A Systematic Review of Aphantasia: Concept, Measurement, Neural Basis, and Theory Development


{kind=link}
Abstract
1. Introduction
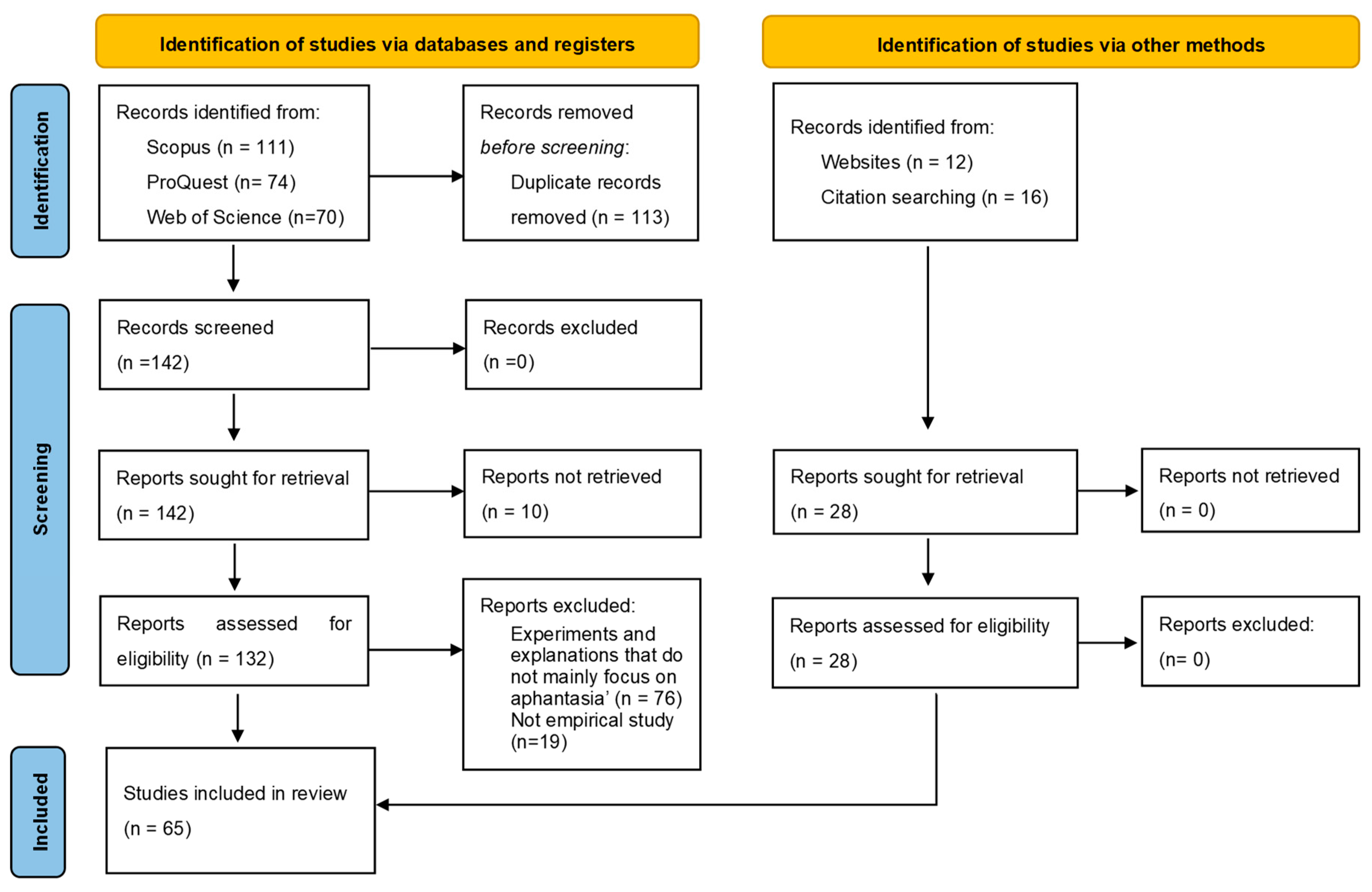
2. Literature Retrieval and Screening
3. Definition, Measurement, and Prevalence of Aphantasia
4. Aphantasia and Cognitive Processing
4.1. Visual and Non-Visual Imagery Ability
4.2. Aphantasia and Memory
4.3. Aphantasia and Object and Spatial Imagery
4.4. Aphantasia and Atemporal and Future Imagination
4.5. Aphantasia and Mental Rotation Task Performance
4.6. Aphantasia and Visual Searching Ability
5. Aphantasia and Disorders and Emotional Processing
5.1. Emotion
5.2. Mental Health
5.3. Post-Traumatic Stress Disorder (PTSD)
5.4. Autism
5.5. Prosopagnosia
6. The Neural Basis of Aphantasia
7. Theory Development
8. Summary and Future Directions
8.1. Clarify Definition and Diagnosis
8.2. Strengthen Behavioral Research
8.3. Discover Neural Bases
8.4. Construct and Refine Theories
8.5. Encourage Direct and Conceptual Replications
9. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Galton, F. Statistics of mental imagery. Mind 1880, 5, 301–318. [Google Scholar] [CrossRef]
- Zeman, A. Aphantasia and hyperphantasia: Exploring imagery vividness extremes. Trends in Cognitive Sciences. Trends Cogn. Sci. 2024, 28, 467–480. [Google Scholar] [CrossRef] [PubMed]
- Keogh, R.; Pearson, J.; Zeman, A. Aphantasia: The science of visual imagery extremes. Handb. Clin. Neurol. 2021, 178, 277–296. [Google Scholar] [PubMed]
- Zeman, A.; Milton, F.; Della Sala, S.; Dewar, M.; Frayling, T.; Gaddum, J.; Hattersley, A.; Heuerman-Williamson, B.; Jones, K.; MacKisack, M.; et al. Phantasia–The psychological significance of lifelong visual imagery vividness extremes. Cortex 2020, 130, 426–440. [Google Scholar] [CrossRef] [PubMed]
- Zeman, A.; Dewar, M.; Della Sala, S. Lives without imagery—Congenital aphantasia. Cortex 2015, 73, 378–380. [Google Scholar] [CrossRef]
- Dawes, A.J.; Keogh, R.; Andrillon, T.; Pearson, J. A cognitive profile of multi-sensory imagery, memory and dreaming in aphantasia. Sci. Rep. 2020, 10, 10022. [Google Scholar] [CrossRef]
- Pearson, J. The human imagination: The cognitive neuroscience of visual mental imagery. Nat. Rev. Neurosci. 2019, 20, 624–634. [Google Scholar] [CrossRef]
- Dijkstra, N.; Bosch, S.E.; van Gerven, M.A. Shared neural mechanisms of visual perception and imagery. Trends Cogn. Sci. 2019, 23, 423–434. [Google Scholar] [CrossRef]
- Luchini, C.; Veronese, N.; Nottegar, A.; Shin, J.I.; Gentile, G.; Granziol, U.; Soysal, P.; Alexinschi, O.; Smith, L.; Solmi, M. Assessing the quality of studies in meta-research: Review/guidelines on the most important quality assessment tools. Pharm. Stat. 2021, 20, 185–195. [Google Scholar] [CrossRef]
- Shaheen, N.; Shaheen, A.; Ramadan, A.; Hefnawy, M.T.; Ramadan, A.; Ibrahim, I.A.; Hassanein, M.E.; Ashour, M.E.; Flouty, O. Appraising systematic reviews: A comprehensive guide to ensuring validity and reliability. Front. Res. Metr. Anal. 2023, 8, 1268045. [Google Scholar] [CrossRef]
- Blomkvist, A. Aphantasia: In search of a theory. Mind Lang. 2022, 38, 866–888. [Google Scholar] [CrossRef]
- Blomkvist, A.; Marks, D.F. Defining and ‘diagnosing’ aphantasia: Condition or individual difference? Cortex 2023, 169, 220–234. [Google Scholar] [CrossRef] [PubMed]
- Keogh, R.; Pearson, J. The blind mind: No sensory visual imagery in aphantasia. Cortex 2018, 105, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Fulford, J.; Milton, F.; Salas, D.; Smith, A.; Simler, A.; Winlove, C.; Zeman, A. The neural correlates of visual imagery vividness—An fMRI study and literature review. Cortex 2018, 105, 26–40. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, C.; Schwarzkopf, D.S.; Silvanto, J. Visual working memory performance in aphantasia. Cortex 2018, 105, 61–73. [Google Scholar] [CrossRef]
- Milton, F.; Fulford, J.; Dance, C.; Gaddum, J.; Heuerman-Williamson, B.; Jones, K.; Knight, K.F.; MacKisack, M.; Winlove, C.; Zeman, A. Behavioral and neural signatures of visual imagery vividness extremes: Aphantasia versus hyperphantasia. Cereb. Cortex Commun. 2021, 2, tgab035. [Google Scholar] [CrossRef]
- Bainbridge, W.A.; Pounder, Z.; Eardley, A.F.; Baker, C.I. Quantifying aphantasia through drawing: Those without visual imagery show deficits in object but not spatial memory. Cortex 2021, 135, 159–172. [Google Scholar] [CrossRef]
