
Potential Impact of DOM on Fish Biodiversity in the Jinshui River Basin Environment

 , , , ,
, , , ,
Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. eDNA Sample Collection
2.2. eDNA Extraction, PCR Amplification, and Sequencing
2.3. Data Processing
2.4. Rarefaction Curve
2.5. Fish Sequence, Taxonomic, and Functional Diversity
2.6. Differences in Diversity Facets Across Areas
2.7. Spectroscopic Analysis
2.8. Evaluation the Impact of DOM and Physicochemical Parameters on Fish Diversity
3. Results
3.1. eDNA Annotation and Rarefaction Curve
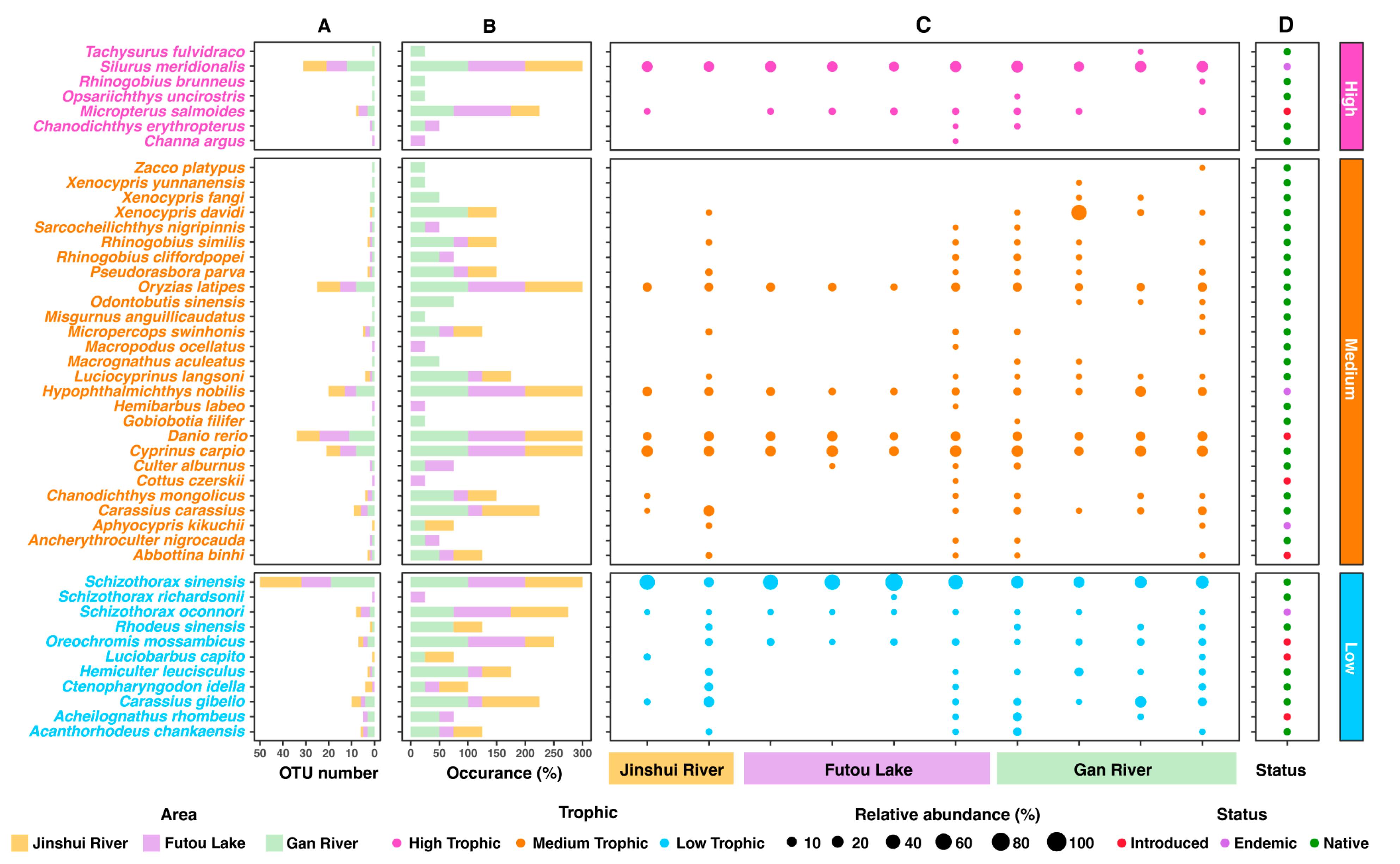
3.2. Fish Trophic Level, Occurrence, and Dominant Species Based on eDNA Analysis
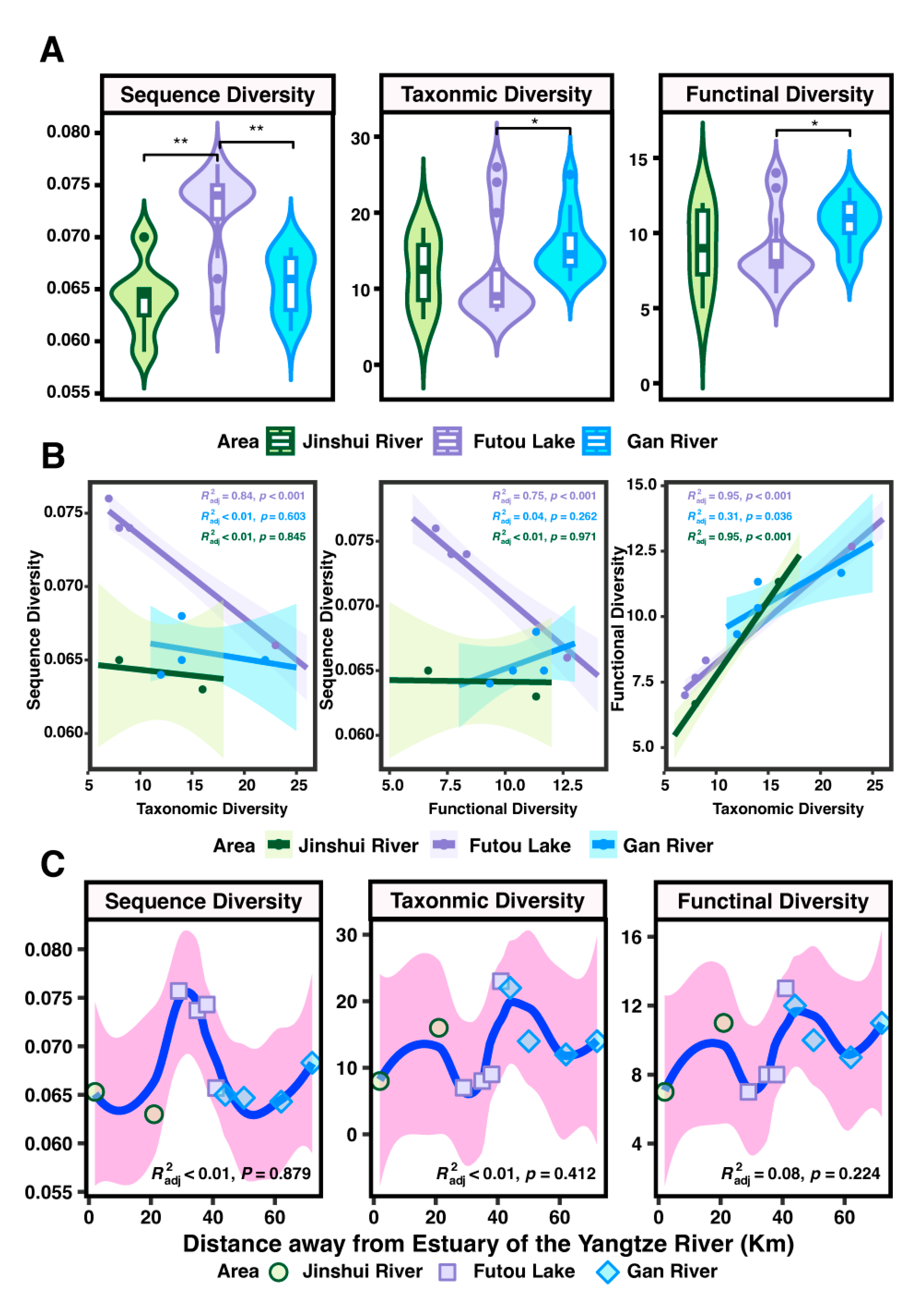
3.3. Diversity Facets: Variations Across Areas
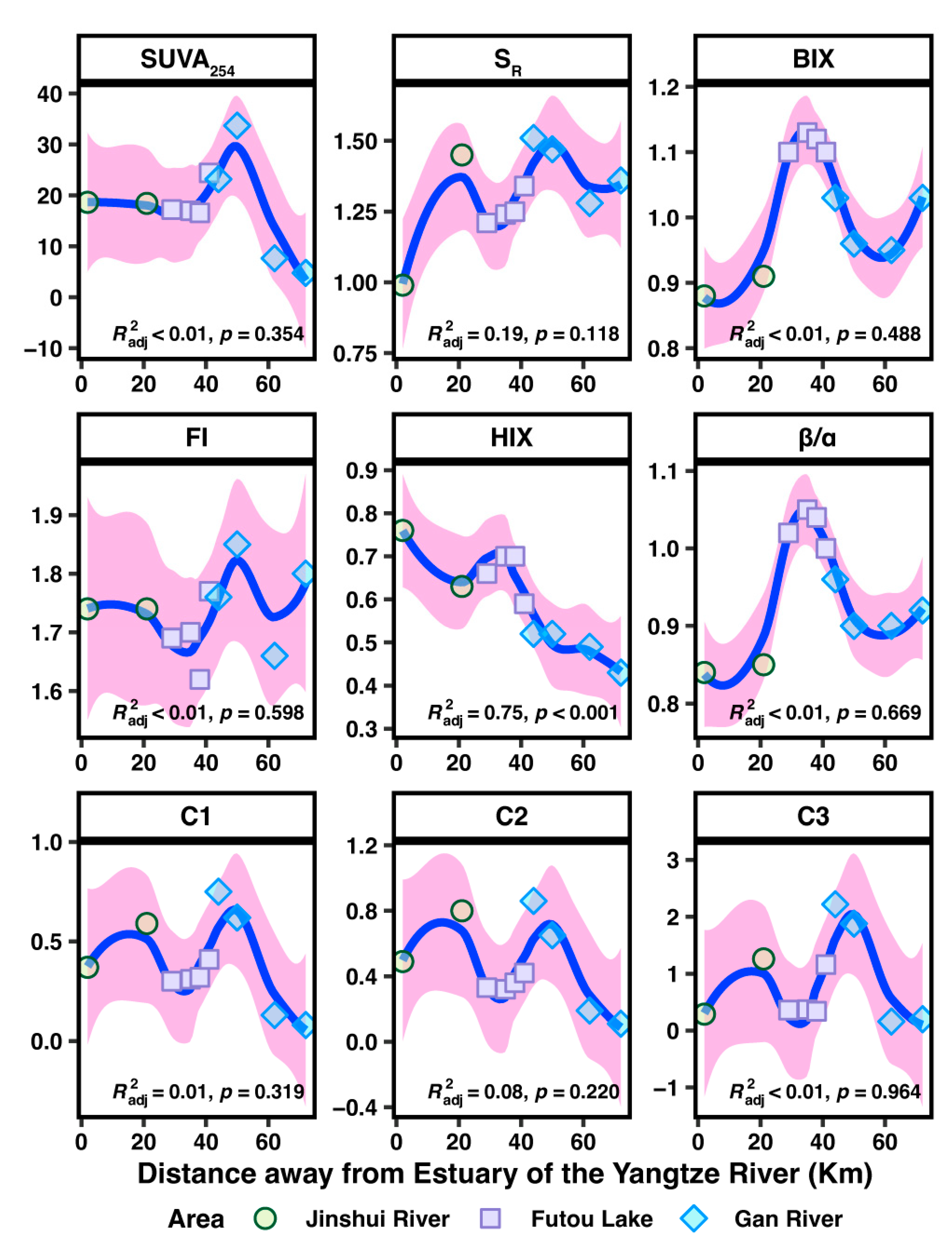
3.4. The Composition, Fluorescence Characteristics, and PARAFAC Components of Dissolved Organic Matter (DOM)
3.5. Impact Evaluation of DOM
4. Discussion
4.1. Impact Evaluation of Physicochemical Parameters and DOM on Fish Diversity Facets
4.2. Factors Determining River Basin Fish Community Structures
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Whitfield, A.K.; Elliott, M. Fishes as Indicators of Environmental and Ecological Changes within Estuaries: A Review of Progress and Some Suggestions for the Future. J. Fish Biol. 2002, 61, 229–250. [Google Scholar] [CrossRef]
- Villéger, S.; Brosse, S.; Mouchet, M.; Mouillot, D.; Vanni, M.J. Functional Ecology of Fish: Current Approaches and Future Challenges. Aquat. Sci. 2017, 79, 783–801. [Google Scholar] [CrossRef]
- Xiang, T.; Dong, X.; Ju, T.; Shi, L.; Grenouillet, G. Anthropogenic Activities and Environmental Filtering Have Reshaped Freshwater Fish Biodiversity Patterns in China over the Past 120 Years. J. Environ. Manag. 2023, 344, 118374. [Google Scholar] [CrossRef]
- Pereira, H.M.; Ferrier, S.; Walters, M.; Geller, G.N.; Jongman, R.H.G.; Scholes, R.J.; Bruford, M.W.; Brummitt, N.; Butchart, S.H.M.; Cardoso, A.C.; et al. Essential Biodiversity Variables. Science 2013, 339, 277–278. [Google Scholar] [CrossRef]
- Paz-Vinas, I.; Loot, G.; Stevens, V.M.; Blanchet, S. Evolutionary Processes Driving Spatial Patterns of Intraspecific Genetic Diversity in River Ecosystems. Mol. Ecol. 2015, 24, 4586–4604. [Google Scholar] [CrossRef]
- Iknayan, K.J.; Tingley, M.W.; Furnas, B.J.; Beissinger, S.R. Detecting Diversity: Emerging Methods to Estimate Species Diversity. Trends Ecol. Evol. 2014, 29, 97–106. [Google Scholar] [CrossRef]
- Jarzyna, M.A.; Jetz, W. Detecting the Multiple Facets of Biodiversity. Trends Ecol. Evol. 2016, 31, 527–538. [Google Scholar] [CrossRef]
- Ji, Y.; Ashton, L.; Pedley, S.M.; Edwards, D.P.; Tang, Y.; Nakamura, A.; Kitching, R.; Dolman, P.M.; Woodcock, P.; Edwards, F.A.; et al. Reliable, Verifiable and Efficient Monitoring of Biodiversity via Metabarcoding. Ecol. Lett. 2013, 16, 1245–1257. [Google Scholar] [CrossRef]
- Bohmann, K.; Evans, A.; Gilbert, M.T.P.; Carvalho, G.R.; Creer, S.; Knapp, M.; Yu, D.W.; De Bruyn, M. Environmental DNA for Wildlife Biology and Biodiversity Monitoring. Trends Ecol. Evol. 2014, 29, 358–367. [Google Scholar] [CrossRef]
- Tsuji, S.; Takahara, T.; Doi, H.; Shibata, N.; Yamanaka, H. The Detection of Aquatic Macroorganisms Using Environmental DNA Analysis—A Review of Methods for Collection, Extraction, and Detection. Environ. DNA 2019, 1, 99–108. [Google Scholar] [CrossRef]
- Wang, S.; Yan, Z.; Hänfling, B.; Zheng, X.; Wang, P.; Fan, J.; Li, J. Methodology of Fish eDNA and Its Applications in Ecology and Environment. Sci. Total Environ. 2021, 755, 142622. [Google Scholar] [CrossRef] [PubMed]
