
Diets, Condition, and Reproductive Fitness of Slimy Sculpin (Uranidea cognata) after Catastrophic Flooding in Trout Streams in Southeastern Minnesota, USA


Abstract
1. Introduction
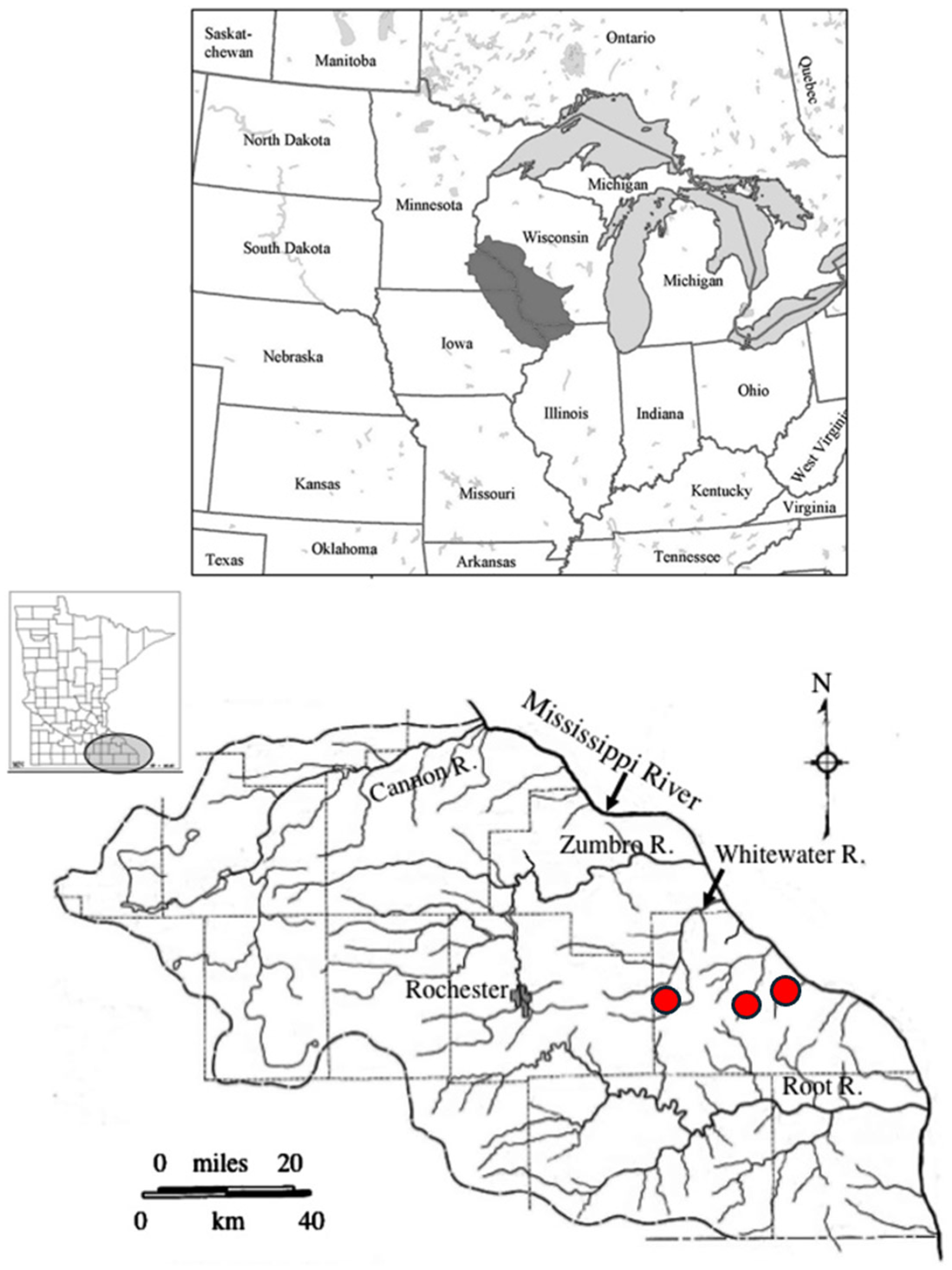
2. Study Sites and Flooding
3. Methods and Materials
3.1. Field Work
3.2. Laboratory Work
3.3. Data Analyses
4. Results
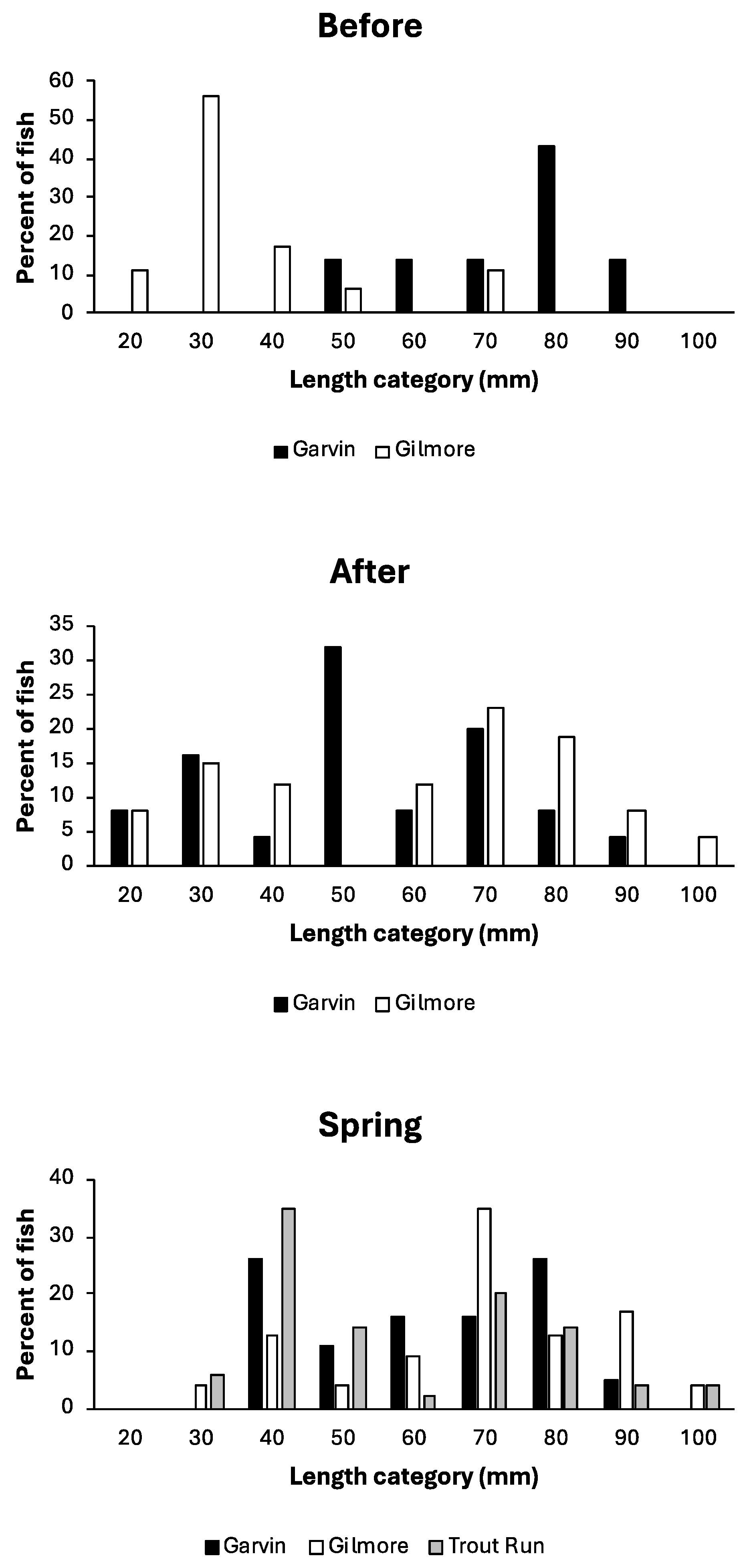
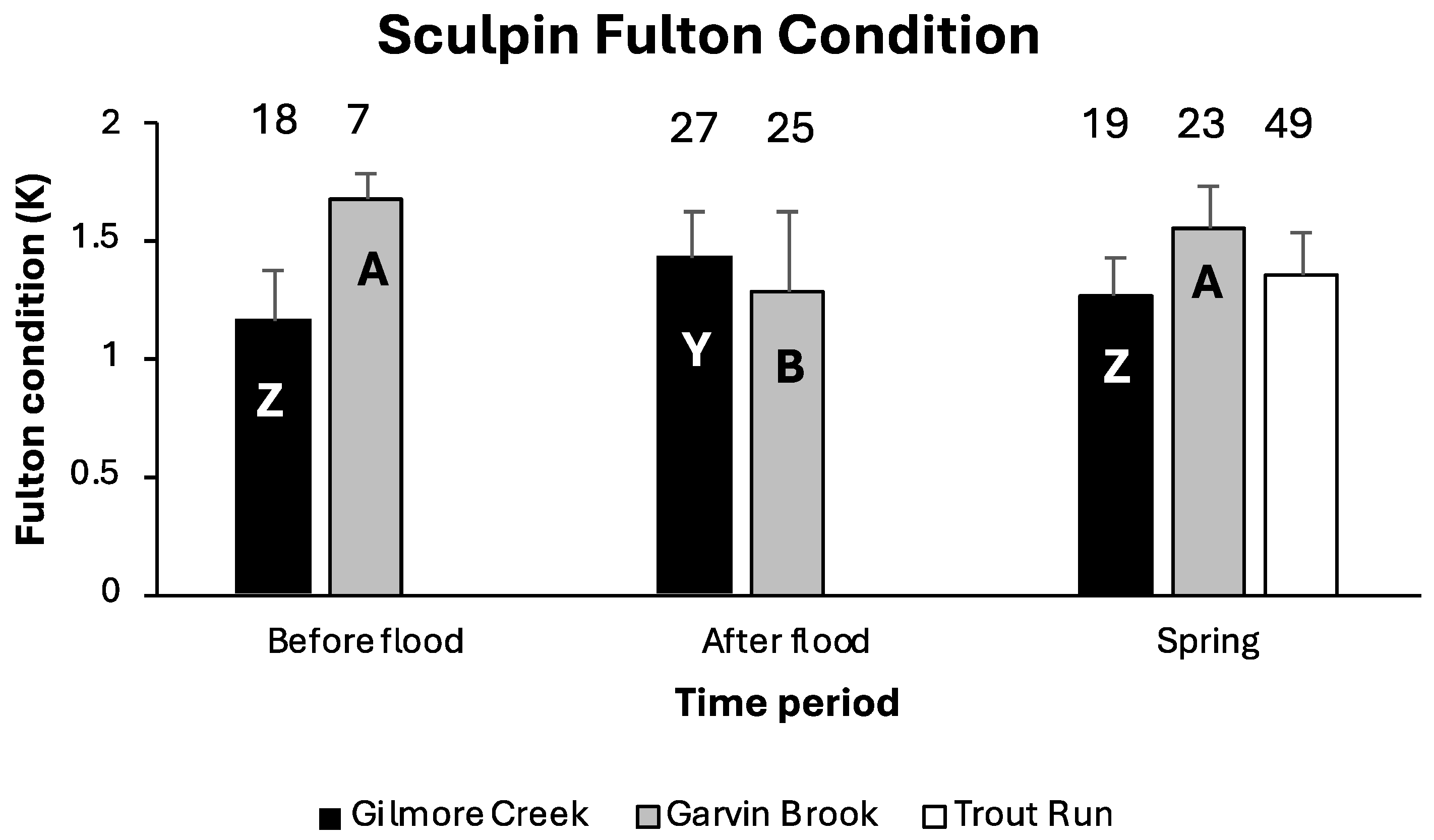
4.1. Sculpin Size and Body Condition
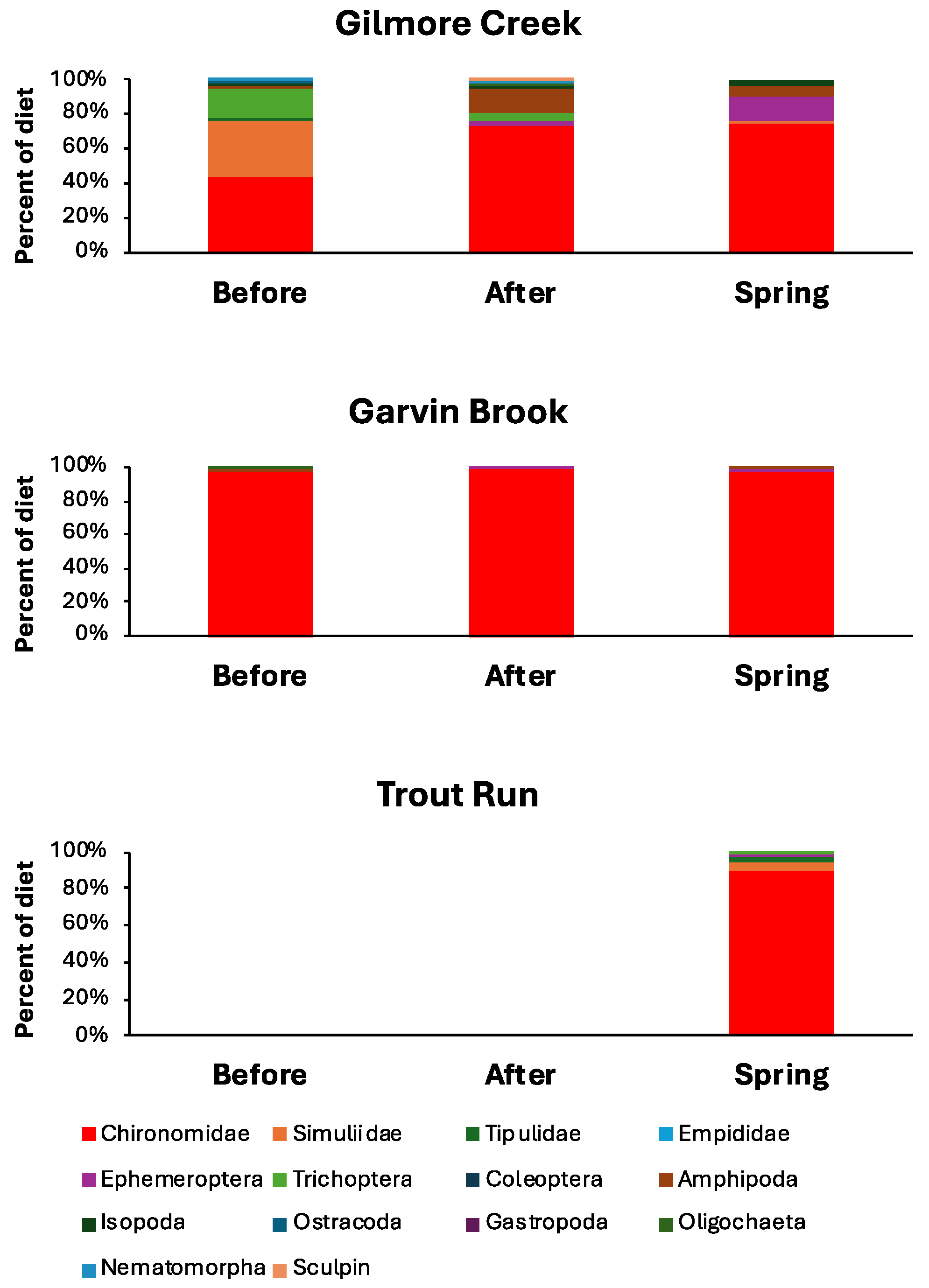
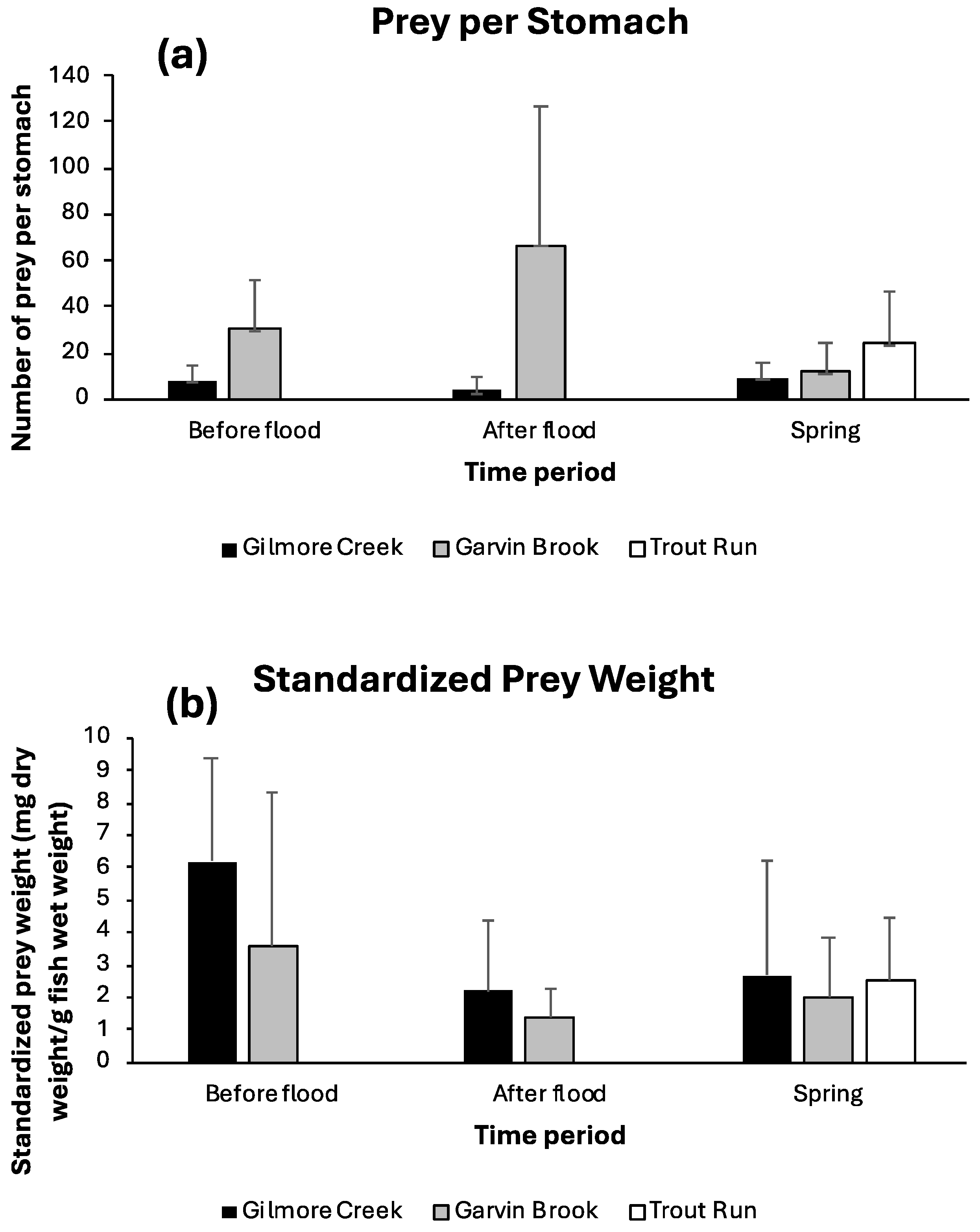
4.2. Sculpin Diets
4.3. Sculpin Size, Condition, and Reproductive Variables during Spring Pre-Pawn
5. Discussion
6. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hickey, J.T.; Salas, J.D. Environmental effects of extreme floods. Hydrometeorol. Impacts Manag. Extreme Floods 1995, 7, 13–17. [Google Scholar]
- Schultz, A.A.; Maughan, O.E.; Bonar, S.A.; Matter, W.J. Effects of flooding on abundance of native and nonnative fishes downstream from a small impoundment. N. Am. J. Fish. Manag. 2003, 23, 503–511. [Google Scholar] [CrossRef]
- Rayner, T.S.; Pusey, B.J.; Pearson, R.G. Seasonal flooding, instream habitat structure and fish assemblages in the Mulgrave River, north-east Queensland: Towards a new conceptual framework for understanding fish-habitat dynamics in small tropical rivers. Mar. Freshw. Res. 2008, 59, 97–116. [Google Scholar] [CrossRef]
- Eby, L.A.; Fagan, W.F.; Minckley, W.L. Variability and dynamics of a desert stream community. Ecol. Appl. 2003, 13, 1566–1579. [Google Scholar] [CrossRef]
- George, S.D.; Baldigo, B.P.; Smith, A.J.; Robinson, G.R. Effects of extreme floods on trout populations and fish communities in a Catskill Mountain river. Freshw. Biol. 2015, 60, 2511–2522. [Google Scholar] [CrossRef]
