


The Zootechnical Performance, Health State Modulation, Morphology, and Intestinal Microbiome of Nile Tilapia Juveniles Fed with a Functional Blend of Immunostimulants Associated with a Diet High in Soybean Meal
 , ,
, ,  ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Diets
2.2. Fish and Experimental Conditions
2.3. Sampling Procedure
2.4. Growth Performance and Survival
(initial body weight × initial body protein)]/protein intake
2.5. Proximate Analysis
2.6. Hematological Parameters
2.7. Immunological Parameters
2.8. Histological Analysis
2.9. Transmission Electron Microscopy (TEM)
2.10. Statistical Analysis
2.11. Metagenomic Analysis
3. Results
3.1. Growth Performance, Survival, and Proximate Composition
3.2. Hematological and Immunological Parameters
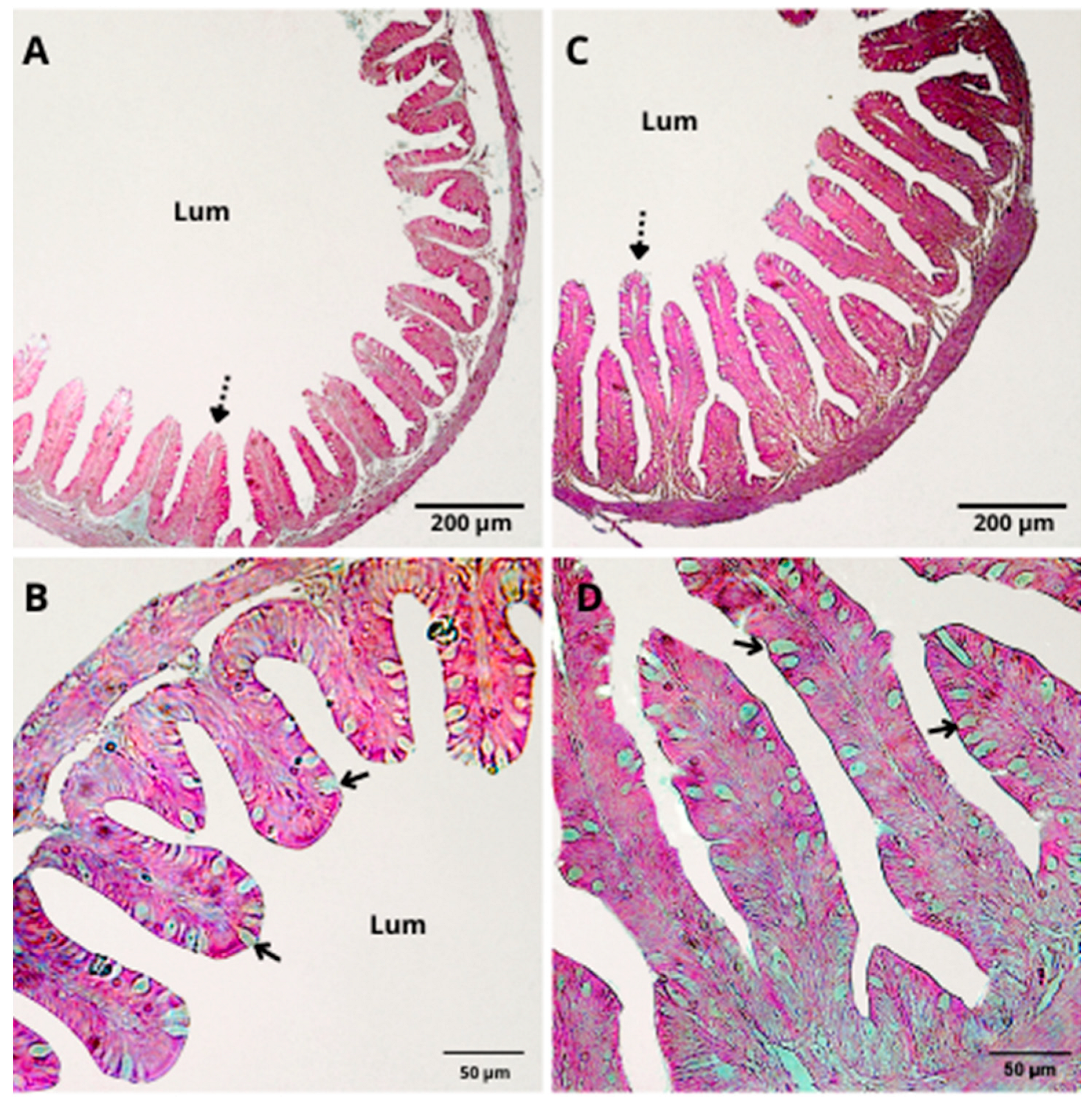
3.3. Histological Analysis
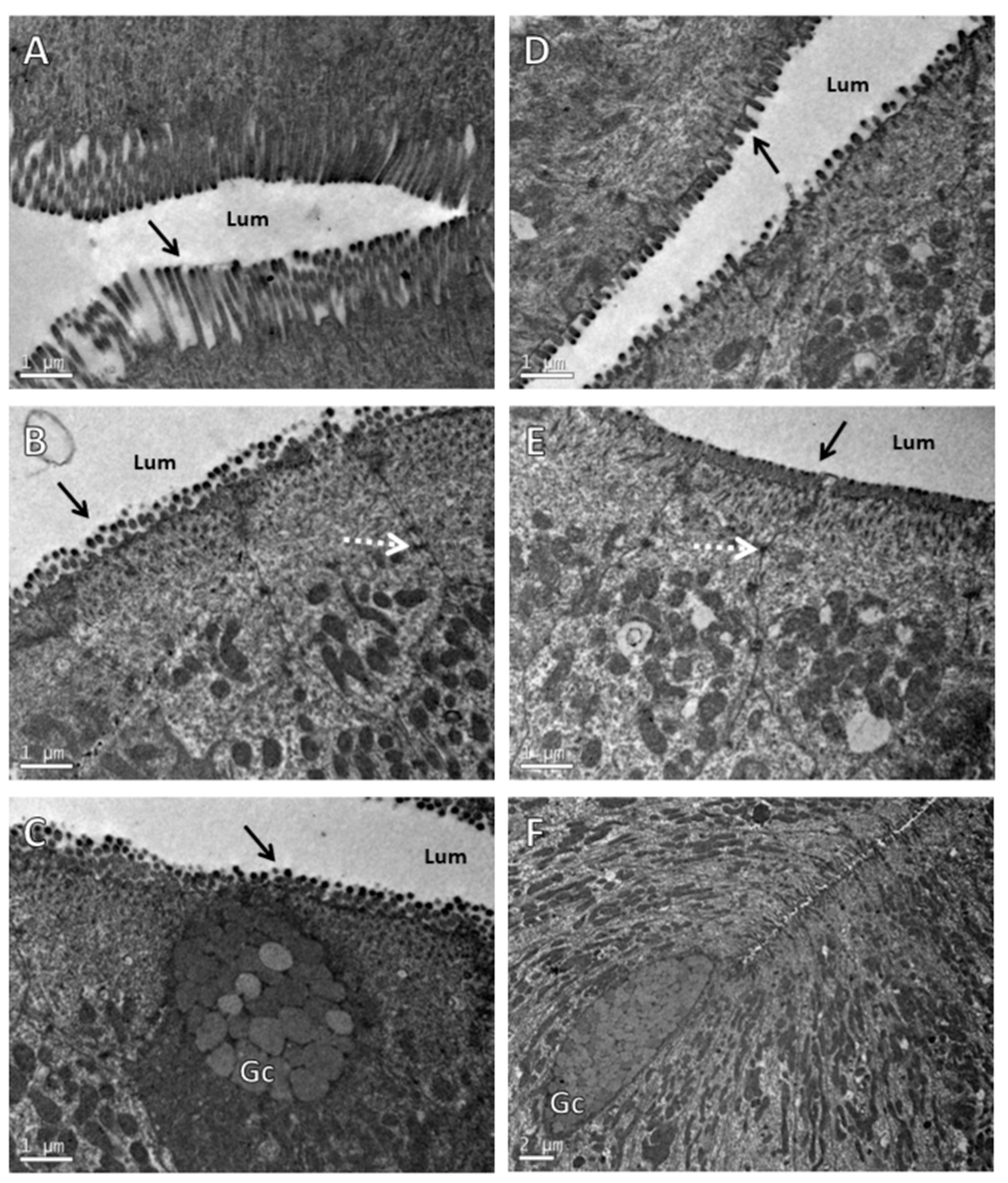
Transmission Electron Microscopy (TEM)
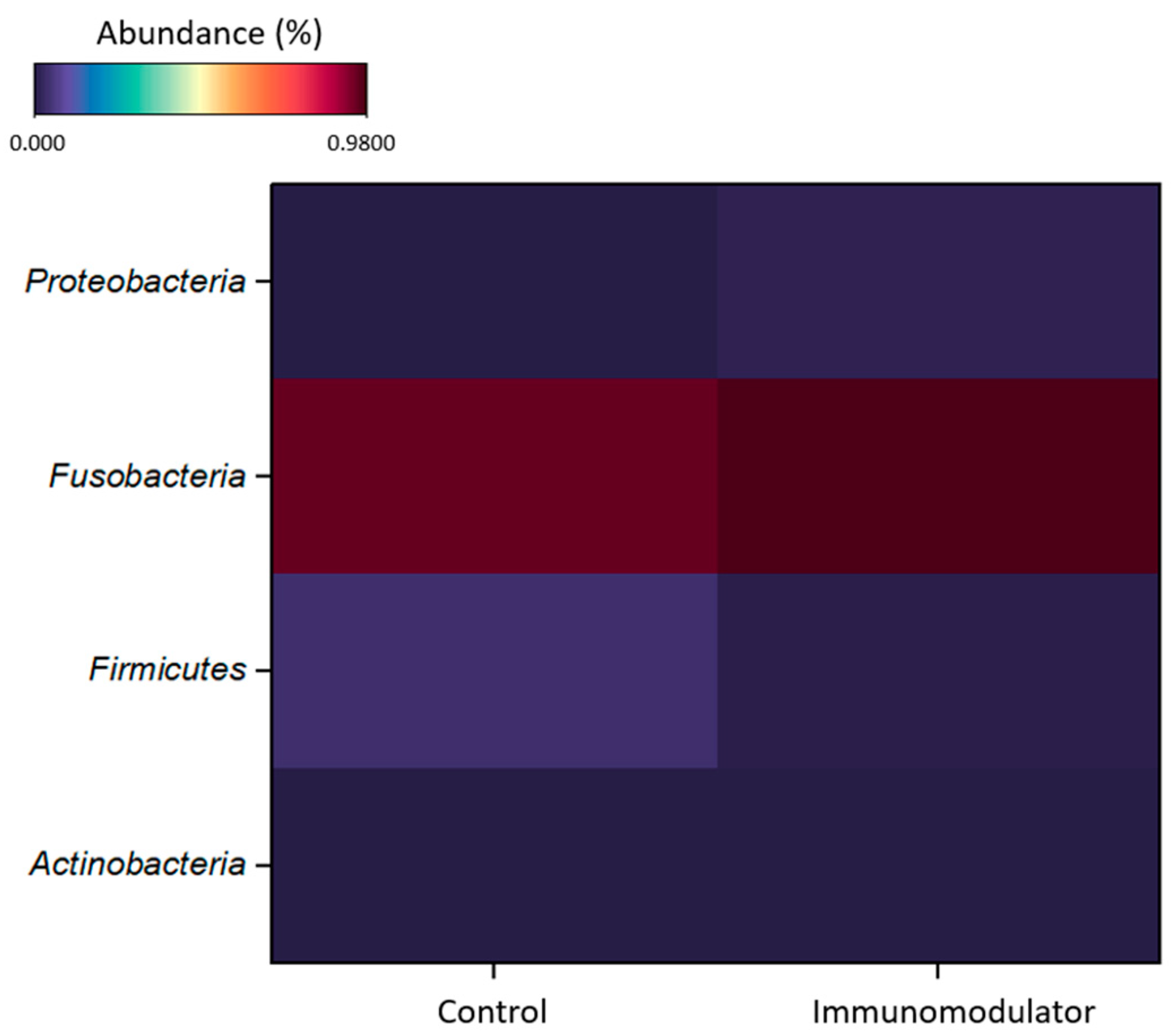
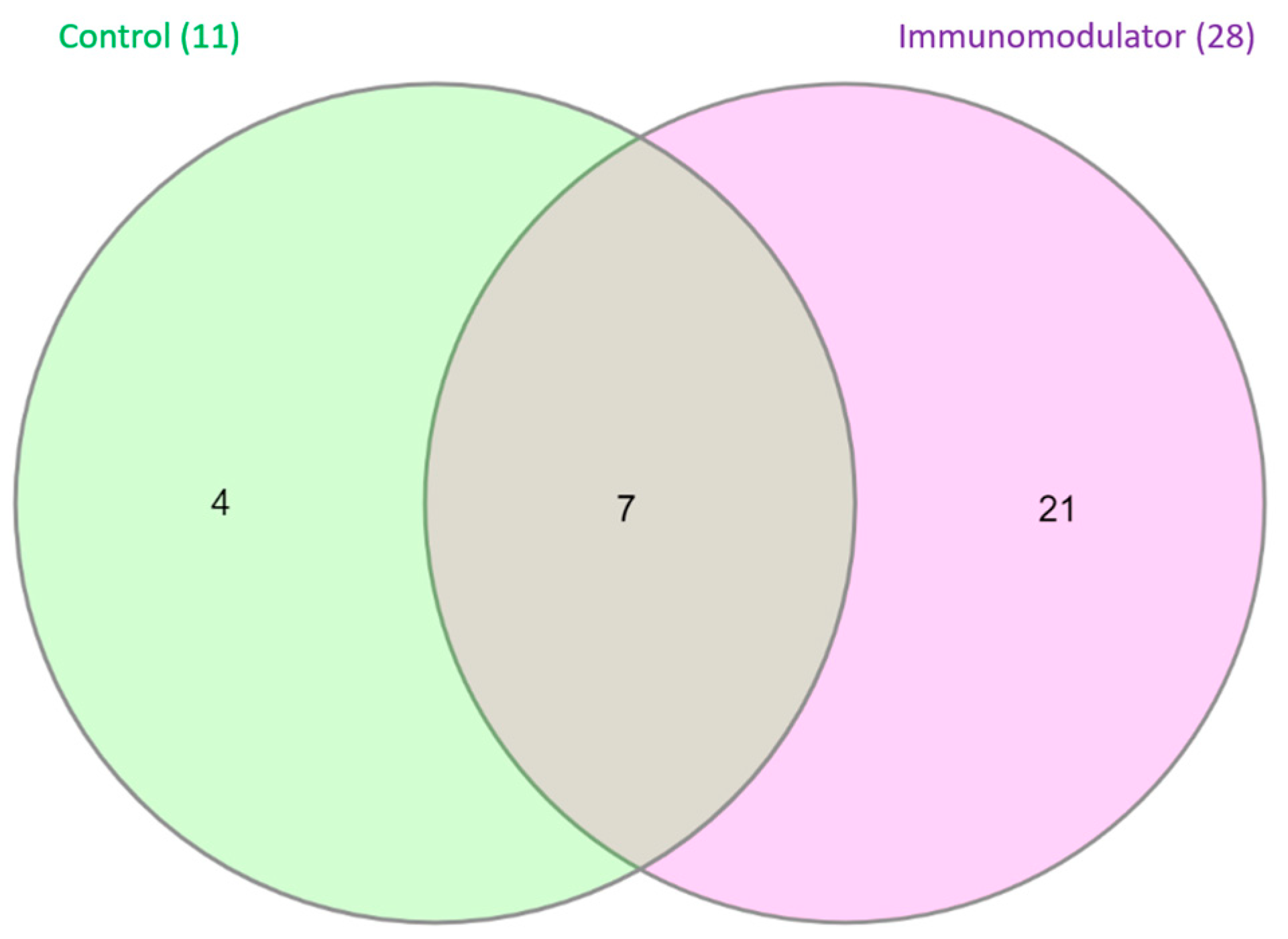
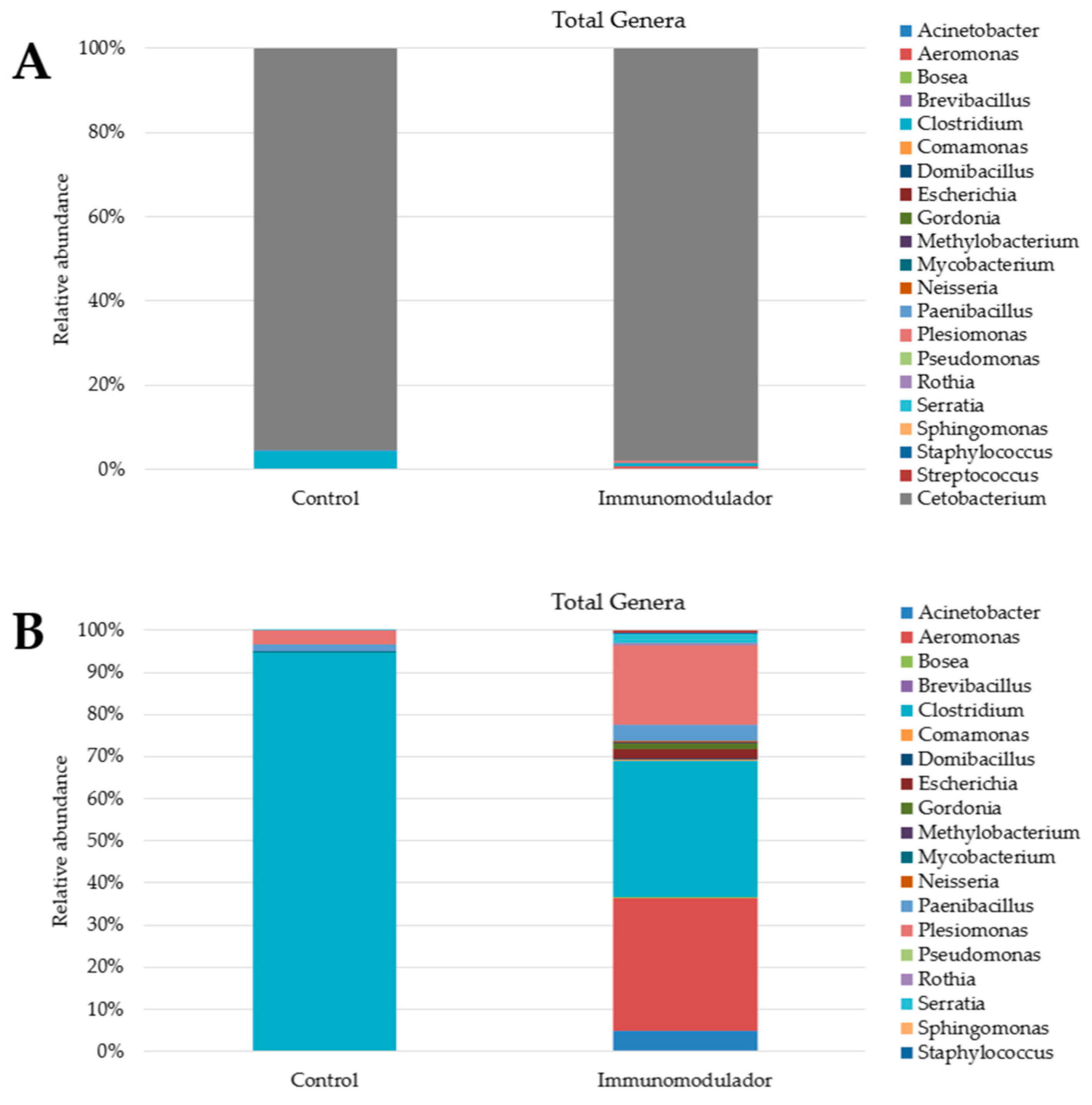
3.4. Metagenomic Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zuberi, A.; Kamran, M.; Younus, N.; Abdel-Tawwab, M. Editorial: Functional Feed Additives: Current Trends. Front. Aquac. 2024, 3, 1385508. [Google Scholar] [CrossRef]
- Dimitroglou, A.; Merrifield, D.L.; Carnevali, O.; Picchietti, S.; Avella, M.; Daniels, C.; Güroy, D.; Davies, S.J. Microbial Manipulations to Improve Fish Health and Production—A Mediterranean Perspective. Fish Shellfish Immunol. 2011, 30, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Aramli, M.S.; Kamangar, B.; Nazari, R.M. Effects of Dietary β-Glucan on the Growth and Innate Immune Response of Juvenile Persian Sturgeon, Acipenser persicus. Fish Shellfish Immunol. 2015, 47, 606–610. [Google Scholar] [CrossRef] [PubMed]
- Cornet, V.; Khuyen, T.D.; Mandiki, S.N.M.; Betoulle, S.; Bossier, P.; Reyes-López, F.E.; Tort, L.; Kestemont, P. GAS1: A New β-Glucan Immunostimulant Candidate to Increase Rainbow Trout (Oncorhynchus mykiss) Resistance to Bacterial Infections with Aeromonas salmonicida achromogenes. Front. Immunol. 2021, 12, 693613. [Google Scholar] [CrossRef] [PubMed]
- Vijayaram, S.; Ringø, E.; Zuorro, A.; van Doan, H.; Sun, Y. Beneficial Roles of Nutrients as Immunostimulants in Aquaculture: A Review. Aquac. Fish 2023, in press. [CrossRef]
- Yang, G.; Zhang, Z.; Kumar, V. Editorial: Functional Feed Additives and Intestinal Health in Aquatic Animals. Front. Physiol. 2024, 15, 1385046. [Google Scholar] [CrossRef] [PubMed]
