Chitosan as a Postharvest Alternative for the Control of Phytophthora capsici in Bell Pepper Fruits

 ,
,  ,
, 
 and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Isolation of Phytophthora spp. in Bell Peppers
2.2. Morphological Identification of Phytophthora spp.
2.3. Pathogenicity Test of Phytophthora spp.
2.4. Molecular Identification and Phylogenetic Analysis of Phytophthora spp.
2.5. Antifungal Activity of Chitosan on the Mycelial Growth of Phytophthora capsici
2.6. Effect of Chitosan on Sporangial Germination
2.7. Postharvest Treatments and Experimental Conditions
2.8. Determination of Lesion Area Using Image Analysis
2.9. Effect of Chitosan on the Incidence of Phytophthora capsica in Bell Peppers
2.10. Effect of Chitosan on the Severity of Phytophthora capsica in Bell Peppers
2.11. Effect of Chitosan on the Fresh and Dry Weight of Bell Peppers Inoculated with Phytophthora capsici
2.12. Determination of Weight Loss in Bell Peppers
2.13. Efficiency of Chitosan Treatments for the Control of Phytophthora capsici
2.14. Statistical Analysis
3. Results
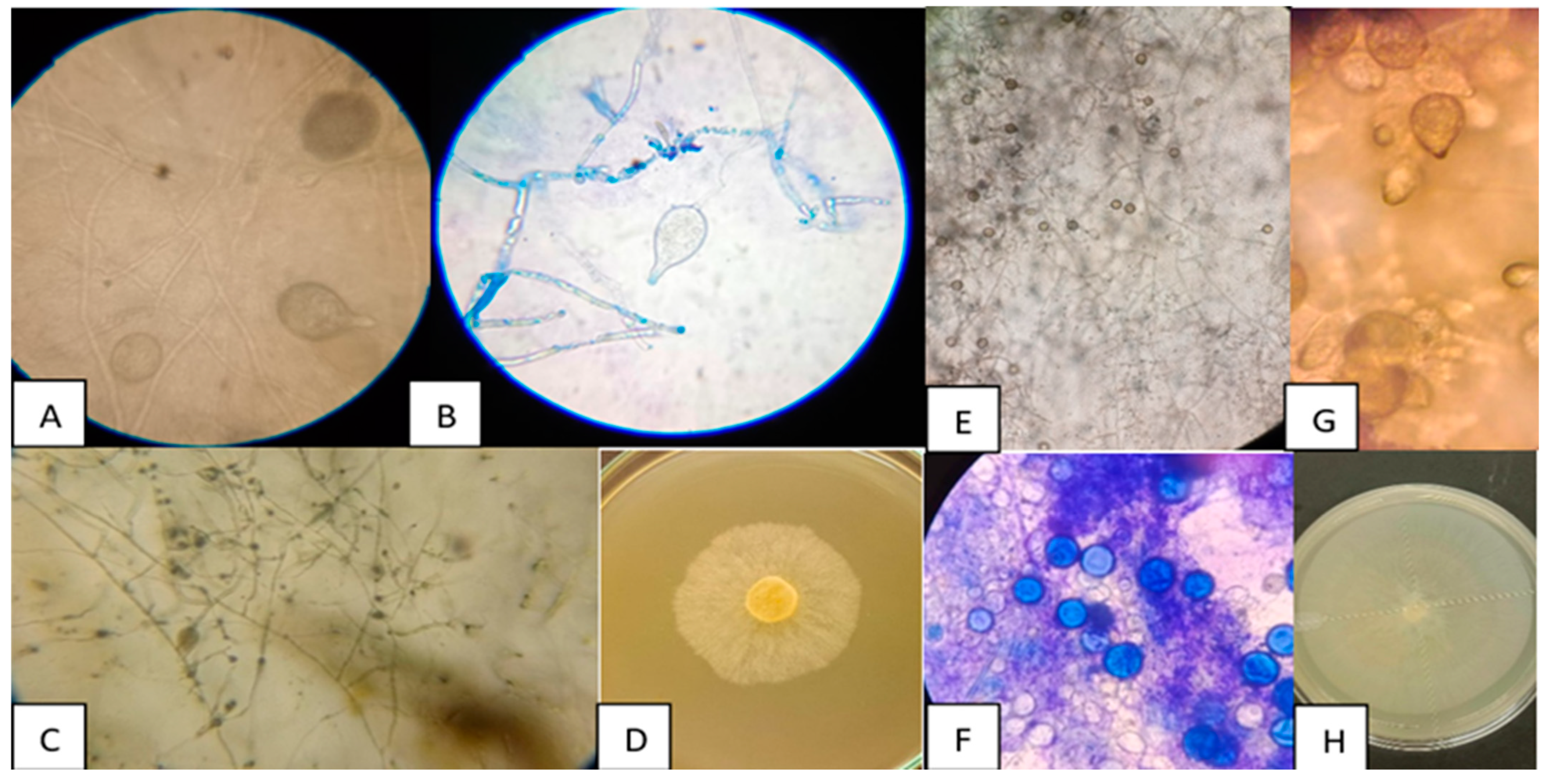
3.1. Morphological Characterisation of Phytophthora spp.
3.2. Pathogenicity of Phytophthora spp. Isolates
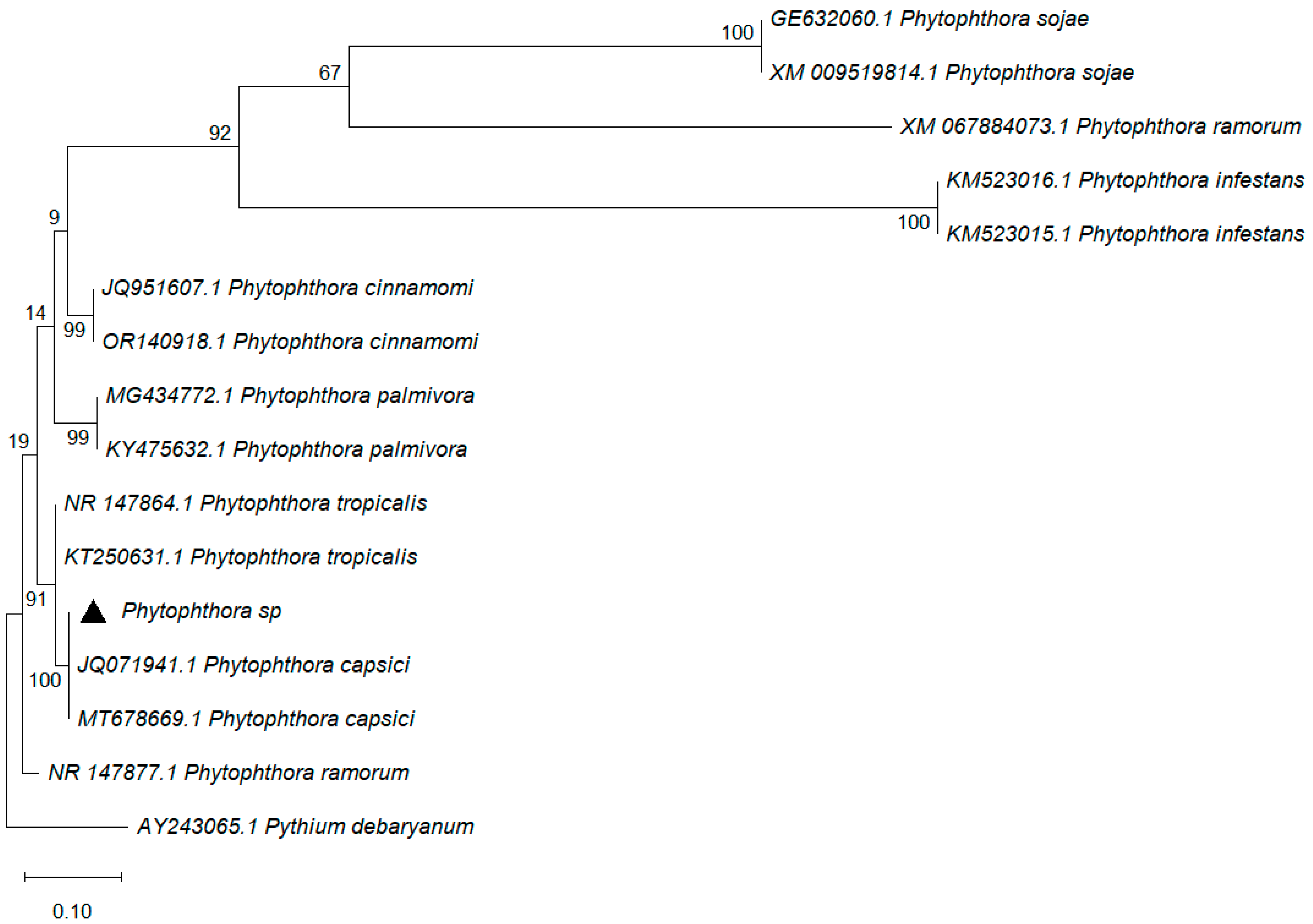
3.3. Molecular Identification of Phytophthora capsici
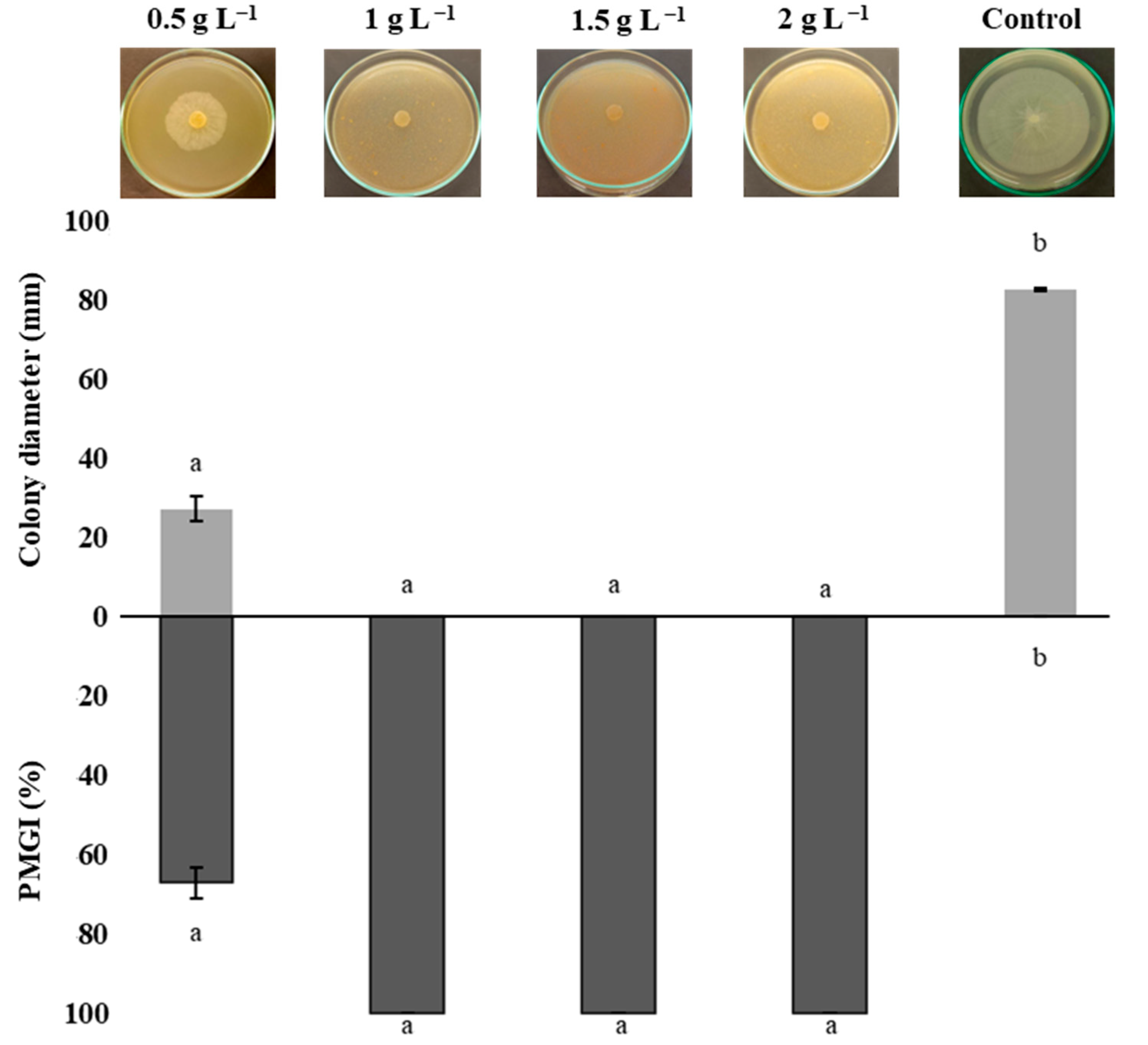
3.4. Mycelial Growth Inhibition of Phytophthora capsici
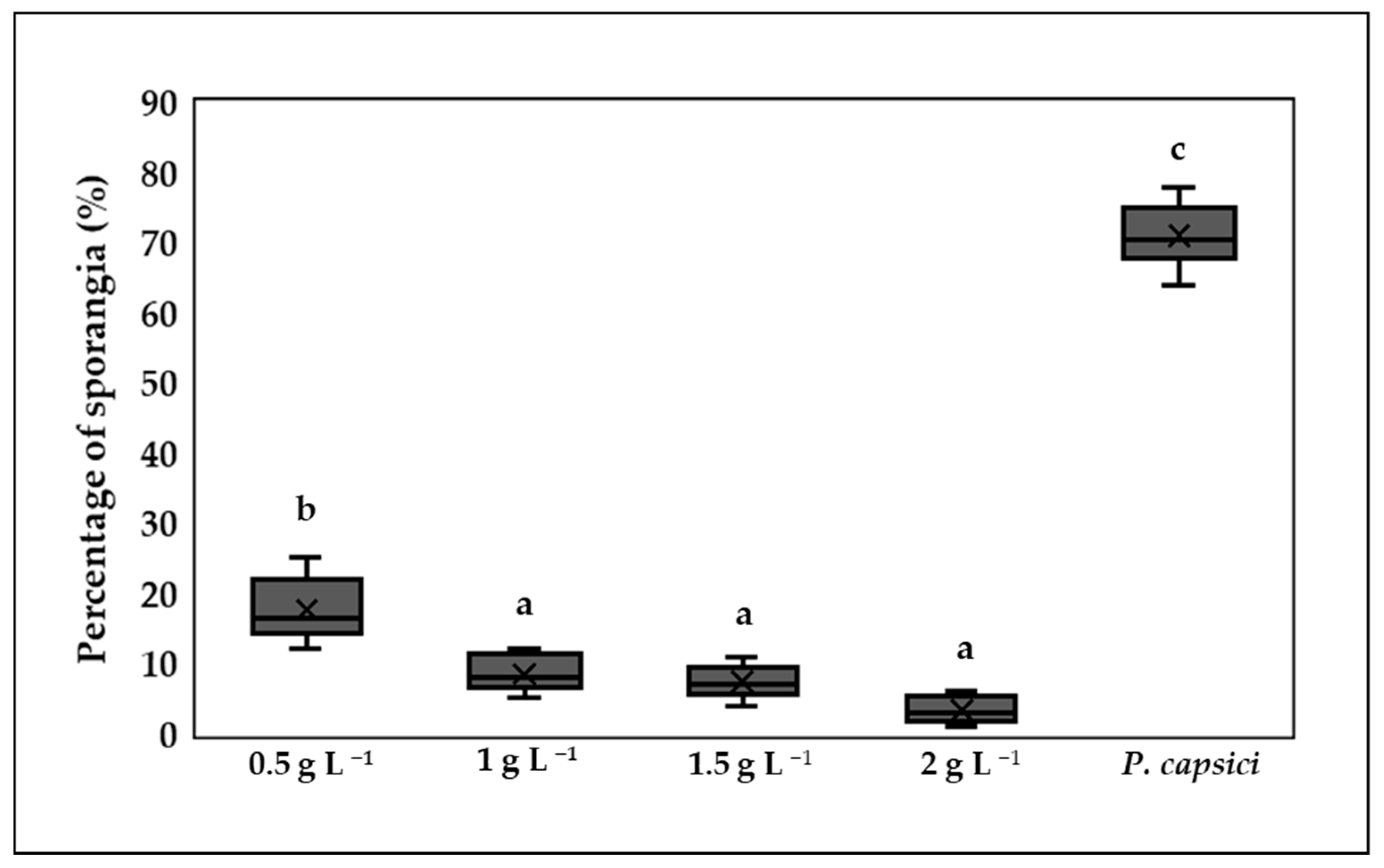
3.5. Inhibition of Sporangium Formation
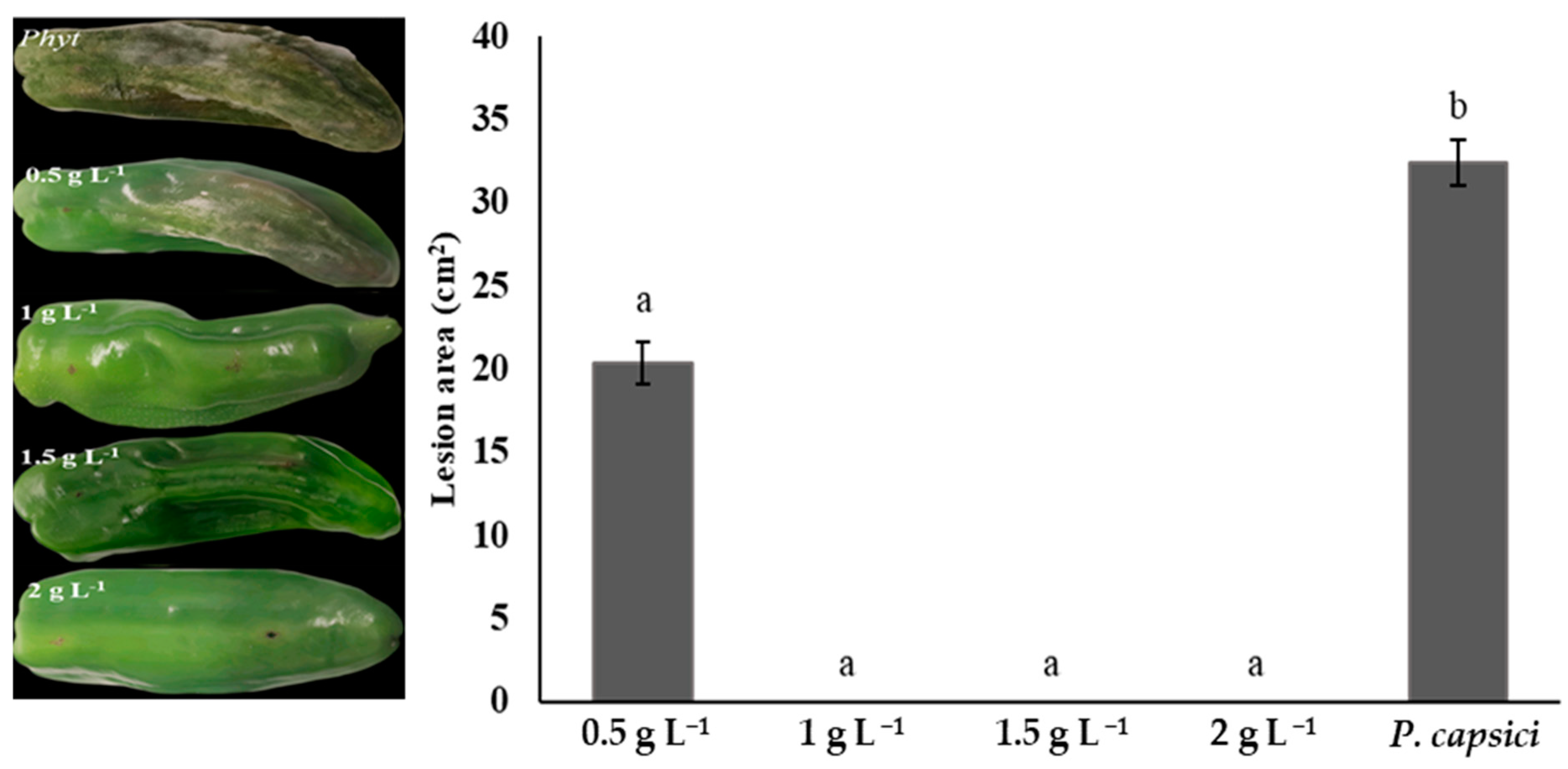
3.6. Lesion Area Measurement in Phytophthora capsica-Infected Bell Peppers
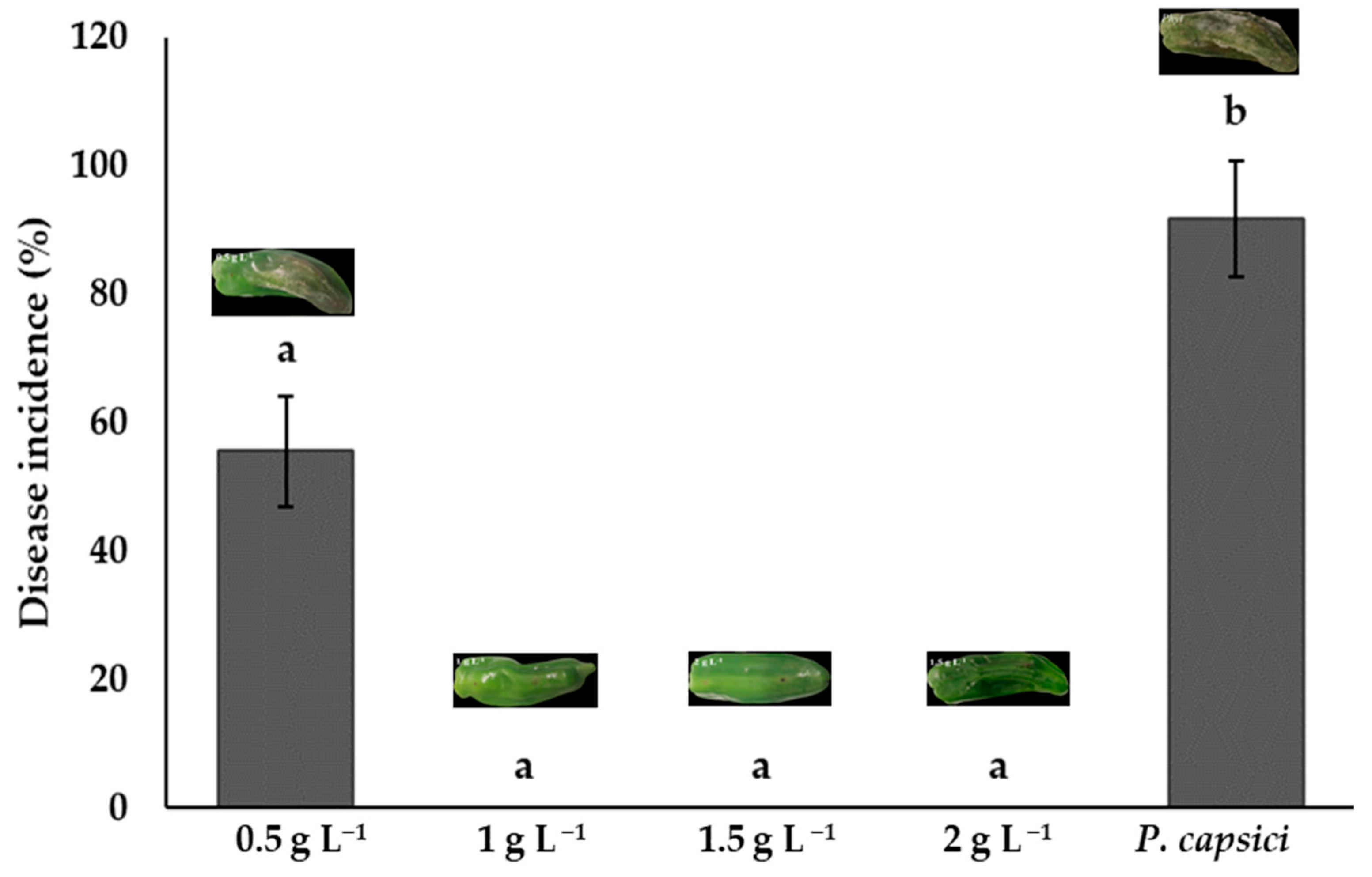
3.7. Disease Incidence and Severity
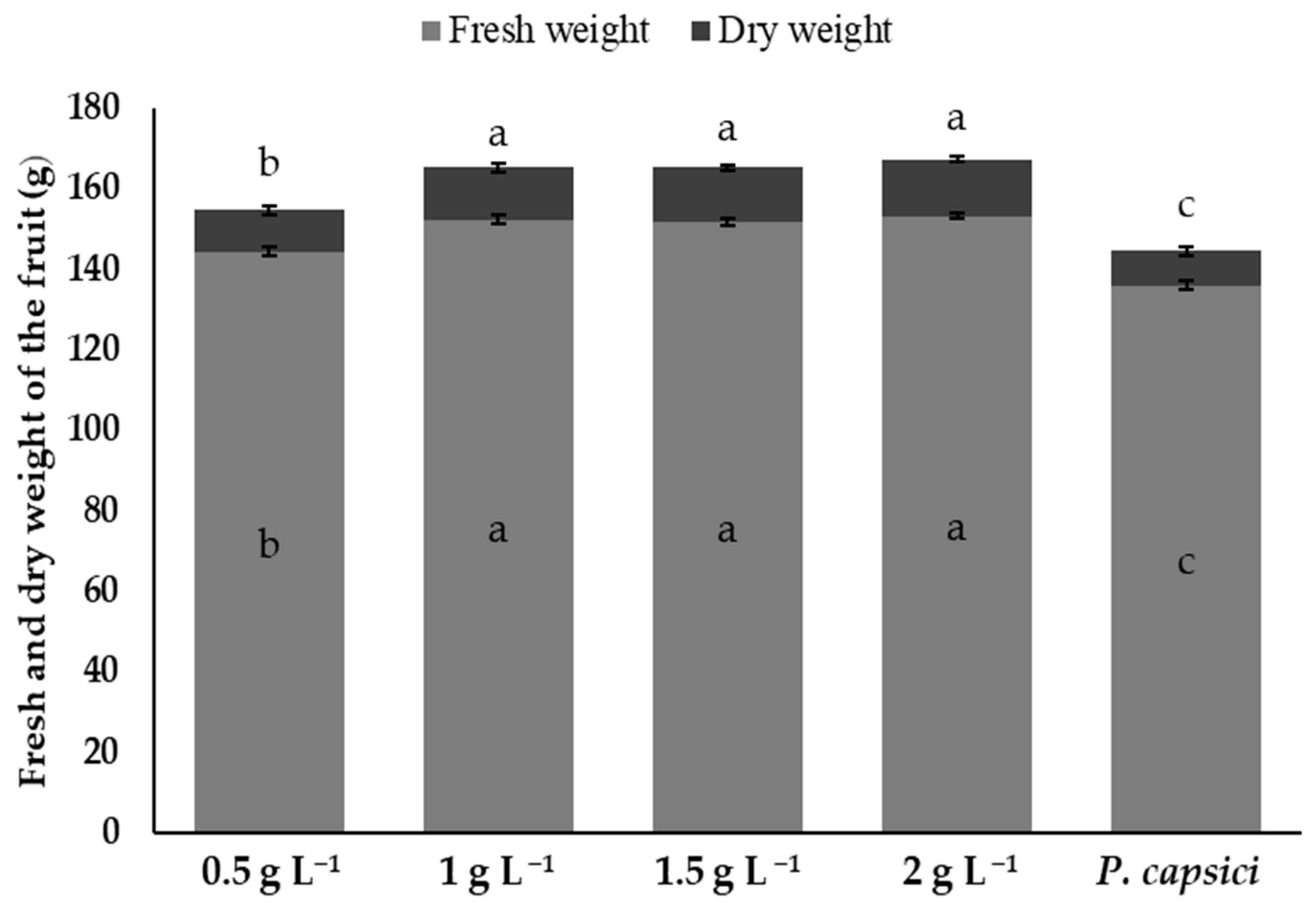
3.8. Fresh and Dry Weight of Fruits
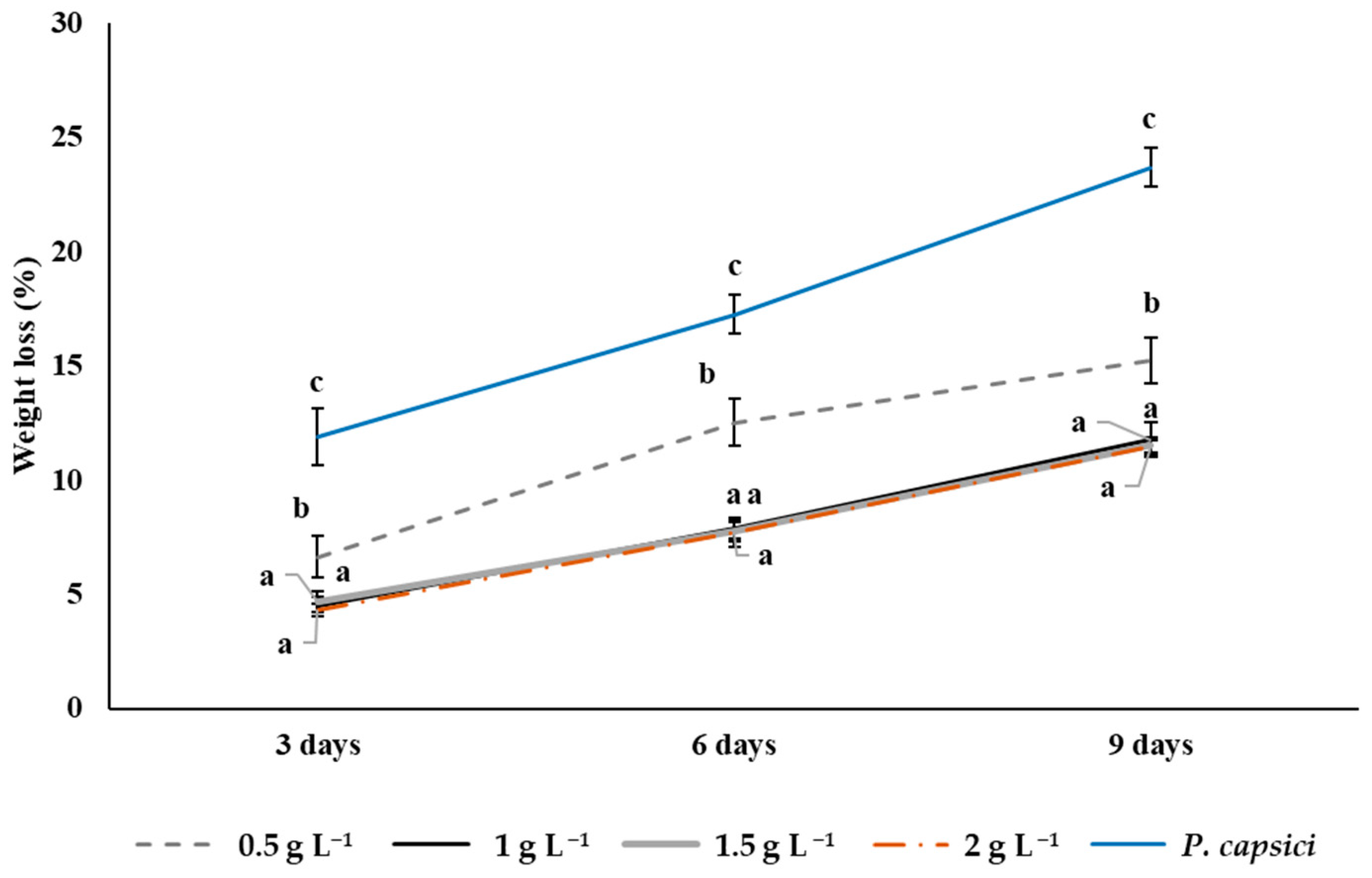
3.9. Weight Loss in Bell Peppers During Storage
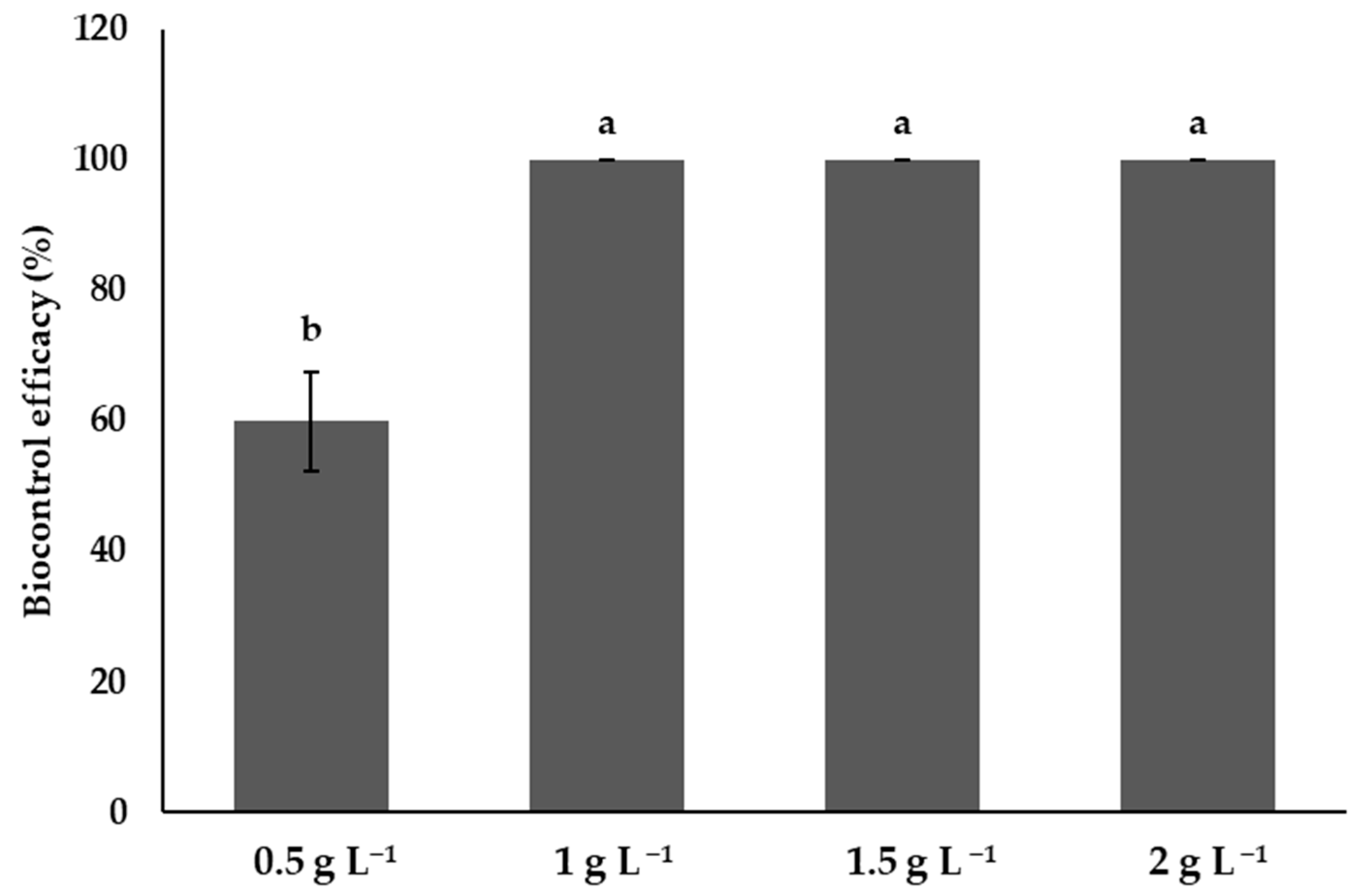
3.10. Efficacy of Chitosan in the Control of Phytophthora capsici
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ivan, I.M.; Popovici, V.; Chițescu, C.L.; Popescu, L.; Luță, E.A.; Ilie, E.I.; Brașoveanu, L.I.; Hotnog, C.M.; Olaru, O.T.; Nițulescu, G.M.; et al. Phytochemical Profile, Antioxidant and Cytotoxic Potential of Capsicum annuum (L.) Dry Hydro-Ethanolic Extract. Pharmaceutics 2024, 16, 245. [Google Scholar] [CrossRef]
- Tahboub, M.B.; Sanogo, S.; Bosland, P.W.; Murray, L. Heat Level in Chile Pepper in Relation to Root and Fruit Infection by Phytophthora capsici. HortScience 2008, 43, 1846–1851. [Google Scholar] [CrossRef]
- Ávila-Oviedo, J.L.; Méndez-Inocencio, C.; Rodríguez-Torres, M.D.; Angoa-Pérez, M.V.; Chávez-Avilés, M.N.; Martínez-Mendoza, E.K.; Oregel-Zamudio, E.; Villar-Luna, E. Antagonistic Effects and Volatile Organic Compound Profiles of Rhizobacteria in the Biocontrol of Phytophthora capsici. Plants 2024, 13, 3224. [Google Scholar] [CrossRef]
- Moradinezhad, F.; Ranjbar, A. Advances in Postharvest Diseases Management of Fruits and Vegetables: A Review. Horticulturae 2023, 9, 1099. [Google Scholar] [CrossRef]
