
The Functional Network of the Visual Cortex Is Altered in Migraine


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
3. Results
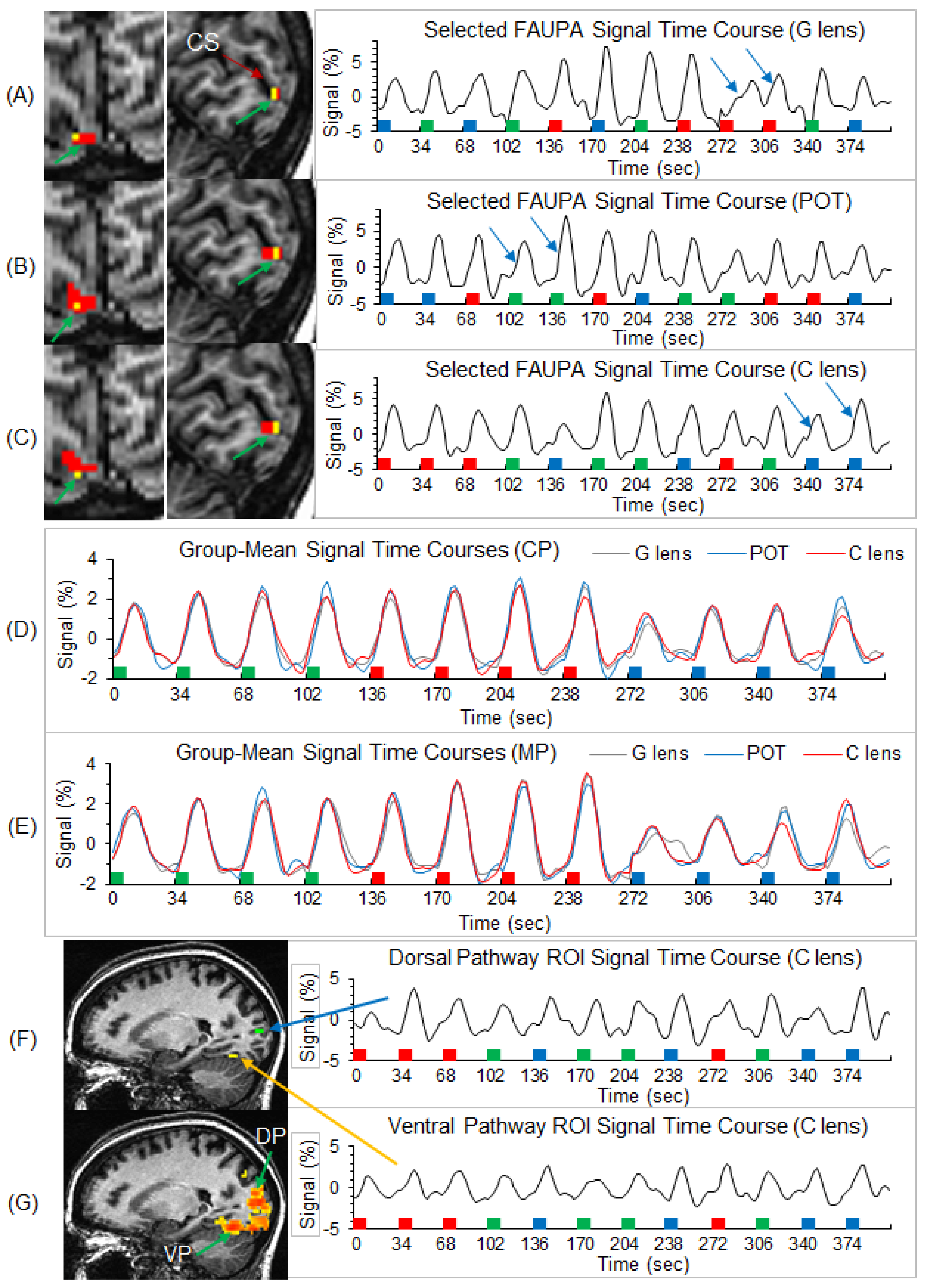
3.1. Identification of Stimulation-Associated FAUPAs
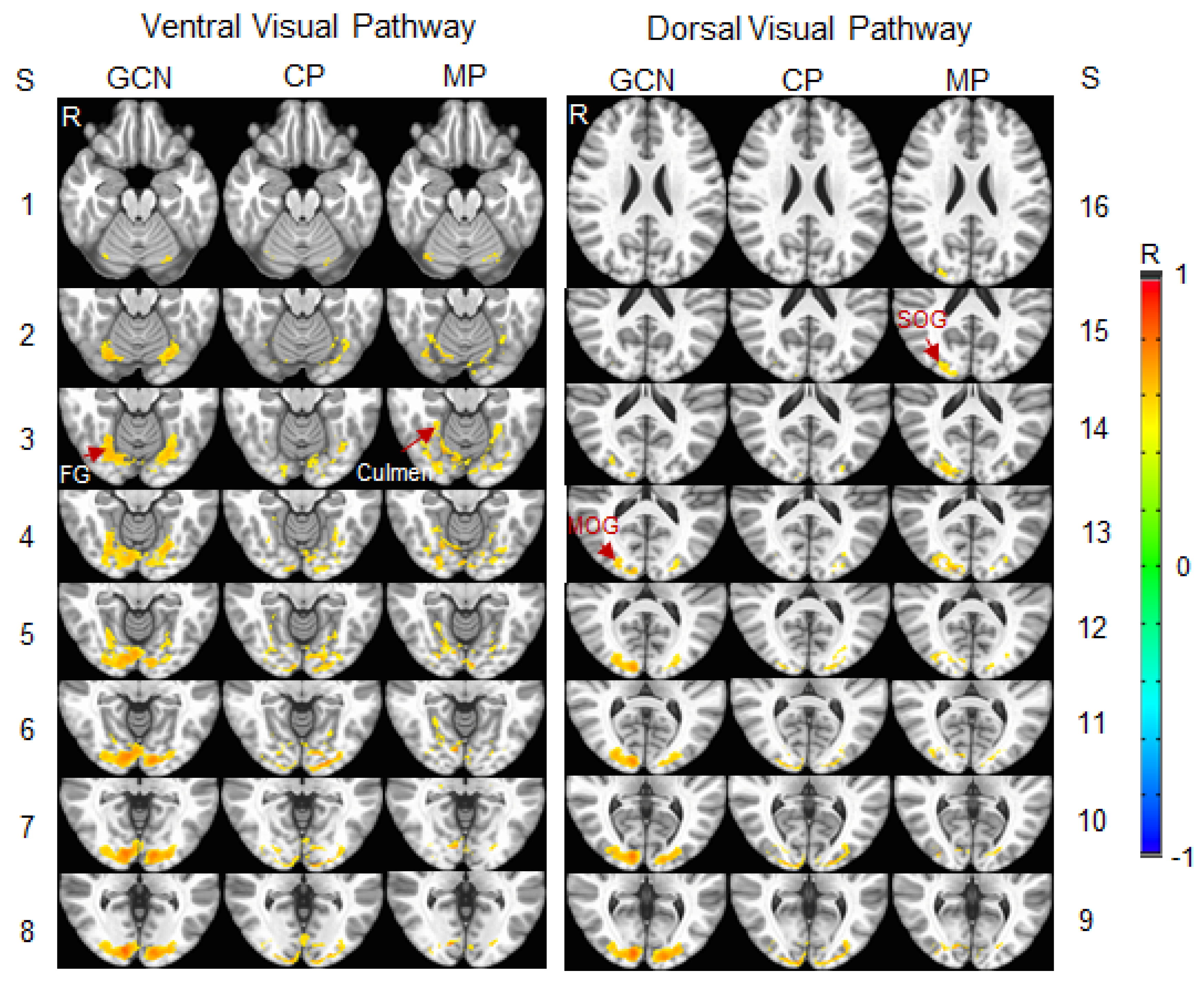
3.2. Stimulation-Evoked Visual Cortical Functional Network
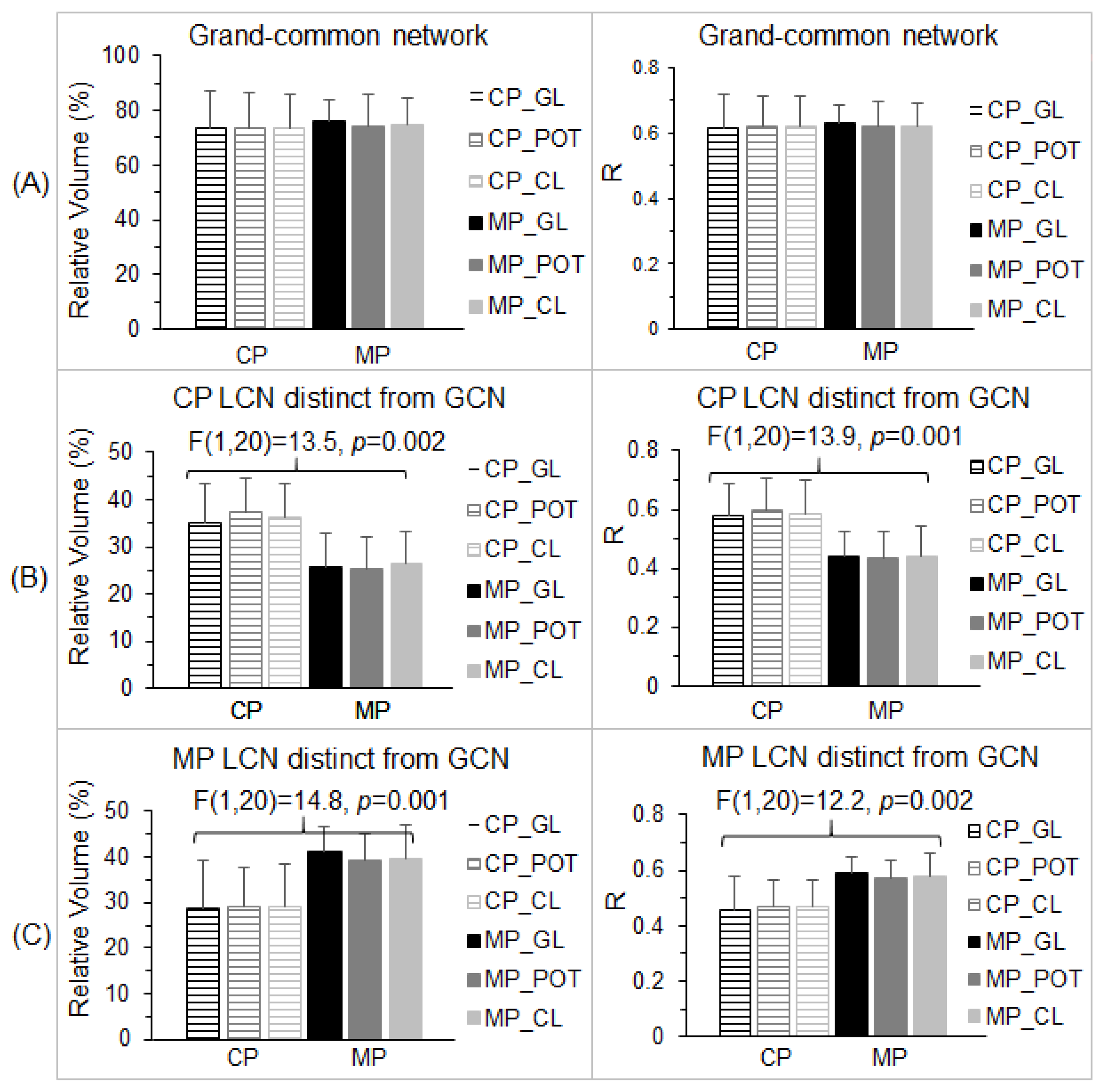
3.3. Altered Visual Functional Network in Migraine
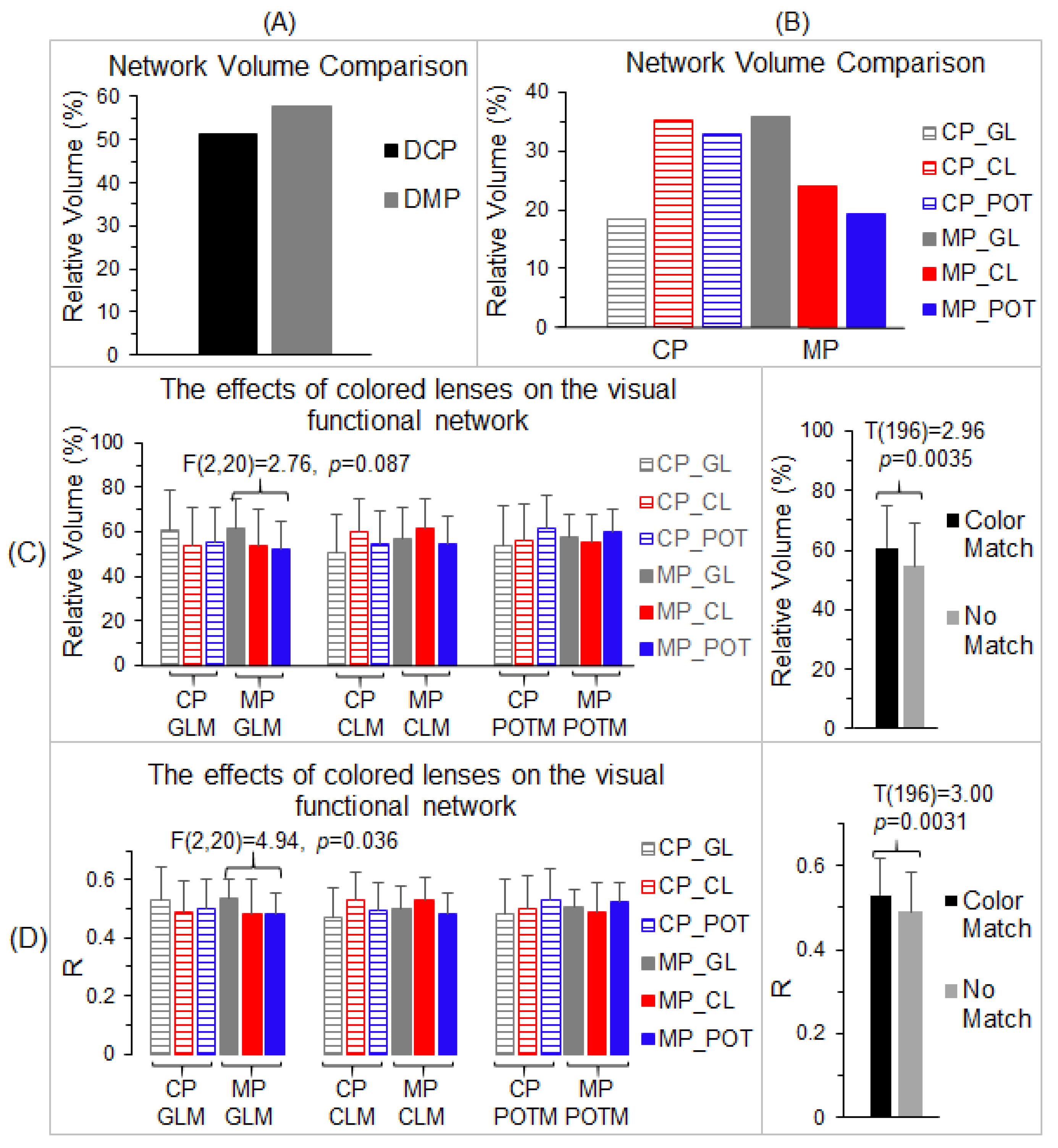
3.4. The Effects of Color on the Visual Functional Network
4. Discussion and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Olesen, J.; Ramadan, N.; Tfelt-Hansen, P.; Welch, K. The Headaches, 3rd ed.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2006. [Google Scholar]
- Marcus, D.A.; Soso, M.J. Migraine and Stripe-Induced Visual Discomfort. Arch. Neurol. 1989, 46, 1129–1132. [Google Scholar] [CrossRef]
- Wilkins, A.; Nimmo-Smith, I.; Tait, A.; McManus, C.; Della Sala, S.; Tilley, A.; Arnold, K.; Barrie, M.; Scott, S. A neurological basis for visual discomfort. Brain 1984, 107 Pt 4, 989–1017. [Google Scholar] [CrossRef] [PubMed]
- Wilkins, A. Visual Stress; Oxford University Press: Oxford, UK, 1995. [Google Scholar]
- Hay, K.M.; Mortimer, M.J.; Barker, D.C.; Debney, L.M.; Good, P.A. 1044 women with migraine: The effect of environmental stimuli. Headache 1994, 34, 166–168. [Google Scholar] [CrossRef]
- Harle, D.E.; Shepherd, A.J.; Evans, B.J. Visual stimuli are common triggers of migraine and are associated with pattern glare. Headache 2006, 46, 1431–1440. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, S.; Lee, T.M.; Kay, A.R.; Tank, D.W. Brain magnetic resonance imaging with contrast dependent on blood oxygenation. Proc. Natl. Acad. Sci. USA 1990, 87, 9868–9872. [Google Scholar] [CrossRef] [Green Version]