- Adams, S.M.; Greeley, M.S.; Law, J.M.; Noga, E.J.; Zelikoff, J.T. Application of multiple sublethal stress indicators to assess the health of fish in Pamlico Sound following extensive flooding. Estuaries 2003, 26, 1365–1382. [Google Scholar] [CrossRef]
- Blewett, D.A.; Stevens, P.W.; Carter, J. Ecological effects of river flooding on abundance and body condition of a large, euryhaline fish. Mar. Ecol. Prog. Ser. 2017, 563, 211–218. [Google Scholar] [CrossRef]
- Pearsons, T.N.; Li, H.W.; Lamberti, G.A. Influence of habitat complexity on resistance to flooding and resilience of stream fish assemblages. Trans. Am. Fish. Soc. 1992, 121, 427–436. [Google Scholar] [CrossRef]
- Mundahl, N.D.; Hunt, A.M. Recovery of stream invertebrates after catastrophic flooding in southeastern Minnesota, USA. J. Freshw. Ecol. 2011, 26, 445–457. [Google Scholar] [CrossRef]
- King, A.J.; Humphries, P.; Lake, P.S. Fish recruitment on floodplains: The roles of patterns of flooding and life history characteristics. Can. J. Fish. Aquat. Sci. 2003, 60, 773–786. [Google Scholar] [CrossRef]
- Pereira, L.S.; Tencatt, L.F.C.; Dias, R.M.; de Oliveira, A.G.; Agostinho, A.A. Effects of long and short flooding years on the feeding ecology of piscivorous fish in floodplain river systems. Hydrobiologia 2017, 795, 65–80. [Google Scholar] [CrossRef]
- Death, R.D. The effect of floods on aquatic invertebrate communities. In Aquatic Insects: Challenges to Populations; Lancaster, J., Briers, R.A., Eds.; Proceedings of the Royal Society’s 24th Symposium; CAB International: Oxfordshire, UK, 2008; pp. 103–121. [Google Scholar]
- Allan, J.D.; Castillo, M.M.; Cappas, K.A. Stream Ecology: Structure and Function of Running Waters, 3rd ed.; Springer Nature Switzerland: Cham, Switzerland, 2021. [Google Scholar]
- Naiman, R.J.; Décamps, H.; McClain, M.E. Riparia: Ecology, Conservation, and Management of Streamside Communities; Elsevier Academic Press: Burlington, MA, USA, 2005. [Google Scholar]
