Potential of Ocular Transmission of SARS-CoV-2: A Review

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Methods
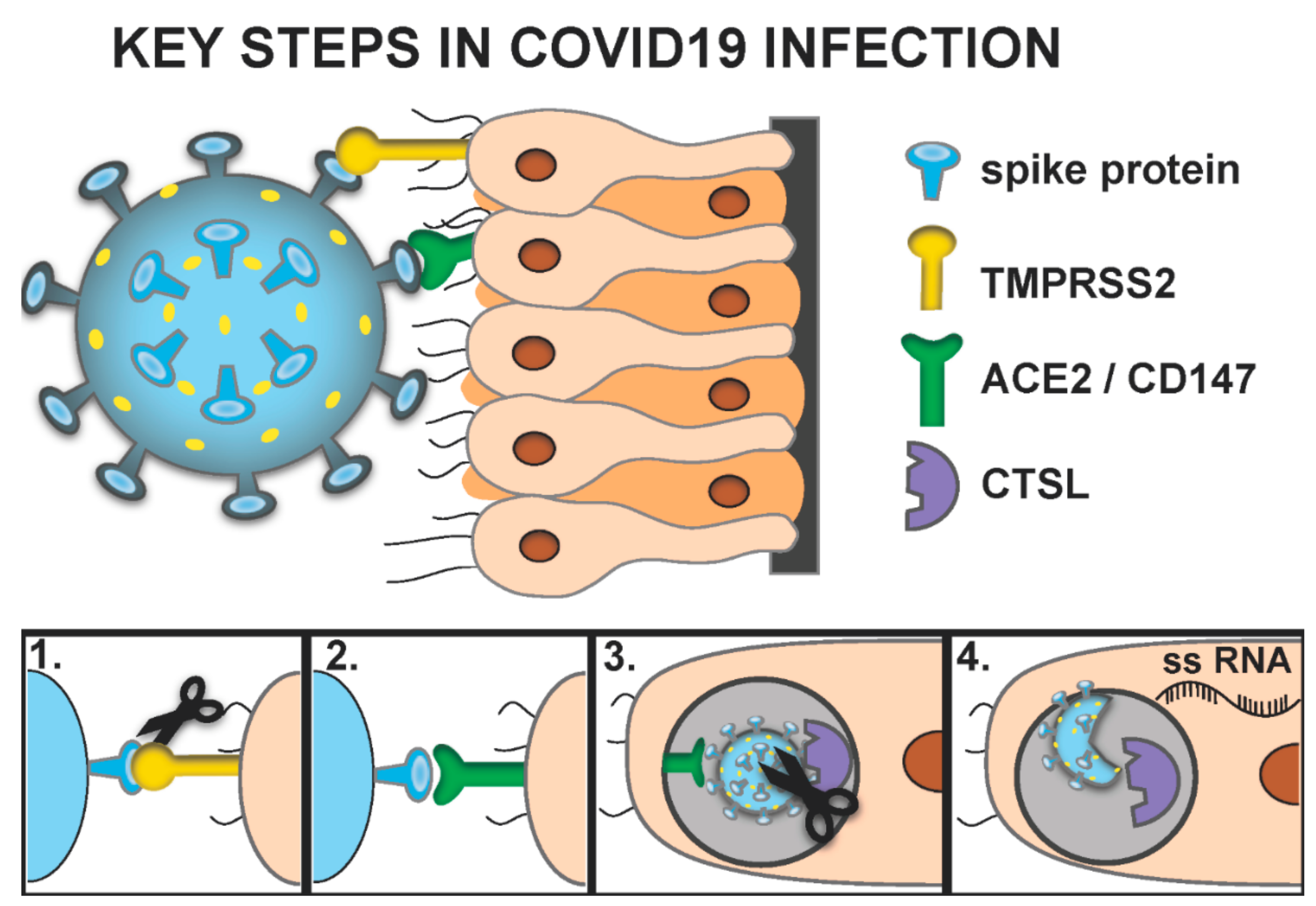
3. What Are the Molecular Mechanisms by Which the SARS-Cov-2 Virus Infect Cells?
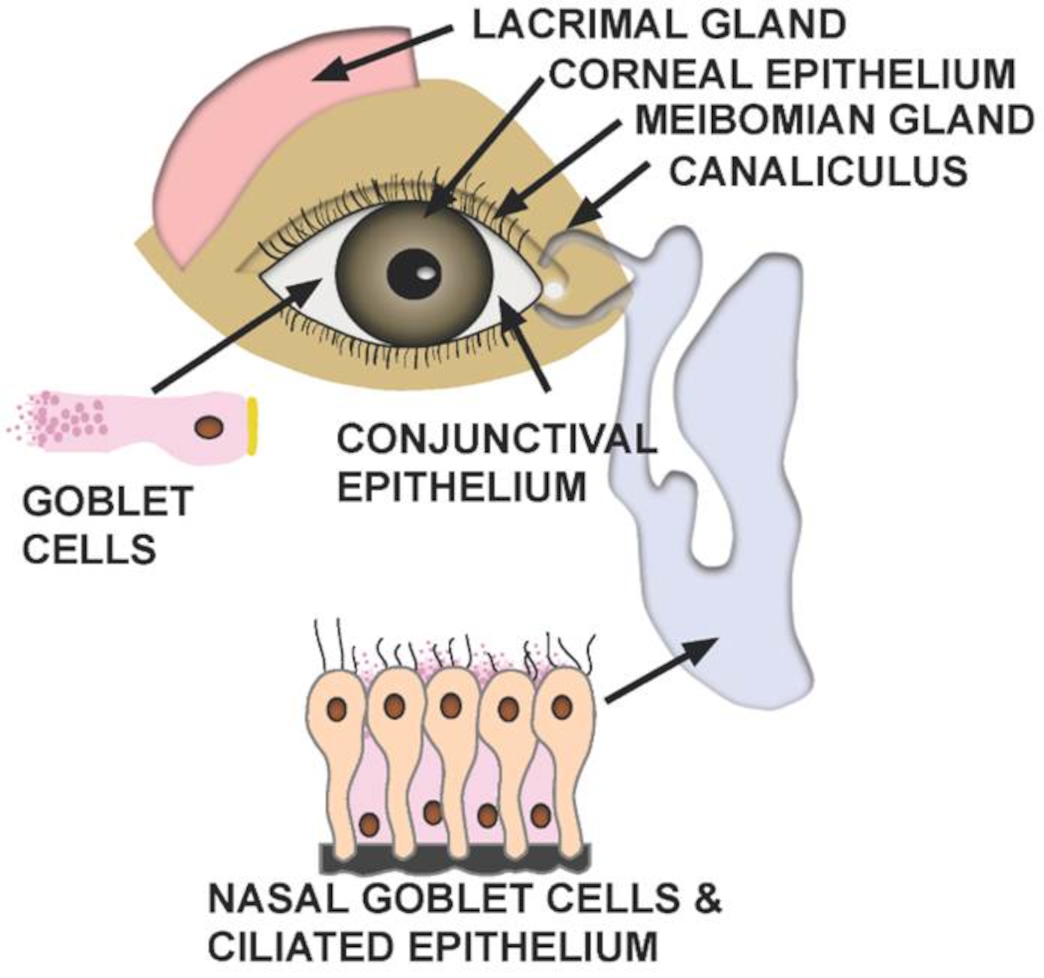