- Kwong, K.K.; Belliveau, J.W.; Chesler, D.A.; Goldberg, I.E.; Weisskoff, R.M.; Poncelet, B.P.; Kennedy, D.N.; Hoppel, B.E.; Cohen, M.S.; Turner, R. Dynamic magnetic resonance imaging of human brain activity during primary sensory stimulation. Proc. Natl. Acad. Sci. USA 1992, 89, 5675–5679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Logothetis, N.K. What we can do and what we cannot do with fMRI. Nature 2008, 453, 869–878. [Google Scholar] [CrossRef]
- Puledda, F.; Ffytche, D.; O’Daly, O.; Goadsby, P.J. Imaging the Visual Network in the Migraine Spectrum. Front. Neurol. 2019, 10, 1325. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Cooper, T.G.; Satana, B.; Kaufman, D.I.; Cao, Y. Visual distortion provoked by a stimulus in migraine associated with hyperneuronal activity. Headache 2003, 43, 664–671. [Google Scholar] [CrossRef]
- Vincent, M.; Pedra, E.; Mourão-Miranda, J.; Bramati, I.E.; Henrique, A.R.; Moll, J. Enhanced Interictal Responsiveness of the Migraineous Visual Cortex to Incongruent Bar Stimulation: A Functional MRI Visual Activation Study. Cephalalgia 2003, 23, 860–868. [Google Scholar] [CrossRef]
- Datta, R.; Aguirre, G.K.; Hu, S.; Detre, J.A.; Cucchiara, B. Interictal cortical hyperresponsiveness in migraine is directly related to the presence of aura. Cephalalgia 2013, 33, 365–374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, J.; Zong, X.; Wilkins, A.; Jenkins, B.; Bozoki, A.; Cao, Y. fMRI evidence that precision ophthalmic tints reduce cortical hyperactivation in migraine. Cephalalgia 2011, 31, 925–936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griebe, M.; Flux, F.; Wolf, M.E.; Hennerici, M.G.; Szabo, K. Multimodal Assessment of Optokinetic Visual Stimulation Response in Migraine With Aura. Headache 2014, 54, 131–141. [Google Scholar] [CrossRef] [PubMed]
- Hougaard, A.; Amin, F.M.; Hoffmann, M.B.; Rostrup, E.; Larsson, H.B.; Asghar, M.S.; Larsen, V.A.; Olesen, J.; Ashina, M. Interhemispheric differences of fMRI responses to visual stimuli in patients with side-fixed migraine aura. Hum. Brain Mapp. 2014, 35, 2714–2723. [Google Scholar] [CrossRef]