- Yin, Y.; Nelson, J.C.; Swenson, G.V.; Langrehr, H.A.; Blackburn, T.A. Tree mortality in the upper Mississippi River and floodplain following an extreme flood in 1993. In Long Term Resource Monitoring Program 1993 Flood Observations (LTRMP 94-S011); National Biological Service, Environmental Management Technical Center: Onalaska, WI, USA, 1994. [Google Scholar]
- Carlson, A.K.; Fincel, M.J.; Longhenry, C.M.; Graeb, B.D.S. Effects of flooding on fishes and aquatic habitats in a Missouri River delta. J. Freshw. Ecol. 2016, 31, 271–288. [Google Scholar] [CrossRef]
- Harrell, H.L. Response of the Devil’s River (Texas) fish community to flooding. Copeia 1978, 1978, 60–68. [Google Scholar] [CrossRef]
- Piniewski, M.; Prudhomme, C.; Acreman, M.C.; Tylec, L.; Oglęcki, P.; Okruszko, T. Responses of fish and invertebrates to floods and droughts in Europe. Ecohydrology 2017, 10, e1793. [Google Scholar] [CrossRef]
- Talbot, J.J.; Bennett, E.M.; Cassell, K.; Hanes, D.M.; Minor, E.C.; Paerl, H.; Raymond, P.A.; Vargas, R.; Vidon, P.G.; Wollheim, W.; et al. The impact of flooding on aquatic ecosystem services. Biogeochemistry 2018, 141, 439–461. [Google Scholar] [CrossRef] [PubMed]
- Edwards, P.A.; Cunjak, R.A. Influence of water temperature and streambed stability on the abundance and distribution of slimy sculpin (Cottus cognatus). Environ. Biol. Fishes 2006, 80, 9–22. [Google Scholar] [CrossRef]
- Erman, D.C.; Andrews, E.D.; Yoder-Williams, M. Effects of winter floods on fishes in the Sierra Nevada. Can. J. Fish. Aquat. Sci. 1988, 45, 2195–2200. [Google Scholar] [CrossRef]
- Harvey, B.C. Susceptibility of young-of-the-year fishes to downstream displacement by flooding. Trans. Am. Fish. Soc. 1987, 116, 851–855. [Google Scholar] [CrossRef]