- Nanay, B. Unconscious mental imagery: Unconscious mental imagery. Philos. Trans. R. Soc. B Biol. Sci. 2021, 376, 20190689. [Google Scholar] [CrossRef]
- Keogh, R.; Pearson, J. Attention driven phantom vision: Measuring the sensory strength of attentional templates and their relation to visual mental imagery and aphantasia: Measuring attentional templates. Philos. Trans. R. Soc. B Biol. Sci. 2021, 376, 20190688. [Google Scholar] [CrossRef]
- Monzel, M.; Mitchell, D.; Macpherson, F.; Pearson, J.; Zeman, A. Aphantasia, dysikonesia, anauralia: Call for a single term for the lack of mental imagery—Commentary on dance et al. (2021) and Hinwar and lambert (2021). Cortex 2022, 150, 149–152. [Google Scholar] [CrossRef]
- Krempel, R.; Monzel, M. Aphantasia and involuntary imagery. Conscious. Cogn. 2024, 120, 103679. [Google Scholar] [CrossRef] [PubMed]
- Marks, D.F. Visual imagery differences in the recall of pictures. Br. J. Psychol. 1973, 64, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Keogh, R.; Pearson, J. Revisiting the blind mind: Still no evidence for sensory visual imagery in individuals with aphantasia. Neurosci. Res. 2024, 201, 27–30. [Google Scholar] [CrossRef] [PubMed]
- Marks, D.F. New directions for mental imagery research. J. Ment. Imag. 1995, 19, 153–167. [Google Scholar]
- Kay, L.; Keogh, R.; Andrillon, T.; Pearson, J. The pupillary light response as a physiological index of aphantasia, sensory and phenomenological imagery strength. eLife 2022, 11, e72484. [Google Scholar] [CrossRef]
- Konigsmark, V.T.; Bergmann, J.; Reeder, R.R. The Ganzflicker experience: High probability of seeing vivid and complex pseudo-hallucinations with imagery but not aphantasia. Cortex 2021, 141, 522–534. [Google Scholar] [CrossRef]
- Reeder, R.R. Ganzflicker reveals the complex relationship between visual mental imagery and pseudo-hallucinatory experiences: A replication and expansion. Collabra Psychol. 2022, 8, 36318. [Google Scholar] [CrossRef]
- Faw, B. Conflicting intuitions may be based on differing abilities: Evidence from mental imaging research. J. Conscious. Stud. 2009, 16, 45–68. [Google Scholar]
- Dance, C.J.; Ipser, A.; Simner, J. The prevalence of aphantasia (imagery weakness) in the general population. Conscious. Cogn. 2022, 97, 103243. [Google Scholar] [CrossRef]
- Monzel, M.; Vetterlein, A.; Reuter, M. No general pathological significance of aphantasia: An evaluation based on criteria for mental disorders. Scand. J. Psychol. 2023, 64, 314–324. [Google Scholar] [CrossRef]
- Takahashi, J.; Saito, G.; Omura, K.; Yasunaga, D.; Sugimura, S.; Sakamoto, S.; Horikawa, T.; Gyoba, J. Diversity of aphantasia revealed by multiple assessments of visual imagery, multisensory imagery, and cognitive style. Front. Psychol. 2023, 14, 1174873. [Google Scholar] [CrossRef] [PubMed]
- Gouveia, L.A. Imagination Capacity in University Students: Aphantasia and Its Possible Changes in Personal Development; Seven Editora: São José dos Pinhais, Brazil, 2023. [Google Scholar] [CrossRef]
- Beran, M.J.; James, B.T.; French, K.; Haseltine, E.L.; Kleider-Offutt, H.M. Assessing aphantasia prevalence and the relation of self-reported imagery abilities and memory task performance. Conscious. Cogn. 2023, 113, 103548. [Google Scholar] [CrossRef] [PubMed]
- Gulyas, E.; Gombos, F.; Sutori, S.; Lovas, A.; Ziman, G.; Kovacs, I. Visual imagery vividness declines across the lifespan. Cortex 2022, 154, 365–374. [Google Scholar] [CrossRef] [PubMed]
- Campos, A. Gender differences in imagery. Personal. Individ. Differ. 2014, 59, 107–111. [Google Scholar] [CrossRef]
- Aydin, Ç. Gender differences in visual imagery: Object and spatial imagery. Dokuz Eylül Üniversitesi Sos. Bilim. Enstitüsü Derg. 2020, 22, 1045–1064. [Google Scholar] [CrossRef]
- Smyth RS, D.; Acharya, P.; Hunt, N.P. Is visual imagery ability higher for orthodontic students than those in other disciplines? A cross-sectional questionnaire-based study. J. Orthod. 2019, 46, 205–211. [Google Scholar] [CrossRef]
- Fielding, D.; Kahui, V.; Wesselbaum, D. Visual imagination and the performance of undergraduate economics students. N. Z. Econ. Pap. 2020, 54, 127–137. [Google Scholar] [CrossRef]
