


Comparison of Bacterial Community Composition in Gut of Chinese Mitten Crabs from Three Distinct Rivers in Korea

 , ,
, , 
 and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Species Identification
2.3. DNA Extraction and Bacterial 16S rDNA Gene Sequencing of CMC Gut
2.4. Bioinformatic Analyses
2.5. Determination of Free Amino Acid Content
2.6. Determination of Free Nucleotide Content
2.7. Determination of Lactic Acid Content
3. Results
3.1. Differences in the Environmental Parameters
3.2. A 16S rDNA Metabarcoding Analysis of the CMC Gut Microbiome
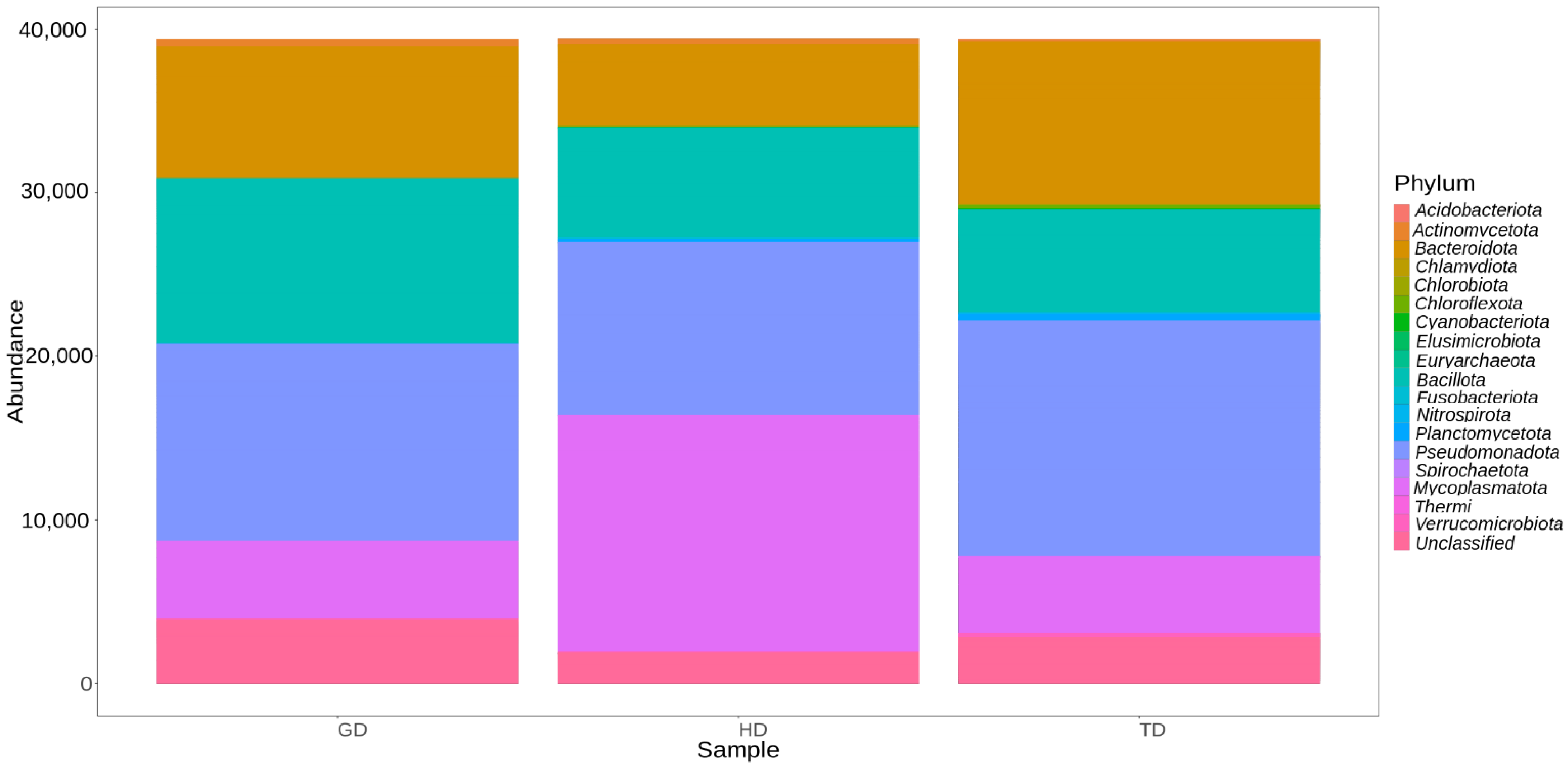
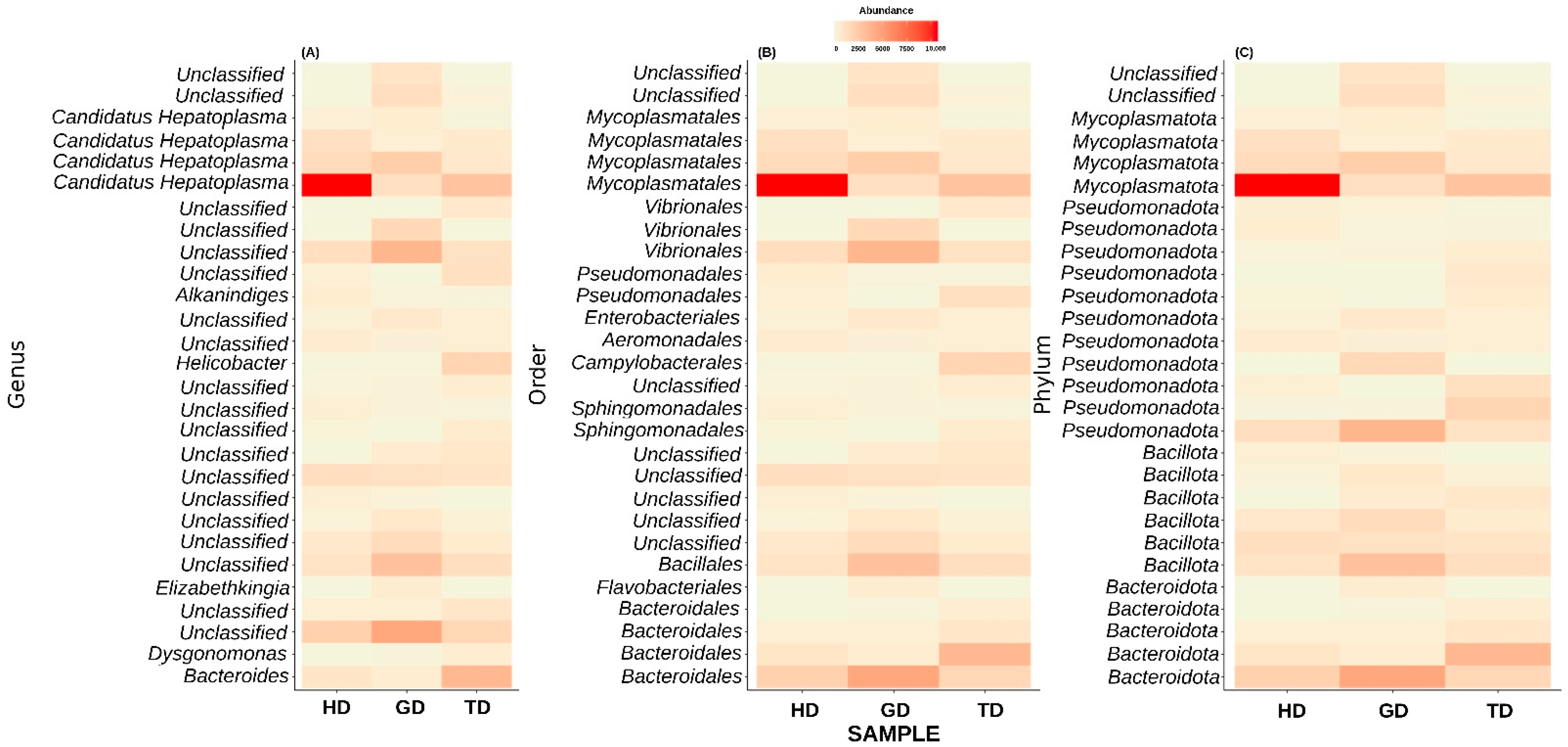
3.3. Microbial Community Composition and Structure
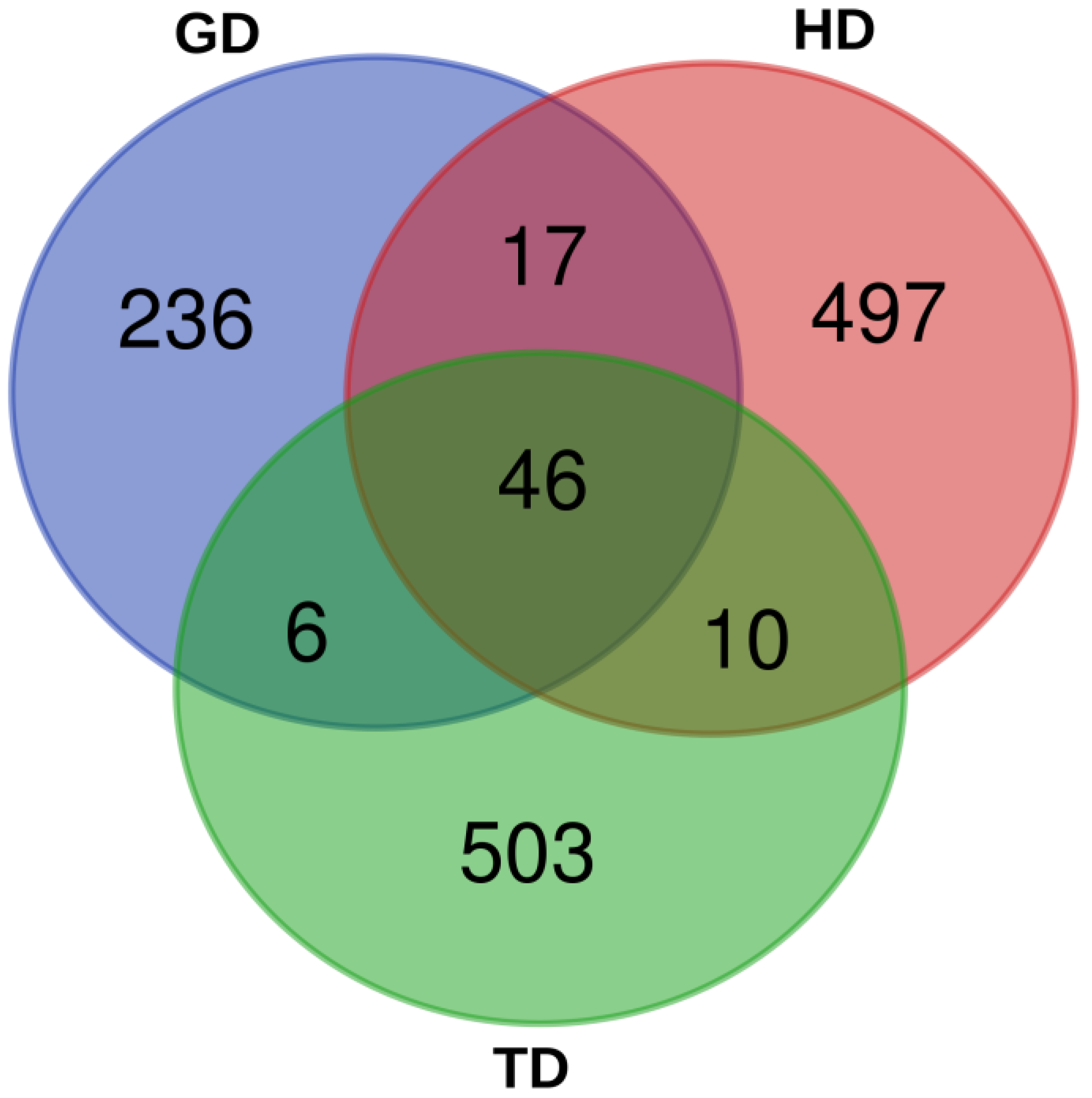
3.4. Microbial Community Similarity among Three Sampling Sites
3.5. Comparison of Free Amino Acids, Nucleotides, and Lactic Acid Content
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shen, H.; Hu, Y.; Zhou, X. Sex-lethal gene of the Chinese mitten crab Eriocheir sinensis: cDNA cloning, induction by eyestalk ablation, and expression of two splice variants in males and females. Dev. Genes Evol. 2014, 224, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Qi, T.; Liu, J.; Liu, Q.; Jiang, S.; Zhang, H.; Wang, Z.; Ding, G.; Tang, B. Adaptively differential expression analysis in gill of Chinese mitten crabs (Eriocheir japonica sinensis) associated with salinity changes. Int. J. Biol. Macromol. 2018, 120, 2242–2246. [Google Scholar] [CrossRef] [PubMed]
- Anger, K. Effects of temperature and salinity on the larval development of the Chinese mitten crab Eriocheir sinensis (Decapoda: Grapsidae). Mar. Ecol. Prog. Ser. 1991, 72, 103–110. [Google Scholar] [CrossRef]