- Sales, N.G.; Wangensteen, O.S.; Carvalho, D.C.; Deiner, K.; Præbel, K.; Coscia, I.; McDevitt, A.D.; Mariani, S. Space-Time Dynamics in Monitoring Neotropical Fish Communities Using eDNA Metabarcoding. Sci. Total Environ. 2021, 754, 142096. [Google Scholar] [CrossRef]
- Van Driessche, C.; Everts, T.; Neyrinck, S.; Halfmaerten, D.; Verschelde, P.; Breine, J.; Bonte, D.; Brys, R. Environmental DNA Metabarcoding Reflects Spatiotemporal Fish Community Shifts in the Scheldt Estuary. Sci. Total Environ. 2024, 934, 173242. [Google Scholar] [CrossRef]
- Hänfling, B.; Lawson Handley, L.; Read, D.S.; Hahn, C.; Li, J.; Nichols, P.; Blackman, R.C.; Oliver, A.; Winfield, I.J. Environmental DNA Metabarcoding of Lake Fish Communities Reflects Long-term Data from Established Survey Methods. Mol. Ecol. 2016, 25, 3101–3119. [Google Scholar] [CrossRef]
- Harper, L.R.; Buxton, A.S.; Rees, H.C.; Bruce, K.; Brys, R.; Halfmaerten, D.; Read, D.S.; Watson, H.V.; Sayer, C.D.; Jones, E.P.; et al. Prospects and Challenges of Environmental DNA (eDNA) Monitoring in Freshwater Ponds. Hydrobiologia 2019, 826, 25–41. [Google Scholar] [CrossRef]
- Zou, K.; Chen, J.; Ruan, H.; Li, Z.; Guo, W.; Li, M.; Liu, L. eDNA Metabarcoding as a Promising Conservation Tool for Monitoring Fish Diversity in a Coastal Wetland of the Pearl River Estuary Compared to Bottom Trawling. Sci. Total Environ. 2020, 702, 134704. [Google Scholar] [CrossRef]
- Rey, A.; Viard, F.; Lizé, A.; Corre, E.; Valentini, A.; Thiriet, P. Coastal Rocky Reef Fish Monitoring in the Context of the Marine Strategy Framework Directive: Environmental DNA Metabarcoding Complements Underwater Visual Census. Ocean. Coast. Manag. 2023, 241, 106625. [Google Scholar] [CrossRef]
- Fraija-Fernández, N.; Bouquieaux, M.-C.; Rey, A.; Mendibil, I.; Cotano, U.; Irigoien, X.; Santos, M.; Rodríguez-Ezpeleta, N. Marine Water Environmental DNA Metabarcoding Provides a Comprehensive Fish Diversity Assessment and Reveals Spatial Patterns in a Large Oceanic Area. Ecol. Evol. 2020, 10, 7560–7584. [Google Scholar] [CrossRef]
- Tsuji, S.; Maruyama, A.; Miya, M.; Ushio, M.; Sato, H.; Minamoto, T.; Yamanaka, H. Environmental DNA Analysis Shows High Potential as a Tool for Estimating Intraspecific Genetic Diversity in a Wild Fish Population. Mol. Ecol. Resour. 2020, 20, 1248–1258. [Google Scholar] [CrossRef]
- Zhang, S.; Lu, Q.; Wang, Y.; Wang, X.; Zhao, J.; Yao, M. Assessment of Fish Communities Using Environmental DNA: Effect of Spatial Sampling Design in Lentic Systems of Different Sizes. Mol. Ecol. Resour. 2020, 20, 242–255. [Google Scholar] [CrossRef]
- Marques, V.; Castagné, P.; Polanco, A.; Borrero-Pérez, G.H.; Hocdé, R.; Guérin, P.; Juhel, J.; Velez, L.; Loiseau, N.; Letessier, T.B.; et al. Use of Environmental DNA in Assessment of Fish Functional and Phylogenetic Diversity. Conserv. Biol. 2021, 35, 1944–1956. [Google Scholar] [CrossRef] [PubMed]
- Su, G.; Logez, M.; Xu, J.; Tao, S.; Villéger, S.; Brosse, S. Human Impacts on Global Freshwater Fish Biodiversity. Science 2021, 371, 835–838. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Liu, X.; Zhao, G.; Zuo, C.; Alofs, K.; Wang, R. Pathways Linking Watershed Development and Riparian Quality to Stream Water Quality and Fish Communities: Insights from 233 Subbasins of the Great Lakes Region. Water Res. 2024, 261, 121964. [Google Scholar] [CrossRef] [PubMed]
- De Sousa, R.S.; Silva, G.C.; Bazzan, T.; De La Torre, F.; Nebo, C.; Siqueira-Silva, D.H.; Cardoso-Silva, S.; Pompêo, M.L.M.; De Paiva, T.C.B.; Da Silva, F.T.; et al. Connections among Land Use, Water Quality, Biodiversity of Aquatic Invertebrates, and Fish Behavior in Amazon Rivers. Toxics 2022, 10, 182. [Google Scholar] [CrossRef]