- Blazhenkova, O.; Pechenkova, E. The two eyes of the blind mind: Object vs. spatial aphantasia? Russ. J. Cogn. Sci. 2019, 6, 51–65. [Google Scholar] [CrossRef]
- Blajenkova, O.; Kozhevnikov, M.; Motes, M.A. Object-spatial imagery: A new self-report imagery questionnaire. Appl. Cogn. Psychol. 2006, 20, 239–263. [Google Scholar] [CrossRef]
- Thorudottir, S.; Sigurdardottir, H.M.; Rice, G.E.; Kerry, S.J.; Robotham, R.J.; Leff, A.P.; Starrfelt, R. The architect who lost the ability to imagine: The cerebral basis of visual imagery. Brain Sci. 2020, 10, 59. [Google Scholar] [CrossRef]
- Knowles, L.; Jones, K.; Zeman, A. Acquired aphantasia in 88 cases: A preliminary report. J. Neurol. Neurosurg. Psychiatry 2021, 92, A6–A7. [Google Scholar] [CrossRef]
- Day, J.; Frayling, T.; Wood, A.; Zeman, A. 130 Does visual imagery vividness have a genetic basis? A genome-wide associa-tion study of 1019 individuals. J. Neurol. Neurosurg. Psychiatry 2022, 93, A51. [Google Scholar] [CrossRef]
- Bumgardner, A.L.; Yuan, K.; Chiu, A.V. I cannot picture it in my mind: Acquired aphantasia after autologous stem cell transplantation for multiple myeloma. Oxf. Med. Case Rep. 2021, 8, 158–160. [Google Scholar] [CrossRef] [PubMed]
- Gaber TA, K.; Eltemamy, M. Post-COVID-19 aphantasia. Prog. Neurol. Psychiatry 2021, 25, 16–17. [Google Scholar] [CrossRef]
- De Vito, S.; Bartolomeo, P. Refusing to imagine? on the possibility of psychogenic aphantasia. A commentary on Zeman et al. (2015). Cortex 2016, 74, 334–335. [Google Scholar] [CrossRef]
- Zeman, A.; Dewar, M.; Della Sala, S. Reflections on aphantasia. Cortex 2016, 74, 336–337. [Google Scholar] [CrossRef]
- Dance, C.J.; Ward, J.; Simner, J. What is the link between mental imagery and sensory sensitivity? insights from aphantasia. Perception 2021, 50, 757–782. [Google Scholar] [CrossRef]
- Takahashi, J.; Gyoba, J. A preliminary single-case study of aphantasia in Japan. Tohoku Psychol. Folia 2021, 79, 26–32. [Google Scholar]
- Bouyer, L.N.; Arnold, D.H. Deep Aphantasia: A visual brain with minimal influence from priors or inhibitory feedback? Front. Psychol. 2024, 15, 1374349. [Google Scholar] [CrossRef]
- Hinwar, R.P.; Lambert, A.J. Anauralia: The silent mind and its association with aphantasia. Front. Psychol. 2021, 12, 744213. [Google Scholar] [CrossRef]
- Wittmann, B.C.; Satirer, Y. Decreased associative processing and memory confidence in aphantasia. Learn. Mem. 2022, 29, 412–420. [Google Scholar] [CrossRef] [PubMed]
- Pounder, Z.; Eardley, A.F.; Loveday, C.; Evans, S. No clear evidence of a difference between individuals who self-report an absence of auditory imagery and typical imagers on auditory imagery tasks. PLoS ONE 2024, 19, e0300219. [Google Scholar] [CrossRef] [PubMed]
- Dawes, A.J.; Keogh, R.; Pearson, J. Multisensory subtypes of aphantasia: Mental imagery as supramodal perception in reverse. Neurosci. Res. 2024, 201, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Ganczarek, J.; Żurawska-Żyła, R.; Rolek, A. “I remember things, but I can’t picture them.” What can a case of aphantasia tell us about imagery and memory? Psychiatr. Psychol. Klin. 2020, 20, 134–141. [Google Scholar] [CrossRef]
- Keogh, R.; Wicken, M.; Pearson, J. Visual working memory in aphantasia: Retained accuracy and capacity with a different strategy. Cortex 2021, 143, 237–253. [Google Scholar] [CrossRef] [PubMed]
- Knight, K.F.; Milton, F. Memory without Imagery: No Evidence of Visual Working Memory Impairment in People with Aphantasia. In Proceedings of the Annual Meeting of the Cognitive Science Society, Toronto, ON, Canada, 27–30 July 2022; Volume 44. [Google Scholar]
- Pounder, Z.; Jacob, J.; Evans, S.; Loveday, C.; Eardley, A.F.; Silvanto, J. Only minimal differences between individuals with congenital aphantasia and those with typical imagery on neuropsychological tasks that involve imagery. Cortex 2022, 148, 180–192. [Google Scholar] [CrossRef]
- Dando, C.J.; Nahouli, Z.; Hart, A.; Pounder, Z. Real-world implications of aphantasia: Episodic recall of eyewitnesses with aphantasia is less complete but no less accurate than typical imagers. R. Soc. Open Sci. 2023, 10, 231007. [Google Scholar] [CrossRef]
- Dawes, A.J.; Keogh, R.; Robuck, S.; Pearson, J. Memories with a blind mind: Remembering the past and imagining the future with aphantasia. Cognition 2022, 227, 105192. [Google Scholar] [CrossRef]
