
Health Insights from Nematode Larval Characterization in Greater Lizardfish, Saurida tumbil (Bloch, 1795) (Teleostei, Synodontidae)

 , and
, and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
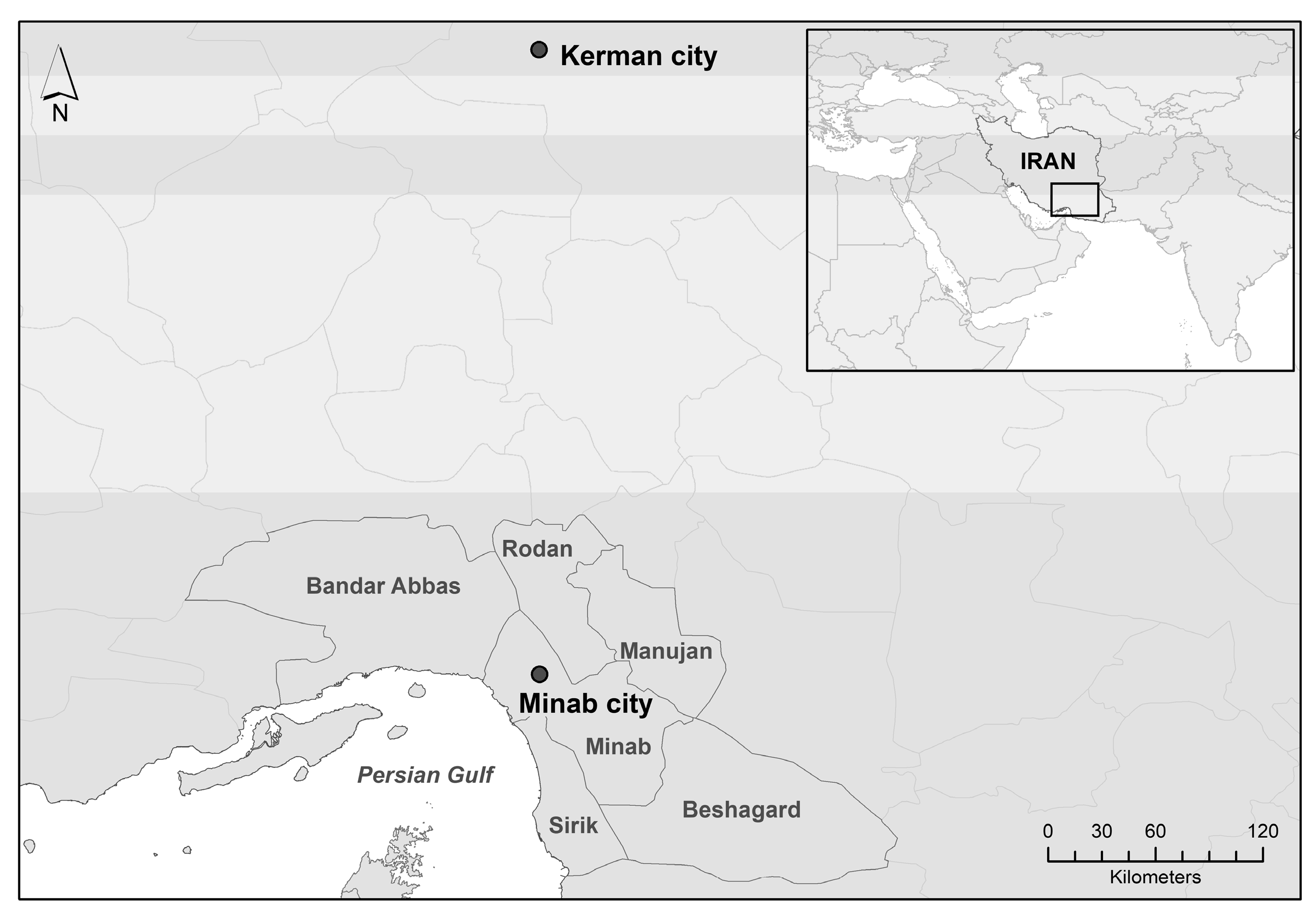
2. Materials and Methods
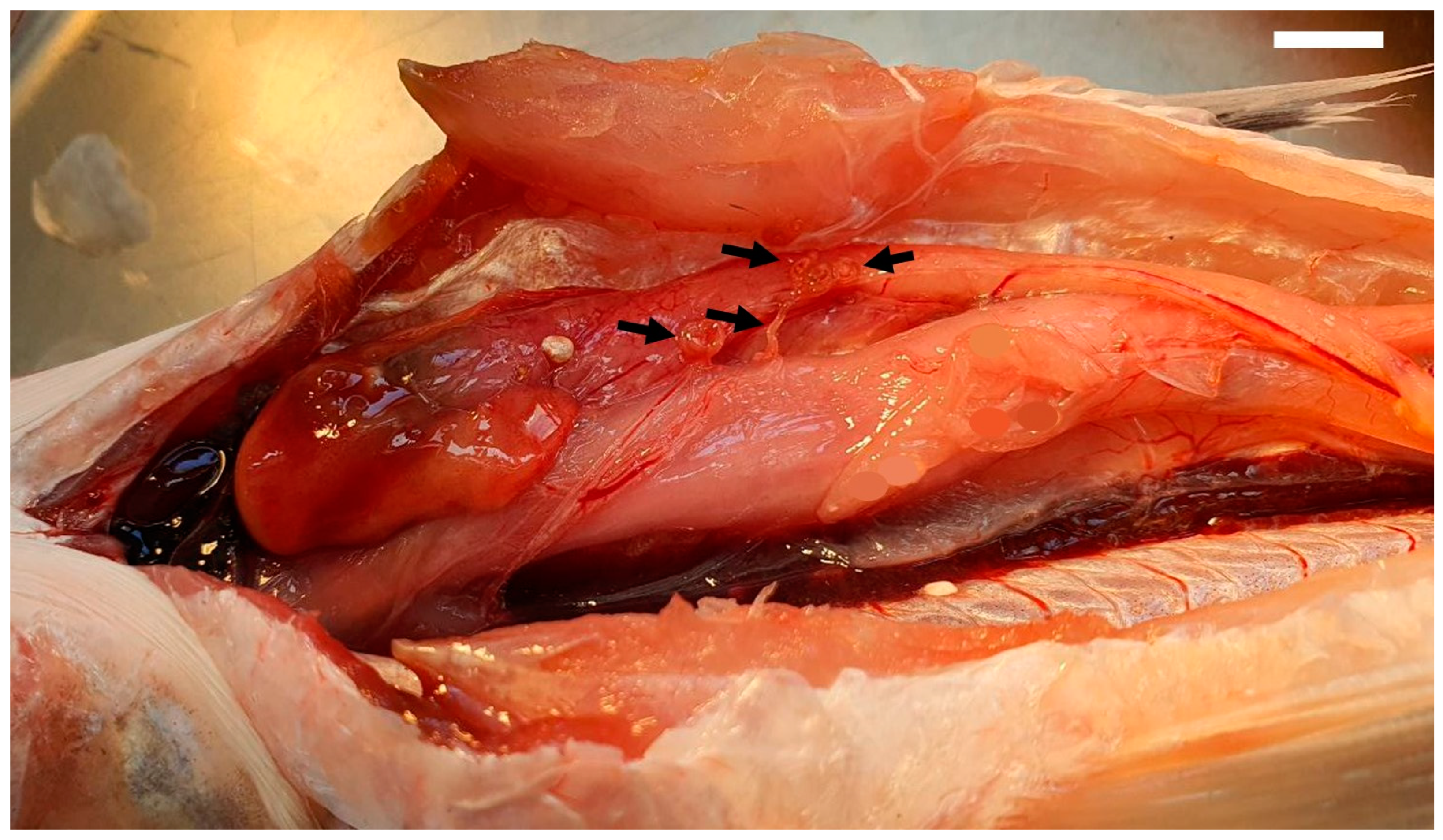
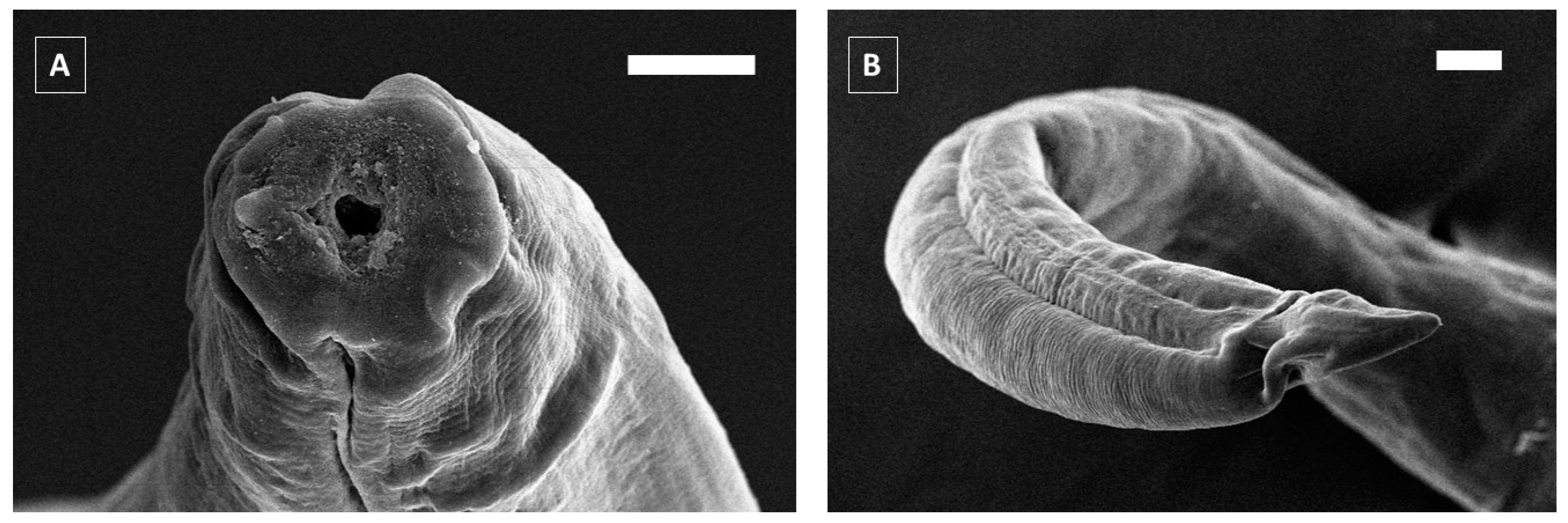
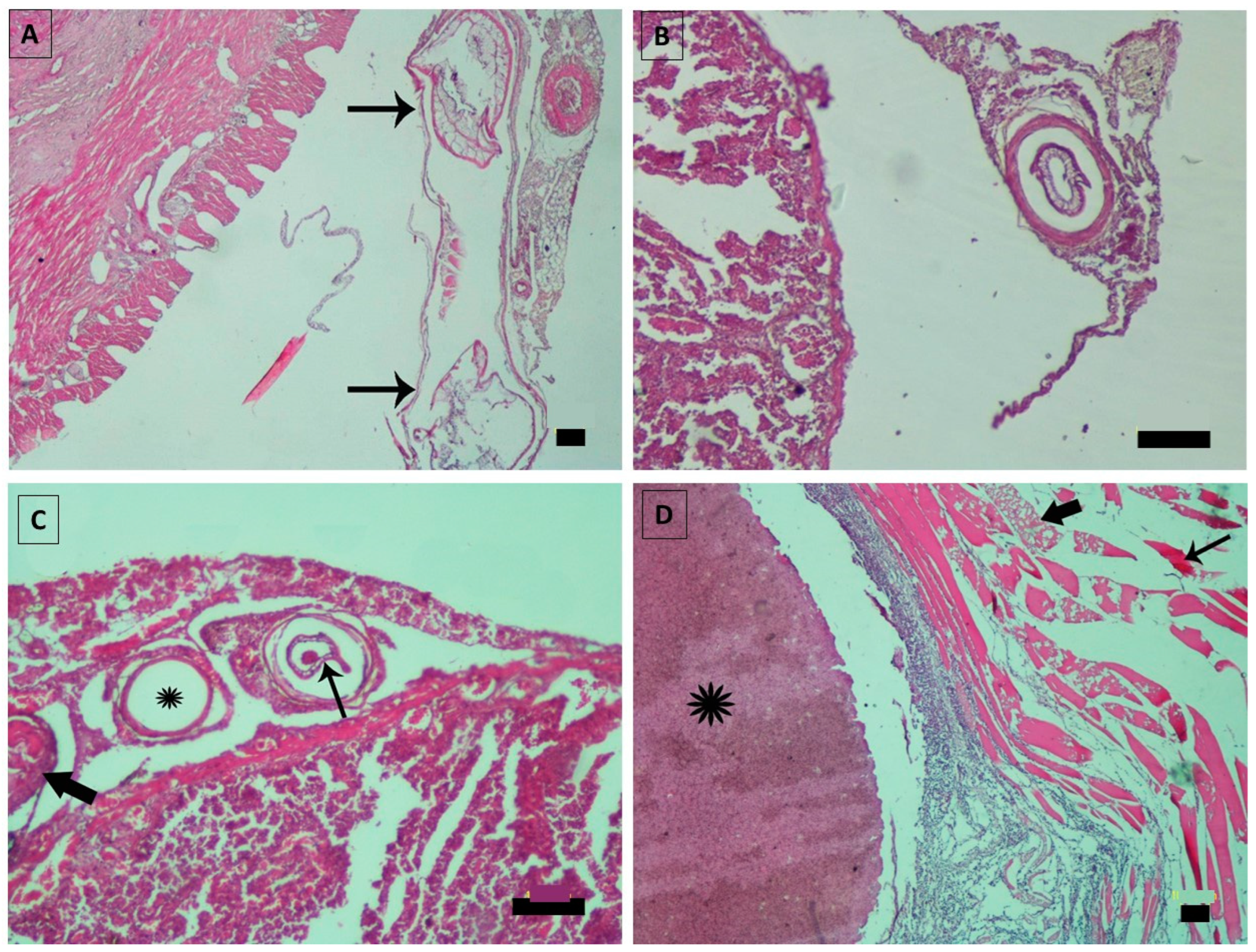
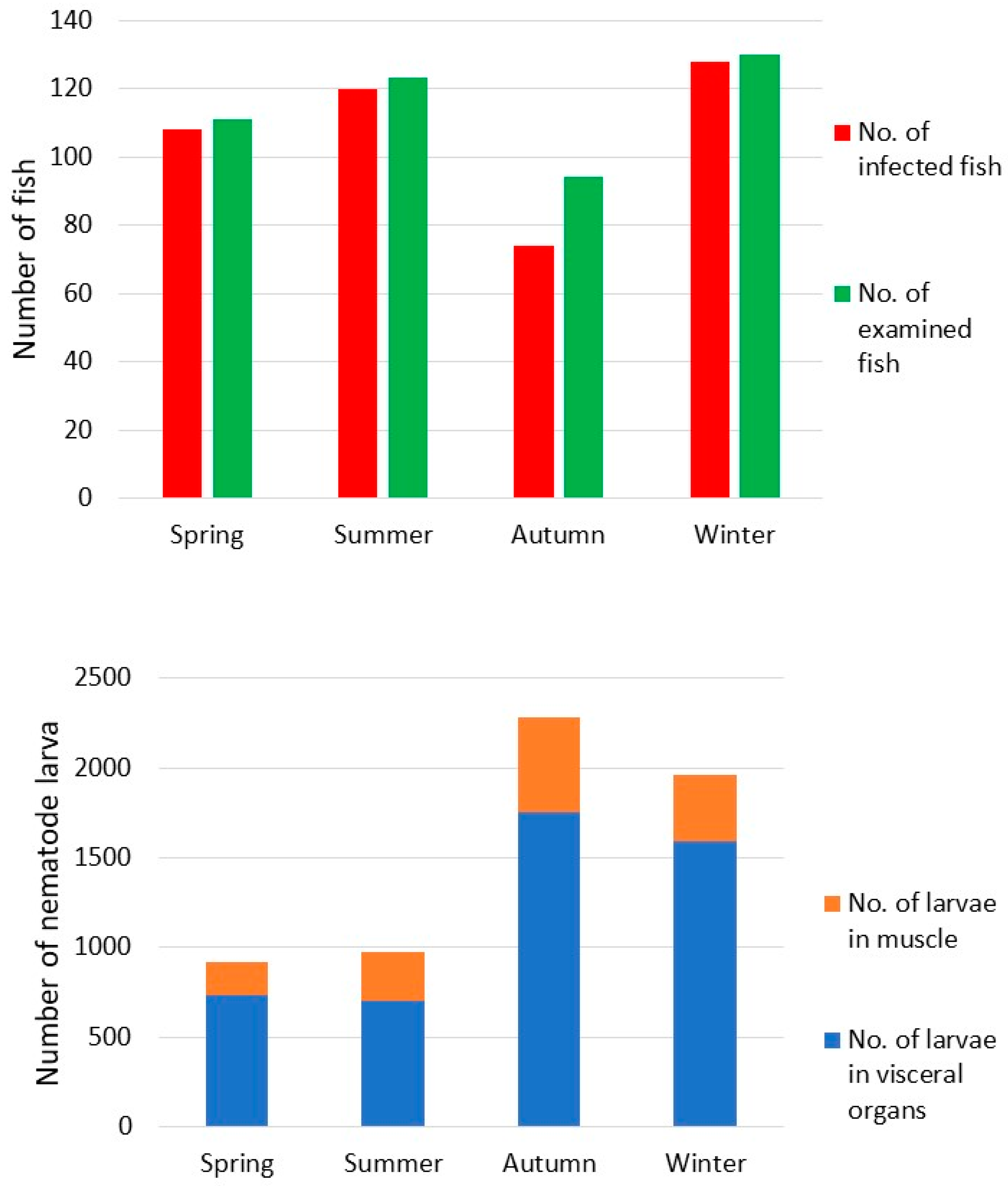
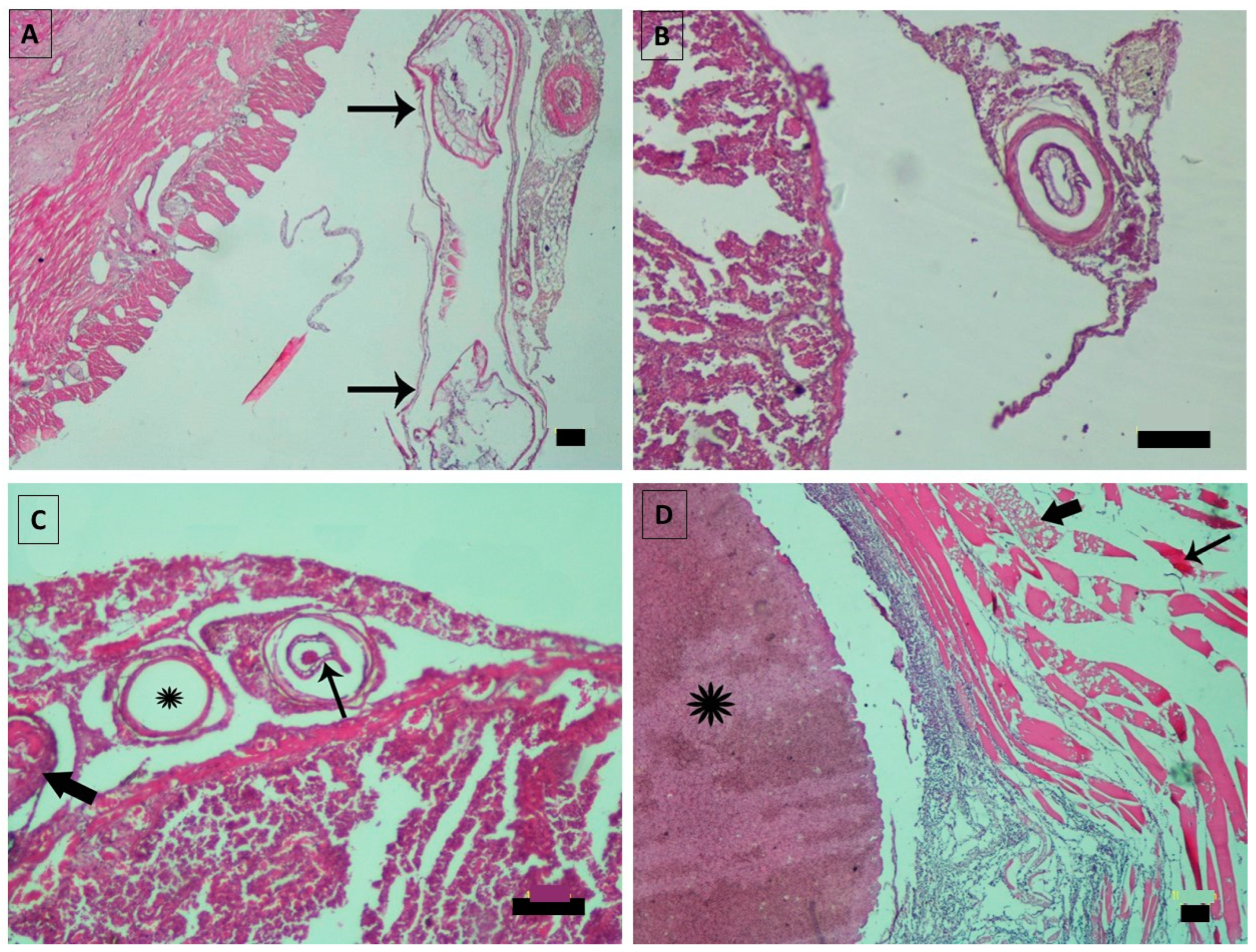
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brunner, E.J.; Jones, P.J.; Friel, S.; Bartley, M. Fish, human health and marine ecosystem health: Policies in collision. Int. J. Epidemiol. 