- Berra, T.M. Freshwater Fish Distribution; Academic Press: San Diego, CA, USA, 2001. [Google Scholar]
- Nelson, J.S. Fishes of the World, 4th ed.; John Wiley & Sons: Hoboken, NJ, USA, 2006. [Google Scholar]
- Baldigo, B.P.; Lawrence, G.B. Composition of fish communities in relation to stream acidification and habitat in the Neversink River, New York. Trans. Am. Fish. Soc. 2000, 129, 60–76. [Google Scholar] [CrossRef]
- Wallace, R.L.; McAllister, D.E.; Rankin, M. Cottus cognatus Richardson, slimy sculpin. In Atlas of North American Freshwater Fishes; Lee, D.S., Gilbert, C.R., Hocutt, C.H., Jenkins, R.E., McAllister, D.E., Stauffer, J.R., Jr., Eds.; North Carolina Biological Survey Publication #1980-12; North Carolina Museum of Natural Sciences: Raleigh, NC, USA, 1980; p. 808. [Google Scholar]
- Page, L.M.; Burr, B.M. A Field Guide to Freshwater Fishes: North America North of Mexico; Houghton Mifflin Company: Boston, MA, USA, 1991. [Google Scholar]
- Gray, M.A.; Munkittrick, K.R. An effects-based assessment of slimy sculpin (Cottus cognatus) populations in agricultural regions of northwestern New Brunswick. Water Qual. Res. J. Can. 2005, 40, 16–27. [Google Scholar] [CrossRef]
- Gray, M.A.; Curry, R.A.; Arciszewski, T.J.; Munkittrick, K.R.; Brasfield, S.M. The biology and ecology of slimy sculpin: A recipe for effective environmental monitoring. Facets 2018, 3, 103–127. [Google Scholar] [CrossRef]
- Keillor, L. Fishing after the flood. Minn. Conserv. Vol. 2010, 73, 8–17. [Google Scholar]
- Cochran, P.A.; Stagg, T.W. Response of a fish assemblage to severe flooding in Gilmore Creek, a southeastern Minnesota trout stream. J. Freshw. Ecol. 2011, 26, 77–84. [Google Scholar] [CrossRef]
- Ney, J.J. Practical use of biological statistics. In Inland Fisheries Management in North America, 2nd ed.; Kohler, C.C., Hubert, W.A., Eds.; American Fisheries Society: Bethesda, MD, USA, 1999; pp. 167–191. [Google Scholar]