4. Can the Eye Serve as a Gateway for SARS-CoV-2 Transmission?
5. Discussion
6. Conclusions
- −
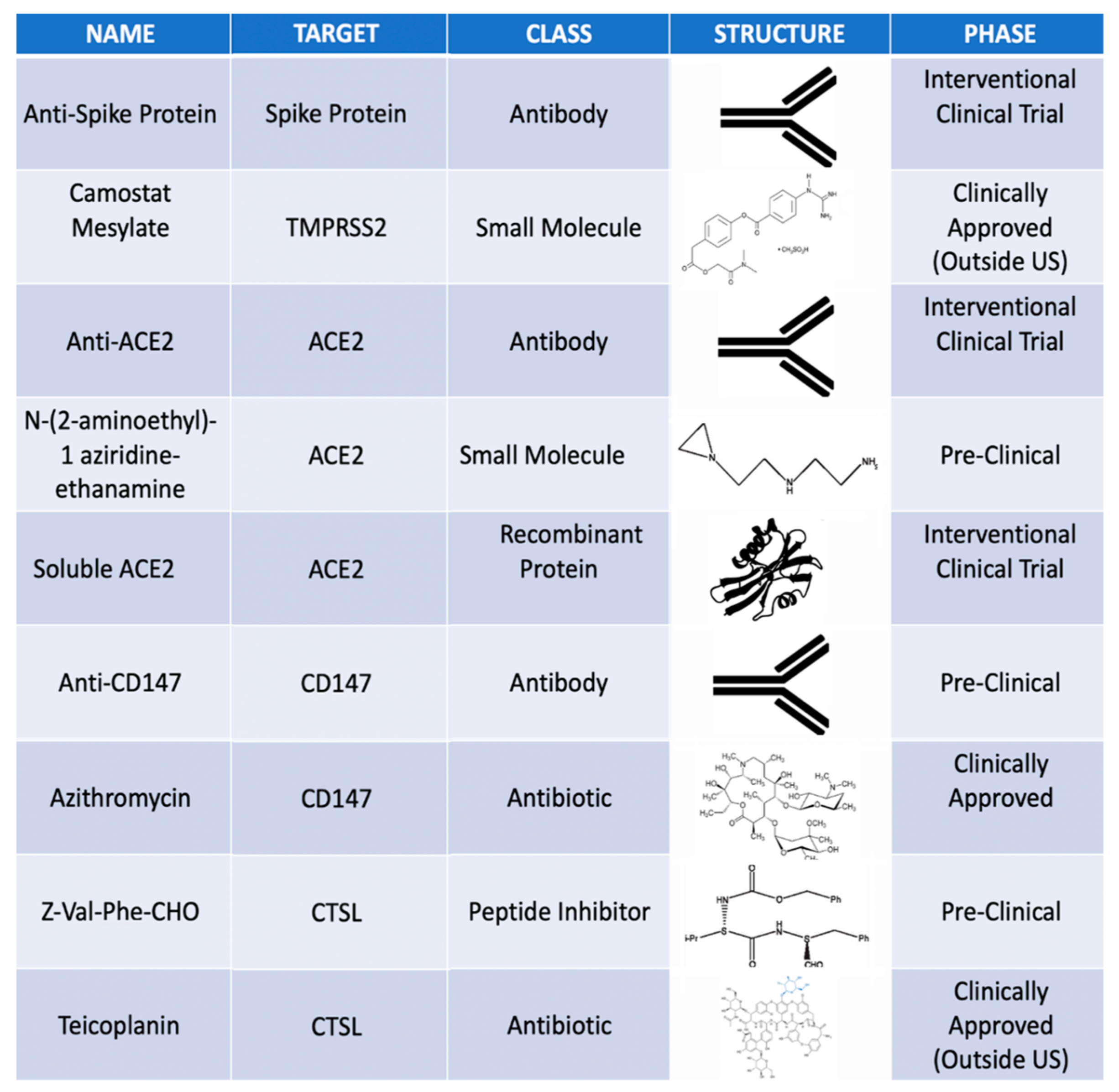
- The key proteins necessary for SARS-CoV-2 infection include Transmembrane Serine Protease 2 (TMPRSS2), CD147, Angiotensin-Converting Enzyme 2 (ACE2) and Cathepsin L (CTSL).