- Elkatatny, N.M.; El Nahas, A.F.; Helal, M.A.; Fahmy, H.A.; Tanekhy, M. The Impacts of Seasonal Variation on the Immune Status of Nile Tilapia Larvae and Their Response to Different Immunostimulants Feed Additives. Fish Shellfish Immunol. 2020, 96, 270–278. [Google Scholar] [CrossRef] [PubMed]
- Menanteau-Ledouble, S.; Skov, J.; Lukassen, M.B.; Rolle-Kampczyk, U.; Haange, S.-B.; Dalsgaard, I.; von Bergen, M.; Nielsen, J.L. Modulation of Gut Microbiota, Blood Metabolites, and Disease Resistance by Dietary β-Glucan in Rainbow Trout (Oncorhynchus mykiss). Anim. Microbiome 2022, 4, 58. [Google Scholar] [CrossRef]
- Machuca, C.; Méndez-Martínez, Y.; Reyes-Becerril, M.; Angulo, C. Yeast β-Glucans as Fish Immunomodulators: A Review. Animals 2022, 12, 2154. [Google Scholar] [CrossRef]
- Medagoda, N.; Chotikachinda, R.; Hasanthi, M.; Lee, K.J. Dietary Supplementation of a Mixture of Nucleotides, β-Glucan and Vitamins C and E Improved the Growth and Health Performance of Olive Flounder, Paralichthys olivaceus. Fishes 2023, 8, 302. [Google Scholar] [CrossRef]
- Abdo, S.E.; Gewaily, M.S.; Abo-Al-Ela, H.G.; Almeer, R.; Soliman, A.A.; Elkomy, A.H.; Dawood, M.A.O.; Nunes, B. Vitamin C Rescues Inflammation, Immunosuppression, and Histopathological Alterations Induced by Chlorpyrifos in Nile Tilapia. Environ. Sci. Pollut. Res. Int. 2021, 28, 28750–28763. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.S.; Koshio, S.; Kestemont, P. Recent Advances of Nucleotide Nutrition Research in Aquaculture: A Review. Rev. Aquac. 2020, 12, 1028–1053. [Google Scholar] [CrossRef]
- Rodrigues, M.V.; Zanuzzo, F.S.; Koch, J.F.A.; de Oliveira, C.A.F.; Sima, P.; Vetvicka, V. Development of Fish Immunity and the Role of β-Glucan in Immune Responses. Molecules 2020, 25, 5378. [Google Scholar] [CrossRef] [PubMed]
- Volman, J.J.; Ramakers, J.D.; Plat, J. Dietary Modulation of Immune Function by β-Glucans. Physiol. Behav. 2008, 94, 276–284. [Google Scholar] [CrossRef]
- Zhang, Y.; Guo, M.; Li, N.; Dong, Z.; Cai, L.; Wu, B.; Xie, J.; Liu, L.; Ren, L.; Shi, B. New Insights into β-Glucan-Enhanced Immunity in Largemouth Bass Micropterus salmoides by Transcriptome and Intestinal Microbial Composition. Front. Immunol. 2022, 13, 1086103. [Google Scholar] [CrossRef]
- Abu-Elala, N.M.; Ali, T.E.S.; Ragaa, N.M.; Ali, S.E.; Abd-Elsalam, R.M.; Younis, N.A.; Abdel-Moneam, D.A.; Hamdien, A.H.; Bonato, M.; Dawood, M.A.O. Analysis of the Productivity, Immunity, and Health Performance of Nile Tilapia (Oreochromis niloticus) Broodstock-Fed Dietary Fermented Extracts Sourced from Saccharomyces cerevisiae (Hilyses): A Field Trial. Animals 2021, 11, 815. [Google Scholar] [CrossRef]