2009, 38, 93–100. [Google Scholar] [CrossRef]
- Pawde, E.D.; Thaware, V.H.; Paul, K. Conservation strategies for fish biodiversity to maintain healthy ecosystem. Int. J. Fish. Aquat. Stud. 2023, 11, 147–149. [Google Scholar] [CrossRef]
- Ortuño Crespo, G.; Dunn, D.C. A review of the impacts of fisheries on open-ocean ecosystems. ICES J. Mar. Sci. 2017, 74, 2283–2297. [Google Scholar] [CrossRef]
- Sommer, U.; Charalampous, E.; Scotti, M.; Moustaka-Gouni, M. Big fish eat small fish: Implications for food chain length? Community Ecol. 2018, 19, 107–115. [Google Scholar] [CrossRef]
- Holmlund, C.M.; Hammer, M. Ecosystem services generated by fish populations. Ecol. Econ. 1999, 29, 253–268. [Google Scholar] [CrossRef]
- Radinger, J.; Matern, S.; Klefoth, T.; Wolter, C.; Feldhege, F.; Monk, C.T.; Arlinghaus, R. Ecosystem-based management outperforms species-focused stocking for enhancing fish populations. Science 2023, 379, 946–951. [Google Scholar] [CrossRef]
- Arthington, A.H.; Dulvy, N.K.; Gladstone, W.; Winfield, I.J. Fish conservation in freshwater and marine realms: Status, threats and management. Aquat. Conserv. Mar. Freshw. Ecosyst. 2016, 26, 838–857. [Google Scholar] [CrossRef]
- Riley, S.C.; Munkittrick, K.R.; Evans, A.N.; Krueger, C.C. Understanding the ecology of disease in Great Lakes fish populations. Aquat. Ecosyst. Health Manag. 2008, 11, 321–334. [Google Scholar] [CrossRef]
- FAO. Fishery and Aquaculture Country Profiles. Iran, 2015. Country Profile Fact Sheets. Fisheries and Aquaculture Division [Online]. Rome. 2023. Updated 1 April 2016. Available online: https://www.fao.org/fishery/en/facp/irn (accessed on 15 December 2023).