- −
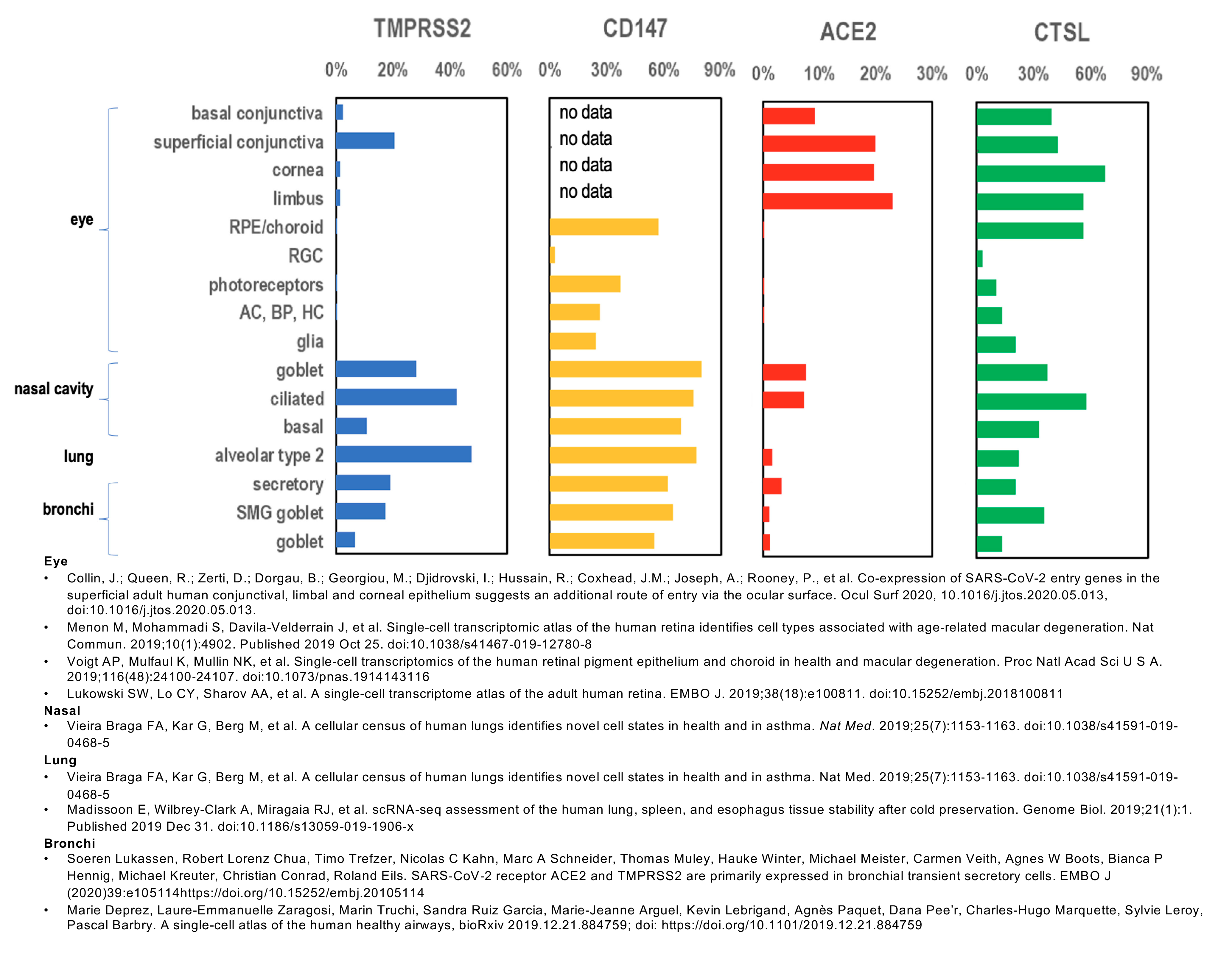
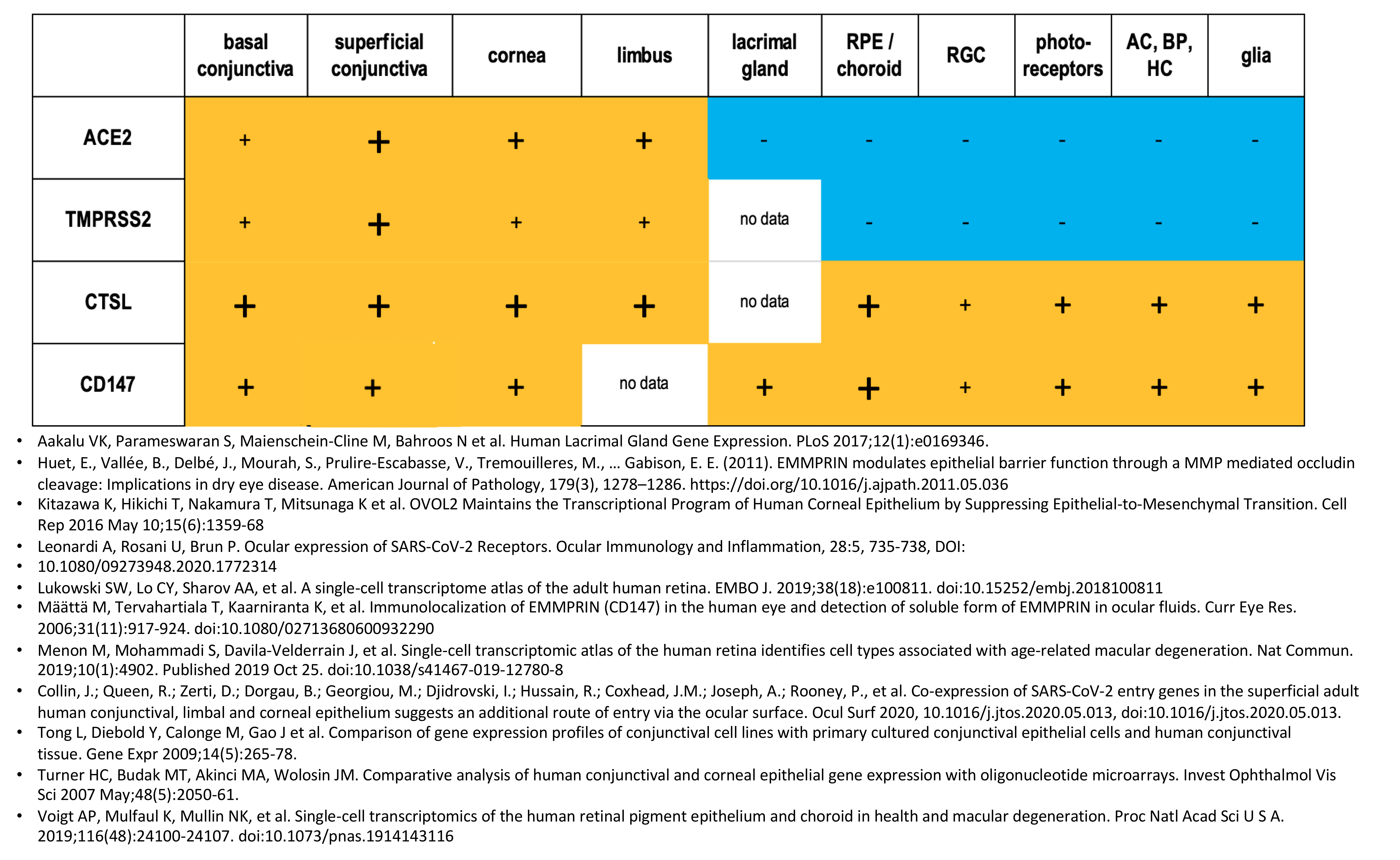
- The ocular surface and retina have variable expression of TMPRSS2, CD147, ACE2 and CTSL and appear to be susceptible to SARS-CoV-2 infection.
- −
- Nasal epithelium is known to highly express TMPRSS2, CD147, ACE2 and CTSL.
- −
