J.; Dainty, J.R.; Nicoletti, C. Selenium Supplementation Has Beneficial and Detrimental Effects on Immunity to Influenza Vaccine in Older Adults. Clin. Nutr. 2017, 36, 407–415. [Google Scholar] [CrossRef]
- Harthill, M. Review: Micronutrient Selenium Deficiency Influences Evolution of Some Viral Infectious Diseases. Biol. Trace Elem. Res. 2011, 143, 1325–1336. [Google Scholar] [CrossRef] [PubMed]
- Martinez, S.S.; Huang, Y.; Acuna, L.; Laverde, E.; Trujillo, D.; Barbieri, M.A.; Tamargo, J.; Campa, A.; Baum, M.K. Role of Selenium in Viral Infections with a Major Focus on SARS-CoV-2. Int. J. Mol. Sci. 2021, 23, 280. [Google Scholar] [CrossRef]
- Larvie, D.Y.; Perrin, M.T.; Donati, G.L.; Armah, S.M. COVID-19 Severity Is Associated with Selenium Intake among Young Adults with Low Selenium and Zinc Intake in North Carolina. Curr. Dev. Nutr. 2023, 7, 100044. [Google Scholar] [CrossRef]
- Schomburg, L. Selenium Deficiency in COVID-19—A Possible Long-Lasting Toxic Relationship. Nutrients 2022, 14, 283. [Google Scholar] [CrossRef]
- Stone, C.A.; Kawai, K.; Kupka, R.; Fawzi, W.W. Role of Selenium in Hiv Infection. Nutr. Rev. 2010, 68, 671–681. [Google Scholar] [CrossRef]
- Steinbrenner, H.; Al-Quraishy, S.; Dkhil, M.A.; Wunderlich, F.; Sies, H. Dietary Selenium in Adjuvant Therapy of Viral and Bacterial Infections12. Adv. Nutr. 2015, 6, 73–82. [Google Scholar] [CrossRef]
- Di Bella, S.; Grilli, E.; Cataldo, M.A.; Petrosillo, N. Selenium Deficiency and HIV Infection. Infect. Dis. Rep. 2010, 2, e18. [Google Scholar] [CrossRef] [PubMed]
- Lipinski, B. Can Selenite Be an Ultimate Inhibitor of Ebola and Other Viral Infections? J. Adv. Med. Med. Res. 2015, 6, 319–324. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nasim, M.J.; Ali, W.; da Silva Júnior, E.N.; Saleem, R.S.Z.; Gaucher, C.; Handzlik, J.; Pedatella, S.; Jacob, C. The Chalcogen Exchange: The Replacement of Oxygen with Sulfur and Selenium to Boost the Activity of Natural Products. Sci 2025, 7, 74. https://doi.org/10.3390/sci7020074
Nasim MJ, Ali W, da Silva Júnior EN, Saleem RSZ, Gaucher C, Handzlik J, Pedatella S, Jacob C. The Chalcogen Exchange: The Replacement of Oxygen with Sulfur and Selenium to Boost the Activity of Natural Products. Sci. 2025; 7(2):74. https://doi.org/10.3390/sci7020074
Chicago/Turabian StyleNasim, Muhammad Jawad, Wesam Ali, Eufrânio N. da Silva Júnior, Rahman Shah Zaib Saleem, Caroline Gaucher, Jadwiga Handzlik, Silvana Pedatella, and Claus Jacob. 2025. "The Chalcogen Exchange: The Replacement of Oxygen with Sulfur and Selenium to Boost the Activity of Natural Products" Sci 7, no. 2: 74. https://doi.org/10.3390/sci7020074
APA StyleNasim, M. J., Ali, W., da Silva Júnior, E. N., Saleem, R. S. Z., Gaucher, C., Handzlik, J., Pedatella, S., & Jacob, C. (2025). The Chalcogen Exchange: The Replacement of Oxygen with Sulfur and Selenium to Boost the Activity of Natural Products. Sci, 7(2), 74. https://doi.org/10.3390/sci7020074