

A Cross-Decadal Change in the Fish and Crustacean Community of Lower Yaquina Bay, Oregon, USA

Abstract
1. Introduction
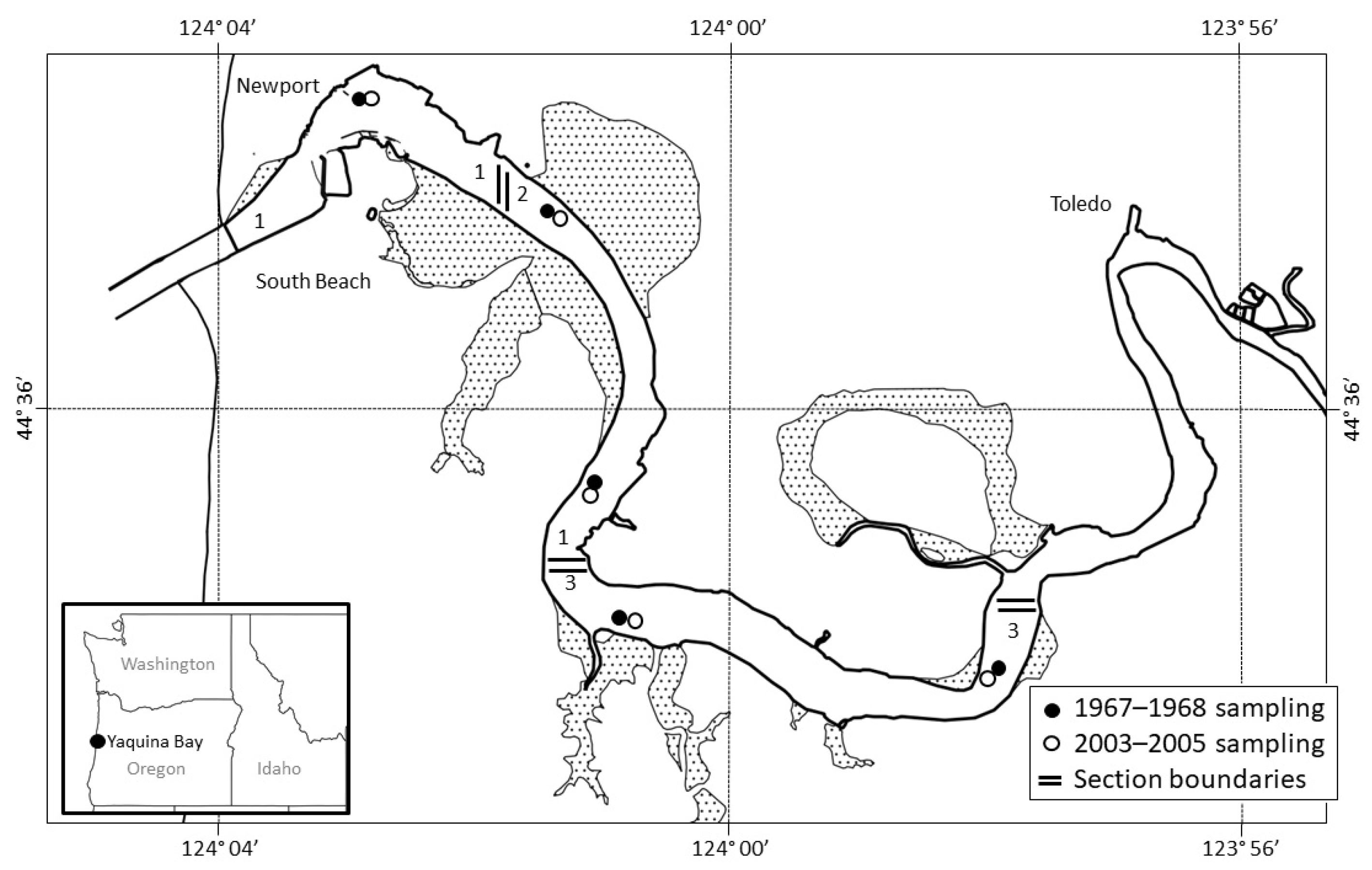
2. Materials and Methods
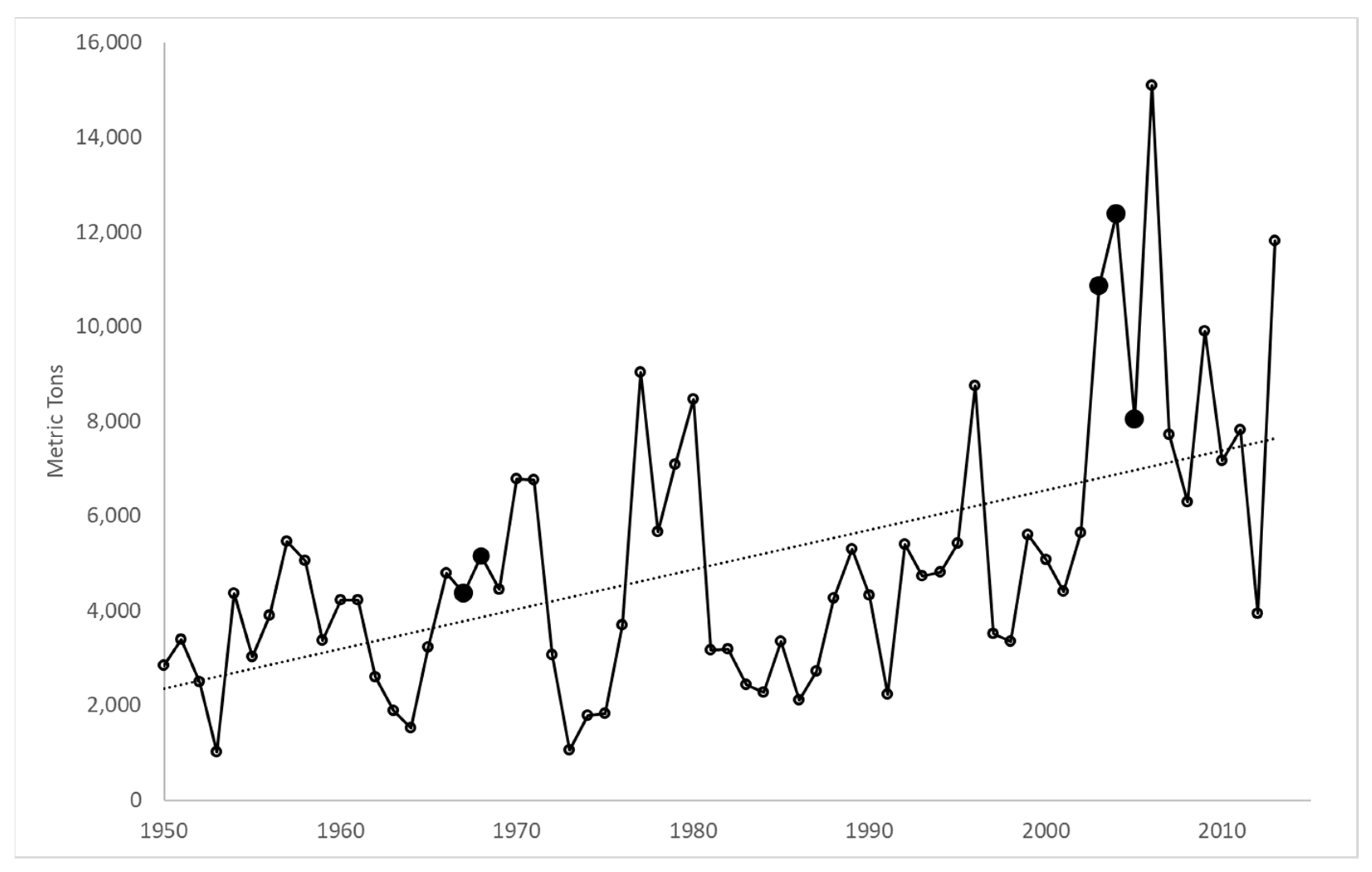
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sobocinski, K.L.; Cordell, J.R.; Simenstad, C.A. Effects of shoreline modifications on supratidal macroinvertebrate fauna on Puget Sound, Washington beaches. Estuaries Coasts 2010, 33, 699–711. [Google Scholar] [CrossRef]
- Long, C.W.; Grow, J.N.; Majoris, J.E.; Hines, A.H. Effects of anthropogenic shoreline hardening and invasion by Phragmites australis on habitat quality for juvenile blue crabs (Callinectes sapidus). J. Exp. Mar. Biol. Ecol. 2011, 409, 215–220. [Google Scholar] [CrossRef]
- Lambert, M.; Ojala-Barbour, R.; Vadas, R., Jr.; McIntyre, A.A.; Quinn, T. Do small overwater structures impact marine habitats and biota? Pac. Con. Biol. 2024, 30, PC22037. Available online: https://www.publish.csiro.au/PC/PC22037. [CrossRef]