- Béné, C.; Arthur, R.; Norbury, H.; Allison, E.H.; Beveridge, M.; Bush, S.; Campling, L.; Leschen, W.; Little, D.; Squires, D. Contribution of fisheries and aquaculture to food security and poverty reduction: Assessing the current evidence. World Dev. 2016, 79, 177–196. [Google Scholar] [CrossRef]
- Rabo, P.; Zarmai, D.; Jwanya, B.; Dikwahal, S. The role of fisheries resources in national development: A review. Int. Lett. Nat. Sci. 2014, 18, 20–28. [Google Scholar] [CrossRef]
- Hamerlynck, O.; Nyingi, W.D.; Paul, J.-L.; Duvail, S. The fish-based farming system: Maintaining ecosystem health and flexible livelihood portfolios. In Farming Systems and Food Security in Africa; Routledge: London, UK, 2019; pp. 354–392. [Google Scholar]
- Funge-Smith, S.; Bennett, A. A fresh look at inland fisheries and their role in food security and livelihoods. Fish Fish. 2019, 20, 1176–1195. [Google Scholar] [CrossRef]
- Sumaila, U.R.; Bellmann, C.; Tipping, A. Fishing for the future: An overview of challenges and opportunities. Mar. Policy 2016, 69, 173–180. [Google Scholar] [CrossRef]
- Johnson, P.T.; Paull, S.H. The ecology and emergence of diseases in fresh waters. Freshw. Biol. 2011, 56, 638–657. [Google Scholar] [CrossRef]
- Preston, D.L.; Mischler, J.A.; Townsend, A.R.; Johnson, P.T. Disease ecology meets ecosystem science. Ecosystems 2016, 19, 737–748. [Google Scholar] [CrossRef]
- Saud, M.; Jawad, L.A.; Park, J.M.; Al Sariri, T.S.; Al Balushi, B.Y. Fish diversity of mangrove ecosystems in Sultanate of Oman. Cah. Biol. Mar 2021, 62, 235–249. [Google Scholar]
- Asadi, H.; Dehghani, R. Atlas of Marine Fishes Persian Gulf and Oman Sea; Iranian Fisheries Research and Training Organization: Tehran, Iran, 1997; p. 226. ISBN 9789645513151. [Google Scholar]
- Ben-Hasan, A.; Daliri, M. Persian Gulf artisanal fisheries: Magnitude, threats, and opportunities. Rev. Fish Biol. Fish. 2023, 33, 541–559. [Google Scholar] [CrossRef]
- Al-Abdulrazzak, D.; Zeller, D.; Belhabib, D.; Tesfamichael, D.; Pauly, D. Total marine fisheries catches in the Persian Gulf from 1950 to 2010. Reg. Stud. Mar. Sci. 2015, 2, 28–34. [Google Scholar] [CrossRef]
- Eagderi, S.; Fricke, R.; Esmaeili, H.R.; Jalili, P. Annotated checklist of the fishes of the Persian Gulf: Diversity and conservation status. Iran. J. Ichthyol. 2019, 6, 1–171. [Google Scholar]
- Froese, R.; Pauly, D. FishBase. World Wide Web Electronic Publication, Version (10/2023). Available online: www.fishbase.org (accessed on 12 December 2023).
- Doustdar, M.; Hashemi, S.; Rahmati, R. The feeding and reproductive habits of Saurida tumbil and Rastrelliger kanagurata in the northern Oman Sea. Iran. J. Fish. Sci. 2022, 21, 816–828. [Google Scholar]
- Yang, X.; Deng, Y.; Qin, J.; Luo, K.; Kang, B.; He, X.; Yan, Y. Dietary shifts in the adaptation to changing marine resources: Insights from a decadal study on greater lizardfish (Saurida tumbil) in the Beibu Gulf, South China Sea. Animals 2024, 14, 798. [Google Scholar] [CrossRef]
- SriHari, M.; Silpa, S.; Pavan-Kumar, A.; Bhushan, S.; Nayak, B.B.; Abidi, Z.J. Stock characterization of Greater Lizardfish, Saurida tumbil (Bloch, 1795) along the west coast of India using morphological and molecular markers. Mar. Biol. Res. 2021, 17, 107–119. [Google Scholar] [CrossRef]
- Zellibooriabadi, M.; Gorgin, S.; Fujimori, Y.; Zare, P.; Niri, A.S.; Susanto, A. Estimation of gillnets selectivity for greater lizardfish, Saurida tumbil (Bloch, 1795) in coastal waters of the Oman Sea. Int. J. Aquat. Biol. 2023, 11, 230–241. [Google Scholar]
- Kalhoro, M.A.; Liu, Q.; Memon, K.H.; Waryani, B.; Soomro, S.H. Maximum sustainable yield of Greater lizardfish Saurida tumbil fishery in Pakistan using the CEDA and ASPIC packages. Acta Oceanol. Sin. 2015, 34, 68–73. [Google Scholar] [CrossRef]
- Ahmadi, M.; Akbarzadeh, A. Phenotypic variation of greater lizard fish (Saurida tumbil) in Northern Persian Gulf (Hormozgan waters). J. Mar. Sci. Technol. 2021, 20, 96–109. [Google Scholar]
- Saberi, M.; Jabaleh, A.; Paighambari, S.Y.; Pormozaffar, S. Determination of Growth, Mortality, and Exploitation Parameters for Saurida tumbil in Coastal Waters off Bandar-e-Jask, Hormozgan Province. Sci. Res. J. Anim. Environ. 2020, 12, 435–442. [Google Scholar] [CrossRef]
- Kalhoro, M.A.; Liu Qun, L.Q.; Tooraj Valinassab, T.V.; Baradi Waryani, B.W.; Abbasi, A.R.; Memon, K.H. Population dynamics of greater lizardfish, Saurida tumbil from Pakistani waters. Pak. J. Zool. 2015, 47, 921–931. [Google Scholar]
- Bahram, S.; Khezri, M.; Javadian, S.R. Evaluation of antioxidant and antimicrobial properties of hydrolyzed protein of Saurida tumbil. Exp. Anim. Biol. 2020, 9, 23–35. [Google Scholar]
- Jaziri, A.A.; Shapawi, R.; Mokhtar, R.A.M.; Noordin, W.N.M.; Huda, N. Microstructural and physicochemical analysis of collagens from the skin of lizardfish (Saurida tumbil Bloch, 1795) extracted with different organic acids. Molecules 2022, 27, 2452. [Google Scholar] [CrossRef]
- Vivekanandan, E. Predatory diversity of two demersal finfish species in the trawling grounds off Veraval. Indian J. Fish. 2001, 48, 133–143. [Google Scholar]
- Abd El-Ghany, A.M.; Nada, M.S.; Nadler, S.A. Morphological and molecular characterization of larval Echinocephalus sp. (Spirurida: Gnathostomatidae), a parasite of the greater lizard fish (Saurida undosquamis) and red porgy or common seabream (Pagrus pagrus). Parasitol. Res. 2023, 122, 2405–2411. [Google Scholar] [CrossRef]
- Morsy, K.; Bashtar, A.-R.; Mostafa, N.; El Deeb, S.; Thabet, S. New host records of three juvenile nematodes in Egypt: Anisakis sp. (Type II), Hysterothylacium patagonense (Anisakidae), and Echinocephalus overstreeti (Gnathostomatidae) from the greater lizard fish Saurida undosquamis of the Red Sea. Parasitol. Res. 2015, 114, 1119–1128. [Google Scholar] [CrossRef] [PubMed]
- FAO Union Nations. FAO Species Fact Sheets: Saurida Tumbil (Bloch, 1795). FAO Fisheries and Aquaculture Department. Available online: https://www.fao.org/figis/pdf/fishery/species/2984/en%3Ftitle%3DFAO%2520Fisheries%2520%2526amp%253B%2520Aquaculture%2520-%2520Aquatic%2520speciessighted04/04/2024 (accessed on 12 December 2023).