- In addition to direct ocular infection, virus carried by tears through the nasolacrimal duct to nasal epithelium may represent a means of ocular inoculation.
- −
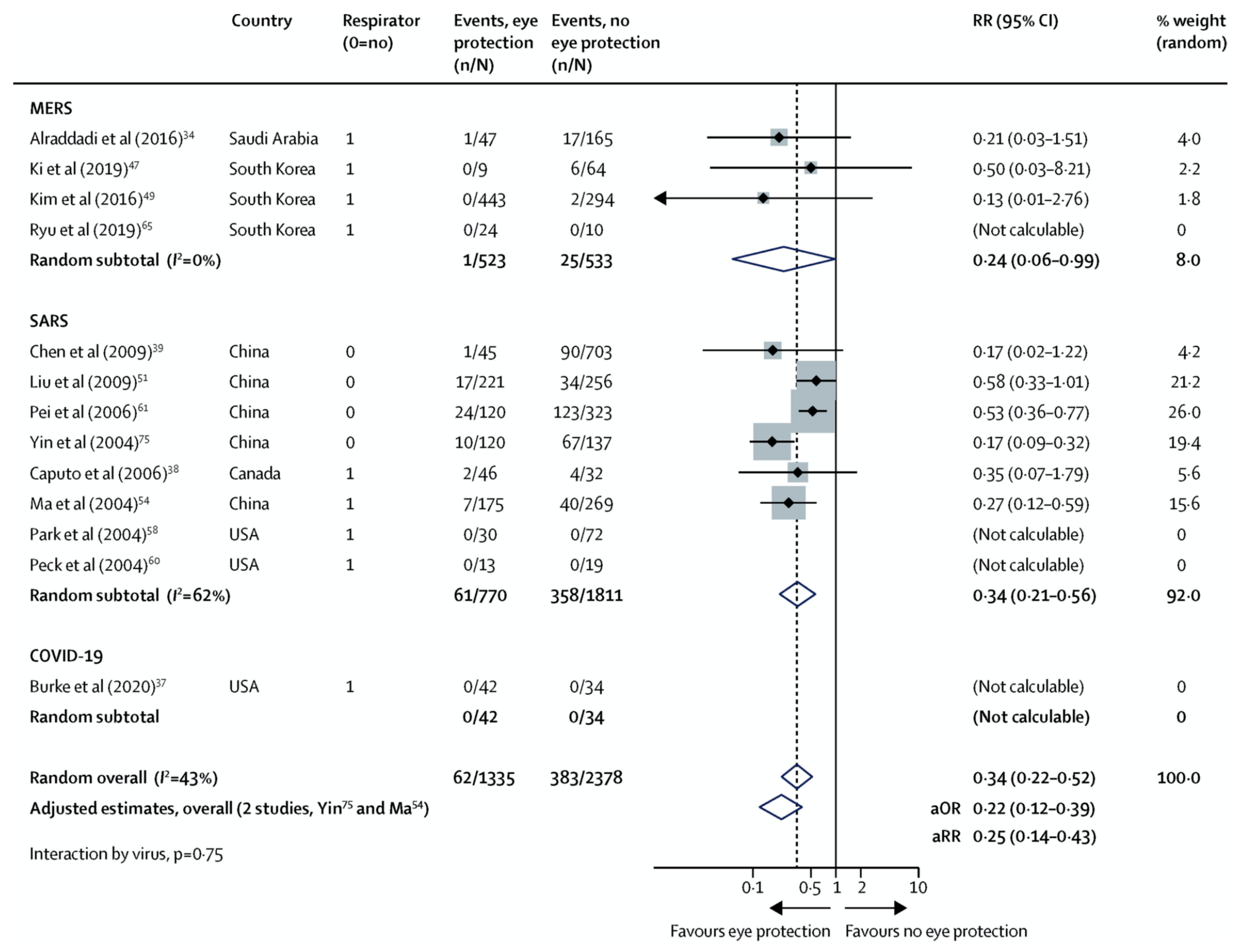
- Efforts to shield the ocular surface and prevent drainage of virus from tears into nasal epithelium may prevent SARS-CoV-2 infection.