- Engle, V.D.; Summers, J.K.; Gaston, G.R. A benthic index of environmental condition of Gulf of Mexico estuaries. Estuaries 1994, 17, 372–384. [Google Scholar] [CrossRef]
- Cao, Y.; Cherr, G.N.; Córdova-Kreylos, A.L.; Fan, T.W.-M.; Green, P.G.; Higashi, R.M.; LaMontagne, M.G.; Scow, K.M.; Vines, C.A.; Yuan, J.; et al. Relationships between sediment microbial communities and pollutants in two California salt marshes. Microb. Ecol. 2006, 52, 619–633. [Google Scholar] [CrossRef]
- Carpenter, S.R.; Caraco, N.F.; Correll, D.L.; Howarth, R.W.; Sharpley, A.N.; Smith, V.H. Non-point pollution of surface waters with phosphorus and nitrogen. Ecol. Appl. 1998, 8, 559–568. [Google Scholar] [CrossRef]
- Rothenberger, M.B.; Swaffield, T.; Calomeni, A.J.; Cabrey, C.D. Multivariate analysis of water quality and plankton assemblages in an urban estuary. Estuaries Coasts 2014, 37, 695–711. [Google Scholar] [CrossRef]
- Van Dolah, R.F.; Calder, D.R.; Knott, D.M. Effects of dredging and open-water disposal on benthic macroinvertebrates in a South Carolina estuary. Estuaries 1984, 7, 28–37. [Google Scholar] [CrossRef]
- Lotze, H.K.; Lenihan, H.S.; Bourque, B.J.; Bradbury, R.H.; Cooke, R.G.; Kay, M.C.; Kidwell, S.M.; Kirby, M.X.; Peterson, C.H.; Jackson, J.B.C. Depletion, degradation, and recovery potential of estuaries and coastal seas. Science 2006, 312, 1806–1809. [Google Scholar] [CrossRef]
- Wolfe, D.A.; Champ, M.A.; Flemer, D.A.; Mearns, A.J. Long-term biological data sets: Their role in research, monitoring, and management of estuarine and coastal marine systems. Estuaries 1987, 10, 181–193. [Google Scholar] [CrossRef]
- DLCD (Oregon Department of Land Conservation and Development). The Oregon Estuary Plan Book. 1987. Available online: www.inforain.org/oregonestuary/ (accessed on 30 June 2015).