- Dukare, A.S.; Paul, S.; Nambi, V.E.; Gupta, R.K.; Singh, R.; Sharma, K.; Vishwakarma, R.K. Exploitation of Microbial Antagonists for the Control of Postharvest Diseases of Fruits: A Review. Crit. Rev. Food Sci. Nutr. 2019, 59, 1498–1513. [Google Scholar] [CrossRef]
- Volynchikova, E.; Kim, K.D. Biological Control of Oomycete Soilborne Diseases Caused by Phytophthora capsici, Phytophthora infestans, and Phytophthora nicotianae in Solanaceous Crops. Mycobiology 2022, 50, 269–293. [Google Scholar] [CrossRef]
- Dunn, A.R.; Lange, H.W.; Smart, C.D. Evaluation of Commercial Bell Pepper Cultivars for Resistance to Phytophthora Blight (Phytophthora capsici). Plant Health Prog. 2014, 15, 19–24. [Google Scholar] [CrossRef]
- Liu, P.; Cai, Y.; Zhang, J.; Wang, R.; Li, B.; Weng, Q.; Chen, Q. Antifungal Activity of Liquiritin in Phytophthora capsici Comprises Not Only Membrane-Damage-Mediated Autophagy, Apoptosis, and Ca2+ Reduction but Also an Induced Defense Responses in Pepper. Ecotoxicol. Environ. Saf. 2021, 209, 111813. [Google Scholar] [CrossRef]
- Lu, X.H.; Hausbeck, M.K.; Liu, X.L.; Hao, J.J. Wild Type Sensitivity and Mutation Analysis for Resistance Risk to Fluopicolide in Phytophthora capsici. Plant Dis. 2011, 95, 1535–1541. [Google Scholar] [CrossRef]
- Barchenger, D.W.; Lamour, K.H.; Bosland, P.W. Challenges and Strategies for Breeding Resistance in Capsicum Annuum to the Multifarious Pathogen, Phytophthora capsici. Front. Plant Sci. 2018, 9, 628. [Google Scholar] [CrossRef]
- Burandt, Q.C.; Deising, H.B.; von Tiedemann, A. Further Limitations of Synthetic Fungicide Use and Expansion of Organic Agriculture in Europe Will Increase the Environmental and Health Risks of Chemical Crop Protection Caused by Copper-Containing Fungicides. Environ. Toxicol. Chem. 2024, 43, 19–30. [Google Scholar] [CrossRef] [PubMed]
- Popescu, P.-A.; Palade, L.M.; Nicolae, I.-C.; Popa, E.E.; Miteluț, A.C.; Drăghici, M.C.; Matei, F.; Popa, M.E. Chitosan-Based Edible Coatings Containing Essential Oils to Preserve the Shelf Life and Postharvest Quality Parameters of Organic Strawberries and Apples during Cold Storage. Foods 2022, 11, 3317. [Google Scholar] [CrossRef] [PubMed]
- Torres-Rodriguez, J.A.; Reyes-Pérez, J.J.; Carranza-Patino, M.S.; Gaibor-Fernández, R.R.; Rivas-García, T.; Rueda-Puente, E.O. Chitosan: Biocontrol Agent of Fusarium oxysporum in Tomato Fruit (Solanum lycopersicum L.). Emir. J. Food Agric. 2024, 36, 1–9. [Google Scholar] [CrossRef]
- Zheng, H.; Deng, W.; Yu, L.; Shi, Y.; Deng, Y.; Wang, D.; Zhong, Y. Chitosan Coatings with Different Degrees of Deacetylation Regulate the Postharvest Quality of Sweet Cherry through Internal Metabolism. Int. J. Biol. Macromol. 2024, 254, 127419. [Google Scholar] [CrossRef]
- Zhang, L.; Yang, Y.; Zhu, Y.; Hu, H.; Jia, Q.; Sun, C.; Zhu, X.; Liu, Y. Antifungal Activity and Mechanism of Chitosan against Fusarium solani Caused Ginger Soft Rot during Postharvest Storage. Postharvest Biol. Technol. 2024, 208, 112680. [Google Scholar] [CrossRef]