- Gao, J.; Koshio, S.; Ishikawa, M.; Yokoyama, S.; Edward, R.; Mamauag, P. Interactive Effects of Vitamin C and E Supplementation on Growth Performance, Fatty Acid Composition and Reduction of Oxidative Stress in Juvenile Japanese Fl Ounder Paralichthys olivaceus Fed Dietary Oxidized Fi Sh Oil. Aquaculture 2014, 422–423, 84–90. [Google Scholar] [CrossRef]
- Abo-Al-Ela, H.G.; El-Nahas, A.F.; Mahmoud, S.; Ibrahim, E.M. Vitamin C Modulates the Immunotoxic Effect of 17α-Methyltestosterone in Nile Tilapia. Biochemistry 2017, 56, 2042–2050. [Google Scholar] [CrossRef]
- Trichet, V.V.; Santigosa, E.; Cochin, E.; Gabaudan, J. The Effect of Vitamin C on Urinary Excretion. In Dietary Nutrients, Additives, and Fish Health; Lee, C.S., Lim, C., Gatlin, D.M., III, Webster, C.D., Eds.; John Wiley & Sons, Ltd.: Haboken, NJ, USA, 2015; pp. 151–171. [Google Scholar] [CrossRef]
- NRC. Nutrient Requirements of Fish and Shrimp; National Academies Press: Washington, DC, USA, 2011; ISBN 978-0-309-16338-5. [Google Scholar]
- El-Sherif, M.S.; El-Feky, A.M.I. Performance of Nile Tilapia (Oreochromis niloticus) Fingerlings. I. Effect of PH. Int. J. Agric. Biol. 2009, 11, 297–300. [Google Scholar]
- Tran-Duy, A.; Schrama, J.W.; van Dam, A.A.; Verreth, J.A.J. Effects of Oxygen Concentration and Body Weight on Maximum Feed Intake, Growth and Hematological Parameters of Nile Tilapia, Oreochromis niloticus. Aquaculture 2008, 275, 152–162. [Google Scholar] [CrossRef]
- AOAC Association of Official Analytical Chemists. Official Methods of Analysis of the AOAC International, 22nd ed.; George, W.L., Jr., Ed.; Oxford University Press: Oxford, UK, 2022; ISBN 0197610137978-0197610138. [Google Scholar]
- Blaxhall, P.C.; Daisley, K.W. Routine Haematological Methods for Use with Fish Blood. J. Fish Biol. 2018, 5, 771–781. [Google Scholar] [CrossRef]
- Amar, E.C.; Kiron, V.; Satoh, S.; Okamoto, N. Effects of Dietary b-carotene on the immune response of rainbow trout Oncorhynchus mykiss. Fish. Sci. 2000, 66, 1068–1075. [Google Scholar] [CrossRef]
- Facimoto, C.T.; Chideroli, R.T.; Gonçalves, D.D.; do Carmo, A.O.; Kalaphotakis, E.; Pereira, U.d.P. Whole-Genome Sequence of Streptococcus agalactiae Strain S13, Isolated from a Fish Eye from a Nile Tilapia Farm in Southern Brazil. Genome Announc. 2017, 5, e00917-17. [Google Scholar] [CrossRef]
- Silva, B.C.; Martins, M.L.; Jatobá, A.; Buglione Neto, C.C.; Vieira, F.N.; Pereira, G.V.; Jerônimo, G.T.; Seiffert, W.Q.; Mouriño, J.L.P. Hematological and Immunological Responses of Nile Tilapia after Polyvalent Vaccine Administration by Different Routes. Pesqui. Veterinária Bras. 2009, 29, 874–880. [Google Scholar] [CrossRef]