- Topić Popović, N.; Strunjak-Perović, I.; Barišić, J.; Kepec, S.; Jadan, M.; Beer-Ljubić, B.; Matijatko, V.; Palić, D.; Klobučar, G.; Babić, S.; et al. Native Prussian Carp (Carassius gibelio) Health Status, Biochemical and Histological Responses to Treated Wastewaters. Environ. Pollut. 2016, 218, 689–701. [Google Scholar] [CrossRef]
- Araújo, C.V.M.; Laissaoui, A.; Silva, D.C.V.R.; Ramos-Rodríguez, E.; González-Ortegón, E.; Espíndola, E.L.G.; Baldó, F.; Mena, F.; Parra, G.; Blasco, J.; et al. Not Only Toxic but Repellent: What Can Organisms’ Responses Tell Us about Contamination and What Are the Ecological Consequences When They Flee from an Environment? Toxics 2020, 8, 118. [Google Scholar] [CrossRef]
- Zhang, H.; Zheng, Y.; Wang, X.C.; Wang, Y.; Dzakpasu, M. Characterization and Biogeochemical Implications of Dissolved Organic Matter in Aquatic Environments. J. Environ. Manag. 2021, 294, 113041. [Google Scholar] [CrossRef]
- Wood, C.M.; Al-Reasi, H.A.; Smith, D.S. The Two Faces of DOC. Aquat. Toxicol. 2011, 105, 3–8. [Google Scholar] [CrossRef]
- Schwartz, M.L.; Curtis, P.J.; Playle, R.C. Influence of Natural Organic Matter Source on Acute Copper, Lead, and Cadmium Toxicity to Rainbow Trout (Oncorhynchus mykiss). Environ. Toxicol. Chem. 2004, 23, 2889–2899. [Google Scholar] [CrossRef]
- Cornelissen, G.; Okkenhaug, G.; Breedveld, G.D.; Sørlie, J.-E. Transport of Polycyclic Aromatic Hydrocarbons and Polychlorinated Biphenyls in a Landfill: A Novel Equilibrium Passive Sampler to Determine Free and Total Dissolved Concentrations in Leachate Water. J. Hydrol. 2009, 369, 253–259. [Google Scholar] [CrossRef]
- Artifon, V.; Zanardi-Lamardo, E.; Fillmann, G. Aquatic Organic Matter: Classification and Interaction with Organic Microcontaminants. Sci. Total Environ. 2019, 649, 1620–1635. [Google Scholar] [CrossRef] [PubMed]
- Baken, S.; Degryse, F.; Verheyen, L.; Merckx, R.; Smolders, E. Metal Complexation Properties of Freshwater Dissolved Organic Matter Are Explained by Its Aromaticity and by Anthropogenic Ligands. Environ. Sci. Technol. 2011, 45, 2584–2590. [Google Scholar] [CrossRef]
- Sun, Y.; Xie, Z.; Wu, K.; Lan, J.; Li, T.; Yuan, D. Speciation, Distribution and Migration Pathways of Polycyclic Aromatic Hydrocarbons in a Typical Underground River System in Southwest China. J. Hydrol. 2021, 596, 125690. [Google Scholar] [CrossRef]
- Reynaud, S.; Deschaux, P. The Effects of Polycyclic Aromatic Hydrocarbons on the Immune System of Fish: A Review. Aquat. Toxicol. 2006, 77, 229–238. [Google Scholar] [CrossRef]
- Honda, M.; Suzuki, N. Toxicities of Polycyclic Aromatic Hydrocarbons for Aquatic Animals. Int. J. Environ. Res. Public Health 2020, 17, 1363. [Google Scholar] [CrossRef]
- Moeckel, C.; Monteith, D.T.; Llewellyn, N.R.; Henrys, P.A.; Pereira, M.G. Relationship between the Concentrations of Dissolved Organic Matter and Polycyclic Aromatic Hydrocarbons in a Typical U.K. Upland Stream. Environ. Sci. Technol. 2014, 48, 130–138. [Google Scholar] [CrossRef]
- Giles, M.A. Electrolyte and Water Balance in Plasma and Urine of Rainbow Trout (Salmo gairdneri) during Chronic Exposure to Cadmium. Can. J. Fish. Aquat. Sci. 1984, 41, 1678–1685. [Google Scholar] [CrossRef]
- Laurén, D.J.; McDonald, D.G. Acclimation to Copper by Rainbow Trout, Salmo gairdneri: Physiology. Can. J. Fish. Aquat. Sci. 1987, 44, 99–104. [Google Scholar] [CrossRef]
- Hollis, L.; McGeer, J.C.; McDonald, D.G.; Wood, C.M. Cadmium Accumulation, Gill Cd Binding, Acclimation, and Physiological Effects during Long Term Sublethal Cd Exposure in Rainbow Trout. Aquat. Toxicol. 1999, 46, 101–119. [Google Scholar] [CrossRef]