- Torres-Rodriguez, J.A.; Reyes-Pérez, J.J.; Castellanos, T.; Angulo, C.; Quiñones-Aguilar, E.E.; Hernandez-Montiel, L.G. A Biopolymer with Antimicrobial Properties and Plant Resistance Inducer against Phytopathogens: Chitosan. Not. Bot. Horti Agrobot. Cluj-Napoca 2021, 49, 12231. [Google Scholar] [CrossRef]
- Gong, W.; Sun, Y.; Tu, T.; Huang, J.; Zhu, C.; Zhang, J.; Salah, M.; Zhao, L.; Xia, X.; Wang, Y. Chitosan Inhibits Penicillium expansum Possibly by Binding to DNA and Triggering Apoptosis. Int. J. Biol. Macromol. 2024, 259, 129113. [Google Scholar] [CrossRef]
- Romanazzi, G.; Feliziani, E.; Baños, S.B.; Sivakumar, D. Shelf Life Extension of Fresh Fruit and Vegetables by Chitosan Treatment. Crit. Rev. Food Sci. Nutr. 2017, 57, 579–601. [Google Scholar] [CrossRef]
- Le, V.T.; Bach, L.G.; Pham, T.T.; Le, N.T.T.; Ngoc, U.T.P.; Tran, D.-H.N.; Nguyen, D.H. Synthesis and Antifungal Activity of Chitosan-Silver Nanocomposite Synergize Fungicide against Phytophthora capsici. J. Macromol. Sci. Part A 2019, 56, 522–528. [Google Scholar] [CrossRef]
- Héctor, L.M.; Elizabeth, V.O.; José, A.J.Z.; Gabriel, L.U. Treatment with chitosan protects habanero pepper against the infection with Phytophthora capsici. Isr. J. Plant Sci. 2010, 58, 61–65. [Google Scholar] [CrossRef]
- Safari, Z.S.; Ding, P.; Nakasha, J.J.; Yusoff, S.F. Controlling Fusarium Oxysporum Tomato Fruit Rot under Tropical Condition Using Both Chitosan and Vanillin. Coatings 2021, 11, 367. [Google Scholar] [CrossRef]
- Petriccione, M.; De Sanctis, F.; Pasquariello, M.S.; Mastrobuoni, F.; Rega, P.; Scortichini, M.; Mencarelli, F. The Effect of Chitosan Coating on the Quality and Nutraceutical Traits of Sweet Cherry During Postharvest Life. Food Bioprocess Technol. 2015, 8, 394–408. [Google Scholar] [CrossRef]
- Khalifa, I.; Barakat, H.; El-Mansy, H.A.; Soliman, S.A. Preserving Apple (Malus domestica var. Anna) Fruit Bioactive Substances Using Olive Wastes Extract-Chitosan Film Coating. Inf. Process. Agric. 2017, 4, 90–99. [Google Scholar] [CrossRef]
- Long, L.T.; Tan, L.V.; Boi, V.N.; Trung, T.S. Antifungal Activity of Water-Soluble Chitosan against Colletotrichum capsici in Postharvest Chili Pepper. J. Food Process. Preserv. 2018, 42, e13339. [Google Scholar] [CrossRef]
- Sikder, M.B.H.; Islam, M.M. Effect of Shrimp Chitosan Coating on Physico-Chemical Properties and Shelf Life Extension of Banana. Int. J. Eng. Technol. Sci. 2019, 6, 41–54. [Google Scholar] [CrossRef]
- Drenth, A.; Sendall, B. Practical Guide to Detection and Identification of Phytophthora. Trop. Plant Prot. 2001, 1, 32–33. [Google Scholar]
- Ro, N.-Y.; Sebastin, R.; Hur, O.-S.; Cho, G.-T.; Geum, B.; Lee, Y.-J.; Kang, B.-C. Evaluation of Anthracnose Resistance in Pepper (Capsicum spp.) Genetic Resources. Horticulturae 2021, 7, 460. [Google Scholar] [CrossRef]