- Kaller, M.D.; Green, C.C.; Haukenes, A.H. Growth and development. In Methods for Fish Biology, 2nd ed.; Midway, S.R., Hasler, C.T., Chakrabarty, P., Eds.; American Fisheries Society: Bethesda, MD, USA, 2022; pp. 347–398. [Google Scholar]
- Hilsenhoff, W.L. Aquatic Insects of Wisconsin: Generic Keys and Notes on Biology, Ecology and Distribution; Technical Bulletin No. 89; Wisconsin Department of Natural Resources: Madison, WI, USA, 1975. [Google Scholar]
- Pennak, R.W. Fresh-Water Invertebrates of the United States, 2nd ed.; John Wiley & Sons: New York, NY, USA, 1978. [Google Scholar]
- Luckenbach, J.A.; Guzmán, J.M. Reproduction. In Methods for Fish Biology, 2nd ed.; Midway, S.R., Hasler, C.T., Chakrabarty, P., Eds.; American Fisheries Society: Bethesda, MD, USA, 2022; pp. 399–448. [Google Scholar]
- Neumann, R.M.; Allen, M.S. Size structure. In Analysis and Interpretation of Freshwater Fisheries Data; Guy, C.S., Brown, M.L., Eds.; American Fisheries Society: Bethesda, MD, USA, 2007; pp. 375–421. [Google Scholar]
- Brower, J.E.; Zar, J.H.; von Ende, C.N. Field and Laboratory Methods for General Ecology, 4th ed.; WCB McGraw-Hill: Boston, MA, USA, 1998. [Google Scholar]
- Mundahl, N.D.; Mundahl, D.E.; Merten, E.C. Success of slimy sculpin reintroductions in Minnesota trout streams: Influence of feeding and diets. Am. Midl. Nat. 2012, 168, 162–183. [Google Scholar] [CrossRef]
- Gray, M.A.; Curry, A.R.; Munkittrick, K.R. Non-lethal sampling methods for assessing environmental impacts using a small-bodied sentinel fish species. Water Qual. Res. J. Can. 2002, 37, 195–211. [Google Scholar] [CrossRef]
- Strachura, S. Six Dead as Flooding Hits SE Minnesota. Minnesota Public Radio News, 19 August 2007. Available online: https://www.mprnews.org/story/2007/08/19/flood (accessed on 5 April 2024).