- Cao, Y.; Welch, K.M.A.; Aurora, S.; Vikingstad, E.M. Functional MRI-BOLD of Visually Triggered Headache in Patients with Migraine. Arch. Neurol. 1999, 56, 548–554. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.; Aurora, S.K.; Nagesh, V.; Patel, S.C.; Welch, K.M. Functional MRI-BOLD of brainstem structures during visually triggered migraine. Neurology 2002, 59, 72–78. [Google Scholar] [CrossRef]
- Hadjikhani, N.; Del Rio, M.S.; Wu, O.; Schwartz, D.; Bakker, D.; Fischl, B.; Kwong, K.K.; Cutrer, F.M.; Rosen, B.R.; Tootell, R.B.H.; et al. Mechanisms of migraine aura revealed by functional MRI in human visual cortex. Proc. Natl. Acad. Sci. USA 2001, 98, 4687–4692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arngrim, N.; Hougaard, A.; Ahmadi, K.; Vestergaard, M.B.; Schytz, H.W.; Amin, F.M.; Larsson, H.B.W.; Olesen, J.; Hoffmann, M.B.; Ashina, M. Heterogenous migraine aura symptoms correlate with visual cortex functional magnetic resonance imaging responses. Ann. Neurol. 2017, 82, 925–939. [Google Scholar] [CrossRef]
- Descamps, B.; Vandemaele, P.; Reyngoudt, H.; Deblaere, K.; Leybaert, L.; Paemeleire, K.; Achten, E. Absence of haemodynamic refractory effects in patients with migraine without aura–an interictal fMRI study. Cephalalgia 2011, 31, 1220–1231. [Google Scholar] [CrossRef]
- Raichle, M.E. Two views of brain function. Trends Cogn. Sci. 2010, 14, 180–190. [Google Scholar] [CrossRef]
- Biswal, B.; Yetkin, F.Z.; Haughton, V.M.; Hyde, J.S. Functional connectivity in the motor cortex of resting human brain using echo-planar mri. Magn. Reson. Med. 1995, 34, 537–541. [Google Scholar] [CrossRef] [PubMed]
- Raichle, M.E. The Restless Brain. Brain Connect. 2011, 1, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Yeo, B.T.; Krienen, F.M.; Sepulcre, J.; Sabuncu, M.; Lashkari, D.; Hollinshead, M.; Roffman, J.; Smoller, J.W.; Zöllei, L.; Polimeni, J.; et al. The organization of the human cerebral cortex estimated by intrinsic functional connectivity. J. Neurophysiol. 2011, 106, 1125–1165. [Google Scholar] [CrossRef] [PubMed]