- Pearcy, W.G.; Myers, S.S. Larval fishes of Yaquina Bay, Oregon: A nursery ground for marine fishes? Fish Bull. 1974, 72, 201–213. [Google Scholar]
- Emmett, R.L.; Llanso, R.; Newton, J.; Thom, R.; Hornberger, M.; Morgan, C.; Levings, C.; Copping, A.; Fishman, P. Geographic signatures of North American West Coast estuaries. Estuaries 2000, 23, 765–792. [Google Scholar] [CrossRef]
- Krygier, E.E.; Pearcy, W.G. The role of estuarine and offshore nursery areas for young English sole, Paraphrys vetulus Girard, of Oregon. Fish Bull. 1986, 84, 119–132. [Google Scholar]
- Boehlert, G.W.; Mundy, B.C. Recruitment dynamics of metamorphosing English sole, Parophorys vetulus, to Yaquina Bay, Oregon. Estuar. Est. Coast. Shelf Sci. 1987, 25, 261–281. [Google Scholar] [CrossRef]
- Armstrong, D.A.; Rooper, C.; Gunderson, D.R. Estuarine production of juvenile Dungeness crab (Cancer magister) and contribution to the Oregon-Washington coastal fishery. Estuaries 2003, 26, 1174–1189. [Google Scholar] [CrossRef]
- Dauble, A.D.; Heppell, S.A.; Johansson, M.L. Settlement patterns of young-of-the-year rockfish among six Oregon estuaries experiencing different levels of human development. Mar. Ecol. Prog. Ser. 2012, 448, 143–154. [Google Scholar] [CrossRef][Green Version]
- Bottin, R.R.; Briggs, M.J. Newport North Marina, Yaquina Bay, Oregon, Design for Wave Protection: Coastal Model Investigation; United States Army Corps of Engineers Technical Report CERC-96-2; 1996. [Google Scholar]
- Good, J.W. Estuary development with implications for management: A case study of Northwest Natural Gas Company’s liquified natural gas (LNG) project at Yaquina Bay, Oregon. Department of Oceanography, Oregon State University; 1975 Corvallis, OR. Port of Newport. History of the Port of Newport. 2012. Available online: www.portofnewport.com (accessed on 3 July 2012).
- USACE. U.S. Army Corps of Engineers: Yaquina Bay. U.S. Army Corps of Engineers. 2012. Available online: www.nwp.usace.army.mil/locations/yaquinabay.asp (accessed on 3 July 2012).
- Oregon Coast Management Program (OCMP) (2016) Estuary Planning. Available online: https://www.oregon.gov/lcd/OCMP/Pages/Estuary-Planning.aspx (accessed on 28 March 2024).
- Brown, C.A.; Power, J.H. Historic and recent patterns of dissolved oxygen in the Yaquina Estuary (Oregon, USA): Importance of anthropogenic activities and oceanic conditions. Estuar. Coast Shelf Sci. 2011, 92, 446–455. [Google Scholar] [CrossRef]
- De Ben, W.A.; Clothier, W.D.; Ditsworth, G.R.; Baumgartner, D.J. Spatio-temporal fluctuations in the distribution and abundance of demersal fish and epibenthic crustaceans in Yaquina Bay, Oregon. Estuaries 1990, 13, 469–478. [Google Scholar] [CrossRef]
- Frolander, H.F. Biological and chemical features of tidal estuaries. J. Water Pollut. Control Fed. 1964, 36, 1037–1048. [Google Scholar]
- Wares, P.G. Biology of the Pile Perch, Rhacochilus vacca, in Yaquina Bay, Oregon; United States Bureau of Sports Fisheries and Wildlife Technical Paper 57; U.S. Fish and Wildlife Service: Washington, DC, USA, 1971.
- Oregon State University Hatfield Marine Science Center Guin Library. Available online: https://yaquina.library.oregonstate.edu/ (accessed on 28 March 2024).