- Monzel, M.; Vetterlein, A.; Reuter, M. Memory deficits in aphantasics are not restricted to autobiographical memory—Perspectives from the dual coding approach. J. Neuropsychol. 2022, 16, 444–461. [Google Scholar] [CrossRef]
- Watkins, N.W. (A)phantasia and severely deficient autobiographical memory: Scientific and personal perspectives. Cortex 2018, 105, 41–52. [Google Scholar] [CrossRef]
- Siena, M.J.; Simons, J. Metacognitive Awareness and the Subjective Experience of Remembering in Aphantasia. J. Cogn. Neurosci. 2024, 36, 1578–1598. [Google Scholar] [CrossRef] [PubMed]
- Kozhevnikov, M.; Kosslyn, S.; Shephard, J. Spatial versus object visualizers: A new characterization of visual cognitive style. Mem. Cogn. 2005, 33, 710–726. [Google Scholar] [CrossRef] [PubMed]
- Crowder, A. Differences in Spatial Visualization Ability and Vividness of Spatial Imagery between People with and without Aphantasia. Doctoral Dissertation, Virginia Commonwealth University, Richmond, VA, USA, 2018. [Google Scholar]
- Furman, M.; Fleitas-Rumak, P.; Lopez-Segura, P.; Furman, M.; Tafet, G.; de Erausquin, G.A.; Ortiz, T. Cortical activity involved in perception and imagery of visual stimuli in a subject with aphantasia. an EEG case report. Neurocase 2022, 28, 344–355. [Google Scholar] [CrossRef] [PubMed]
- Palermo, L.; Boccia, M.; Piccardi, L.; Nori, R. Congenital lack and extraordinary ability in object and spatial imagery: An investigation on sub-types of aphantasia and hyperphantasia. Conscious. Cogn. 2022, 103, 103360. [Google Scholar] [CrossRef] [PubMed]
- Keogh, R.; Pearson, J. The perceptual and phenomenal capacity of mental imagery. Cognition 2017, 162, 124–132. [Google Scholar] [CrossRef]
- Shepard, R.N.; Metzler, J. Mental rotation of three-dimensional objects. Science 1971, 171, 701–703. [Google Scholar] [CrossRef]
- Zhao, B.; Della Sala, S.; Zeman, A.; Gherri, E. Spatial transformation in mental rotation tasks in aphantasia. Psychon. Bull. Rev. 2022, 29, 2096–2107. [Google Scholar] [CrossRef]
- Kay, L.; Keogh, R.; Pearson, J. Slower but more accurate mental rotation performance in aphantasia linked to differences in cognitive strategies. Conscious. Cogn. 2024, 121, 103694. [Google Scholar] [CrossRef]
- Eardley, A.F.; Pring, L. Spatial processing, mental imagery, and creativity in individuals with and without sight. Eur. J. Cogn. Psychol. 2007, 19, 37–58. [Google Scholar] [CrossRef]
- Monzel, M.; Keidel, K.; Reuter, M. Imagine, and you will find—Lack of attentional guidance through visual imagery in aphantasics. Atten. Percept. Psychophys. 2021, 83, 2486–2497. [Google Scholar] [CrossRef]
- Monzel, M.; Reuter, M. Where’s Wanda? the influence of visual imagery vividness on visual search speed measured by means of hidden object pictures. Atten. Percept. Psychophys. 2024, 86, 22–27. [Google Scholar] [CrossRef] [PubMed]
- Pearson, J.; Clifford, C.W.; Tong, F. The functional impact of mental imagery on conscious perception. Curr. Biol. 2008, 18, 982–986. [Google Scholar] [CrossRef] [PubMed]
- Moriya, J. Visual mental imagery influences attentional guidance in a visual-search task. Atten. Percept. Psychophys. 2018, 80, 1127–1142. [Google Scholar] [CrossRef] [PubMed]
- Speed, L.J.; Eekhof, L.S.; Mak, M. The role of visual imagery in story reading: Evidence from aphantasia. Conscious. Cogn. 2024, 118, 103645. [Google Scholar] [CrossRef]
- Wicken, M.; Keogh, R.; Pearson, J. The critical role of mental imagery in human emotion: Insights from fear-based imagery and aphantasia. Proc. R. Soc. B-Biol. Sci. 2021, 288, 20210267. [Google Scholar] [CrossRef] [PubMed]
- Monzel, M.; Keidel, K.; Reuter, M. Is it really empathy? the potentially confounding role of mental imagery in self-reports of empathy. J. Res. Personal. 2023, 103, 104354. [Google Scholar] [CrossRef]
- Hashim, S.; Pulcini, C.; Jansari, A.; Küssner, M.B.; Omigie, D. The Experience of Music in Aphantasia: Emotion, Reward, and Everyday Functions. Music Sci. 2024, 7, 20592043231216259. [Google Scholar] [CrossRef]
- Keogh, R.; Wicken, M.; Pearson, J. Fewer intrusive memories in aphantasia: Using the trauma film paradigm as a laboratory model of PTSD. PsyArxiv 2023. [Google Scholar] [CrossRef]