Supplementary Materials
Funding
Conflicts of interest
References
- Peiris, J.S.; Yuen, K.Y.; Osterhaus, A.D.; Stohr, K. The severe acute respiratory syndrome. N. Engl. J. Med. 2003, 349, 2431–2441. [Google Scholar] [CrossRef] [PubMed]
- Raboud, J.; Shigayeva, A.; McGeer, A.; Bontovics, E.; Chapman, M.; Gravel, D.; Henry, B.; Lapinsky, S.; Loeb, M.; McDonald, L.C.; et al. Risk factors for SARS transmission from patients requiring intubation: A multicentre investigation in Toronto, Canada. PLoS ONE 2010, 5, e10717. [Google Scholar] [CrossRef]
- Sungnak, W.; Huang, N.; Becavin, C.; Berg, M.; Queen, R.; Litvinukova, M.; Talavera-Lopez, C.; Maatz, H.; Reichart, D.; Sampaziotis, F.; et al. SARS-CoV-2 entry factors are highly expressed in nasal epithelial cells together with innate immune genes. Nat. Med. 2020, 26, 681–687. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Chun, Y.S.; Lee, S.H.; Mun, S.K.; Jung, H.S.; Lee, S.H.; Son, Y.; Kim, J.C. Ocular surface reconstruction with autologous nasal mucosa in cicatricial ocular surface disease. Am. J. Ophthalmol. 2010, 149, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Leonardi, A.; Rosani, U.; Brun, P. Ocular Surface Expression of SARS-CoV-2 Receptors. Ocul. Immunol. Inflamm. 2020, 28, 735–738. [Google Scholar] [CrossRef] [PubMed]
- Ulrich, H.; Pillat, M.M. CD147 as a Target for COVID-19 Treatment: Suggested Effects of Azithromycin and Stem Cell Engagement. Stem Cell Rev. Rep. 2020. [Google Scholar] [CrossRef]
- Ou, X.; Liu, Y.; Lei, X.; Li, P.; Mi, D.; Ren, L.; Guo, L.; Guo, R.; Chen, T.; Hu, J.; et al. Characterization of spike glycoprotein of SARS-CoV-2 on virus entry and its immune cross-reactivity with SARS-CoV. Nat. Commun. 2020, 11, 1620. [Google Scholar] [CrossRef]
- Yan, R.; Zhang, Y.; Li, Y.; Xia, L.; Guo, Y.; Zhou, Q. Structural basis for the recognition of SARS-CoV-2 by full-length human ACE2. Science 2020, 367, 1444–1448. [Google Scholar] [CrossRef]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Kruger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.H.; Nitsche, A.; et al. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 2020, 181, 271–280.e278. [Google Scholar] [CrossRef]
- Simmons, G.; Gosalia, D.N.; Rennekamp, A.J.; Reeves, J.D.; Diamond, S.L.; Bates, P. Inhibitors of cathepsin L prevent severe acute respiratory syndrome coronavirus entry. Proc. Natl. Acad. Sci. USA 2005, 102, 11876–11881. [Google Scholar] [CrossRef]
- Zhou, L.; Xu, Z.; Castiglione, G.M.; Soiberman, U.S.; Eberhart, C.G.; Duh, E.J. ACE2 and TMPRSS2 are expressed on the human ocular surface, suggesting susceptibility to SARS-CoV-2 infection. Biorxiv 2020. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.N.; Wang, Q.; Liu, T.; Dou, S.Q.; Qi, X.; Jiang, H.; Qi, B.X.; Zhang, B.; Zhou, Q.J. [Expression analysis of 2019-nCoV related ACE2 and TMPRSS2 in eye tissues]. Zhonghua Yan Ke Za Zhi 2020, 56, E011. [Google Scholar] [CrossRef]
- Heurich, A.; Hofmann-Winkler, H.; Gierer, S.; Liepold, T.; Jahn, O.; Pohlmann, S. TMPRSS2 and ADAM17 cleave ACE2 differentially and only proteolysis by TMPRSS2 augments entry driven by the severe acute respiratory syndrome coronavirus spike protein. J. Virol. 2014, 88, 1293–1307. [Google Scholar] [CrossRef] [PubMed]
- Asselta, R.; Paraboschi, E.M.; Mantovani, A.; Duga, S. ACE2 and TMPRSS2 variants and expression as candidates to sex and country differences in COVID-19 severity in Italy. Medrxiv 2020. [Google Scholar] [CrossRef]
- Redzic, J.S.; Armstrong, G.S.; Isern, N.G.; Jones, D.N.; Kieft, J.S.; Eisenmesser, E.Z. The retinal specific CD147 Ig0 domain: From molecular structure to biological activity. J. Mol. Biol. 2011, 411, 68–82. [Google Scholar] [CrossRef]
- Huet, E.; Vallee, B.; Delbe, J.; Mourah, S.; Pruliere-Escabasse, V.; Tremouilleres, M.; Kadomatsu, K.; Doan, S.; Baudouin, C.; Menashi, S.; et al. EMMPRIN modulates epithelial barrier function through a MMP-mediated occludin cleavage: Implications in dry eye disease. Am. J. Pathol. 2011, 179, 1278–1286. [Google Scholar] [CrossRef]