- Schwaiger, J.; Wanke, R.; Adam, S.; Pawert, M.; Hönnen, W.; Triebskorn, R. The Use of Histopathological Indicators to Evaluate Contaminant-Related Stress in Fish. J. Aquat. Ecosyst. Stress Recovery 1997, 6, 75–86. [Google Scholar] [CrossRef]
- Brum, A.; Cardoso, L.; Chagas, E.C.; Chaves, F.C.M.; Mouriño, J.L.P.; Martins, M.L. Histological Changes in Nile Tilapia Fed Essential Oils of Clove Basil and Ginger after Challenge with Streptococcus agalactiae. Aquaculture 2018, 490, 98–107. [Google Scholar] [CrossRef]
- Schmidt, É.C.; dos Santos, R.; Horta, P.A.; Maraschin, M.; Bouzon, Z.L. Effects of UVB Radiation on the Agarophyte Gracilaria domingensis (Rhodophyta, Gracilariales): Changes in Cell Organization, Growth and Photosynthetic Performance. Micron 2010, 41, 919–930. [Google Scholar] [CrossRef] [PubMed]
- Langille, M.G.I.; Zaneveld, J.; Caporaso, J.G.; McDonald, D.; Knights, D.; Reyes, J.A.; Clemente, J.C.; Burkepile, D.E.; Vega Thurber, R.L.; Knight, R.; et al. Predictive Functional Profiling of Microbial Communities Using 16S RRNA Marker Gene Sequences. Nat. Biotechnol. 2013, 31, 814–821. [Google Scholar] [CrossRef] [PubMed]
- Babicki, S.; Arndt, D.; Marcu, A.; Liang, Y.; Grant, J.R.; Maciejewski, A.; Wishart, D.S. Heatmapper: Web-Enabled Heat Mapping for All. Nucleic Acids Res. 2016, 44, W147–W153. [Google Scholar] [CrossRef]
- Koch, J.F.A.; de Oliveira, C.A.F.; Zanuzzo, F.S. Dietary β-Glucan (MacroGard®) Improves Innate Immune Responses and Disease Resistance in Nile Tilapia Regardless of the Administration Period. Fish Shellfish Immunol. 2021, 112, 56–63. [Google Scholar] [CrossRef]
- Sabioni, R.E.; Zanuzzo, F.S.; Gimbo, R.Y.; Urbinati, E.C. β-Glucan Enhances Respiratory Activity of Leukocytes Suppressed by Stress and Modulates Blood Glucose Levels in Pacu (Piaractus mesopotamicus). Fish Physiol. Biochem. 2020, 46, 629–640. [Google Scholar] [CrossRef] [PubMed]
- Reda, R.M.; Selim, K.M.; Mahmoud, R.; El-Araby, I.E. Effect of Dietary Yeast Nucleotide on Antioxidant Activity, Non-Specific Immunity, Intestinal Cytokines, and Disease Resistance in Nile Tilapia. Fish Shellfish Immunol. 2018, 80, 281–290. [Google Scholar] [CrossRef] [PubMed]
- Shiau, S.Y.; Gabaudan, J.; Lin, Y.H. Dietary Nucleotide Supplementation Enhances Immune Responses and Survival to Streptococcus Iniae in Hybrid Tilapia Fed Diet Containing Low Fish Meal. Aquac. Rep. 2015, 2, 77–81. [Google Scholar] [CrossRef]