- Fernando, C.H.; Furtado, J.I.; Gussev, A.V.; Hanek, G.; Kakonge, S.A. Methods for the Study of Freshwater Fish Parasites; University of Waterloo Publishing: Waterloo, ON, Canada, 1972; Volume 12, p. 76. [Google Scholar]
- Bron, J.E.; Wiegertjes, G.; Piazzon, M.C.; Bobadilla, A. Fish Parasites: A Handbook of Protocols for Their Isolation, Culture and Transmission; 5m Books Ltd.: Great Easton, UK, 2021. [Google Scholar]
- Shamsi, S.; Poupa, A.; Justine, J.L. Characterisation of Ascaridoid larvae from marine fish off New Caledonia, with description of new Hysterothylacium larval types XIII and XIV. Parasitol. Int. 2015, 64, 397–404. [Google Scholar] [CrossRef] [PubMed]
- Cannon, L.R.G. Some larval ascaridoids from south-eastern Queensland marine fishes. Int. J. Parasitol. 1977, 7, 233–243. [Google Scholar] [CrossRef]
- Li, L.; Zhao, W.T.; Guo, Y.N.; Zhang, L.P. Nematode parasites infecting in the starry batfish Halieutaea stellata (Vahl) (Lophiiformes: Ogcocephalidae) from the East and South China Sea. J. Fish Dis. 2016, 39, 515–529. [Google Scholar] [CrossRef] [PubMed]
- Berland, B. Identification of larval nematodes from fish. In Nematode Problems in North Atlantic Fish; Moller, H., Ed.; International Council for the Exploration of the Sea: Copenhagen, Denmark, 1989; pp. 15–22. [Google Scholar]
- Shamsi, S. Morphometric and molecular descriptions of three new species of Hysterothylacium (Nematoda: Raphidascarididae) from Australian marine fish. J. Helminthol. 2017, 91, 613–624. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D.; Gibson, T.J.; Plewniac, F.; Jeanmougin, F.; Higgins, D.G. The Clustal X windows interface:flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 24, 4876–4882. [Google Scholar] [CrossRef] [PubMed]
- Bush, A.O.; Lafferty, K.D.; Lotz, J.M.; Shostak, A.W. Parasitology meets ecology on its own terms: Margolis et al revisited. J. Parasitol. 1997, 83, 575–583. [Google Scholar] [CrossRef]
- Hartwich, G. Keys to genera of the Ascaridoidea. In CIH Keys to the Nematode Parasites of Vertebrates; Anderson, R.C., Willmott, A.G.C.S., Eds.; Commonwealth Agricultural Bureaux: Oxfordshire, UK, 1974; Volume 2, pp. 1–15. [Google Scholar]
- Shamsi, S. The occurrence of Anisakis spp. in Australian waters: Past, present, and future trends. Parasitol. Res. 2021, 120, 3007–3033. [Google Scholar] [CrossRef] [PubMed]
- Shamsi, S.; Sprohnle-Barrera, C.; Shafaet Hossen, M. Occurrence of Anisakis spp. (Nematoda: Anisakidae) in a pygmy sperm whale Kogia breviceps (Cetacea: Kogiidae) in Australian waters. Dis. Aquat. Org. 2019, 134, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Mattiucci, S.; Nascetti, G. Advances and trends in the molecular systematics of anisakid nematodes, with implications for their evolutionary ecology and host—Parasite co-evolutionary processes. Adv. Parasitol. 2008, 66, 47–148. [Google Scholar]
- Anderson, R.C. Nematode Parasites of Vertebrates: Their Development and Transmission, 2nd ed.; CABI Publishing: Oxfordshire, UK, 2000. [Google Scholar]
- Gregori, M.; Roura, Á.; Abollo, E.; González, Á.F.; Pascual, S. Anisakis simplex complex (Nematoda: Anisakidae) in zooplankton communities from temperate NE Atlantic waters. J. Nat. Hist. 2015, 49, 755–773. [Google Scholar] [CrossRef]
- Marcogliese, D.J. The role of zooplankton in the transmission of helminth-parasites to fish. Rev. Fish Biol. Fish. 1995, 5, 336–371. [Google Scholar] [CrossRef]
- Smith, J.W.; Wootten, R. Anisakis and anisakiasis. In Advances in Parasitology; Academic Press Inc. (London) Ltd.: London, UK, 1978; Volume 16, pp. 93–163. [Google Scholar]
- Abollo, E.; Gestal, C.; Pascual, S. Anisakis infestation in marine fish and cephalopods from Galician waters: An updated perspective. Parasitol. Res. 2001, 87, 492–499. [Google Scholar] [PubMed]
- Klimpel, S.; Kuhn, T.; Münster, J.; Dörge, D.D.; Klapper, R.; Kochmann, J. Parasites of Marine Fish and Cephalopods: A Practical Guide; Springer Nature Switzerland AG: Cham, Switzerland, 2019. [Google Scholar]
- Nagasawa, K.; Moravec, F. Larval anisakid nematodes from four species of squid (Cephalopoda: Teuthoidea) from the central and western North Pacific Ocean. J. Nat. Hist. 2002, 36, 883–891. [Google Scholar] [CrossRef]
- Desowitz, R.S. Human and experimental anisakiasis in the United States. [Hokkaido Igaku Zasshi] Hokkaido J. Med. Sci. 1986, 61, 358–371. [Google Scholar] [PubMed]
- Shamsi, S.; Barton, D.P. A critical review of anisakidosis cases occurring globally. Parasitol. Res. 2023, 122, 1733–1745. [Google Scholar] [CrossRef] [PubMed]
- European Food Safety Authority. Scientific Opinion on risk assessment of parasites in fishery products. EFSA J. 2010, 8, 1543. [Google Scholar]
- Najjari, M.; Sadjjadi, S.M.; Khodadadi, H.; Farzaneh, M.R.; Mattiucci, S. Anisakis spp, DNA detection in paraffin-embedded tissue biopsies recovered from patients with gastritis using real-time PCR in Bushehr, Persian Gulf, Iran. Mol. Biochem. Parasitol. 2022, 251, 111494. [Google Scholar] [CrossRef] [PubMed]
- Kent, A.J.; Pert, C.C.; Briers, R.A.; Diele, K.; Rueckert, S. Increasing intensities of Anisakis simplex third-stage larvae (L3) in Atlantic salmon of coastal waters of Scotland. Parasites Vectors 2020, 13, 62. [Google Scholar] [CrossRef] [PubMed]
- Marino, F.; Lanteri, G.; Passantino, A.; De Stefano, C.; Costa, A.; Gaglio, G.; Macri, F. Experimental susceptibility of gilthead sea bream, Sparus aurata, via challenge with Anisakis pegreffii larvae. Biomed Res. Int. 2013, 2013, 701828. [Google Scholar] [CrossRef]