- Frolander, H.F.; Miller, C.B.; Flynn, M.J. Seasonal cycles of abundance in zooplankton populations of Yaquina Bay Oregon. Mar. Biol. 1973, 21, 277–288. [Google Scholar] [CrossRef]
- Krygier, E.E.; Johnson, W.C.; Bond, C.E. Records of California tonguefish, threadfin shad and smooth alligatorfish from Yaquina Bay, Oregon. Cal. Fish Game. 1973, 59, 140–142. [Google Scholar]
- Richardson, S.L.; Pearcy, W.G. Coastal and oceanic fish larvae in an area of upwelling off Yaquina Bay, Oregon. Fish Bull. 1977, 75, 125–145. [Google Scholar]
- Barton, M. Comparative distribution and habitat preferences of 2 species of stichaeoid fishes in Yaquina Bay, Oregon. J. Exp. Mar. Biol. Ecol. 1982, 59, 77–87. [Google Scholar] [CrossRef]
- Bond, C.E.; Kan, T.T.; Myers, K.W. Notes on the marine life of the river lamprey, Lampetra ayresi in Yaquina Bay, Oregon, and the Columbia River estuary. Fish Bull. 1983, 81, 165–167. [Google Scholar]
- Gallagher, M.B.; Heppell, S.S. Essential habitat identification for age-0 rockfish along the central Oregon coast. Mar. Coastal. Fish 2010, 2, 60–72. [Google Scholar] [CrossRef]
- Simpson, E.H. Measurement of diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Shannon, C.E.; Weaver, W. The Mathematical Theory of Communication; University of Illinois Press: Champaign, IL, USA, 1949. [Google Scholar]
- Margalef, R. Perspectives in Ecological Theory; University of Chicago Press: Chicago, IL, USA, 1968. [Google Scholar]
- Pielou, E.C. An Introduction to Mathematical Ecology; Wiley: New York, NY, USA, 1969. [Google Scholar]
- Pauley, G.B.; Armstrong, D.A.; Van Citter, R.; Thomas, G.L. Species profiles: Life histories and environmental requirements of coastal fishes and invertebrates (Pacific Southwest)—Dungeness Crab. USFWS Biol. Rep. 1989, 82, TR EL-82-4. [Google Scholar]
- Peterson, M.S.; Comyns, B.H.; Hendon, J.R.; Bond, P.J.; Duff, G.A. Habitat use by early life-history stages of fishes and crustaceans along a changing estuarine landscape: Differences between natural and altered shoreline sites. Wetl. Ecol. Manag. 2000, 8, 209–219. [Google Scholar] [CrossRef]
- Bilkovic, D.M.; Roggero, M.M. Effects of coastal development on nearshore estuarine nekton communities. Mar. Ecol. Prog. Ser. 2008, 358, 27–39. [Google Scholar] [CrossRef]
- Kimball, M.E.; Pfirrmann, B.W.; Allen, D.M.; Ogburn-Matthews, V.; Kenny, P.D. Intertidal creek pool nekton assemblages: Long-term patterns in diversity and abundance in a warm-temperate estuary. Estuaries Coasts 2023, 46, 860–877. [Google Scholar] [CrossRef]
- Schwartzkopf, B.D.; Dauble, A.D.; Lindsley, A.J.; Heppell, S.A. Temporal and habitat differences in the juvenile demersal fish community at a marine-dominated northeast Pacific estuary. Fish Res. 2020, 227, 105557. [Google Scholar] [CrossRef]
- NOAA. Fisheries Statistics: Commercial landings, Dungeness crab in Oregon from 1967–2010, NOAA Fisheries OST. 2012. Available online: www.st.nmfs.noaa.gov/st1/commercial/landings/annual_landings.html (accessed on 3 July 2012).