- Bowyer, P.H.; El-Haroun, E.R.; Hassaan, M.; Salim, H.; Davies, S.J. Dietary Nucleotides Enhance Growth Performance, Feed Efficiency and Intestinal Functional Topography in European Seabass (Dicentrarchus labrax). Aquac. Res. 2019, 50, 1921–1930. [Google Scholar] [CrossRef]
- Álvarez-Rodríguez, M.; Pereiro, P.; Reyes-López, F.E.; Tort, L.; Figueras, A.; Novoa, B. Analysis of the Long-Lived Responses Induced by Immunostimulants and Their Effects on a Viral Infection in Zebrafish (Danio rerio). Front. Immunol. 2018, 9, 388727. [Google Scholar] [CrossRef] [PubMed]
- Gandar, A.; Laffaille, P.; Canlet, C.; Tremblay-Franco, M.; Gautier, R.; Perrault, A.; Gress, L.; Mormède, P.; Tapie, N.; Budzinski, H.; et al. Adaptive Response under Multiple Stress Exposure in Fish: From the Molecular to Individual Level. Chemosphere 2017, 188, 60–72. [Google Scholar] [CrossRef]
- Xu, C.M.; Yu, H.R.; Li, L.Y.; Li, M.; Qiu, X.Y.; Fan, X.Q.; Fan, Y.L.; Shan, L.L. Effects of Dietary Vitamin C on the Growth Performance, Biochemical Parameters, and Antioxidant Activity of Coho Salmon Oncorhynchus kisutch (Walbaum, 1792) Postsmolts. Genet. Res. 2022, 2022, 6866578. [Google Scholar] [CrossRef] [PubMed]
- Izquierdo, M.; Jiménez, J.I.; Saleh, R.; Hernández-Cruz, C.M.; Domínguez, D.; Zamorano, M.J.; Hamre, K. Interaction between Taurine, Vitamin E and Vitamin C in Microdiets for Gilthead Seabream (Sparus aurata) Larvae. Aquaculture 2019, 498, 246–253. [Google Scholar] [CrossRef]
- Jomova, K.; Alomar, S.Y.; Alwasel, S.H.; Nepovimova, E.; Kuca, K.; Valko, M. Several Lines of Antioxidant Defense against Oxidative Stress: Antioxidant Enzymes, Nanomaterials with Multiple Enzyme-Mimicking Activities, and Low-Molecular-Weight Antioxidants. Arch. Toxicol. 2024, 98, 1323–1367. [Google Scholar] [CrossRef]
- Nicholson, J.K.; Holmes, E.; Kinross, J.; Burcelin, R.; Gibson, G.; Jia, W.; Pettersson, S. Host-Gut Microbiota Metabolic Interactions. Science 2012, 336, 1262–1267. [Google Scholar] [CrossRef]
- Vu, V.; Muthuramalingam, K.; Singh, V.; Choi, C.; Kim, Y.M.; Unno, T.; Cho, M. Schizophyllum Commune-Derived β-Glucan Improves Intestinal Health Demonstrating Protective Effects against Constipation and Common Metabolic Disorders. Appl. Biol. Chem. 2022, 65, 9. [Google Scholar] [CrossRef]
- Subramanian, S.; MacKinnon, S.L.; Ross, N.W. A Comparative Study on Innate Immune Parameters in the Epidermal Mucus of Various Fish Species. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2007, 148, 256–263. [Google Scholar] [CrossRef] [PubMed]