- Monzel, M.; Dance, C.; Azañón, E.; Simner, J. Aphantasia within the framework of neurodivergence: Some preliminary data and the curse of the confidence gap. Conscious. Cogn. 2023, 115, 103567. [Google Scholar] [CrossRef]
- Cavedon-Taylor, D. Aphantasia and psychological disorder: Current connections, defining the imagery deficit and future directions. Front. Psychol. 2022, 13, 822989. [Google Scholar] [CrossRef]
- Sitek, E.J.; Konieczna, S. Does progressive aphantasia exist? the hypothetical role of aphantasia in the diagnosis of neurodegenerative diseases. Behav. Brain Sci. 2022, 45, 49–51. [Google Scholar] [CrossRef] [PubMed]
- Pearson, J.; Naselaris, T.; Holmes, E.A.; Kosslyn, S.M. Mental imagery: Functional mechanisms and clinical applications. Trends Cogn. Sci. 2015, 19, 590–602. [Google Scholar] [CrossRef] [PubMed]
- Yehuda, R.; Hoge, C.W.; McFarlane, A.C.; Vermetten, E.; Lanius, R.A.; Nievergelt, C.M.; Hobfoll, S.E.; Koenen, K.C.; Neylan, T.C.; Hyman, S.E. Post-traumatic stress disorder. Nat. Rev. Dis. Primers 2015, 1, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Dance, C.J.; Jaquiery, M.; Eagleman, D.M.; Porteous, D.; Zeman, A.; Simner, J. What is the relationship between aphantasia, synaesthesia and autism? Conscious. Cogn. 2021, 89, 103087. [Google Scholar] [CrossRef]
- Crespi, B.; Leach, E.; Dinsdale, N.; Mokkonen, M.; Hurd, P. Imagination in human social cognition 2021, autism, and psychotic-affective conditions. Cognition 2016, 150, 181–199. [Google Scholar] [CrossRef]
- Lord, C.; Elsabbagh, M.; Baird, G.; Veenstra-Vanderweele, J. Autism spectrum disorder. Lancet 2018, 392, 508–520. [Google Scholar] [CrossRef]
- Dance, C.J.; Hole, G.; Simner, J. The role of visual imagery in face recognition and the construction of facial composites. Evidence from Aphantasia. Cortex 2023, 167, 318–334. [Google Scholar] [CrossRef]
- Liu, J.; Bartolomeo, P. Probing the unimaginable: The impact of aphantasia on distinct domains of visual mental imagery and visual perception. Cortex 2023, 166, 338–347. [Google Scholar] [CrossRef]
- Svart, N.; Starrfelt, R. Is it just face blindness? exploring developmental comorbidity in individuals with Self-Reported developmental prosopagnosia. Brain Sci. 2022, 12, 230. [Google Scholar] [CrossRef]
- Monzel, M.; Vetterlein, A.; Hogeterp, S.A.; Reuter, M. No increased prevalence of prosopagnosia in aphantasia: Visual recognition deficits are small and not restricted to faces. Perception 2023, 52, 03010066231180712. [Google Scholar] [CrossRef]
- Spagna, A.; Hajhajate, D.; Liu, J.; Bartolomeo, P. Visual mental imagery engages the left fusiform gyrus, but not the early visual cortex: A meta-analysis of neuroimaging evidence. Neurosci. Biobehav. Rev. 2021, 122, 201–217. [Google Scholar] [CrossRef] [PubMed]
- Dijkstra, N. Uncovering the Role of the Early Visual Cortex in Visual Mental Imagery. Vision 2024, 8, 29. [Google Scholar] [CrossRef] [PubMed]
- Monzel, M.; Leelaarporn, P.; Lutz, T.; Schultz, J.; Brunheim, S.; Reuter, M.; McCormick, C. Hippocampal-occipital connectivity reflects autobiographical memory deficits in aphantasia. bioRxiv 2023. [Google Scholar] [CrossRef]
- Dupont, W.; Papaxanthis, C.; Madden-Lombardi, C.; Lebon, F. Explicit and implicit motor simulations are impaired in individuals with aphantasia. Brain Commun. 2024, 6, fcae072. [Google Scholar] [CrossRef] [PubMed]
- Dupont, W.; Papaxanthis, C.; Lebon, F.; Madden-Lombardi, C. Mental simulations and action language are impaired in individuals with aphantasia. J. Cogn. Neurosci. 2024, 36, 261–271. [Google Scholar] [CrossRef]
- Bartolomeo, P. The relationship between visual perception and visual mental imagery: A reappraisal of the neuropsychological evidence. Cortex 2002, 38, 357–378. [Google Scholar] [CrossRef]
- Bartolomeo, P.; Hajhajate, D.; Liu, J.; Spagna, A. Assessing the causal role of early visual areas in visual mental imagery. Nat. Rev. Neurosci. 2020, 21, 517. [Google Scholar] [CrossRef]
- Bartolomeo, P. Visual agnosia and imagery after Lissauer. Brain 2021, 144, 2557–2559. [Google Scholar] [CrossRef]
- Bartolomeo, P.; Bachoud-Lévi, A.C.; De Gelder, B.; Denes, G.; Dalla Barba, G.; Brugières, P.; Degos, J.D. Multiple-domain dissociation between impaired visual perception and preserved mental imagery in a patient with bilateral extrastriate lesions. Neuropsychologia 1998, 36, 239–249. [Google Scholar] [CrossRef]