- Power, M.E.; Parker, M.S.; Dietrich, W.E. Seasonal reassembly of a river food web: Floods, droughts, and impacts of fish. Ecol. Monogr. 2008, 78, 263–282. [Google Scholar] [CrossRef]
- Clarke, A.D.; Telmer, K.H.; Shrimpton, J.M. Movement patterns of fish revealed by otolith microchemistry: A comparison of putative migratory and resident species. Environ. Biol. Fish. 2015, 98, 1583–1597. [Google Scholar] [CrossRef]
- Weinstein, S.Y.; Coombs, J.A.; Nislow, K.H.; Riley, C.; Roy, A.H.; Whiteley, A.R. Evaluating the effects of barriers on slimy sculpin movement and population connectivity using novel sibship-based and traditional genetic metrics. Trans. Am. Fish. Soc. 2019, 148, 1117–1131. [Google Scholar] [CrossRef]
- Mundahl, N.D. Recovery of slimy sculpin (Uranidea cognata) after an autumn fish kill in the headwaters of a Minnesota trout stream. Water 2024, 16, 283. [Google Scholar] [CrossRef]
- Marsden, J.E.; Tobi, H. Sculpin predation on lake trout eggs in interstices: Skull compression as a novel foraging mechanism. Copeia 2014, 2014, 654–658. [Google Scholar] [CrossRef]
- Waters, T. The Streams and Rivers of Minnesota; University of Minnesota Press: Minneapolis, MN, USA, 1977. [Google Scholar]
- Ali, M.; Nicieza, A.; Wootton, R.J. Compensatory growth in fishes: A response to growth depression. Fish Fish. 2003, 4, 147–190. [Google Scholar] [CrossRef]
- McGinley, E.J.; Raesley, R.L.; Seddon, W.L. The effects of embeddedness on the seasonal feeding of mottled sculpin. Am. Midl. Nat. 2013, 170, 213–228. [Google Scholar] [CrossRef]
- Hards, A.R.; Gray, M.A.; Noël, S.C.; Cunjak, R.A. Utility of condition indices as predictors of lipid content in slimy sculpin (Cottus cognatus). Diversity 2019, 11, 71. [Google Scholar] [CrossRef]