- Mainero, C.; Bs, J.B.; Hadjikhani, N. Altered functional magnetic resonance imaging resting-state connectivity in periaqueductal gray networks in migraine. Ann. Neurol. 2011, 70, 838–845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russo, A.; Tessitore, A.; Giordano, A.; Corbo, D.; Marcuccio, L.; De Stefano, M.; Salemi, F.; Conforti, R.; Esposito, F.; Tedeschi, G. Executive resting-state network connectivity in migraine without aura. Cephalalgia 2012, 32, 1041–1048. [Google Scholar] [CrossRef]
- Xue, T.; Yuan, K.; Zhao, L.; Yu, D.H.; Zhao, L.M.; Dong, T.; Cheng, P.; von Deneen, K.M.; Qin, W.; Tian, J. Intrinsic Brain Network Abnormalities in Migraines without Aura Revealed in Resting-State fMRI. PLoS ONE 2012, 7, e52927. [Google Scholar] [CrossRef] [Green Version]
- Tessitore, A.; Russo, A.; Giordano, A.; Conte, F.; Corbo, D.; De Stefano, M.; Cirillo, S.; Cirillo, M.; Esposito, F.; Tedeschi, G. Disrupted default mode network connectivity in migraine without aura. J. Headache Pain 2013, 14, 89. [Google Scholar] [CrossRef] [Green Version]
- Niddam, D.M.; Lai, K.-L.; Fuh, J.-L.; Chuang, C.-Y.N.; Chen, W.-T.; Wang, S.-J. Reduced functional connectivity between salience and visual networks in migraine with aura. Cephalalgia 2016, 36, 53–66. [Google Scholar] [CrossRef] [Green Version]
- Amin, F.M.; Hougaard, A.; Magon, S.; Asghar, M.S.; Ahmad, N.N.; Rostrup, E.; Sprenger, T.; Ashina, M. Change in brain network connectivity during PACAP38-induced migraine attacks. Neurology 2015, 86, 180–187. [Google Scholar] [CrossRef]
- Tedeschi, G.; Russo, A.; Conte, F.; Corbo, D.; Caiazzo, G.; Giordano, A.; Conforti, R.; Esposito, F.; Tessitore, A. Increased interictal visual network connectivity in patients with migraine with aura. Cephalalgia 2015, 36, 139–147. [Google Scholar] [CrossRef]