- Bartolomeo, P. The neural correlates of visual mental imagery: An ongoing debate. Cortex 2008, 44, 107–108. [Google Scholar] [CrossRef]
- Moro, V.; Berlucchi, G.; Lerch, J.; Tomaiuolo, F.; Aglioti, S.M. Selective deficit of mental visual imagery with intact primary visual cortex and visual perception. Cortex 2008, 44, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Riley, S.N.; Davies, J. Vividness as the similarity between generated imagery and an internal model. Brain Cogn. 2023, 169, 105988. [Google Scholar] [CrossRef] [PubMed]
- Arnold, D.H.; Andresen, I.; Anderson, N.; Saurels, B.W. Commonalities between the berger rhythm and spectra differences driven by cross-modal attention and imagination. Conscious. Cogn. 2023, 107, 103436. [Google Scholar] [CrossRef] [PubMed]
- Zeman, A.Z.; Della Sala, S.; Torrens, L.A.; Gountouna, V.E.; McGonigle, D.J.; Logie, R.H. Loss of imagery phenomenology with intact visuo-spatial task performance: A case of ‘blind imagination’. Neuropsychologia 2010, 48, 145–155. [Google Scholar] [CrossRef] [PubMed]
- Hajhajate, D.; Kaufmann, B.C.; Liu, J.; Siuda-Krzywicka, K.; Bartolomeo, P. The connectional anatomy of visual mental imagery: Evidence from a patient with left occipito-temporal damage. Brain Struct. Funct. 2022, 227, 3075–3083. [Google Scholar] [CrossRef]
- Spagna, A.; Heidenry, Z.; Miselevich, M.; Lambert, C.; Eisenstadt, B.E.; Tremblay, L.; Liu, Z.; Liu, J.; Bartolomeo, P. Visual mental imagery: Evidence for a heterarchical neural architecture. Phys. Life Rev. 2024, 48, 113–131. [Google Scholar] [CrossRef]
- Liu, J.; Zhan, M.; Hajhajate, D.; Spagna, A.; Dehaene, S.; Cohen, L.; Bartolomeo, P. Ultra-high field fMRI of visual mental imagery in typical imagers and aphantasic individuals. bioRxiv 2023. [Google Scholar] [CrossRef]
- Boccia, M.; Sulpizio, V.; Bencivenga, F.; Guariglia, C.; Galati, G. Neural representations underlying mental imagery as unveiled by representation similarity analysis. Brain Struct. Funct. 2021, 226, 1511–1531. [Google Scholar] [CrossRef]
- McCarthy, G.; Puce, A.; Gore, J.C.; Allison, T. Face-specific processing in the human fusiform gyrus. J. Cogn. Neurosci. 1997, 9, 605–610. [Google Scholar] [CrossRef]
- Pearson, J. Reply to: Assessing the causal role of early visual areas in visual mental imagery. Nat. Rev. Neurosci. 2020, 21, 517–518. [Google Scholar] [CrossRef]
- Kosslyn, S.M.; Pascual-Leone, A.; Felician, O.; Camposano, S.; Keenan, J.P.; Thompson, W.L.; Ganis, G.; Sukel, K.E.; Alpert, N.M. The role of area 17 in visual imagery: Convergent evidence from PET and rTMS. Science 1999, 284, 167–170. [Google Scholar] [CrossRef] [PubMed]
- Bergmann, J.; Genç, E.; Kohler, A.; Singer, W.; Pearson, J. Smaller primary visual cortex is associated with stronger, but less precise mental imagery. Cereb. Cortex 2016, 26, 3838–3850. [Google Scholar] [CrossRef] [PubMed]
- Keogh, R.; Bergmann, J.; Pearson, J. Cortical excitability controls the strength of mental imagery. eLife 2020, 9, e50232. [Google Scholar] [CrossRef] [PubMed]
- Meng, M.; Chang, S.; Zhang, X.; Pearson, J. Imageless imagery in aphantasia: Decoding non-sensory imagery in aphantasia. Res. Sq. 2023; preprint. [Google Scholar] [CrossRef]
- Cabbai, G.; Racey, C.; Simner, J.; Dance, C.; Ward, J.; Forster, S. Sensory representations in primary visual cortex are not sufficient for subjective imagery. bioRxiv 2024. [Google Scholar] [CrossRef]
- Arcangeli, M. Aphantasia demystified. Synthese 2023, 201, 31. [Google Scholar] [CrossRef]
- Monzel, M.; Handlogten, J.; Reuter, M. No verbal overshadowing in aphantasia: The role of visual imagery for the verbal overshadowing effect. Cognition 2024, 245, 105732. [Google Scholar] [CrossRef]
- Park, T. Why successful performance in imagery tasks does not require the manipulation of mental imagery. Avant 2019, 10, 1–11. [Google Scholar] [CrossRef]
- Toftness, A.R. Clarifying Aphantasia. Doctoral Dissertation, Iowa State University, Ames, IA, USA, 2022. [Google Scholar]
- Purkart, R.; Delem, M.; Cavalli, E.; Ranson, V.; Andrey, C.; Versace, R.; Plancher, G. Are There Unconscious Visual Images in Aphantasia? Development of an Implicit Priming Paradigm. Available online: https://ssrn.com/abstract=4755823 (accessed on 10 September 2024).