- Jeong, H.; Jeon, D. An Integrated Ecological-Economic System Dynamics Model Analysis on the Ecosystem Restoration Policy (II): Extensions and Relaxations of the Model of King Crabs in the Imjin River, Korea. Korean Syst. Dyn. Rev. 2006, 7, 97–120. [Google Scholar]
- Wang, H.Z.; Wang, H.J.; Liang, X.M.; Cui, Y.D. Stocking models of Chinese mitten crab (Eriocheir japonica sinensis) in Yangtze lakes. Aquaculture 2006, 255, 456–465. [Google Scholar] [CrossRef]
- Qiu, J.; Luo, C.; Ren, L.; Li, W.; Dai, T.; Wang, G.; Sun, X.; Moua, K.C.; Sima, Y.; Xu, S. Black soldier fly larvae replace traditional iced trash fish diet to enhance the delicious flavor of Chinese mitten crab (Eriocheir sinensis). Front. Mar. Sci. 2023, 9, 1089421. [Google Scholar] [CrossRef]
- Jiang, H.; Bao, J.; Xing, Y.; Cao, G.; Li, X.; Chen, Q. Metabolomic and metagenomic analyses of the Chinese mitten crab Eriocheir sinensis after challenge with Metschnikowia bicuspidata. Front. Microbiol. 2022, 13, 990737. [Google Scholar] [CrossRef] [PubMed]
- Yoo, B.; Myung, N. Eco-Friendly Chinese Mitten Crab Production Process Using Microbiota; Ministry of SMEs and Startups: Sejong-si, Republic of Korea, 2012.
- FIRA Seed Release Management System. Available online: https://seed.fira.or.kr/index.jsp (accessed on 19 December 2023).
- Tao, H.; Du, B.; Wang, H.; Dong, H.; Yu, D.; Ren, L.; Sima, Y.; Xu, S. Intestinal microbiome affects the distinctive flavor of Chinese mitten crabs in commercial farms. Aquaculture 2018, 483, 38–45. [Google Scholar] [CrossRef]
- An, H.; Choi, T.; Kim, C. Comparative Transcriptome Analysis of Eriocheir sinensis from Wild Habitats in Han River, Korea. Life 2022, 12, 2027. [Google Scholar] [CrossRef] [PubMed]