- Elvy, J.E.; Symonds, J.E.; Hilton, Z.; Walker, S.P.; Tremblay, L.A.; Casanovas, P.; Herbert, N.A. The Relationship of Feed Intake, Growth, Nutrient Retention, and Oxygen Consumption to Feed Conversion Ratio of Farmed Saltwater Chinook Salmon (Oncorhynchus tshawytscha). Aquaculture 2022, 554, 738184. [Google Scholar] [CrossRef]
- Carballo, C.; Pinto, P.I.S.; Mateus, A.P.; Berbel, C.; Guerreiro, C.C.; Martinez-Blanch, J.F.; Codoñer, F.M.; Mantecon, L.; Power, D.M.; Manchado, M. Yeast β-Glucans and Microalgal Extracts Modulate the Immune Response and Gut Microbiome in Senegalese Sole (Solea senegalensis). Fish Shellfish Immunol. 2019, 92, 31–39. [Google Scholar] [CrossRef] [PubMed]
- de Souza, F.P.; de Lima, E.C.S.; Pandolfi, V.C.F.; Leite, N.G.; Furlan-Murari, P.J.; Leal, C.N.S.; Mainardi, R.M.; Suphoronski, S.A.; Favero, L.M.; Koch, J.F.A.; et al. Effect of β-Glucan in Water on Growth Performance, Blood Status and Intestinal Microbiota in Tilapia under Hypoxia. Aquac. Rep. 2020, 17, 100369. [Google Scholar] [CrossRef]
- Ofek, T.; Lalzar, M.; Laviad-Shitrit, S.; Izhaki, I.; Halpern, M. Comparative Study of Intestinal Microbiota Composition of Six Edible Fish Species. Front. Microbiol. 2021, 12, 760266. [Google Scholar] [CrossRef]
- Zhao, N.; Guo, J.; Zhang, B.; Liu, K.; Liu, Y.; Shen, Y.; Li, J. Heterogeneity of the Tissue-Specific Mucosal Microbiome of Normal Grass Carp (Ctenopharyngodon idella). Mar. Biotechnol. 2022, 24, 366–379. [Google Scholar] [CrossRef]
- Salger, S.A.; Reza, J.; Dec, C.A.; Wahab, M.A.; Baltzegar, D.A.; Mur, A.T.; Borsk, R.J. Enhanced Biodiversity of Gut Flora and Feed Efficiency in Pond Cultured Tilapia under Reduced Frequency Feeding Strategies. PLoS ONE 2020, 15, e0236100. [Google Scholar] [CrossRef]
- Zhang, Z.; Fan, Z.; Yi, M.; Liu, Z.; Ke, X.; Gao, F.; Cao, J.; Wang, M.; Chen, G.; Lu, M. Characterization of the Core Gut Microbiota of Nile Tilapia (Oreochromis niloticus): Indication of a Putative Novel Cetobacterium Species and Analysis of Its Potential Function on Nutrition. Arch. Microbiol. 2022, 204, 690. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient, g·kg−1 | Control | Immunostimulant |
|---|---|---|
| Soybean meal | 485.0 | 485.0 |
| Poultry meal | 115.0 | 115.0 |
| Broken rice | 244.0 | 224.0 |
| Corn | 145.5 | 125.5 |
| Soybean oil | 7.0 | 7.0 |
| Choline chloride | 2.0 | 2.0 |
| Premix 1 | 1.5 | 1.5 |
| Functional immunostimulant mixture 2 | 0.0 | 40.0 |
| Centesimal Composition, % | ||
| Moisture | 12.8 | 11.0 |
| Crude protein 3 | 35.0 | 35.8 |
| Ether extract 3 | 5.5 | 5.2 |