- Mousseau, T.A.; Collins, N.C.; Cabana, G. A comparative study of sexual selection and reproductive investment in the slimy sculpin, Cottus cognatus. Oikos 1987, 51, 156–162. [Google Scholar] [CrossRef]
- Diana, J.S. Biology and Ecology of Fishes, 2nd ed.; Cooper Publishing Group: Traverse City, MI, USA, 2004. [Google Scholar]
- Petty, J.T.; Grossman, G.D. Patch selection by mottled sculpin (Pisces: Cottidae) in a southern Appalachian stream. Freshw. Biol. 1996, 35, 261–276. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stream | Total Length (mm) | Wet Mass (g) |
|---|---|---|
| Gilmore Creek | 27–101 | 0.2–10.1 |
| Garvin Brook | 25–100 | 0.1–15.6 |
| Trout Run | 38–101 | 0.8–18.0 |
| F2,167 = 2.62 | F2,167 = 2.98 | |
| p = 0.076 | p = 0.053 |
| Taxa | Gilmore Creek | Garvin Brook | Trout Run | |
|---|---|---|---|---|
| DIPTERA | Chironomidae | X | X | X |
| Simuliidae | X | X | X | |
| Tipula | X | |||
| Dicranota | X | |||
| Empididae | X | X | ||
| EPHEMEROPTERA | X | X | X | |
| TRICHOPTERA | Brachycentrus | X | ||
| Hydropsyche | X | X | ||
| Limnephilus | X | X | ||
| Glossosoma | X | |||
| COLEOPTERA | Agabus | X | ||
| Unid. beetle | X | |||
| CRUSTACEA | Amphipoda | X | X | |
| Isopoda | X | X | ||
| Ostracoda | X | |||
| GASTROPODA | X | |||
| OLIGOCHAETA | X | X | X | |
| NEMATOMORPHA | X | |||
| FISH | X |
| Period/Variable | Garvin | Gilmore | Trout Run |
|---|---|---|---|
| Before flooding | |||
| Taxa/fish | 1.7 (0.8) | 2.3 (0.8) | - |
| Shannon diversity | 0.075 | 0.624 | - |
| % empty stomachs | 0.0 | 0.0 | - |
| After flooding | |||
| Taxa/fish | 1.2 (0.6) | 1.4 (0.8) | - |
| Shannon diversity | 0.016 | 0.458 | - |
| % empty stomachs | 4.0 | 11.1 | - |
| Spring | |||
| Taxa/fish | 1.3 (0.5) | 2.2 (1.0) | 2.0 (1.1) |
| Shannon diversity | 0.075 | 0.387 | 0.197 |
| % empty stomachs | 4.3 | 5.3 | 4.1 |