- Coppola, G.; di Renzo, A.; Tinelli, E.; Lepre, C.; Di Lorenzo, C.; Di Lorenzo, G.; Scapeccia, M.; Parisi, V.; Serrao, M.; Colonnese, C.; et al. Thalamo-cortical network activity between migraine attacks: Insights from MRI-based microstructural and functional resting-state network correlation analysis. J. Headache Pain 2016, 17, 100. [Google Scholar] [CrossRef] [PubMed]
- Hougaard, A.; Amin, F.M.; Larsson, H.B.; Rostrup, E.; Ashina, M. Increased intrinsic brain connectivity between pons and somatosensory cortex during attacks of migraine with aura. Hum. Brain Mapp. 2017, 38, 2635–2642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farago, P.; Tuka, B.; Toth, E.; Szabo, N.; Kiraly, A.; Csete, G.; Szok, D.; Tajti, J.; Pardutz, A.; Vecsei, L.; et al. Interictal brain activity differs in migraine with and without aura: Resting state fMRI study. J. Headache Pain 2017, 18, 8. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Lan, L.; Zeng, F.; Makris, N.; Hwang, J.; Guo, T.; Wu, F.; Gao, Y.; Dong, M.; Liu, M.; et al. The altered right frontoparietal network functional connectivity in migraine and the modulation effect of treatment. Cephalalgia 2017, 37, 161–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Su, J.; Wang, M.; Zhao, Y.; Zhang, Q.-T.; Yao, Q.; Lu, H.; Zhang, H.; Li, G.-F.; Wu, Y.-L.; et al. The sensorimotor network dysfunction in migraineurs without aura: A resting-state fMRI study. J. Neurol. 2017, 264, 654–663. [Google Scholar] [CrossRef]
- Lisicki, M.; D’Ostilio, K.; Coppola, G.; de Noordhout, A.M.; Parisi, V.; Schoenen, J.; Magis, D. Increased functional connectivity between the right temporo-parietal junction and the temporal poles in migraine without aura. Cephalalgia Rep. 2018, 1, 4823. [Google Scholar] [CrossRef] [Green Version]
- Xiong, Z.; Tian, C.; Zeng, X.; Huang, J.; Wang, R. The Relationship of Functional Connectivity of the Sensorimotor and Visual Cortical Networks Between Resting and Task States. Front. Neurosci. 2020, 14, 2720. [Google Scholar] [CrossRef]
- Huang, J. Human brain functional areas of unitary pooled activity discovered with fMRI. Sci. Rep. 2018, 8, 2388. [Google Scholar] [CrossRef] [PubMed]