- Long, X.; Wu, X.; Zhao, L.; Ye, H.; Cheng, Y.; Zeng, C. Effects of salinity on gonadal development, osmoregulation and metabolism of adult male Chinese mitten crab, Eriocheir sinensis. PLoS ONE 2017, 12, e0179036. [Google Scholar] [CrossRef]
- Yuan, Q.; Wang, Q.; Zhang, T.; Li, Z.; Liu, J. Effects of water temperature on growth, feeding and molting of juvenile Chinese mitten crab Eriocheir sinensis. Aquaculture 2017, 468, 169–174. [Google Scholar] [CrossRef]
- Hooper, L.V.; Midtvedt, T.; Gordon, J.I. How host-microbial interactions shape the nutrient environment of the mammalian intestine. Annu. Rev. Nutr. 2002, 22, 283–307. [Google Scholar] [CrossRef] [PubMed]
- Hooper, L.V.; Macpherson, A.J. Immune adaptations that maintain homeostasis with the intestinal microbiota. Nat. Rev. Immunol. 2010, 10, 159–169. [Google Scholar] [CrossRef] [PubMed]
- Crump, B.C.; Hopkinson, C.S.; Sogin, M.L.; Hobbie, J.E. Microbial biogeography along an estuarine salinity gradient: Combined influences of bacterial growth and residence time. Appl. Environ. Microbiol. 2004, 70, 1494–1505. [Google Scholar] [CrossRef]
- Herlemann, D.P.R.; Labrenz, M.; Jürgens, K.; Bertilsson, S.; Waniek, J.J.; Andersson, A.F. Transitions in bacterial communities along the 2000 km salinity gradient of the Baltic Sea. ISME J. 2011, 5, 1571–1579. [Google Scholar] [CrossRef]
- Zhang, M.; Sun, Y.; Liu, Y.; Qiao, F.; Chen, L.; Liu, W.-T.; Du, Z.; Li, E. Response of gut microbiota to salinity change in two euryhaline aquatic animals with reverse salinity preference. Aquaculture 2016, 454, 72–80. [Google Scholar] [CrossRef]
- Kivistik, C.; Knobloch, J.; Käiro, K.; Tammert, H.; Kisand, V.; Hildebrandt, J.-P.; Herlemann, D.P.R. Impact of salinity on the gastrointestinal bacterial community of Theodoxus fluviatilis. Front. Microbiol. 2020, 11, 683. [Google Scholar] [CrossRef] [PubMed]