- Raeder, U.; Broda, P. Rapid Preparation of DNA from Filamentous Fungi. Lett. Appl. Microbiol. 1985, 1, 17–20. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple Sequence Alignment with High Accuracy and High Throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef]
- Talavera, G.; Castresana, J. Improvement of Phylogenies after Removing Divergent and Ambiguously Aligned Blocks from Protein Sequence Alignments. Syst. Biol. 2007, 56, 564–577. [Google Scholar] [CrossRef] [PubMed]
- Dereeper, A.; Guignon, V.; Blanc, G.; Audic, S.; Buffet, S.; Chevenet, F.; Dufayard, J.-F.; Guindon, S.; Lefort, V.; Lescot, M.; et al. Phylogeny.Fr: Robust Phylogenetic Analysis for the Non-Specialist. Nucleic Acids Res. 2008, 36, W465–W469. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Tikhonov, V.E.; Stepnova, E.A.; Babak, V.G.; Yamskov, I.A.; Palma-Guerrero, J.; Jansson, H.-B.; Lopez-Llorca, L.V.; Salinas, J.; Gerasimenko, D.V.; Avdienko, I.D.; et al. Bactericidal and Antifungal Activities of a Low Molecular Weight Chitosan and Its N-/2(3)-(Dodec-2-Enyl)Succinoyl/-Derivatives. Carbohydr. Polym. 2006, 64, 66–72. [Google Scholar] [CrossRef]
- González-Peña Fundora, D.; Falcón-Rodríguez, A.B.; Costales Menéndez, D.; Foroud, N.A.; Vaillant Flores, D.; Aispuro-Hernández, E.; Martínez-Téllez, M.Á. Chitosan Induces Tomato Basal Resistance against Phytophthora Nicotianae and Inhibits Pathogen Development. Can. J. Plant Pathol. 2022, 44, 400–414. [Google Scholar] [CrossRef]
- Cui, Q.; Li, X.; Hu, S.; Yang, D.; Abozeid, A.; Yang, Z.; Jiang, J.; Ren, Z.; Li, D.; Li, D.; et al. The Critical Role of Phenylpropanoid Biosynthesis Pathway in Lily Resistance Against Gray Mold. Int. J. Mol. Sci. 2024, 25, 11068. [Google Scholar] [CrossRef] [PubMed]
- Khaliq, G.; Muda Mohamed, M.T.; Ali, A.; Ding, P.; Ghazali, H.M. Effect of Gum Arabic Coating Combined with Calcium Chloride on Physico-Chemical and Qualitative Properties of Mango (Mangifera indica L.) Fruit during Low Temperature Storage. Sci. Hortic. 2015, 190, 187–194. [Google Scholar] [CrossRef]
- Petriccione, M.; Mastrobuoni, F.; Pasquariello, M.S.; Zampella, L.; Nobis, E.; Capriolo, G.; Scortichini, M. Effect of Chitosan Coating on the Postharvest Quality and Antioxidant Enzyme System Response of Strawberry Fruit during Cold Storage. Foods 2015, 4, 501–523. [Google Scholar] [CrossRef]
- Abbott, W.S. A Method of Computing the Effectiveness of an Insecticide. J. Econ. Entomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]
- Meng, D.; Garba, B.; Ren, Y.; Yao, M.; Xia, X.; Li, M.; Wang, Y. Antifungal Activity of Chitosan against Aspergillus ochraceus and Its Possible Mechanisms of Action. Int. J. Biol. Macromol. 2020, 158, 1063–1070. [Google Scholar] [CrossRef]
- Li, Z.; Long, W.; Zheng, J.; Lei, J. Isolation and Identification of Phytophthora capsici in Guangdong Province and Measurement of Their Pathogenicity and Physiological Race Differentiation. Front. Agric. China 2007, 1, 377–381. [Google Scholar] [CrossRef]