| Mineral matter 3 | 5.4 | 7.8 |
| Control | Immunostimulant | p Value | |
|---|---|---|---|
| Final weight, g | 18.46 ± 1.28 | 29.53 ± 1.62 | 0.0008 |
| Weight gain, g | 16.79 ± 0.99 | 27.55 ± 1.63 | 0.0006 |
| Daily weight gain, g | 0.33 ± 0.01 | 0.55 ± 0.04 | 0.0006 |
| Food conversion | 1.22 ± 0.06 | 0.96 ± 0.03 | 0.0089 |
| Protein retention (PR, %) | 25.57 ± 4.16 | 52.58 ± 6.16 | 0.0033 |
| Survival, % | 75.33 ± 29.97 | 91.00 ± 12.57 | 0.5347 |
| Centesimal Composition (g·kg−1) | |||
|---|---|---|---|
| Control | Immunostimulant | p Value | |
| Moisture | 72.55 ± 0.512 | 73.26 ± 0.829 | 0.1901 |
| Crude protein | 54.47 ± 3.273 | 56.76 ± 2.296 | 0.119 |
| Ether extract | 25.05 ± 0.093 | 27.11 ± 1.61 | 0.0973 |
| Mineral matter | 1.70 ± 0.305 | 2.21 ± 0.184 | 0.0376 |
| Hematological Parameters | Control | Immunostimulant | p Value |
|---|---|---|---|
| Erythrocyte (cell mL−1) 106 | 1.53 ± 0.10 | 1.65 ± 0.54 | 0.1608 |
| Hematocrit (%) | 28.83 ± 1.12 | 30.81 ± 3.51 | 0.4073 |
| Hemoglobin (g.dL−1) | 14.95 ± 7.53 | 16.40 ± 9.6 | 0.7385 |
| MCV (10−4 fL) | 1.89 ± 0.88 | 1.89 ± 2.80 | >0.9999 |
| MCH (105 pg) | 0.96 ± 4.3 | 1.05 ± 0.47 | 0.8159 |
| MCHC (g dL−1) | 46.27 ± 15.07 | 57.07 ± 30.38 | 0.6107 |
| Immunological Parameters | |||
| Total protein (mg·mL−1) | 46.29 ± 8.00 | 36.07 ± 2.19 | 0.0999 |
| Total Ig (mg·mL−1) | 21.39 ± 10.22 | 16.93 ± 2.39 | 0.5025 |
| Agglutinating (Log2) | 4.25 ± 0.57 | 5.085 ± 0.50 | 0.1318 |
| MIC (Log2) | 3.55 ± 0.00 | 3.55 ± 0.00 | 0.9999 |
| Control | Immunostimulant | p Value | |
|---|---|---|---|
| Intestinal Morphology | |||
| Number of intestinal folds | 36.22 ± 8.95 | 43.33 ± 1.15 | 0.2441 |
| Length (µm) | 22.76 ± 8.29 | 37.88 ± 4.20 | 0.0481 |
| Width (µm) | 10.56 ± 2.85 | 17.29 ± 2.89 | 0.0380 |
| Number of goblet cells (×10) | 311.11 ± 69.60 | 501.55 ± 83.82 | 0.0389 |
| Total area (×103 µm2) | 31.19 ± 15.91 | 38.76 ± 9.95 | 0.3190 |
| Total perimeter (×103 µm) | 4.35 ± 1.16 | 5.02 ± 0.57 | 0.3142 |
| Histological alterations | |||
| Eosinophilic infiltrate | 1.11 ± 0.19 | 1.22 ± 0.83 | 0.2302 |
| Loose melanomacrophages | 0.08 ± 0.19 | 0.33 ± 0.57 | 0.3739 |
| Histological Alterations in Liver | Control | Immunostimulant | p Value |
|---|---|---|---|
| Cordonal appearance | 1.44 ± 0.50 | 2.00 ± 0.66 | 0.315 |
| Balloon appearance in the hepatocytes | 2.11 ± 0.50 | 1.77 ± 0.19 | 0.348 |
| Congestion of the pancreas | 1.55 ± 0.19 | 1.44 ± 0.19 | 0.518 |
| Congestion of the sinusoids | 1.00 ± 0.33 | 1.44 ± 0.69 | 0.373 |
| Eosinophilic infiltrate | 1.44 ± 0.38 | 1.77 ± 0.19 | 0.250 |