- Campbell, P.G.; Twiss, M.R.; Wilkinson, K.J. Accumulation of Natural Organic Matter on the Surfaces of Living Cells: Implications for the Interaction of Toxic Solutes with Aquatic Biota. Can. J. Fish. Aquat. Sci. 1997, 54, 2543–2554. [Google Scholar] [CrossRef]
- Galvez, F.; Donini, A.; Playle, R.C.; Smith, D.S.; O’Donnell, M.J.; Wood, C.M. A Matter of Potential Concern: Natural Organic Matter Alters the Electrical Properties of Fish Gills. Environ. Sci. Technol. 2008, 42, 9385–9390. [Google Scholar] [CrossRef] [PubMed]
- Crémazy, A.; Braz-Mota, S.; Brix, K.V.; Duarte, R.M.; Val, A.L.; Wood, C.M. Investigating the Mechanisms of Dissolved Organic Matter Protection against Copper Toxicity in Fish of Amazon’s Black Waters. Sci. Total Environ. 2022, 843, 157032. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, A.Y.O.; Playle, R.C.; Val, A.L.; Wood, C.M. Physiological Action of Dissolved Organic Matter in Rainbow Trout in the Presence and Absence of Copper: Sodium Uptake Kinetics and Unidirectional Flux Rates in Hard and Softwater. Aquat. Toxicol. 2004, 70, 63–81. [Google Scholar] [CrossRef] [PubMed]
- Burnison, B.K.; Meinelt, T.; Playle, R.; Pietrock, M.; Wienke, A.; Steinberg, C.E.W. Cadmium Accumulation in Zebrafish (Danio rerio) Eggs Is Modulated by Dissolved Organic Matter (DOM). Aquat. Toxicol. 2006, 79, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Duarte, R.M.; Smith, D.S.; Val, A.L.; Wood, C.M. Dissolved Organic Carbon from the Upper Rio Negro Protects Zebrafish (Danio rerio) against Ionoregulatory Disturbances Caused by Low pH Exposure. Sci. Rep. 2016, 6, 20377. [Google Scholar] [CrossRef]
- Wood, C.M.; Matsuo, A.Y.O.; Wilson, R.W.; Gonzalez, R.J.; Patrick, M.L.; Playle, R.C.; Luis Val, A. Protection by Natural Blackwater against Disturbances in Ion Fluxes Caused by Low pH Exposure in Freshwater Stingrays Endemic to the Rio Negro. Physiol. Biochem. Zool. 2003, 76, 12–27. [Google Scholar] [CrossRef]
- Chen, S.; Ke, R.; Zha, J.; Wang, Z.; Khan, S.U. Influence of Humic Acid on Bioavailability and Toxicity of Benzo[k]Fluoranthene to Japanese Medaka. Environ. Sci. Technol. 2008, 42, 9431–9436. [Google Scholar] [CrossRef]
- Liu, Y.; Yang, C.; Cheng, P.; He, X.; Zhu, Y.; Zhang, Y. Influences of Humic Acid on the Bioavailability of Phenanthrene and Alkyl Phenanthrenes to Early Life Stages of Marine Medaka (Oryzias melastigma). Environ. Pollut. 2016, 210, 211–216. [Google Scholar] [CrossRef]
- Zhao, Q.; Zhu, L. Effect of Humic Acid on Prometryn Bioaccumulation and the Induction of Oxidative Stress in Zebrafish (Danio rerio). RSC Adv. 2016, 6, 16790–16797. [Google Scholar] [CrossRef]
- Ye, S.; Li, Z.; Zhang, T.; Liu, J.; Xie, S. Assessing Fish Distribution and Threats to Fish Biodiversity in the Yangtze River Basin, China. Ichthyol. Res. 2014, 61, 183–188. [Google Scholar] [CrossRef]
- Qian, M.-M.; Wang, Z.-Y.; Zhou, Q.; Wang, J.; Shao, Y.; Qiao, Q.; Fan, J.-T.; Yan, Z.-G. Environmental DNA Unveiling the Fish Community Structure and Diversity Features in the Yangtze River Basin. Environ. Res. 2023, 239, 117198. [Google Scholar] [CrossRef] [PubMed]
- Zhong, W.; Zhang, J.; Wang, Z.; Lin, J.; Huang, X.; Liu, W.; Li, H.; Pellissier, L.; Zhang, X. Holistic Impact Evaluation of Human Activities on the Coastal Fish Biodiversity in the Chinese Coastal Environment. Environ. Sci. Technol. 2022, 56, 6574–6583. [Google Scholar] [CrossRef] [PubMed]