- Rooks, M.G.; Garrett, W.S. Gut microbiota, metabolites and host immunity. Nat. Rev. Immunol. 2016, 16, 341–352. [Google Scholar] [CrossRef]
- Garcia-Garcia, E.; Galindo-Villegas, J.; Mulero, V. Mucosal immunity in the gut: The non-vertebrate perspective. Dev. Comp. Immunol. 2013, 40, 278–288. [Google Scholar] [CrossRef]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, A.R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Kumar, P.S.; Brooker, M.R.; Dowd, S.E.; Camerlengo, T. Target region selection is a critical determinant of community fingerprints generated by 16S pyrosequencing. PLoS ONE 2011, 6, e20956. [Google Scholar] [CrossRef] [PubMed]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- McMurdie, P.J.; Holmes, S. phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [PubMed]
- Bliek, T.; Frans, V.D.K.; Marc, G. RNA-seq lesson. ScienceParkStudyGroup. Available online: https://github.com/ScienceParkStudyGroup/rnaseq-lesson (accessed on 7 November 2023).
- Qin, K.X.; Ruan, T.S.; Chen, Y.H.; Liang, G.L.; Wang, H.; Mu, C.K.; Wang, C.L. Effects of temporary rearing time under salinity 7 on the non-volatile flavorings and fatty acids of Eriocheir sinensis. J. Food Compos. Anal. 2022, 107, 104366. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, C.; Wang, H.; Ma, H.; Huang, Y.Q.; Lu, J.X.; Li, X.C.; Zhang, X.W. Involvement of a newly identified atypical type II crustin (SpCrus5) in the antibacterial immunity of mud crab Scylla paramamosain. Fish Shellfish Immunol. 2018, 75, 346–356. [Google Scholar] [CrossRef]
- Wu, X.; Chang, G.L.; Cheng, Y.; Zeng, C.; Southgate, P.C.; Lu, J. Effects of dietary phospholipid and highly unsaturated fatty acid on the gonadal development, tissue proximate composition, lipid class and fatty acid composition of precocious Chinese mitten crab, Eriocheir sinensis. Aquac. Nutr. 2010, 16, 25–36. [Google Scholar] [CrossRef]