- Soto-Plancarte, A.; Rodríguez-Alvarado, G.; Fernández-Pavía, Y.L.; Pedraza-Santos, M.E.; López-Pérez, L.; Díaz-Celaya, M.; Fernández-Pavía, S.P. Protocolos de aislamiento y diagnóstico de Phytophthora spp. enfoque aplicado a la investigación. Rev. Mex. Cienc. Agrí. 2017, 8, 1867–1880. [Google Scholar] [CrossRef]
- Meyer, M.D.; Hausbeck, M.K. Age-Related Resistance to Phytophthora Fruit Rot in ‘Dickenson Field’ Processing Pumpkin and ‘Golden Delicious’ Winter Squash Fruit. Plant Dis. 2013, 97, 446–552. [Google Scholar] [CrossRef] [PubMed]
- Foster, J.M.; Hausbeck, M.K. Resistance of Pepper to Phytophthora Crown, Root, and Fruit Rot Is Affected by Isolate Virulence. Plant Dis. 2010, 94, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Van Tran, Q.; Ha, C.V.; Vvedensky, V.V.; Han, V.-C. Current Status and Characterization of Phytophthora Species Associated with Gummosis of Citrus in Northern Vietnam. J. Phytopathol. 2023, 171, 478–488. [Google Scholar] [CrossRef]
- Kuswinanti, T.; Patandjengi, B.; Melina; Hardina, N. Morphological and Molecular Characterization of Phytophthora Palmivora Isolates and Their Virulence Test on Cocoa Clone Sulawesi 2. J. Fitopatol. Indones. 2023, 19, 145–155. [Google Scholar] [CrossRef]
- Karyath Palliyath, G.; Kilingar Subrahmanya, M.; Antony, G.; Binod Bihari, S.; Hegde, V.; Muliyar Krishna, R. A Rapid in Vitro Leaf Inoculation Assay to Investigate Phytophthora palmivora–Coconut Interactions. J. Phytopathol. 2021, 169, 316–328. [Google Scholar] [CrossRef]
- Reyes-Tena, A.; Rodríguez-Alvarado, G.; Fernández-Pavía, S.P.; Pedraza-Santos, M.E.; Larsen, J.; Vázquez-Marrufo, G.; Reyes-Tena, A.; Rodríguez-Alvarado, G.; Fernández-Pavía, S.P.; Pedraza-Santos, M.E.; et al. Caracterización morfológica de aislados de Phytophthora capsici provenientes de Jalisco y Michoacán, México. Rev. Mex. Fitopatol. 2021, 39, 75–93. [Google Scholar] [CrossRef]
- Bhunjun, C.S.; Phillips, A.J.L.; Jayawardena, R.S.; Promputtha, I.; Hyde, K.D. Importance of Molecular Data to Identify Fungal Plant Pathogens and Guidelines for Pathogenicity Testing Based on Koch’s Postulates. Pathogens 2021, 10, 1096. [Google Scholar] [CrossRef]
- Abdul Haq, M.; Shahzad, S.; Lodhi, A.M.; Rajput, A.Q. Morphological and Molecular Characterization of Four Phytophthora Species with the First Report of Phytophthora Lacustris from Pakistan. Plant Biosyst.-Int. J. Deal. Asp. Plant Biol. 2024, 158, 457–463. [Google Scholar] [CrossRef]
- Seddaiu, S.; Linaldeddu, B.T. First Report of Phytophthora acerina, P. plurivora, and P. pseudocryptogea Associated with Declining Common Alder Trees in Italy. Plant Dis. 2020, 104, 1874. [Google Scholar] [CrossRef]
- Mohammed, M.A.; Syeda, J.T.M.; Wasan, K.M.; Wasan, E.K. An Overview of Chitosan Nanoparticles and Its Application in Non-Parenteral Drug Delivery. Pharmaceutics 2017, 9, 53. [Google Scholar] [CrossRef]
- Wei, L.; Chen, Y.; Tan, W.; Li, Q.; Gu, G.; Dong, F.; Guo, Z. Synthesis, Characterization, and Antifungal Activity of Pyridine-Based Triple Quaternized Chitosan Derivatives. Molecules 2018, 23, 2604. [Google Scholar] [CrossRef]