- Huang, J. Method and System for Determining Brain-State Dependent Functional Areas of Unitary Pooled Activity and Associated Dynamic Networks with Functional Magnetic Resonance Imaging. U.S. Patent and Trademark Office US-20,200,069,237, 5 March 2020. [Google Scholar]
- Huang, J. Dynamic activity of human brain task-specific networks. Sci. Rep. 2020, 10, 7851. [Google Scholar] [CrossRef]
- Huang, J.; Beach, P.; Bozoki, A.; Zhu, D.C. Alzheimer’s Disease Progressively Alters the Face-Evoked Visual-Processing Network. J. Alzheimer’s Dis. 2020, 77, 1025–1042. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Beach, P.; Bozoki, A.; Zhu, D.C. Alzheimer’s Disease Progressively Reduces Visual Functional Network Connectivity. J. Alzheimer’s Dis. Rep. 2021, 5, 549–562. [Google Scholar] [CrossRef] [PubMed]
- Headache Classification Committee of the International Headache Society (IHS). The International Classification of Headache Disorders, 3rd edition (beta version). Cephalalgia 2013, 33, 629–808. [Google Scholar] [CrossRef] [Green Version]
- Wilkins, A.; Milroy, R.; Nimmo-Smith, I.; Wright, A.; Tyrrell, R.; Holland, K.; Martin, J.; Bald, J.; Yale, S.; Miles, T.; et al. Preliminary observations concerning treatment of visual discomfort and associated perceptual distortion. Ophthalmic Physiol. Opt. 1992, 12, 257–263. [Google Scholar] [CrossRef] [PubMed]
- Wilkins, A.; Sihra, N. A colorizer for use in determing an optimal ophthalmic tint. Color Res. Appl. 2000, 26, 246–253. [Google Scholar] [CrossRef]
- Hunt, R. Measuring Color; Fountain Press Ltd.: London, UK, 2001. [Google Scholar]
- Aldrich, A.; Lovell-Patel, R.; Allen, P.; Wilkins, A. The Repeatability of Colorimetry is Precise(ly) as Expected. Neuro Ophthalmol. Vis. Neurosci. 2018, 3, 1–6. [Google Scholar] [CrossRef]
- Wilkins, A.J.; Patel, R.; Adjamian, P.; Evans, B.J. Tinted spectacles and visually sensitive migraine. Cephalalgia 2002, 22, 711–719. [Google Scholar] [CrossRef]