- Tang, L.; Wang, H.; Wang, C.L.; Mu, C.K.; Wei, H.L.; Yao, H.Z.; Ye, C.Y.; Chen, L.Z.; Shi, C. Temperature potentially induced distinctive flavor of mud crab Scylla paramamosain mediated by gut microbiota. Sci. Rep. 2020, 10, 3720. [Google Scholar] [CrossRef] [PubMed]
- Kinne, O. Salinity: Animals-invertebrates. In Marine Ecology, Environmental Factors, Part 2; John Wiley and Sons: New York, NY, USA, 1971; Volume 1, pp. 821–996. [Google Scholar]
- Wang, H.; Tang, L.; Wei, H.; Mu, C.; Wang, C. “Butter Crab”: An environment-induced phenotypic variation of Scylla paramamosain with special nutrition and flavour. Aquac. Res. 2019, 50, 541–549. [Google Scholar] [CrossRef]
- Malik, A.; Kim, C.-B. Role of transportome in the gills of Chinese mitten crabs in response to salinity change: A meta-analysis of rna-seq datasets. Biology 2021, 10, 39. [Google Scholar] [CrossRef]
- Li, K.; Guan, W.; Wei, G.; Liu, B.; Xu, J.; Zhao, L.; Zhang, Y. Phylogenetic analysis of intestinal bacteria in the Chinese mitten crab (Eriocheir sinensis). J. Appl. Microbiol. 2007, 103, 675–682. [Google Scholar] [CrossRef]
- Chen, X.; Di, P.; Wang, H.; Li, B.; Pan, Y.; Yan, S.; Wang, Y. Bacterial community associated with the intestinal tract of Chinese mitten crab (Eriocheir sinensis) farmed in Lake Tai, China. PLoS ONE 2015, 10, e0123990. [Google Scholar] [CrossRef] [PubMed]
- Givens, C.E.; Burnett, K.G.; Burnett, L.E.; Hollibaugh, J.T. Microbial communities of the carapace, gut, and hemolymph of the Atlantic blue crab, Callinectes sapidus. Mar. Biol. 2013, 160, 2841–2851. [Google Scholar] [CrossRef]