- Perrin, D.; Michaelian, K. Memory as mental time travel. In The Routledge Handbook of Philosophy of Memory; Routledge: New York, NY, USA, 2017; pp. 228–239. [Google Scholar]
- Ungerleider, L.G.; Haxby, J.V. ‘What’and ‘where’in the human brain. Curr. Opin. Neurobiol. 1994, 4, 157–165. [Google Scholar] [CrossRef]
- Goodale, M.A.; Milner, A.D. Separate visual pathways for perception and action. Trends Neurosci. 1992, 15, 20–25. [Google Scholar] [CrossRef]
- Milner, A.D.; Goodale, M.A. Two visual systems re-viewed. Neuropsychologia 2008, 46, 774–785. [Google Scholar] [CrossRef]
- de Haan, E.H.F.; Jackson, S.R.; Schenk, T. Where are we now with ‘What’ and ‘How’? Cortex 2018, 98, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Grünbaum, T. The two visual systems hypothesis and contrastive underdetermination. Synthese 2021, 198 (Suppl. S17), S4045–S4068. [Google Scholar] [CrossRef]
- Bergmann, J.; Ortiz-Tudela, J. Feedback signals in visual cortex during episodic and schematic memory retrieval and their potential implications for aphantasia. Neurosci. Biobehav. Rev. 2023, 152, 105335. [Google Scholar] [CrossRef] [PubMed]
- Suica, Z.; Behrendt, F.; Gäumann, S.; Gerth, U.; Schmidt-Trucksäss, A.; Ettlin, T.; Schuster-Amft, C. Imagery ability assessments: A cross-disciplinary systematic review and quality evaluation of psychometric properties. BMC Med. 2022, 20, 166. [Google Scholar] [CrossRef] [PubMed]
- Schwarzkopf, D.S. What is the true range of mental imagery? Cortex 2024, 170, 21–25. [Google Scholar] [CrossRef]
- Larner, A.J.; Leff, A.P.; Nachev, P.C. Phantasia, aphantasia, and hyperphantasia: Empirical data and conceptual considerations. Neurosci. Biobehav. Rev. 2024, 164, 105819. [Google Scholar] [CrossRef]
- Monzel, M.; Mitchell, D.; Macpherson, F.; Pearson, J.; Zeman, A. Proposal for a consistent definition of aphantasia and hyperphantasia: A response to Lambert and Sibley (2022) and Simner and Dance (2022). Cortex 2022, 152, 74–76. [Google Scholar] [CrossRef]
- Lambert, A.J.; Sibley, C.G. On the importance of consistent terminology for describing sensory imagery and its absence: A response to Monzel et al.(2022). Cortex 2022, 152, 153–156. [Google Scholar] [CrossRef]
- Simner, J.; Dance, C.J. Dysikonesia or aphantasia? understanding the impact and history of names. A reply to monzel et al. (2022). Cortex 2022, 153, 220–223. [Google Scholar] [CrossRef]
- D’Angiulli, A. Vividness, Consciousness and Mental Imagery: A Start on Connecting the Dots. Brain Sci. 2020, 10, 500. [Google Scholar] [CrossRef]
- Küssner, M.B.; Taruffi, L. Modalities and causal routes in music-induced mental imagery. Trends Cogn. Sci. 2023, 27, 114–115. [Google Scholar] [CrossRef] [PubMed]
- Lau, H.; Michel, M.; LeDoux, J.E.; Fleming, S.M. The mnemonic basis of subjective experience. Nat. Rev. Psychol. 2022, 1, 479–488. [Google Scholar] [CrossRef]
- Muraki, E.J.; Speed, L.J.; Pexman, P.M. Insights into embodied cognition and mental imagery from aphantasia. Nat. Rev. Psychol. 2023, 2, 591–605. [Google Scholar] [CrossRef]
- Cohen, M.X. Where does EEG come from and what does it mean? Trends Neurosci. 2017, 40, 208–218. [Google Scholar] [CrossRef] [PubMed]
- Baillet, S. Magnetoencephalography for brain electrophysiology and imaging. Nat. Neurosci. 2017, 20, 327–339. [Google Scholar] [CrossRef] [PubMed]
- Xie, S.; Kaiser, D.; Cichy, R.M. Visual imagery and perception share neural representations in the alpha frequency band. Curr. Biol. 2020, 30, 2621–2627. [Google Scholar] [CrossRef]
- Cichy, R.M.; Oliva, A. A M/EEG-fMRI fusion primer: Resolving human brain responses in space and time. Neuron 2020, 107, 772–781. [Google Scholar] [CrossRef]
- Kriegeskorte, N.; Mur, M.; Bandettini, P.A. Representational similarity analysis-connecting the branches of systems neuroscience. Front. Syst. Neurosci. 2008, 2, 249. [Google Scholar] [CrossRef]
- Haynes, J.D. A primer on pattern-based approaches to fMRI: Principles, pitfalls, and perspectives. Neuron 2015, 87, 257–270. [Google Scholar] [CrossRef]