- Joung, C.; Noh, H.; Jung, J.; Song, H.Y.; Bae, H.; Pahk, K.; Kim, W.K. A Novel CD147 Inhibitor, SP-8356, Attenuates Pathological Fibrosis in Alkali-Burned Rat Cornea. Int. J. Mol. Sci. 2020, 21, 2990. [Google Scholar] [CrossRef]
- Mission, W.-C.J. Report of the WHO-China Joint Mission on Coronavirus Disease 2019 (COVID-19); World Health Organization: Geneva, Switzerland, 2020. [Google Scholar]
- Marinho, P.M.; Marcos, A.A.A.; Romano, A.C.; Nascimento, H.; Belfort, R., Jr. Retinal findings in patients with COVID-19. Lancet 2020, 395, 1610. [Google Scholar] [CrossRef]
- Aakalu, V.K.; Parameswaran, S.; Maienschein-Cline, M.; Bahroos, N.; Shah, D.; Ali, M.; Krishnakumar, S. Human Lacrimal Gland Gene Expression. PLoS ONE 2017, 12, e0169346. [Google Scholar] [CrossRef]
- Kitazawa, K.; Hikichi, T.; Nakamura, T.; Mitsunaga, K.; Tanaka, A.; Nakamura, M.; Yamakawa, T.; Furukawa, S.; Takasaka, M.; Goshima, N.; et al. OVOL2 Maintains the Transcriptional Program of Human Corneal Epithelium by Suppressing Epithelial-to-Mesenchymal Transition. Cell Rep. 2016, 15, 1359–1368. [Google Scholar] [CrossRef]
- Lukowski, S.W.; Lo, C.Y.; Sharov, A.A.; Nguyen, Q.; Fang, L.; Hung, S.S.; Zhu, L.; Zhang, T.; Grunert, U.; Nguyen, T.; et al. A single-cell transcriptome atlas of the adult human retina. EMBO J. 2019, 38, e100811. [Google Scholar] [CrossRef] [PubMed]
- Maatta, M.; Tervahartiala, T.; Kaarniranta, K.; Tang, Y.; Yan, L.; Tuukkanen, J.; Sorsa, T. Immunolocalization of EMMPRIN (CD147) in the human eye and detection of soluble form of EMMPRIN in ocular fluids. Curr. Eye Res. 2006, 31, 917–924. [Google Scholar] [CrossRef] [PubMed]
- Menon, M.; Mohammadi, S.; Davila-Velderrain, J.; Goods, B.A.; Cadwell, T.D.; Xing, Y.; Stemmer-Rachamimov, A.; Shalek, A.K.; Love, J.C.; Kellis, M.; et al. Single-cell transcriptomic atlas of the human retina identifies cell types associated with age-related macular degeneration. Nat. Commun. 2019, 10, 4902. [Google Scholar] [CrossRef] [PubMed]
- Tong, L.; Diebold, Y.; Calonge, M.; Gao, J.; Stern, M.E.; Beuerman, R.W. Comparison of gene expression profiles of conjunctival cell lines with primary cultured conjunctival epithelial cells and human conjunctival tissue. Gene Expr. 2009, 14, 265–278. [Google Scholar] [CrossRef]
- Turner, H.C.; Budak, M.T.; Akinci, M.A.; Wolosin, J.M. Comparative analysis of human conjunctival and corneal epithelial gene expression with oligonucleotide microarrays. Invest. Ophthalmol. Vis. Sci. 2007, 48, 2050–2061. [Google Scholar] [CrossRef]
- Voigt, A.P.; Mulfaul, K.; Mullin, N.K.; Flamme-Wiese, M.J.; Giacalone, J.C.; Stone, E.M.; Tucker, B.A.; Scheetz, T.E.; Mullins, R.F. Single-cell transcriptomics of the human retinal pigment epithelium and choroid in health and macular degeneration. Proc. Natl. Acad. Sci. USA 2019, 116, 24100–24107. [Google Scholar] [CrossRef] [PubMed]
- Sugita, S.; Horie, S.; Nakamura, O.; Maruyama, K.; Takase, H.; Usui, Y.; Takeuchi, M.; Ishidoh, K.; Koike, M.; Uchiyama, Y.; et al. Acquisition of T regulatory function in cathepsin L-inhibited T cells by eye-derived CTLA-2alpha during inflammatory conditions. J. Immunol. 2009, 183, 5013–5022. [Google Scholar] [CrossRef]
- Robbins, S.G.; Detrick, B.; Hooks, J.J. Ocular tropisms of murine coronavirus (strain JHM) after inoculation by various routes. Invest. Ophthalmol. Vis. Sci. 1991, 32, 1883–1893. [Google Scholar]
- Nagata, N.; Iwata-Yoshikawa, N.; Taguchi, F. Studies of severe acute respiratory syndrome coronavirus pathology in human cases and animal models. Vet. Pathol. 2010, 47, 881–892. [Google Scholar] [CrossRef]
- Martina, B.E.; Haagmans, B.L.; Kuiken, T.; Fouchier, R.A.; Rimmelzwaan, G.F.; Van Amerongen, G.; Peiris, J.S.; Lim, W.; Osterhaus, A.D. Virology: SARS virus infection of cats and ferrets. Nature 2003, 425, 915. [Google Scholar] [CrossRef]
- Andries, K.; Pensaert, M.B. Immunofluorescence studies on the pathogenesis of hemagglutinating encephalomyelitis virus infection in pigs after oronasal inoculation. Am. J. Vet. Res. 1980, 41, 1372–1378. [Google Scholar] [PubMed]
- Doherty, M.J. Ocular manifestations of feline infectious peritonitis. J. Am. Vet. Med. Assoc. 1971, 159, 417–424. [Google Scholar] [PubMed]