- Ma, Z.; Garrido-Maestu, A.; Jeong, K.C. Application, Mode of Action, and in Vivo Activity of Chitosan and Its Micro- and Nanoparticles as Antimicrobial Agents: A Review. Carbohydr. Polym. 2017, 176, 257–265. [Google Scholar] [CrossRef] [PubMed]
- Torres-Rodriguez, J.A.; Reyes-Pérez, J.J.; Carranza-Patiño, M.S.; Herrera-Feijoo, R.J.; Preciado-Rangel, P.; Hernandez-Montiel, L.G. Biocontrol of Fusarium solani: Antifungal Activity of Chitosan and Induction of Defence Enzymes. Plants 2025, 14, 431. [Google Scholar] [CrossRef]
- Rabea, E.I.; Badawy, M.E.-T.; Stevens, C.V.; Smagghe, G.; Steurbaut, W. Chitosan as Antimicrobial Agent: Applications and Mode of Action. Biomacromolecules 2003, 4, 1457–1465. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Moya, F.; Suarez-Fernandez, M.; Lopez-Llorca, L.V. Molecular Mechanisms of Chitosan Interactions with Fungi and Plants. Int. J. Mol. Sci. 2019, 20, 332. [Google Scholar] [CrossRef]
- Silva Júnior, S.; Stamford, N.P.; Lima, M.A.B.; Arnaud, T.M.S.; Pintado, M.M.; Sarmento, B.F. Characterization and Inhibitory Activity of Chitosan on Hyphae Growth and Morphology of Botrytis Cinerea Plant Pathogen. Int. J. Appl. Res. Nat. Prod. 2014, 7, 31–38. [Google Scholar]
- Vitti, A.; Coviello, L.; Triunfo, M.; Guarnieri, A.; Scieuzo, C.; Salvia, R.; Falabella, P.; Nuzzaci, M. Actividad Antifúngica in Vitro y Eficacia Del Recubrimiento Comestible in Vivo Del Quitosano Derivado de Insectos Contra Botrytis cinerea En Fresa. Int. J. Biol. Macromol. 2024, 279, 135158. [Google Scholar] [CrossRef]
- Oliveira Junior, E.N.D.; Melo, I.S.D.; Franco, T.T. Changes in Hyphal Morphology Due to Chitosan Treatment in Some Fungal Species. Braz. Arch. Biol. Technol. 2012, 55, 637–646. [Google Scholar] [CrossRef]
- Jiménez-Mejía, R.; Arceo-Martínez, M.T.; Loeza-Lara, P.D. Quitosano: Actividad antimicrobiana y mecanismos de acción. E-CUCBA 2018, 9, 17–23. [Google Scholar]
- Huang, X.; You, Z.; Luo, Y.; Yang, C.; Ren, J.; Liu, Y.; Wei, G.; Dong, P.; Ren, M. Actividad Antifúngica Del Quitosano Contra Phytophthora infestans, Patógeno Del Tizón Tardío de La Papa. Int. J. Biol. Macromol. 2021, 166, 1365–1376. [Google Scholar] [CrossRef]
- Xu, J.; Zhao, X.; Han, X.; Du, Y. Antifungal Activity of Oligochitosan against Phytophthora capsici and Other Plant Pathogenic Fungi in Vitro. Pestic. Biochem. Physiol. 2007, 87, 220–228. [Google Scholar] [CrossRef]
- Abdel-Rahman, F.A.; Monir, G.A.; Hassan, M.S.S.; Ahmed, Y.; Refaat, M.H.; Ismail, I.A.; El-Garhy, H.A.S. Exogenously Applied Chitosan and Chitosan Nanoparticles Improved Apple Fruit Resistance to Blue Mold, Upregulated Defense-Related Genes Expression, and Maintained Fruit Quality. Horticulturae 2021, 7, 224. [Google Scholar] [CrossRef]
- Chen, C.; Cai, N.; Chen, J.; Peng, X.; Wan, C. Chitosan-Based Coating Enriched with Hairy Fig (Ficus hirta Vahl.) Fruit Extract for “Newhall” Navel Orange Preservation. Coatings 2018, 8, 445. [Google Scholar] [CrossRef]