- Miya, M.; Sato, Y.; Fukunaga, T.; Sado, T.; Poulsen, J.Y.; Sato, K.; Minamoto, T.; Yamamoto, S.; Yamanaka, H.; Araki, H.; et al. MiFish, a Set of Universal PCR Primers for Metabarcoding Environmental DNA from Fishes: Detection of More than 230 Subtropical Marine Species. R. Soc. Open Sci. 2015, 2, 150088. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. Fastp: An Ultra-Fast All-in-One FASTQ Preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Magoč, T.; Salzberg, S.L. FLASH: Fast Length Adjustment of Short Reads to Improve Genome Assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef]
- Stackebrandt, E.; Goebel, B.M. Taxonomic Note: A Place for DNA-DNA Reassociation and 16S rRNA Sequence Analysis in the Present Species Definition in Bacteriology. Int. J. Syst. Evol. Microbiol. 1994, 44, 846–849. [Google Scholar] [CrossRef]
- Edgar, R.C. UPARSE: Highly Accurate OTU Sequences from Microbial Amplicon Reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Manel, S.; Guerin, P.-E.; Mouillot, D.; Blanchet, S.; Velez, L.; Albouy, C.; Pellissier, L. Global Determinants of Freshwater and Marine Fish Genetic Diversity. Nat. Commun. 2020, 11, 692. [Google Scholar] [CrossRef]
- Murphy, K.R.; Hambly, A.; Singh, S.; Henderson, R.K.; Baker, A.; Stuetz, R.; Khan, S.J. Organic Matter Fluorescence in Municipal Water Recycling Schemes: Toward a Unified PARAFAC Model. Environ. Sci. 2011, 45, 2909–2916. [Google Scholar] [CrossRef]
- Jørgensen, L. Global Trends in the Fluorescence Characteristics and Distribution of Marine Dissolved Organic Matter. Mar. Chem. 2011, 126, 139–148. [Google Scholar] [CrossRef]
- Yu, H.; Liang, H.; Qu, F.; Han, Z.; Shao, S.; Chang, H.; Li, G. Impact of Dataset Diversity on Accuracy and Sensitivity of Parallel Factor Analysis Model of Dissolved Organic Matter Fluorescence Excitation-Emission Matrix. Sci. Rep. 2015, 5, 10207. [Google Scholar] [CrossRef] [PubMed]
- Zang, C.; Huang, S.; Wu, M.; Du, S.; Scholz, M.; Gao, F.; Lin, C.; Guo, Y.; Dong, Y. Comparison of Relationships Between pH, Dissolved Oxygen and Chlorophyll a for Aquaculture and Non-Aquaculture Waters. Water Air Soil Pollut. 2011, 219, 157–174. [Google Scholar] [CrossRef]
- Whiles, M.R.; Huryn, A.D.; Taylor, B.W.; Reeve, J.D. Influence of Handling Stress and Fasting on Estimates of Ammonium Excretion by Tadpoles and Fish: Recommendations for Designing Excretion Experiments. Limnol. Oceanogr. Methods 2009, 7, 1–7. [Google Scholar] [CrossRef]
- Edwards, T.M.; Puglis, H.J.; Kent, D.B.; Durán, J.L.; Bradshaw, L.M.; Farag, A.M. Ammonia and Aquatic Ecosystems—A Review of Global Sources, Biogeochemical Cycling, and Effects on Fish. Sci. Total Environ. 2024, 907, 167911. [Google Scholar] [CrossRef]
- Fu, X.; Zheng, M.; Su, J.; Xi, B.; Wei, D.; Wang, X. Spatiotemporal Patterns and Threshold of Chlorophyll-a in Lake Taihu Based on Microcystins. Environ. Sci. Pollut. Res. 2023, 30, 49327–49338. [Google Scholar] [CrossRef]
- Howarth, R.; Chan, F.; Conley, D.J.; Garnier, J.; Doney, S.C.; Marino, R.; Billen, G. Coupled Biogeochemical Cycles: Eutrophication and Hypoxia in Temperate Estuaries and Coastal Marine Ecosystems. Front. Ecol. Environ. 2011, 9, 18–26. [Google Scholar] [CrossRef]
- Fourtune, L.; Paz-Vinas, I.; Loot, G.; Prunier, J.G.; Blanchet, S. Lessons from the Fish: A Multi-Species Analysis Reveals Common Processes Underlying Similar Species-Genetic Diversity Correlations. Freshw. Biol. 2016, 61, 1830–1845. [Google Scholar] [CrossRef]
- Wang, L.; Lyons, J.; Kanehl, P.; Bannerman, R. Impacts of Urbanization on Stream Habitat and Fish Across Multiple Spatial Scales. Environ. Manag. 2001, 28, 255–266. [Google Scholar] [CrossRef]
- Rahel, F.J.; Olden, J.D. Assessing the Effects of Climate Change on Aquatic Invasive Species. Conserv. Biol. 2008, 22, 521–533. [Google Scholar] [CrossRef]