- Sigleo, A.C.; Frick, W.E. Seasonal variations in river discharge and nutrient export to a Northeastern Pacific estuary. Coast. Shelf Sci. 2007, 73, 368–378. [Google Scholar] [CrossRef]
- Brown, C.A.; Ozretich¸, R.J. Coupling between the coastal ocean and Yaquina Bay, Oregon: Importance of oceanic inputs relative to other nitrogen sources. Estuaries Coasts 2009, 32, 219–237. [Google Scholar] [CrossRef]
- Camara, M.D.; Evans, S.; Langdon, C.J. Parental relatedness and survival of Pacific oysters from a naturalized population. J. Shellfish Res. 2008, 27, 323–336. [Google Scholar] [CrossRef]
- D’Andrea, A.F.; DeWitt, T.H. Geochemical ecosystem engineering by the mud shrimp Upogebia pugettensis (Crustacea: Thalassinidae) in Yaquina Bay, Oregon: Density-dependent effects on organic matter remineralization and nutrient cycling. Limnol. Oceanogr. 2009, 54, 1911–1932. [Google Scholar] [CrossRef]
- Repetto, M.; Griffen, B.D. Physiological consequences of parasite infection in the burrowing mud shrimp, Upogebia pugettensis, a widespread ecosystem engineer. Mar. Fresh Res. 2012, 63, 60–67. [Google Scholar] [CrossRef]
- Kaldy, J.E. Production ecology of the non-indigenous seagrass, dwarf eelgrass (Zostera japonica Ascher & Graeb), in a Pacific Northwest estuary, USA. Hydrobiologia 2006, 553, 201–217. [Google Scholar]
- Boese, B.L.; Robbins, B.D. Effects of erosion and macroalgae on intertidal eelgrass (Zostera marina) in a northeastern Pacific estuary (USA). Bot. Mar. 2008, 51, 247–257. [Google Scholar] [CrossRef]
- Hessing-Lewis, M.L.; Hacker, S.D.; Menge, B.A. Context-dependent eelgrass-macroalgae interactions along an estuarine gradient in the Pacific Northwest, USA. Estuaries Coasts 2011, 34, 1169–1181. [Google Scholar] [CrossRef]
- Petrie, M.E.; Ryer, C.H. Laboratory and field evidence for structural habitat affinity of young-of-the-year lingcod. Trans. Am. Fish Soc. 2006, 135, 1622–1630. [Google Scholar] [CrossRef]
- Johnson, L.L.; Ylitalo, G.M.; Arkoosh, M.R. Contaminant exposure in outmigrant juvenile salmon from Pacific Northwest estuaries of the United States. Environ. Monit. Assess 2007, 124, 167–194. [Google Scholar] [CrossRef] [PubMed]
- Schwartzkopf, B.D.; Heppell, S.A. A feeding ecology-based approach to evaluating nursery potential of estuaries for Black Rockfish. Mar. Coast. Fish 2020, 12, 124–141. [Google Scholar] [CrossRef]
- Spencer, M.L.; Stoner, A.W.; Ryer, C.H.; Munk, J.E. A towed camera sled for estimating abundance of juvenile flatfishes and habitat characteristics: Comparison with beam trawls and divers. Est. Coast. Shelf. Sci. 2005, 64, 497–503. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Environmental Data | 1967, 1968 | 2003, 2004, 2005 |
|---|---|---|
| Mean Monthly Salinity Range | 8–28 | 14–36 |
| Maximum Salinity | 34 | 36.4 |
| Summer Monthly Salinity Range | 16–34 | 24–36.4 |
| Annual Rainfall (cm) | 158, 282 | 176, 150, 158 |
| Summer Bottom Temperature Range (°C) | 21–23 | 12–17 |
| Coldest Bottom Temperature (°C) | 6 | 9 |
| Common Name | Scientific Name | 1967–1968 | 2003–2005 | ||||
|---|---|---|---|---|---|---|---|
| Section | Section | ||||||
| 1 | 2 | 3 | 1 | 2 | 3 | ||