- Rungrassamee, W.; Klanchui, A.; Chaiyapechara, S.; Maibunkaew, S.; Tangphatsornruang, S.; Jiravanichpaisal, P.; Karoonuthaisiri, N. Bacterial population in intestines of the black tiger shrimp (Penaeus monodon) under different growth stages. PLoS ONE 2013, 8, e60802. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Tian, J.; Wang, G.; Li, W.; Zou, H. Characterization of bacterial community in the stomach of yellow catfish (Pelteobagrus fulvidraco). World J. Microbiol. Biotechnol. 2012, 28, 2165–2174. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Stingl, U.; Anton-Erxleben, F.; Geisler, S.; Brune, A.; Zimmer, M. “Candidatus Hepatoplasma crinochetorum,” a new, stalk-forming lineage of Mollicutes colonizing the midgut glands of a terrestrial isopod. Appl. Environ. Microbiol. 2004, 70, 6166–6172. [Google Scholar] [CrossRef] [PubMed]
- Bouchon, D.; Zimmer, M.; Dittmer, J. The terrestrial isopod microbiome: An all-in-one toolbox for animal–microbe interactions of ecological relevance. Front. Microbiol. 2016, 7, 1472. [Google Scholar] [CrossRef] [PubMed]
- Fraune, S.; Zimmer, M. Host-specificity of environmentally transmitted Mycoplasma-like isopod symbionts. Environ. Microbiol. 2008, 10, 2497–2504. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.S.; Zang, Y.N.; Song, K.; Ma, Y.C.; Dai, T.H.; Serwadda, A. A meta-transcriptomics survey reveals changes in the microbiota of the Chinese mitten crab Eriocheir sinensis infected with Hepatopancreatic necrosis disease. Front. Microbiol. 2017, 8, 732. [Google Scholar] [CrossRef] [PubMed]
- Jeon, I.J. Han River Chinese Mitten Crab with Shells Turned to Black...Fisherman “Never Seen Anything Like It”. The JoongAng. Available online: https://www.joongang.co.kr/article/25194822#home (accessed on 23 September 2023).