| Lymphocytic infiltrate | 0.55 ± 0.69 | 0.22 ± 0.19 | 0.467 |
| Hypertrophy of the hepatocytes | 2.11 ± 0.50 | 1.55 ± 0.86 | 0.351 |
| Macrosteatosis | 2.22 ± 0.50 | 2.00 ± 0.57 | 0.643 |
| Microsteatosis | 0.22 ± 0.38 | 0.33 ± 0.57 | 0.795 |
| Necrosis | 2.11 ± 0.83 | 1.56 ± 0.50 | 0.382 |
| Histological Alterations in Spleen | Control | Immunostimulant | p Value |
| Integrity of the white pulp | 2.83 ± 0.28 | 2.83 ± 0.28 | >0.999 |
| Integrity of the red pulp | 2.83 ± 0.28 | 2.83 ± 0.28 | >0.999 |
| Centers of melanomacrophages | 1.167 ± 0.76 | 1.00 ± 0.00 | 0.724 |
| Loose melanomacrophages | 0.83 ± 0.28 | 1.00 ± 0.00 | 0.373 |
| Necrosis | 0.83 ± 0.76 | 0.83 ± 0.76 | >0.999 |
| Samples | Read Count | OTU | Chao1 | Shannon | Simpson | Good’s Coverage |
|---|---|---|---|---|---|---|
| Control | 16.628 | 11 | 21 | 0.21 | 0.089 | 99.98% |
| Immunomodulator | 32.904 | 28 | 32.5 | 0.14 | 0.042 | 99.95% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sá, L.S.d.; Jerônimo, G.T.; Soligo, T.; Yamashita, E.; Machado Fracalossi, D.; Martins, M.L.; Mouriño, J.L.P. The Zootechnical Performance, Health State Modulation, Morphology, and Intestinal Microbiome of Nile Tilapia Juveniles Fed with a Functional Blend of Immunostimulants Associated with a Diet High in Soybean Meal. Fishes 2024, 9, 212. https://doi.org/10.3390/fishes9060212
Sá LSd, Jerônimo GT, Soligo T, Yamashita E, Machado Fracalossi D, Martins ML, Mouriño JLP. The Zootechnical Performance, Health State Modulation, Morphology, and Intestinal Microbiome of Nile Tilapia Juveniles Fed with a Functional Blend of Immunostimulants Associated with a Diet High in Soybean Meal. Fishes. 2024; 9(6):212. https://doi.org/10.3390/fishes9060212
Chicago/Turabian StyleSá, Lúvia Souza de, Gabriela Tomas Jerônimo, Thiago Soligo, Eduardo Yamashita, Débora Machado Fracalossi, Maurício Laterça Martins, and José Luiz Pedreira Mouriño. 2024. "The Zootechnical Performance, Health State Modulation, Morphology, and Intestinal Microbiome of Nile Tilapia Juveniles Fed with a Functional Blend of Immunostimulants Associated with a Diet High in Soybean Meal" Fishes 9, no. 6: 212. https://doi.org/10.3390/fishes9060212
APA StyleSá, L. S. d., Jerônimo, G. T., Soligo, T., Yamashita, E., Machado Fracalossi, D., Martins, M. L., & Mouriño, J. L. P. (2024). The Zootechnical Performance, Health State Modulation, Morphology, and Intestinal Microbiome of Nile Tilapia Juveniles Fed with a Functional Blend of Immunostimulants Associated with a Diet High in Soybean Meal. Fishes, 9(6), 212. https://doi.org/10.3390/fishes9060212