- Macri, F.; Lanteri, G.; Rapisarda, G.; Costa, A.; Marino, F. Anisakis pegreffii experimental challenge in Dicentrarchus labrax: An endoscopic study. Aquaculture 2012, 338, 297–299. [Google Scholar] [CrossRef]
- Pons-Bordas, C.; Hazenberg, A.; Hernandez-Gonzalez, A.; Pool, R.V.; Covelo, P.; Sanchez-Hermosin, P.; Lopez, A.; Saavedra, C.; Fraija-Fernandez, N.; Fernandez, M.; et al. Recent increase of ulcerative lesions caused by Anisakis spp. in cetaceans from the north-east Atlantic. J. Helminthol. 2020, 94, e127. [Google Scholar] [CrossRef] [PubMed]
- Hrabar, J.; Bocina, I.; Kurilj, A.G.; Duras, M.; Mladineo, I. Gastric lesions in dolphins stranded along the Eastern Adriatic coast. Dis. Aquat. Org. 2017, 125, 125–139. [Google Scholar] [CrossRef]
- De Benedetto, G.; Giannetto, A.; Riolo, K.; Iaria, C.; Brianti, E.; Gaglio, G. Anisakis pegreffii Larvae in Sphyraena viridensis and Description of Granulomatous Lesions. Animals 2021, 11, 3449. [Google Scholar] [CrossRef] [PubMed]
- Deardorff, T.L.; Overstreet, R.M. Review of Hysterothylacium and Iheringascaris (both previously = Thynnascaris) (Nematoda: Anisakidae) from the northern Gulf of Mexico. Proc. Biol. Soc. Wash. 1981, 93, 1035–1079. [Google Scholar]
- Nadler, S.A.; D’Amelio, S.; Dailey, M.D.; Paggi, L.; Siu, S.; Sakanari, J.A. Molecular phylogenetics and diagnosis of Anisakis, Pseudoterranova, and Contracaecum from Northern Pacific marine mammals. J. Parasitol. 2005, 91, 1413–1429. [Google Scholar] [CrossRef] [PubMed]
- Shamsi, S.; Gasser, R.; Beveridge, I. Description and genetic characterisation of Hysterothylacium (Nematoda: Raphidascarididae) larvae parasitic in Australian marine fishes. Parasitol. Int. 2013, 62, 320–328. [Google Scholar] [CrossRef]
- Klimpel, S.; Rückert, S. Life cycle strategy of Hysterothylacium aduncum to become the most abundant anisakid fish nematode in the North Sea. Parasitol. Res. 2005, 97, 141–149. [Google Scholar] [CrossRef]
- Rokicki, J. The possibility of completing the life cycle of Hysterothylacium aduncum (Rudolphi, 1802) and Contracaecum rudolphii (Hartwich, 1964) (Nematoda) at the waters of Vistula Lagoon. Wiad. Parazytol. 2005, 51, 239–241. [Google Scholar] [PubMed]
- Gonzalez, L. The life cycle of Hysterothylacium aduncum (Nematoda: Anisakidae) in Chilean marine farms. Aquaculture 1998, 162, 173–186. [Google Scholar] [CrossRef]
- Roca-Geronès, X.; Montoliu, I.; Godínez-González, C.; Fisa, R.; Shamsi, S. Morphological and genetic characterization of Hysterothylacium Ward & Magath, 1917 (Nematoda: Raphidascarididae) larvae in horse mackerel, blue whiting and anchovy from Spanish Atlantic and Mediterranean waters. J. Fish Dis. 2018, 41, 1463–1475. [Google Scholar] [CrossRef] [PubMed]
- Vermeil, C.; Petter, A.; Morin, O.; Le Bodic, M.; Daniel, C.; Guegan, J.; Kerneis, J. Do the eosinophilic granulomas observed in Brittany represent a form of anisakiasis? The larvae of Thynnascaris aduncum do not produce these granulomas experimentally. Bull. Soc. Pathol. Exot. Ses Fil. 1975, 68, 79–83. [Google Scholar]
- Overstreet, R.M.; Meyer, G.W. Hemorrhagic lesions in stomach of rhesus-monkey caused by a piscine ascaridoid nematode. J. Parasitol. 1981, 67, 226–235. [Google Scholar] [CrossRef] [PubMed]
- Yagi, K.; Nagasawa, K.; Ishikura, H.; Nakagawa, A.; Sato, N.; Kikuchi, K.; Ishikura, H. Female worm Hysterothylacium aduncum excreted from human: A case report. Jpn. J. Parasitol. 1996, 45, 12–23. [Google Scholar]
- Shamsi, S. Recent advances in our knowledge of Australian anisakid nematodes. Int. J. Parasitol. Parasites Wildl. 2014, 3, 178–187. [Google Scholar] [CrossRef] [PubMed]
- Shamsi, S. Seafood-borne parasitic diseases: A “one-health” approach is needed. Fishes 2019, 4, 9. [Google Scholar] [CrossRef]
- Beig, J.; Lane, R.J.; Lane, M.R. Gastric anisakiasis: A rare cause of abdominal pain. Intern. Med. J. 2019, 49, 129. [Google Scholar] [CrossRef] [PubMed]
- Sohn, W.M.; Na, B.K.; Kim, T.H.; Park, T.J. Anisakiasis: Report of 15 gastric cases caused by Anisakis Type I larvae and a brief review of Korean anisakiasis cases. Korean J. Parasitol. 2015, 53, 465–470. [Google Scholar] [CrossRef] [PubMed]
- Woo, P.T.K. Fish Diseases and Disorders. Volume 1: Protozoan and Metazoan Infections; CAB International: Wallingford, UK, 1995. [Google Scholar]
- Wootten, R. The Parasitology of Teleosts. In Fish Pathol; Wiley-Blackwell: Hoboken, NJ, USA, 2012; pp. 292–338. [Google Scholar] [CrossRef]
- Feist, S.W.; Longshaw, M. Histopathology of fish parasite infections—Importance for populations. J. Fish Biol. 2008, 73, 2143–2160. [Google Scholar] [CrossRef]
- Gazzonis, A.L.; Cavallero, S.; Zanzani, S.A.; Olivieri, E.; Malandra, R.; Ranghieri, V.; D’Amelio, S.; Manfredi, M.T. Anisakis sp. and Hysterothylacium sp. larvae in anchovies (Engraulis encrasicolus) and chub mackerel (Scomber colias) in the Mediterranean Sea: Molecular identification and risk factors. Food Control 2017, 80, 366–373. [Google Scholar] [CrossRef]
- De Benedetto, G.; Corti, I.; Malandra, R.; Riolo, K.; Giannetto, A.; Gaglio, G. Unusual Localization of Hysterothylacium Incurvum in Xiphias gladius (Linnaeus 1758) Caught in the Atlantic Ocean. Pathogens 2022, 11, 1315. [Google Scholar] [CrossRef] [PubMed]
- The European Food Safety Authority. EFSA Evaluates Parasites in Fish. 2010. Available online: https://www.efsa.europa.eu/en/press/news/biohaz100414 (accessed on 15 December 2023).