- Goodwin, C.S.; Armstrong, J.A.; Chilvers, T.; Peters, M.; Collins, M.D.; Sly, L.; McConnell, W.; Harper, W.E. Transfer of Campylobacter pylori and Campylobacter mustelae to Helicobacter gen. nov. as Helicobacter pylori comb. nov. and Helicobacter mustelae comb, nov., respectively. Int. J. Syst. Evol. Microbiol. 1989, 39, 397–405. [Google Scholar] [CrossRef]
- Ryan, K.J.; Ray, C.G. Sherris Medical Microbiology, 4th ed.; McGraw Hill: New York, NY, USA, 2004. [Google Scholar]
- Dunn, B.E.; Cohen, H.; Blaser, M.J. Helicobacter pylori. Clin. Microbiol. Rev. 1997, 10, 720–741. [Google Scholar] [CrossRef]
- Abdel-Moein, K.A.; Saeed, H.; Samir, A. Novel detection of Helicobacter pylori in fish: A possible public health concern. Acta Trop. 2015, 152, 141–144. [Google Scholar] [CrossRef] [PubMed]
- Hua, J.-S.; Zheng, P.-Y.; Ho, B. Species differentiation and identification in the genus of Helicobacter. World J. Gastroenterol. 1999, 5, 7. [Google Scholar] [CrossRef] [PubMed]
- Rust, M.; Schweinitzer, T.; Josenhans, C. Helicobacter flagella, motility and chemotaxis. In Helicobacter pylori: Molecular Genetics and Cellular Biology; Yamaoka, Y., Ed.; Horizon Scientific Press: Norfolk, UK, 2008; pp. 61–86. [Google Scholar]
- Shen, H.; Ma, Y.; Hu, Y. Near-full-length genome sequence of a novel reovirus from the Chinese mitten crab, Eriocheir sinensis. Genome Announc. 2015, 3, 10–1128. [Google Scholar] [CrossRef] [PubMed]
- Spaargaren, D.H.; Haefner, P.A., Jr. Interactions of ovary and hepatopancreas during the reproductive cycle of Crangon crangon (L.). II. Biochemical relationships. J. Crustac. Biol. 1994, 14, 6–19. [Google Scholar] [CrossRef]
- Boucard, C.G.V.; Levy, P.; Ceccaldi, H.J.; Brogren, C.H. Developmental changes in concentrations of vitellin, vitellogenin, and lipids in hemolymph, hepatopancreas, and ovaries from different ovarian stages of Indian white prawn Fenneropenaeus indicus. J. Exp. Mar. Biol. Ecol. 2002, 281, 63–75. [Google Scholar] [CrossRef]
- Lalitha, K.V.; Sonaji, E.R.; Manju, S.; Jose, L.; Gopal, T.K.S.; Ravisankar, C.N. Microbiological and biochemical changes in pearl spot (Etroplus suratensis Bloch) stored under modified atmospheres. J. Appl. Microbiol. 2005, 99, 1222–1228. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, T.; Asakawa, A.; Yamaguchi, K.; Konosu, S. Studies on flavor components in boiled crabs. III. Sugars, organic acids, and minerals in the extracts. Nippon Suisan Gakkaishi 1979, 45, 1325–1329. [Google Scholar] [CrossRef][Green Version]
- Tao, H.; Liu, H.J.; Cheng, Y.Q.; Sima, Y.H.; Yin, W.M.; Xu, S.Q. Parental environmental exposure leads to glycometabolic disturbances that affect fertilization of eggs in the silkworm Bombyx mori: The parental transcript legacy. J. Comp. Physiol. B 2015, 185, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Wang, X.; Huang, Y.; Bu, X.; Xiao, S.; Qin, C.; Qiao, F.; Qin, J.G.; Chen, L. Effects of dietary T-2 toxin on gut health and gut microbiota composition of the juvenile Chinese mitten crab (Eriocheir sinensis). Fish Shellfish Immunol. 2020, 106, 574–582. [Google Scholar] [CrossRef]
- Du, X.; Zhang, W.; He, J.; Zhao, M.; Wang, J.; Dong, X.; Fu, Y.; Xie, X.; Miao, S. The impact of rearing salinity on flesh texture, taste, and fatty acid composition in largemouth bass Micropterus salmoides. Foods 2022, 11, 3261. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Total Read Bases (bp) | Total Reads | GC (%) | AT(%) | Q20 (%) | Q30 (%) |
|---|---|---|---|---|---|---|
| HD | 595,188,972 | 1,977,372 | 50.33 | 49.67 | 90.52 | 81.68 |