- Lai, Y.L.; Jacoby, R.O.; Bhatt, P.N.; Jonas, A.M. Keratoconjunctivitis associated with sialodacryoadenitis in rats. Investig. Ophthalmol. 1976, 15, 538–541. [Google Scholar]
- Guan, W.J.; Ni, Z.Y.; Hu, Y.; Liang, W.H.; Ou, C.Q.; He, J.X.; Liu, L.; Shan, H.; Lei, C.L.; Hui, D.S.C.; et al. Clinical Characteristics of Coronavirus Disease 2019 in China. N. Engl. J. Med. 2020, 382, 1708–1720. [Google Scholar] [CrossRef]
- Lu, C.-W.; Liu, X.-F.; Jia, Z.-F. 2019-nCoV transmission through the ocular surface must not be ignored. Lancet 2020, 395, e39. [Google Scholar] [CrossRef]
- Vieira Braga, F.A.; Kar, G.; Berg, M.; Carpaij, O.A.; Polanski, K.; Simon, L.M.; Brouwer, S.; Gomes, T.; Hesse, L.; Jiang, J.; et al. A cellular census of human lungs identifies novel cell states in health and in asthma. Nat. Med. 2019, 25, 1153–1163. [Google Scholar] [CrossRef]
- Madissoon, E.; Wilbrey-Clark, A.; Miragaia, R.J.; Saeb-Parsy, K.; Mahbubani, K.T.; Georgakopoulos, N.; Harding, P.; Polanski, K.; Huang, N.; Nowicki-Osuch, K.; et al. scRNA-seq assessment of the human lung, spleen, and esophagus tissue stability after cold preservation. Genome Biol. 2019, 21, 1. [Google Scholar] [CrossRef]
- Lukassen, S.; Chua, R.L.; Trefzer, T.; Kahn, N.C.; Schneider, M.A.; Muley, T.; Winter, H.; Meister, M.; Veith, C.; Boots, A.W.; et al. SARS-CoV-2 receptor ACE2 and TMPRSS2 are primarily expressed in bronchial transient secretory cells. EMBO J. 2020, 39, e105114. [Google Scholar] [CrossRef]
- Deprez, M.; Zaragosi, L.-E.; Truchi, M.; Garcia, S.R.; Arguel, M.-J.; Lebrigand, K.; Paquet, A.; Pee’r, D.; Marquette, C.-H.; Leroy, S.; et al. A single-cell atlas of the human healthy airways. Biorxiv 2019. [Google Scholar] [CrossRef]
- Alraddadi, B.M.; Al-Salmi, H.S.; Jacobs-Slifka, K.; Slayton, R.B.; Estivariz, C.F.; Geller, A.I.; Al-Turkistani, H.H.; Al-Rehily, S.S.; Alserehi, H.A.; Wali, G.Y.; et al. Risk Factors for Middle East Respiratory Syndrome Coronavirus Infection among Healthcare Personnel. Emerg. Infect. Dis. 2016, 22, 1915–1920. [Google Scholar] [CrossRef]
- Kim, C.J.; Choi, W.S.; Jung, Y.; Kiem, S.; Seol, H.Y.; Woo, H.J.; Choi, Y.H.; Son, J.S.; Kim, K.H.; Kim, Y.S.; et al. Surveillance of the Middle East respiratory syndrome (MERS) coronavirus (CoV) infection in healthcare workers after contact with confirmed MERS patients: Incidence and risk factors of MERS-CoV seropositivity. Clin. Microbiol. Infect. 2016, 22, 880–886. [Google Scholar] [CrossRef] [PubMed]
- Ryu, B.; Cho, S.I.; Oh, M.D.; Lee, J.K.; Lee, J.; Hwang, Y.O.; Yang, J.S.; Kim, S.S.; Bang, J.H. Seroprevalence of Middle East respiratory syndrome coronavirus (MERS-CoV) in public health workers responding to a MERS outbreak in Seoul, Republic of Korea, in 2015. West. Pac. Surveill. Response J. 2019, 10, 46–48. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.Q.; Ling, W.H.; Lu, C.Y.; Hao, Y.T.; Lin, Z.N.; Ling, L.; Huang, J.; Li, G.; Yan, G.M. Which preventive measures might protect health care workers from SARS? BMC Public Health 2009, 9, 81. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.C.; Chang, W.H.; Jiang, B.C.; Maa, S.H.; Chien, D.K.; Tang, W.R. Mortality Factor Survey of Severe Acute Respiratory Syndrome in Taiwan. Int. J. Gerontol. 2009, 3, 31–38. [Google Scholar] [CrossRef][Green Version]
- Pei, L.Y.; Gao, Z.C.; Yang, Z.; Wei, D.G.; Wang, S.X.; Ji, J.M.; Jiang, B.G. Investigation of the influencing factors on severe acute respiratory syndrome among health care workers. Beijing Da Xue Xue Bao Yi Xue Ban 2006, 38, 271–275. [Google Scholar] [PubMed]
- Yin, W.W.; Gao, L.D.; Lin, W.S.; Gao, L.D.; Lin, W.S.; Du, L.; Zhang, X.C.; Zou, Q.; Li, L.H.; Liang, W.J.; et al. Effectiveness of personal protective measures in prevention of nosocomial transmission of severe acute respiratory syndrome. Zhonghua Liu Xing Bing Xue Za Zhi 2004, 25, 18–22. (In Chinese) [Google Scholar]
- Caputo, K.M.; Byrick, R.; Chapman, M.G.; Orser, B.J.; Orser, B.A. Intubation of SARS patients: Infection and perspectives of healthcare workers. Can. J. Anaesth. 2006, 53, 122–129. [Google Scholar] [CrossRef]