- Xing, K.; Zhu, X.; Peng, X.; Qin, S. Chitosan Antimicrobial and Eliciting Properties for Pest Control in Agriculture: A Review. Agron. Sustain. Dev. 2015, 35, 569–588. [Google Scholar] [CrossRef]
- Shcherban, A.B. Chitosan and Its Derivatives as Promising Plant Protection Tools. Vavilov J. Genet. Breed. 2023, 27, 1010–1021. [Google Scholar] [CrossRef]
- Meng, X.; Yang, L.; Kennedy, J.F.; Tian, S. Effects of Chitosan and Oligochitosan on Growth of Two Fungal Pathogens and Physiological Properties in Pear Fruit. Carbohydr. Polym. 2010, 81, 70–75. [Google Scholar] [CrossRef]
- Abebe, Z.; Tola, Y.B.; Mohammed, A. Effects of Edible Coating Materials and Stages of Maturity at Harvest on Storage Life and Quality of Tomato (Lycopersicon esculentum Mill.) Fruits. Afr. J. Agric. Res. 2017, 12, 550–565. [Google Scholar] [CrossRef]
- Shah, S.; Hashmi, M.S. Chitosan–Aloe Vera Gel Coating Delays Postharvest Decay of Mango Fruit. Hortic. Environ. Biotechnol. 2020, 61, 279–289. [Google Scholar] [CrossRef]
- Yunita, R.; Amin, N.N.; Damayanti, T.A. Pemanfaatan Kitosan Untuk Mengendalikan Antraknosa Pada Pepaya (Colletotrichum gloeosporioides) Dan Meningkatkan Daya Simpan Buah. J. Fitopatol. Indones. 2012, 8, 97. [Google Scholar] [CrossRef]
- Hernández-López, G.; Ventura-Aguilar, R.I.; Correa-Pacheco, Z.N.; Bautista-Baños, S.; Barrera-Necha, L.L. Recubrimiento Comestible de Quitosano Nanoestructurado Cargado Con α-Pineno Para La Preservación de La Calidad Poscosecha de Capsicum annuum L. y El Control de Alternaria alternata. Int. J. Biol. Macromol. 2020, 165, 1881–1888. [Google Scholar] [CrossRef] [PubMed]
- Correa-Pacheco, Z.N.; Bautista-Baños, S.; Corona-Rangel, M.L.; Ventura-Aguilar, R.I.; Barrera-Necha, L.L.; Cuevas-Gómez, K.D. Effect of Chitosan-Based Natural Products Nanocoatings on Green Bell Peppers During Storage. Food Bioprocess Technol. 2023, 16, 1703–1715. [Google Scholar] [CrossRef]
- Sharma, S.; Krishna, H.; Barman, K.; Kole, B.; Singh, S.K.; Behera, T.K. Synergistic Effect of Polyamine Treatment and Chitosan Coating on Postharvest Senescence and Enzyme Activity of Bell Pepper (Capsicum annuum L.) Fruit. S. Afr. J. Bot. 2022, 151, 175–184. [Google Scholar] [CrossRef]
- Narsaiah, K.; Wilson, R.A.; Gokul, K.; Mandge, H.M.; Jha, S.N.; Bhadwal, S.; Anurag, R.K.; Malik, R.K.; Vij, S. Effect of Bacteriocin-Incorporated Alginate Coating on Shelf-Life of Minimally Processed Papaya (Carica papaya L.). Postharvest Biol. Technol. 2015, 100, 212–218. [Google Scholar] [CrossRef]
- Rastegar, S.; Hassanzadeh Khankahdani, H.; Rahimzadeh, M. Effectiveness of Alginate Coating on Antioxidant Enzymes and Biochemical Changes during Storage of Mango Fruit. J. Food Biochem. 2019, 43, e12990. [Google Scholar] [CrossRef]
- Chaple, S.; Vishwasrao, C.; Ananthanarayan, L. Edible Composite Coating of Methyl Cellulose for Post-Harvest Extension of Shelf-Life of Finger Hot Indian Pepper (Pusa jwala). J. Food Process. Preserv. 2017, 41, e12807. [Google Scholar] [CrossRef]