| GD | 495,387,606 | 1,645,806 | 50.47 | 49.53 | 90.76 | 81.81 |
| TD | 535,309,236 | 1,778,436 | 50.83 | 49.17 | 90.45 | 81.15 |
| S. No. | Blast Hit | Accession | % Id | E-Value | Source |
|---|---|---|---|---|---|
| 1 | uncultured bacterium | GU293201.1 | 99.57 | 0 | Pelteobagrus fulvidraco (Yellow catfish) Gut |
| 2 | uncultured bacterium | HG792250.1 | 99.57 | 0 | CMC intestine |
| 3 | uncultured bacterium | HG792243.1 | 99.57 | 0 | CMC intestine |
| 4 | uncultured bacterium | HG792210.1 | 99.57 | 0 | CMC intestine |
| 5 | uncultured bacterium | HG792239.1 | 99.78 | 0 | CMC intestine |
| 6 | uncultured Mycoplasmataceae bacterium | KC918305.1 | 97.63 | 0 | Callinectes sapidus (Blue Crab) gut |
| 7 | uncultured bacterium | HG792233.1 | 99.35 | 0 | CMC intestine |
| 8 | uncultured bacterium | HG792215.1 | 99.57 | 0 | CMC intestine |
| 9 | uncultured bacterium | GU293190.1 | 99.78 | 0 | Pelteobagrus fulvidraco (Yellow catfish) Gut |
| 10 | uncultured bacterium | HG792248.1 | 99.55 | 0 | CMC intestine |
| 11 | Pseudomonas paralcaligenes MRCP1333 | MT604974.1 | 98.49 | 0 | Human feces |
| 12 | uncultured bacterium | DQ856511.1 | 99.35 | 0 | CMC intestine |
| 13 | uncultured bacterium | HG792238.1 | 99.78 | 0 | CMC intestine |
| 14 | uncultured bacterium | HG792243.1 | 99.14 | 0 | CMC intestine |
| 15 | Shewanella sp. | MK757953 | 99.57 | 0 | Lake sediment |
| 16 | uncultured bacterium | HG792201.1 | 99.35 | 0 | CMC intestine |
| 17 | Bacterium 35(2013) strain 35 | KF954020.1 | 99.57 | 0 | CMC intestine |
| 18 | Mollicutes bacterium A0039 | OR648474.1 | 99.57 | 0 | NA |
| 19 | uncultured bacterium | HG792211.1 | 99.55 | 0 | CMC intestine |
| 20 | uncultured bacterium | MW548812.1 | 99.78 | 0 | Phosphogypsum resuspension experiences |
| 21 | uncultured Mycoplasmataceae bacterium | KC918305.1 | 96.55 | 0 | Callinectes sapidus (Blue Crab) gut |
| 22 | uncultured bacterium | DQ856548.1 | 99.54 | 0 | CMC intestine |
| 23 | uncultured bacterium | HG792201.1 | 99.14 | 0 | CMC intestine |
| 24 | Acinetobacter soli S-X6A | KJ806412 | 99.14 | 0 | Jiulong River |
| 25 | uncultured bacterium | DQ856517.1 | 97.83 | 0 | CMC intestine |
| 27 | uncultured bacterium | HG792211.1 | 99.32 | 0 | CMC intestine |
| 28 | uncultured bacterium | JX937698.1 | 98.27 | 0 | Penaeus monodon (Black Tiger Shrimp) Gut |
| 32 | bacterium TSA331-4 | HG792169.1 | 99.35 | 0 | CMC intestine |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
An, H.-E.; Malik, A.; Lee, J.; Mun, M.-H.; Lee, K.H.; Yoo, H.Y.; Kim, C.-B. Comparison of Bacterial Community Composition in Gut of Chinese Mitten Crabs from Three Distinct Rivers in Korea. Fishes 2024, 9, 144. https://doi.org/10.3390/fishes9040144
An H-E, Malik A, Lee J, Mun M-H, Lee KH, Yoo HY, Kim C-B. Comparison of Bacterial Community Composition in Gut of Chinese Mitten Crabs from Three Distinct Rivers in Korea. Fishes. 2024; 9(4):144. https://doi.org/10.3390/fishes9040144
Chicago/Turabian StyleAn, Hyung-Eun, Adeel Malik, Jeongho Lee, Min-Ho Mun, Kang Hyun Lee, Hah Young Yoo, and Chang-Bae Kim. 2024. "Comparison of Bacterial Community Composition in Gut of Chinese Mitten Crabs from Three Distinct Rivers in Korea" Fishes 9, no. 4: 144. https://doi.org/10.3390/fishes9040144
APA StyleAn, H.-E., Malik, A., Lee, J., Mun, M.-H., Lee, K. H., Yoo, H. Y., & Kim, C.-B. (2024). Comparison of Bacterial Community Composition in Gut of Chinese Mitten Crabs from Three Distinct Rivers in Korea. Fishes, 9(4), 144. https://doi.org/10.3390/fishes9040144