- López Muñoz, I.; Bernard, O. Modeling the Influence of Temperature, Light Intensity and Oxygen Concentration on Microalgal Growth Rate. Processes 2021, 9, 496. [Google Scholar] [CrossRef]
- Matthews, W.J.; Marsh-Matthews, E. Effects of Drought on Fish across Axes of Space, Time and Ecological Complexity. Freshw. Biol. 2003, 48, 1232–1253. [Google Scholar] [CrossRef]
- Hansen, A.M.; Kraus, T.E.C.; Pellerin, B.A.; Fleck, J.A.; Downing, B.D.; Bergamaschi, B.A. Optical Properties of Dissolved Organic Matter (DOM): Effects of Biological and Photolytic Degradation. Limnol. Oceanogr. 2016, 61, 1015–1032. [Google Scholar] [CrossRef]
- Hu, A.; Choi, M.; Tanentzap, A.J.; Liu, J.; Jang, K.-S.; Lennon, J.T.; Liu, Y.; Soininen, J.; Lu, X.; Zhang, Y.; et al. Ecological Networks of Dissolved Organic Matter and Microorganisms under Global Change. Nat. Commun. 2022, 13, 3600. [Google Scholar] [CrossRef] [PubMed]
- Malzahn, A.M.; Aberle, N.; Clemmesen, C.; Boersma, M. Nutrient Limitation of Primary Producers Affects Planktivorous Fish Condition. Limnol. Oceanogr. 2007, 52, 2062–2071. [Google Scholar] [CrossRef]
- Boulion, V.V. Relationship between Phytoplankton, Heterotrophic Plankton, and Planktivorous Fish Productions in Different Water Bodies. Dokl. Biol. Sci. 2023, 513, S5–S9. [Google Scholar] [CrossRef]
- Suárez-Castro, A.F.; Raymundo, M.; Bimler, M.; Mayfield, M.M. Using Multi-Scale Spatially Explicit Frameworks to Understand the Relationship between Functional Diversity and Species Richness. Ecography 2022, 2022, e05844. [Google Scholar] [CrossRef]
- Helms, J.R.; Stubbins, A.; Ritchie, J.D.; Minor, E.C.; Kieber, D.J.; Mopper, K. Absorption Spectral Slopes and Slope Ratios as Indicators of Molecular Weight, Source, and Photobleaching of Chromophoric Dissolved Organic Matter. Limnol. Oceanogr. 2008, 53, 955–969. [Google Scholar] [CrossRef]
- Bai, Y.; Zhang, S.; Mu, E.; Zhao, Y.; Cheng, L.; Zhu, Y.; Yuan, Y.; Wang, Y.; Ding, A. Characterizing the Spatiotemporal Distribution of Dissolved Organic Matter (DOM) in the Yongding River Basin: Insights from Flow Regulation. J. Environ. Manag. 2023, 325, 116476. [Google Scholar] [CrossRef]
- Zhang, Y.; Van Dijk, M.A.; Liu, M.; Zhu, G.; Qin, B. The Contribution of Phytoplankton Degradation to Chromophoric Dissolved Organic Matter (CDOM) in Eutrophic Shallow Lakes: Field and Experimental Evidence. Water Res. 2009, 43, 4685–4697. [Google Scholar] [CrossRef]
- Liu, Q.; Jiang, Y.; Tian, Y.; Hou, Z.; He, K.; Fu, L.; Xu, H. Impact of Land Use on the DOM Composition in Different Seasons in a Subtropical River Flowing through a Region Undergoing Rapid Urbanization. J. Clean. Prod. 2019, 212, 1224–1231. [Google Scholar] [CrossRef]
- Ahoutou, M.K.; Yao, E.K.; Djeha, R.Y.; Kone, M.; Tambosco, K.; Duval, C.; Hamlaoui, S.; Bernard, C.; Bouvy, M.; Marie, B.; et al. Impacts of Nutrient Loading and Fish Grazing on the Phytoplankton Community and Cyanotoxin Production in a Shallow Tropical Lake: Results from Mesocosm Experiments. MicrobiologyOpen 2022, 11, e1278. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Wu, X.; Zhi, G.; Yang, Y.; Wu, L.; Zhang, Y.; Zheng, B.; Qadeer, A.; Zheng, J.; Deng, W.; et al. Fluorescence Characteristics of DOM and Its Influence on Water Quality of Rivers and Lakes in the Dianchi Lake Basin. Ecol. Indic. 2022, 142, 109088. [Google Scholar] [CrossRef]
- Meinelt, T.; Kroupova, H.; Stüber, A.; Rennert, B.; Wienke, A.; Steinberg, C.E.W. Can Dissolved Aquatic Humic Substances Reduce the Toxicity of Ammonia and Nitrite in Recirculating Aquaculture Systems? Aquaculture 2010, 306, 378–383. [Google Scholar] [CrossRef]