- Ruelas-Chacon, X.; Contreras-Esquivel, J.C.; Montañez, J.; Aguilera-Carbo, A.F.; Reyes-Vega, M.L.; Peralta-Rodriguez, R.D.; Sanchéz-Brambila, G. Guar Gum as an Edible Coating for Enhancing Shelf-Life and Improving Postharvest Quality of Roma Tomato (Solanum lycopersicum L.). J. Food Qual. 2017, 2017, 8608304. [Google Scholar] [CrossRef]
- Hernández-Muñoz, P.; Almenar, E.; Ocio, M.J.; Gavara, R. Effect of Calcium Dips and Chitosan Coatings on Postharvest Life of Strawberries (Fragaria x ananassa). Postharvest Biol. Technol. 2006, 39, 247–253. [Google Scholar] [CrossRef]
- Bhanushree, L.S.; Vasudeva, K.R.; Suresha, G.J.; Sadananda, G.K.; Mohamad Tayeebulla, H.; Halesh, G.K. Influence of Chitosan on Postharvest Behavior of Papaya (Carica papaya L.) Fruits under Different Storage Conditions. J. Pharmacogn. Phytochem. 2018, 7, 2010–2014. [Google Scholar]
- Chien, P.-J.; Sheu, F.; Yang, F.-H. Effects of Edible Chitosan Coating on Quality and Shelf Life of Sliced Mango Fruit. J. Food Eng. 2007, 78, 225–229. [Google Scholar] [CrossRef]
- Velickova, E.; Winkelhausen, E.; Kuzmanova, S.; Alves, V.D.; Moldão-Martins, M. Impact of Chitosan-Beeswax Edible Coatings on the Quality of Fresh Strawberries (Fragaria ananassa cv Camarosa) under Commercial Storage Conditions. LWT-Food Sci. Technol. 2013, 52, 80–92. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate | Severity | Damage Diameter (cm) | Fresh Weight | Dry Weight |
|---|---|---|---|---|
| Phy1 | 5 | 13.17 ± 5.85 a | 139.24 ± 4.35 c | 13.66 ± 0.47 b |
| Phy2 | 4 | 11.67 ± 2.58 a | 149.32 ± 2.74 b | 14.90 ± 0.45 b |
| Control | 0 | 0.00 ± 0.00 b | 160.03 ± 3.43 a | 16.15 ± 1.30 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Torres-Rodriguez, J.A.; Reyes Pérez, J.J.; Ramos, L.T.L.; Gonzalo-Matute, L.; Rueda-Puente, E.O.; Hernandez-Montiel, L.G. Chitosan as a Postharvest Alternative for the Control of Phytophthora capsici in Bell Pepper Fruits. Sci 2025, 7, 37. https://doi.org/10.3390/sci7020037
Torres-Rodriguez JA, Reyes Pérez JJ, Ramos LTL, Gonzalo-Matute L, Rueda-Puente EO, Hernandez-Montiel LG. Chitosan as a Postharvest Alternative for the Control of Phytophthora capsici in Bell Pepper Fruits. Sci. 2025; 7(2):37. https://doi.org/10.3390/sci7020037
Chicago/Turabian StyleTorres-Rodriguez, Juan Antonio, Juan José Reyes Pérez, Luis Tarquino Llerena Ramos, Leonardo Gonzalo-Matute, Edgar Omar Rueda-Puente, and Luis Guillermo Hernandez-Montiel. 2025. "Chitosan as a Postharvest Alternative for the Control of Phytophthora capsici in Bell Pepper Fruits" Sci 7, no. 2: 37. https://doi.org/10.3390/sci7020037
APA StyleTorres-Rodriguez, J. A., Reyes Pérez, J. J., Ramos, L. T. L., Gonzalo-Matute, L., Rueda-Puente, E. O., & Hernandez-Montiel, L. G. (2025). Chitosan as a Postharvest Alternative for the Control of Phytophthora capsici in Bell Pepper Fruits. Sci, 7(2), 37. https://doi.org/10.3390/sci7020037