| English sole | Parophrys vetulus | 379 | 8349 | 2267 | 76 | 996 | 714 |
| Snake prickleback | Lumpenus sagitta | 37 | 2547 | 445 | 0 | 10 | 13 |
| Shiner surfperch | Cymatogaster aggregata | 105 | 1116 | 1119 | 30 | 66 | 300 |
| Dungeness crab | Cancer magister | 323 | 867 | 298 | 130 | 1780 | 2189 |
| Blacktail bay shrimp | Crangon nigricauda | 155 | 1326 | 3 | 27 | 297 | 265 |
| Pile surfperch | Rhacochilus vacca | 58 | 829 | 394 | 4 | 218 | 139 |
| Buffalo sculpin | Enophrys bison | 1170 | 85 | 1 | 0 | 8 | 6 |
| Starry flounder | Platichthys stellatus | 211 | 692 | 317 | 2 | 72 | 42 |
| White seaperch | Phanerodon furcatus | 53 | 587 | 121 | 11 | 1 | 0 |
| California bay shrimp | Crangon franciscorum | 11 | 87 | 603 | 0 | 0 | 0 |
| Opossum shrimp | Neomysis mercedis | 4 | 42 | 590 | 0 | 0 | 0 |
| Pacific staghorn sculpin | Leptocottus armatus | 30 | 395 | 96 | 0 | 57 | 156 |
| Surf smelt | Hypomesus pretiosus | 4 | 28 | 333 | 16 | 161 | 36 |
| Sand sole | Psettichthys melanosticus | 189 | 114 | 36 | 0 | 0 | 0 |
| Speckled sanddab | Citharichthys stigmaeus | 239 | 96 | 1 | 1 | 252 | 30 |
| Striped seaperch | Embiotoca lateralis | 202 | 120 | 1 | 0 | 0 | 0 |
| Walleye surfperch | Hyperprosopon argenteum | 1 | 53 | 226 | 0 | 0 | 0 |
| Kelp greenling | Hexagrammos decagrammus | 154 | 20 | 1 | 0 | 6 | 1 |
| Saddleback gunnel | Pholis ornata | 2 | 85 | 9 | 1 | 6 | 4 |
| Pacific tomcod | Microgadus proximus | 43 | 40 | 4 | 0 | 57 | 9 |
| Northern anchovy | Engraulis mordax | 0 | 2 | 80 | 0 | 0 | 0 |
| Longfin smelt | Spirinchus thaleichthys | 2 | 1 | 76 | 0 | 0 | 0 |
| Red rock crab | Cancer productus | 41 | 3 | 0 | 1 | 28 | 26 |
| Pacific herring | Clupea pallasi | 0 | 8 | 35 | 0 | 19 | 102 |
| Black rockfish | Sebastes melanops | 22 | 14 | 5 | 2 | 27 | 1 |
| American shad | Alosa sapidissima | 1 | 0 | 36 | 0 | 0 | 27 |
| Bay pipefish | Syngnathus leptorhynchus | 3 | 10 | 21 | 0 | 10 | 3 |
| Lingcod | Ophiodon elongatus | 13 | 9 | 2 | 0 | 5 | 3 |
| California coastal shrimp | Heptacarpus paludicola | 8 | 9 | 0 | 0 | 0 | 0 |
| Tube snout | Aulorhynchus flavidus | 1 | 14 | 0 | 10 | 1 | 0 |
| Cabezon | Scorpaenichthys marmoratus | 11 | 4 | 0 | 0 | 1 | 0 |
| Rock greenling | Hexagrammos lagocephalus | 7 | 1 | 1 | 0 | 0 | 0 |
| Redtail surfperch | Amphistichus rhodoterus | 2 | 6 | 1 | 0 | 0 | 0 |
| C-O sole | Pleuronichthys coenosus | 6 | 0 | 0 | 0 | 0 | 0 |
| Three-spined stickleback | Gasterosteus aculeatus | 0 | 0 | 6 | 0 | 0 | 0 |
| Shortspine shrimp | Heptacarpus brevirostris | 1 | 5 | 0 | 0 | 0 | 0 |
| Whitebait smelt | Allosmerus elongatus | 3 | 1 | 0 | 0 | 0 | 0 |
| Tubenose poacher | Pallasina barbata | 3 | 1 | 0 | 0 | 0 | 0 |
| Big skate | Raja binoculata | 1 | 2 | 0 | 0 | 0 | 0 |
| Penpoint gunnel | Apodichthys flavidus | 3 | 0 | 0 | 0 | 0 | 0 |
| California spot prawn | Pandalus platyceros | 1 | 0 | 1 | 0 | 0 | 0 |
| Bay shrimp | Lissocrangon stylirostris | 2 | 0 | 0 | 0 | 0 | 0 |
| Bay goby | Lepidogobius lepidus | 0 | 2 | 0 | 0 | 0 | 0 |
| Pacific sand lance | Ammodytes hexapterus | 1 | 0 | 1 | 0 | 0 | 0 |
| Longnose skate | Raja rhina | 1 | 1 | 0 | 0 | 0 | 0 |