- Wilkins, A.J.; Evans, B.J.; Brown, J.A.; Busby, A.E.; Wingfield, A.E.; Jeanes, R.J.; Bald, J. Double-masked placebo-controlled trial of precision spectral filters in children who use coloured overlays. Ophthalmic Physiol. Opt. 1994, 14, 365–370. [Google Scholar] [CrossRef]
- Wilkins, A.J.; Sihra, N.; Myers, A. Increasing reading speed by using colours: Issues concerning reliability and specificity, and their theoretical and practical implications. Perception 2005, 34, 109–120. [Google Scholar] [CrossRef]
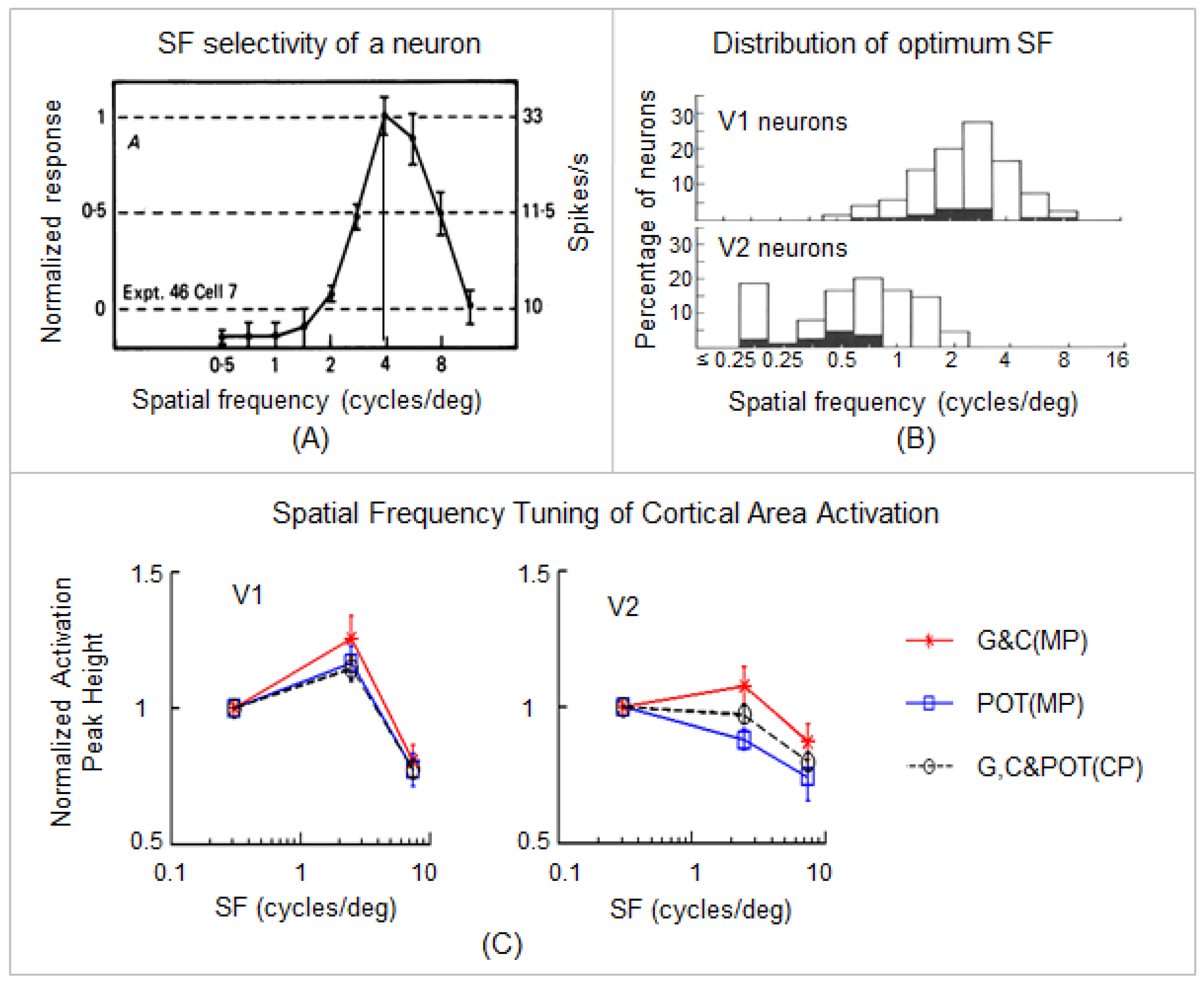
- Foster, K.H.; Gaska, J.P.; Nagler, M.; Pollen, D.A. Spatial and temporal frequency selectivity of neurones in visual cortical areas V1 and V2 of the macaque monkey. J. Physiol. 1985, 365, 331–363. [Google Scholar] [CrossRef]
- Cox, R. AFNI: Software for Analysis and Visualization of Functional Magnetic Resonance Neuroimages. Comput. Biomed. Res. 1996, 29, 162–173. [Google Scholar] [CrossRef]
- Huang, J.; Zhu, D.C. Visually stressful striped patterns alter human visual cortical functional connectivity. Hum. Brain Mapp. 2017, 38, 5474–5484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friston, K.J.; Holmes, A.P.; Worsley, K.J.; Poline, J.-P.; Frith, C.D.; Frackowiak, R.S.J. Statistical parametric maps in functional imaging: A general linear approach. Hum. Brain Mapp. 1995, 2, 189–210. [Google Scholar] [CrossRef]
- Worsley, K.J. Statistical analysis of activation images. In Functional MRI: An Introduction to Methods; Jezzard, P., Matthews, P.M., Smith, S.M., Eds.; Oxford University Press: New York, NY, USA, 2001; pp. 251–270. [Google Scholar]
- Khalil, N.M.; Nicotra, A.; Wilkins, A.J. Asymmetry of visual function in migraine with aura: Correlation with lateralisation of headache and aura. Cephalalgia 2010, 31, 213–221. [Google Scholar] [CrossRef]
- Wilkins, A.J.; Haigh, S.M.; Mahroo, O.A.; Plant, G.T. Photophobia in migraine: A symptom cluster? Cephalalgia 2021, 41, 1240–1248. [Google Scholar] [CrossRef] [PubMed]