| Variable | Sex | Gilmore Creek | Garvin Brook | Trout Run | F | p |
|---|---|---|---|---|---|---|
| n | F | 9 | 11 | 28 | ||
| M | 9 | 10 | 21 | |||
| Total length | F | 74 (8) AB | 73 (7) B | 77 (5) A | 3.52 | 0.038 |
| (mm) | M | 84 (5) | 92 (8) | 91 (8) | 2.20 | 0.125 |
| K | F | 1.25 (0.16) B | 1.62 (0.11) A | 1.33 (0.15) B | 21.06 | <0.001 |
| M | 1.29 (0.16) B | 1.55 (0.21) A | 1.41 (0.18) AB | 5.01 | 0.012 | |
| GSI (%) | F | 14.74 (1.75) | 17.28 (1.63) | 15.99 (2.13) | 3.33 | 0.052 |
| M | 1.82 (0.21) | 1.93 (0.42) | 1.94 (0.57) | 0.15 | 0.864 | |
| HSI (%) | F | 2.72 (0.43) | 3.28 (0.49) | 2.95 (0.21) | 2.65 | 0.103 |
| M | 1.47 (0.18) | 1.51 (0.29) | 1.53 (0.06) | 0.11 | 0.896 | |
| Oocyte number | F | 221 (61) | 271 (61) | 223 (60) | 2.18 | 0.134 |
| Mean oocyte wet mass (mg) | F | 3.7 (0.3) C | 4.1 (0.6) B | 4.8 (1.0) A | 4.14 | 0.027 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mundahl, N.D. Diets, Condition, and Reproductive Fitness of Slimy Sculpin (Uranidea cognata) after Catastrophic Flooding in Trout Streams in Southeastern Minnesota, USA. Fishes 2024, 9, 197. https://doi.org/10.3390/fishes9060197
Mundahl ND. Diets, Condition, and Reproductive Fitness of Slimy Sculpin (Uranidea cognata) after Catastrophic Flooding in Trout Streams in Southeastern Minnesota, USA. Fishes. 2024; 9(6):197. https://doi.org/10.3390/fishes9060197
Chicago/Turabian StyleMundahl, Neal D. 2024. "Diets, Condition, and Reproductive Fitness of Slimy Sculpin (Uranidea cognata) after Catastrophic Flooding in Trout Streams in Southeastern Minnesota, USA" Fishes 9, no. 6: 197. https://doi.org/10.3390/fishes9060197
APA StyleMundahl, N. D. (2024). Diets, Condition, and Reproductive Fitness of Slimy Sculpin (Uranidea cognata) after Catastrophic Flooding in Trout Streams in Southeastern Minnesota, USA. Fishes, 9(6), 197. https://doi.org/10.3390/fishes9060197