- Polanía, R.; Nitsche, M.A.; Ruff, C.C. Studying and modifying brain function with non-invasive brain stimulation. Nat. Neurosci. 2018, 21, 174–187. [Google Scholar] [CrossRef]
- Monzel, M.; Karneboge, J.; Reuter, M. The role of dopamine in visual imagery—An experimental pharmacological study. J. Neurosci. Res. 2024, 102, e25262. [Google Scholar] [CrossRef] [PubMed]
- Fried, E.I. Theories and models: What they are, what they are for, and what they are about. Psychol. Inq. 2020, 31, 336–344. [Google Scholar] [CrossRef]
- Gigerenzer, G. Personal reflections on theory and psychology. Theory Psychol. 2010, 20, 733–743. [Google Scholar] [CrossRef]
- Cavedon-Taylor, D. Mental imagery: Pulling the plug on perceptualism. Philos. Stud. 2021, 178, 3847–3868. [Google Scholar] [CrossRef]
- Cavedon-Taylor, D. Untying the knot: Imagination, perception and their neural substrates. Synthese 2021, 199, 7203–7230. [Google Scholar] [CrossRef]
- Cavedon-Taylor, D. Predictive processing and perception: What does imagining have to do with it? Conscious. Cogn. 2022, 106, 103419. [Google Scholar] [CrossRef]
- D’Aloisio-Montilla, N. Imagery and overflow: We see more than we report. Philos. Psychol. 2017, 30, 545–570. [Google Scholar] [CrossRef]
- Duch, W. Imagery agnosia and its phenomenology. Ann. Psychol. 2021, 24, 307–323. [Google Scholar] [CrossRef]
- Fox-Muraton, M. Aphantasia and the language of imagination: A wittgensteinian exploration. Anal. I Egzystencja 2021, 55, 5–24. [Google Scholar] [CrossRef]
- Fox-Muraton, M. A world without imagination? consequences of aphantasia for an existential account of self. Hist. Eur. Ideas 2021, 47, 414–428. [Google Scholar] [CrossRef]
- Lorenzatti, J.J. Aphantasia: A philosophical approach. Philos. Psychol. 2023, 1–29. [Google Scholar] [CrossRef]
- MacKisack, M.; Aldworth, S.; Macpherson, F.; Onians, J.; Winlove, C.; Zeman, A. Plural imagination: Diversity in mind and making. Art J. 2022, 81, 70–87. [Google Scholar] [CrossRef]
- Whiteley CM, K. Aphantasia, imagination and dreaming. Philos. Stud. 2021, 178, 2111–2132. [Google Scholar] [CrossRef]
- Open Science Collaboration. Estimating the reproducibility of psychological science. Science 2015, 349, aac4716. [Google Scholar] [CrossRef] [PubMed]
- Tackett, J.L.; Brandes, C.M.; King, K.M.; Markon, K.E. Psychology’s replication crisis and clinical psychological science. Annu. Rev. Clin. Psychol. 2019, 15, 579–604. [Google Scholar] [CrossRef] [PubMed]
- Nosek, B.A.; Hardwicke, T.E.; Moshontz, H.; Allard, A.; Corker, K.S.; Dreber, A.; Fidler, F.; Hilgard, J.; Struhl, M.K.; Nuijten, M.B.; et al. Replicability, robustness, and reproducibility in psychological science. Annu. Rev. Psychol. 2022, 73, 719–748. [Google Scholar] [CrossRef] [PubMed]
- Miłkowski, M.; Hensel, W.M.; Hohol, M. Replicability or reproducibility? On the replication crisis in computational neuroscience and sharing only relevant detail. J. Comput. Neurosci. 2018, 45, 163–172. [Google Scholar] [CrossRef]
- Balas, B. Mid-level Characteristics of Drawings Made by Observers with Aphantasia. J. Percept. Imaging 2024, 7, 1–8. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jin, F.; Hsu, S.-M.; Li, Y. A Systematic Review of Aphantasia: Concept, Measurement, Neural Basis, and Theory Development. Vision 2024, 8, 56. https://doi.org/10.3390/vision8030056
Jin F, Hsu S-M, Li Y. A Systematic Review of Aphantasia: Concept, Measurement, Neural Basis, and Theory Development. Vision. 2024; 8(3):56. https://doi.org/10.3390/vision8030056
Chicago/Turabian StyleJin, Feiyang, Shen-Mou Hsu, and Yu Li. 2024. "A Systematic Review of Aphantasia: Concept, Measurement, Neural Basis, and Theory Development" Vision 8, no. 3: 56. https://doi.org/10.3390/vision8030056
APA StyleJin, F., Hsu, S.-M., & Li, Y. (2024). A Systematic Review of Aphantasia: Concept, Measurement, Neural Basis, and Theory Development. Vision, 8(3), 56. https://doi.org/10.3390/vision8030056