| Copper rockfish | Sebastes caurinus | 1 | 0 | 0 | 0 | 1 | 1 |
| Arrow goby | Clevlandia ios | 0 | 1 | 0 | 0 | 0 | 0 |
| Scalyhead sculpin | Artedius harringtoni | 1 | 0 | 0 | 0 | 0 | 0 |
| Whitespotted greenling | Hexagrammos stelleri | 1 | 0 | 0 | 0 | 0 | 0 |
| Tidepool snailfish | Liparis florae | 1 | 0 | 0 | 0 | 0 | 0 |
| Green sturgeon | Acipenser medirostris | 0 | 0 | 1 | 0 | 0 | 0 |
| Pacific hake | Merluccius productus | 0 | 1 | 0 | 0 | 0 | 0 |
| Wolf eel | Anarrhichthys ocellatus | 1 | 0 | 0 | 0 | 0 | 0 |
| Brown Irish lord | Hemilepidotus spinosus | 1 | 0 | 0 | 0 | 0 | 0 |
| Silver surfperch | Hyperprosopon elliptiicum | 1 | 0 | 0 | 0 | 0 | 0 |
| Vermilion rockfish | Sebastes miniatus | 1 | 0 | 0 | 0 | 0 | 0 |
| Chinook salmon | Oncorhynchus tshawytscha | 0 | 0 | 1 | 0 | 0 | 0 |
| Coho salmon | Oncorhynchus kisutch | 0 | 0 | 1 | 0 | 0 | 0 |
| Prickly sculpin | Cottus asper | 0 | 0 | 1 | 0 | 0 | 0 |
| Dock shrimp | Pandalus danae | 1 | 0 | 0 | 0 | 0 | 0 |
| Porcelain crab | Petrolisthes cinctipes | 0 | 0 | 0 | 0 | 2 | 12 |
| Hermit crab | Pagurus sp. | 0 | 0 | 0 | 0 | 0 | 2 |
| Kelp crab | Pugettia producta | 0 | 0 | 0 | 0 | 6 | 1 |
| Red Irish lord | Hemilepidotus hemilepidotus | 0 | 0 | 0 | 0 | 2 | 0 |
| Pacific sanddab | Citharichthys sordidus | 0 | 0 | 0 | 0 | 1 | 0 |
| Eulachon | Thaleichthys pacificus | 0 | 0 | 0 | 0 | 1 | 0 |
| Totals | 3512 | 17,573 | 7134 | 311 | 4091 | 4082 | |
| CPUE of Species in Top 95% of Catch | 1967–1968 | 2003–2005 |
|---|---|---|
| All Individuals | 644.64 | 57.99 |
| Epibenthic Crustaceans (all) | 134.6 | 146.5 |
| Epibenthic Crustaceans (minus Dungeness crab) | 102.6 | 33.73 |
| Demersal Fishes | 542.05 | 24.27 |
| 1967–1968 | 2003–2005 | |||||
|---|---|---|---|---|---|---|
| Bay Section | 1 | 2 | 3 | 1 | 2 | 3 |
| Species Richness | 50 | 40 | 36 | 13 | 28 | 24 |
| Simpson’s Index | 0.15 | 0.26 | 0.15 | 0.26 | 0.26 | 0.33 |
| Shannon’s Index | 2.43 | 1.90 | 2.29 | 1.70 | 1.80 | 1.62 |
| Margalef’s Index | 6.00 | 3.99 | 3.94 | 2.09 | 3.25 | 2.77 |
| Shannon’s Evenness | 0.62 | 0.52 | 0.64 | 0.66 | 0.54 | 0.51 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Heppell, S.A.; Heppell, S.S.; Arbuckle, N.S.; Gallagher, M.B. A Cross-Decadal Change in the Fish and Crustacean Community of Lower Yaquina Bay, Oregon, USA. Fishes 2024, 9, 125. https://doi.org/10.3390/fishes9040125
Heppell SA, Heppell SS, Arbuckle NS, Gallagher MB. A Cross-Decadal Change in the Fish and Crustacean Community of Lower Yaquina Bay, Oregon, USA. Fishes. 2024; 9(4):125. https://doi.org/10.3390/fishes9040125
Chicago/Turabian StyleHeppell, Scott A., Selina S. Heppell, N. Scarlett Arbuckle, and M. Brett Gallagher. 2024. "A Cross-Decadal Change in the Fish and Crustacean Community of Lower Yaquina Bay, Oregon, USA" Fishes 9, no. 4: 125. https://doi.org/10.3390/fishes9040125
APA StyleHeppell, S. A., Heppell, S. S., Arbuckle, N. S., & Gallagher, M. B. (2024). A Cross-Decadal Change in the Fish and Crustacean Community of Lower Yaquina Bay, Oregon, USA. Fishes, 9(4), 125. https://doi.org/10.3390/fishes9040125