- Wilkins, A.J.; Binnie, C.D.; Darby, C.E. Interhemispheric differences in photosensitive epilepsy. I. Pattern sensitivity thresholds. Electroencephalogr. Clin. Neurophysiol. 1981, 52, 461–468. [Google Scholar] [CrossRef]
- Binnie, C.D.; Wilkins, A.J.; De Korte, R.A. Interhemispheric differences in photosensitive epilepsy. II. Intermittent photic stimulation. Electroencephalogr. Clin. Neurophysiol. 1981, 52, 469–472. [Google Scholar] [CrossRef]
- Silvestro, M.; Tessitore, A.; Di Nardo, F.; di Clemente, F.S.; Trojsi, F.; Cirillo, M.; Esposito, F.; Tedeschi, G.; Russo, A. Functional connectivity changes in complex migraine aura: Beyond the visual network. Eur. J. Neurol. 2021, 1–10. [Google Scholar] [CrossRef]
- Friston, K.J. Functional and Effective Connectivity: A Review. Brain Connect. 2011, 1, 13–36. [Google Scholar] [CrossRef]
- Ning, Y.; Zheng, R.; Li, K.; Zhang, Y.; Lyu, D.; Jia, H.; Ren, Y.; Zou, Y. The altered Granger causality connection among pain-related brain networks in migraine. Medicine 2018, 97, e0102. [Google Scholar] [CrossRef] [PubMed]
- De Tommaso, M.; Trotta, G.; Vecchio, E.; Ricci, K.; Siugzdaite, R.; Stramaglia, S. Brain networking analysis in migraine with and without aura. J. Headache Pain 2017, 18, 98. [Google Scholar] [CrossRef] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, J.; Wilkins, A. The Functional Network of the Visual Cortex Is Altered in Migraine. Vision 2021, 5, 57. https://doi.org/10.3390/vision5040057
Huang J, Wilkins A. The Functional Network of the Visual Cortex Is Altered in Migraine. Vision. 2021; 5(4):57. https://doi.org/10.3390/vision5040057
Chicago/Turabian StyleHuang, Jie, and Arnold Wilkins. 2021. "The Functional Network of the Visual Cortex Is Altered in Migraine" Vision 5, no. 4: 57. https://doi.org/10.3390/vision5040057
APA StyleHuang, J., & Wilkins, A. (2021). The Functional Network of the Visual Cortex Is Altered in Migraine. Vision, 5(4), 57. https://doi.org/10.3390/vision5040057