- Peuranen, S.; Vuorinen, P.J.; Vuorinen, M.; Hollender, A. The Effects of Iron, Humic Acids and Low pH on the Gills and Physiology of Brown Trout (Salmo trutta). Ann. Zool. Fenn. 1994, 31, 389–396. [Google Scholar]
- Hammock, D.; Huang, C.C.; Mort, G.; Swinehart, J.H. The Effect of Humic Acid on the Uptake of Mercury(II), Cadmium(II), and Zinc(II) by Chinook Salmon (Oncorhynchus tshawytscha) Eggs. Arch. Environ. Contam. Toxicol. 2003, 44, 83–88. [Google Scholar] [CrossRef]
- Yamin, G.; Borisover, M.; Cohen, E.; Van Rijn, J. Accumulation of Humic-like and Proteinaceous Dissolved Organic Matter in Zero-Discharge Aquaculture Systems as Revealed by Fluorescence EEM Spectroscopy. Water Res. 2017, 108, 412–421. [Google Scholar] [CrossRef]
- Duque, G.; Gamboa-García, D.E.; Molina, A.; Cogua, P. Effect of Water Quality Variation on Fish Assemblages in an Anthropogenically Impacted Tropical Estuary, Colombian Pacific. Environ. Sci. Pollut. Res. 2020, 27, 25740–25753. [Google Scholar] [CrossRef]
- Ramírez-Herrejón, J.; Moncayo-Estrada, R.; Balart, E.; García Camacho, L.; Vital Rodríguez, B.; Alvarado Villanueva, R.; Ortega Murillo, R.; Caraveo-Patiño, J. Trophic Interrelations between Introduced Common Carp, Cyprinus Carpio (Actinopterygii: Cypriniformes: Cyprinidae), and Fish Community in a Eutrophic Shallow Lake. Acta Ichthyol. Piscat. 2014, 44, 45–58. [Google Scholar] [CrossRef]
- Wu, J.; Mao, R.; Li, M.; Xia, J.; Song, J.; Cheng, D.; Sun, H. Assessment of Aquatic Ecological Health Based on Determination of Biological Community Variability of Fish and Macroinvertebrates in the Weihe River Basin, China. J. Environ. Manag. 2020, 267, 110651. [Google Scholar] [CrossRef]
- Wang, C.; Zhang, H.; Lei, P.; Xin, X.; Zhang, A.; Yin, W. Evidence on the Causes of the Rising Levels of CODMn along the Middle Route of the South-to-North Diversion Project in China: The Role of Algal Dissolved Organic Matter. J. Environ. Sci. 2022, 113, 281–290. [Google Scholar] [CrossRef]
- Weishaar, J.L.; Aiken, G.R.; Bergamaschi, B.A.; Fram, M.S.; Fujii, R.; Mopper, K. Evaluation of Specific Ultraviolet Absorbance as an Indicator of the Chemical Composition and Reactivity of Dissolved Organic Carbon. Environ. Sci. Technol. 2003, 37, 4702–4708. [Google Scholar] [CrossRef] [PubMed]
- Huguet, A.; Vacher, L.; Relexans, S.; Saubusse, S.; Froidefond, J.M.; Parlanti, E. Properties of Fluorescent Dissolved Organic Matter in the Gironde Estuary. Org. Geochem. 2009, 40, 706–719. [Google Scholar] [CrossRef]
- Parlanti, E.; Wörz, K.; Geoffroy, L.; Lamotte, M. Dissolved Organic Matter Fluorescence Spectroscopy as a Tool to Estimate Biological Activity in a Coastal Zone Submitted to Anthropogenic Inputs. Org. Geochem. 2000, 31, 1765–1781. [Google Scholar] [CrossRef]
- Cory, R.M.; McKnight, D.M. Fluorescence Spectroscopy Reveals Ubiquitous Presence of Oxidized and Reduced Quinones in Dissolved Organic Matter. Environ. Sci. Technol. 2005, 39, 8142–8149. [Google Scholar] [CrossRef]
- McKnight, D.M.; Boyer, E.W.; Westerhoff, P.K.; Doran, P.T.; Kulbe, T.; Andersen, D.T. Spectrofluorometric Characterization of Dissolved Organic Matter for Indication of Precursor Organic Material and Aromaticity. Limnol. Oceanogr. 2001, 46, 38–48. [Google Scholar] [CrossRef]
- Ohno, T. Fluorescence Inner-Filtering Correction for Determining the Humification Index of Dissolved Organic Matter. Environ. Sci. Technol. 2002, 36, 742–746. [Google Scholar] [CrossRef]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2—Approximately Maximum-Likelihood Trees for Large Alignments. PLoS ONE 2010, 5, e9490. [Google Scholar] [CrossRef]