- Roden, C. The New Book of Middle Eastern Food; Knopf: New York, NY, USA, 2008. [Google Scholar]
- Bargain, O. Globalization and cultural spillover in trade: Evidence from the Japanese food culture. Rev. World Econ. 2024, 160, 55–73. [Google Scholar] [CrossRef]
- Tian, C.; Luan, W.; Wang, H. Exotic Food, Food Environment, and Geographical Patterns: Big Data Analytics From Japanese Cuisine in China. Front. Earth Sci. 2022, 10, 944927. [Google Scholar] [CrossRef]
- Savvaidis, I.N.; Al Katheeri, A.; Lim, S.-H.E.; Lai, K.-S.; Abushelaibi, A. Traditional foods, food safety practices, and food culture in the Middle East. In Food Safety in the Middle East; Elsevier: Amsterdam, The Netherlands, 2022; pp. 1–31. [Google Scholar]
- Ahmed, N. Tourism Impact on the Cultural Heritage of Countries in the Middle East. J. Appl. Geogr. Stud. 2023, 3, 41–53. [Google Scholar] [CrossRef]
- Derek, M. Ethnic Cuisine in Urban Space. Gastron. Urban Space Chang. Chall. Geogr. Perspect. 2020, 225–237. [Google Scholar] [CrossRef]
- Shamsi, S.; Ghadam, M.; Suthar, J.; Ebrahimzadeh Mousavi, H.; Soltani, M.; Mirzargar, S. Occurrence of ascaridoid nematodes in selected edible fish from the Persian Gulf and description of Hysterothylacium larval type XV and Hysterothylacium persicum n. sp. (Nematoda: Raphidascarididae). Int. J. Food Microbiol. 2016, 236, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Naderi Beni, A.; Marriner, N.; Sharifi, A.; Azizpour, J.; Kabiri, K.; Djamali, M.; Kirman, A. Climate change: A driver of future conflicts in the Persian Gulf Region? Heliyon 2021, 7, e06288. [Google Scholar] [CrossRef] [PubMed]
- Le Quesne, W. Too Hot to Handle? Adapting to the Impacts of Marine Climate Change in the Gulf. Marine Science Gove.UK. 2022. Available online: https://marinescience.blog.gov.uk/2022/11/15/too-hot-to-handle-adapting-to-the-impacts-of-marine-climate-change-in-the-gulf/ (accessed on 4 April 2024).
- Højgaard, D.P. Impact of temperature, salinity and light on hatching of eggs of Anisakis simplex (Nematoda, Anisakidae), isolated by a new method, and some remarks on survival of larvae. Sarsia 1998, 83, 21–28. [Google Scholar] [CrossRef]
- Habibi, F.; Shamsi, S. Preliminary report of occurrence of Corynosoma spp. (Acanthocephala: Polymorphidae) in Southern Caspian sprat (Clupeonella grimmi). Parasitol. Res. 2018, 17, 3327–3331. [Google Scholar] [CrossRef]
- Zuo, S.; Kania, P.W.; Mehrdana, F.; Marana, M.H.; Buchmann, K. Contracaecum osculatum and other anisakid nematodes in grey seals and cod in the Baltic Sea: Molecular and ecological links. J. Helminthol. 2017, 92, 81–89. [Google Scholar] [CrossRef]
- Zuo, S.; Huwer, B.; Bahlool, Q.; Al-Jubury, A.; Christensen, N.D.; Korbut, R.; Kania, P.; Buchmann, K. Host size-dependent anisakid infection in Baltic cod Gadus morhua associated with differential food preferences. Dis. Aquat. Org. 2016, 120, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Dezfuli, B.S.; Pironi, F.; Shinn, A.P.; Manera, M.; Giari, L. Histopathology and ultrastructure of Platichthys flesus naturally infected with Anisakis simplex sl larvae (Nematoda: Anisakidae). J. Parasitol. 2007, 93, 1416–1423. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ghahvei, Y.; Mirzaei, M.; Azizi, S.; Hashemnia, S.; Shamsi, S. Health Insights from Nematode Larval Characterization in Greater Lizardfish, Saurida tumbil (Bloch, 1795) (Teleostei, Synodontidae). Fishes 2024, 9, 143. https://doi.org/10.3390/fishes9040143
Ghahvei Y, Mirzaei M, Azizi S, Hashemnia S, Shamsi S. Health Insights from Nematode Larval Characterization in Greater Lizardfish, Saurida tumbil (Bloch, 1795) (Teleostei, Synodontidae). Fishes. 2024; 9(4):143. https://doi.org/10.3390/fishes9040143
Chicago/Turabian StyleGhahvei, Younes, Mohammad Mirzaei, Shahrzad Azizi, Shadi Hashemnia, and Shokoofeh Shamsi. 2024. "Health Insights from Nematode Larval Characterization in Greater Lizardfish, Saurida tumbil (Bloch, 1795) (Teleostei, Synodontidae)" Fishes 9, no. 4: 143. https://doi.org/10.3390/fishes9040143
APA StyleGhahvei, Y., Mirzaei, M., Azizi, S., Hashemnia, S., & Shamsi, S. (2024). Health Insights from Nematode Larval Characterization in Greater Lizardfish, Saurida tumbil (Bloch, 1795) (Teleostei, Synodontidae). Fishes, 9(4), 143. https://doi.org/10.3390/fishes9040143