- Li, Q.; Guan, X.; Wu, P.; Wang, X.; Zhou, L.; Tong, Y.; Ren, R.; Leung, K.S.M.; Lau, E.H.Y.; Wong, J.Y.; et al. Early Transmission Dynamics in Wuhan, China, of Novel Coronavirus-Infected Pneumonia. N. Engl. J. Med. 2020, 382, 1199–1207. [Google Scholar] [CrossRef]
- Park, J.E.; Li, K.; Barlan, A.; Fehr, A.R.; Perlman, S.; McCray, P.B., Jr.; Gallagher, T. Proteolytic processing of Middle East respiratory syndrome coronavirus spikes expands virus tropism. Proc. Natl. Acad. Sci. USA 2016, 113, 12262–12267. [Google Scholar] [CrossRef]
- Peck, A.J.; Newbern, E.C.; Feikin, D.R.; Issakbaeva, E.T.; Park, B.J.; Fehr, J.; LaMonte, A.C.; Le, T.P.; Burger, T.L.; Rhodes, L.V., 3rd; et al. Lack of SARS transmission and U.S. SARS case-patient. Emerg. Infect. Dis. 2004, 10, 217–224. [Google Scholar] [CrossRef]
- Burke, R.M.; Balter, S.; Barnes, E.; Barry, V.; Bartlett, K.; Beer, K.D.; Benowitz, I.; Biggs, H.M.; Bruce, H.; Bryant-Genevier, J.; et al. Enhanced Contact Investigations for Nine Early Travel-Related Cases of SARS-CoV-2 in the United States. Medrxiv 2020. [Google Scholar] [CrossRef]
- Chu, D.K.; Akl, E.A.; Duda, S.; Solo, K.; Yaacoub, S.; Schünemann, H.J.; Chu, D.K.; Akl, E.A.; El-harakeh, A.; Bognanni, A.; et al. Physical distancing, face masks, and eye protection to prevent person-to-person transmission of SARS-CoV-2 and COVID-19: A systematic review and meta-analysis. Lancet 2020, 395, 1973–1987. [Google Scholar] [CrossRef]
- Bhaskar, M.E.; Arun, S. SARS-CoV-2 Infection Among Community Health Workers in India Before and After Use of Face Shields. J. Am. Med. Aossc. 2020. [Google Scholar] [CrossRef] [PubMed]
- Varga, Z.; Flammer, A.J.; Steiger, P.; Haberecker, M.; Andermatt, R.; Zinkernagel, A.S.; Mehra, M.R.; Schuepbach, R.A.; Ruschitzka, F.; Moch, H. Endothelial cell infection and endotheliitis in COVID-19. Lancet 2020, 395, 1417–1418. [Google Scholar] [CrossRef]
- Hui, K.P.Y.; Ching, R.H.H.; Chan, S.K.H.; Nicholls, J.M.; Sachs, N.; Clevers, H.; Peiris, J.S.M.; Chan, M.C.W. Tropism, replication competence, and innate immune responses of influenza virus: An analysis of human airway organoids and ex-vivo bronchus cultures. Lancet Respir Med. 2018, 6, 846–854. [Google Scholar] [CrossRef]
- Dai, H.; Zhang, X.; Xia, J.; Zhang, T.; Shang, Y.; Huang, R.; Liu, R.; Wang, D.; Li, M.; Wu, J.; et al. High-resolution Chest CT Features and Clinical Characteristics of Patients Infected with COVID-19 in Jiangsu, China. Int. J. Infect. Dis. 2020. [Google Scholar] [CrossRef]
- Dye, B.R.; Hill, D.R.; Ferguson, M.A.; Tsai, Y.H.; Nagy, M.S.; Dyal, R.; Wells, J.M.; Mayhew, C.N.; Nattiv, R.; Klein, O.D.; et al. In vitro generation of human pluripotent stem cell derived lung organoids. Elife 2015, 4, e05098. [Google Scholar] [CrossRef]
- Foster, J.W.; Wahlin, K.; Adams, S.M.; Birk, D.E.; Zack, D.J.; Chakravarti, S. Cornea organoids from human induced pluripotent stem cells. Sci. Rep. 2017, 7, 41286. [Google Scholar] [CrossRef]
- Monteil, V.; Kwon, H.; Prado, P.; Hagelkruys, A.; Wimmer, R.A.; Stahl, M.; Leopoldi, A.; Garreta, E.; Del Pozo, C.H.; Prosper, F.; et al. Inhibition of SARS-CoV-2 Infections in Engineered Human Tissues Using Clinical-Grade Soluble Human ACE2. Cell 2020, e7, 905–913.e7. [Google Scholar] [CrossRef]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barnett, B.P.; Wahlin, K.; Krawczyk, M.; Spencer, D.; Welsbie, D.; Afshari, N.; Chao, D. Potential of Ocular Transmission of SARS-CoV-2: A Review. Vision 2020, 4, 40. https://doi.org/10.3390/vision4030040
Barnett BP, Wahlin K, Krawczyk M, Spencer D, Welsbie D, Afshari N, Chao D. Potential of Ocular Transmission of SARS-CoV-2: A Review. Vision. 2020; 4(3):40. https://doi.org/10.3390/vision4030040
Chicago/Turabian StyleBarnett, Brad P., Karl Wahlin, Michal Krawczyk, Doran Spencer, Derek Welsbie, Natalie Afshari, and Daniel Chao. 2020. "Potential of Ocular Transmission of SARS-CoV-2: A Review" Vision 4, no. 3: 40. https://doi.org/10.3390/vision4030040
APA StyleBarnett, B. P., Wahlin, K., Krawczyk, M., Spencer, D., Welsbie, D., Afshari, N., & Chao, D. (2020). Potential of Ocular Transmission of SARS-CoV-2: A Review. Vision, 4(3), 40. https://doi.org/10.3390/vision4030040