- Revell, L.J. Phytools: An R Package for Phylogenetic Comparative Biology (and Other Things). Methods Ecol. Evol. 2012, 3, 217–223. [Google Scholar] [CrossRef]
- Poelen, J.H.; Simons, J.D.; Mungall, C.J. Global Biotic Interactions: An Open Infrastructure to Share and Analyze Species-Interaction Datasets. Ecol. Inform. 2014, 24, 148–159. [Google Scholar] [CrossRef]
- Csardi, G.; Nepusz, T. The Igraph Software Package for Complex Network Research. InterJournal 2005, Complex Systems, 1695. Available online: https://igraph.org (accessed on 28 May 2024).
- Murphy, K.; Stedmon, C.; Wenig, P.; Bro, R. OpenFluor—A Spectral Database of Auto-Fluorescence by Organic Compounds in the Environment. Anal. Methods 2014, 6, 658. [Google Scholar] [CrossRef]
- Wang, S. Suspect Screening to Support Source Identification and Risk Assessment of Organic Micropollutants in the Aquatic Environment of a Sub-Saharan African Urban Center. Water Res. 2022, 220, 118706. [Google Scholar] [CrossRef] [PubMed]
- Wang, H. Spectral and Isotopic Characteristics of Particulate Organic Matter in a Subtropical Estuary under the Influences of Human Disturbance. J. Mar. Syst. 2020, 203, 103264. [Google Scholar] [CrossRef]
- Krylov, I.N.; Drozdova, A.N.; Labutin, T.A. Albatross R Package to Study PARAFAC Components of DOM Fluorescence from Mixing Zones of Arctic Shelf Seas. Chemom. Intell. Lab. Syst. 2020, 207, 104176. [Google Scholar] [CrossRef]
- Eder, A. Pathways and Composition of Dissolved Organic Carbon in a Small Agricultural Catchment during Base Flow Conditions. Ecohydrol. Hydrobiol. 2022, 22, 96–112. [Google Scholar] [CrossRef]
- Lee, D. Characteristics of Intracellular Algogenic Organic Matter and Its Reactivity with Hydroxyl Radicals. Water Res. 2018, 144, 13–25. [Google Scholar] [CrossRef]
- Murphy, K.R.; Ruiz, G.M.; Dunsmuir, W.T.M.; Waite, T.D. Optimized Parameters for Fluorescence-Based Verification of Ballast Water Exchange by Ships. Environ. Sci. Technol. 2006, 40, 2357–2362. [Google Scholar] [CrossRef]
- Stedmon, C.; Thomas, D.; Papadimitriou, S.; Granskog, M.; Dieckmann, G. Using Fluorescence to Characterize Dissolved Organic Matter in Antarctic Sea Ice Brines. J. Geophys. Res. 2011, 116, G03027. [Google Scholar] [CrossRef]
- Du, Y. Composition of Dissolved Organic Matter Controls Interactions with La and Al Ions: Implications for Phosphorus Immobilization in Eutrophic Lakes. Environ. Pollut. 2019, 248, 36–47. [Google Scholar] [CrossRef]
- Brogi, S.R.; Derrien, M.; Hur, J. In-Depth Assessment of the Effect of Sodium Azide on the Optical Properties of Dissolved Organic Matter. J. Fluoresc. 2019, 29, 877–885. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheng, S.; Zhang, J.; Xue, H.; Yang, Q.; Liu, J.; Zhang, L.; Liu, N.; Wang, Y.; Meng, F. Potential Impact of DOM on Fish Biodiversity in the Jinshui River Basin Environment. Fishes 2024, 9, 489. https://doi.org/10.3390/fishes9120489
Cheng S, Zhang J, Xue H, Yang Q, Liu J, Zhang L, Liu N, Wang Y, Meng F. Potential Impact of DOM on Fish Biodiversity in the Jinshui River Basin Environment. Fishes. 2024; 9(12):489. https://doi.org/10.3390/fishes9120489
Chicago/Turabian StyleCheng, Shoutao, Jiasheng Zhang, Hao Xue, Qi Yang, Jing Liu, Lingsong Zhang, Na Liu, Yeyao Wang, and Fansheng Meng. 2024. "Potential Impact of DOM on Fish Biodiversity in the Jinshui River Basin Environment" Fishes 9, no. 12: 489. https://doi.org/10.3390/fishes9120489
APA StyleCheng, S., Zhang, J., Xue, H., Yang, Q., Liu, J., Zhang, L., Liu, N., Wang, Y., & Meng, F. (2024). Potential Impact of DOM on Fish Biodiversity in the Jinshui River Basin Environment. Fishes, 9(12), 489. https://doi.org/10.3390/fishes9120489