

In Silico Evaluation of the PCR Performance of Different Tests for Detection of WSSV


Abstract
:1. Introduction
2. Materials and Methods
2.1. Analysis In Silico of Primers for WSSV Detection
2.2. In Silico Analysis
2.2.1. Primer Specificity
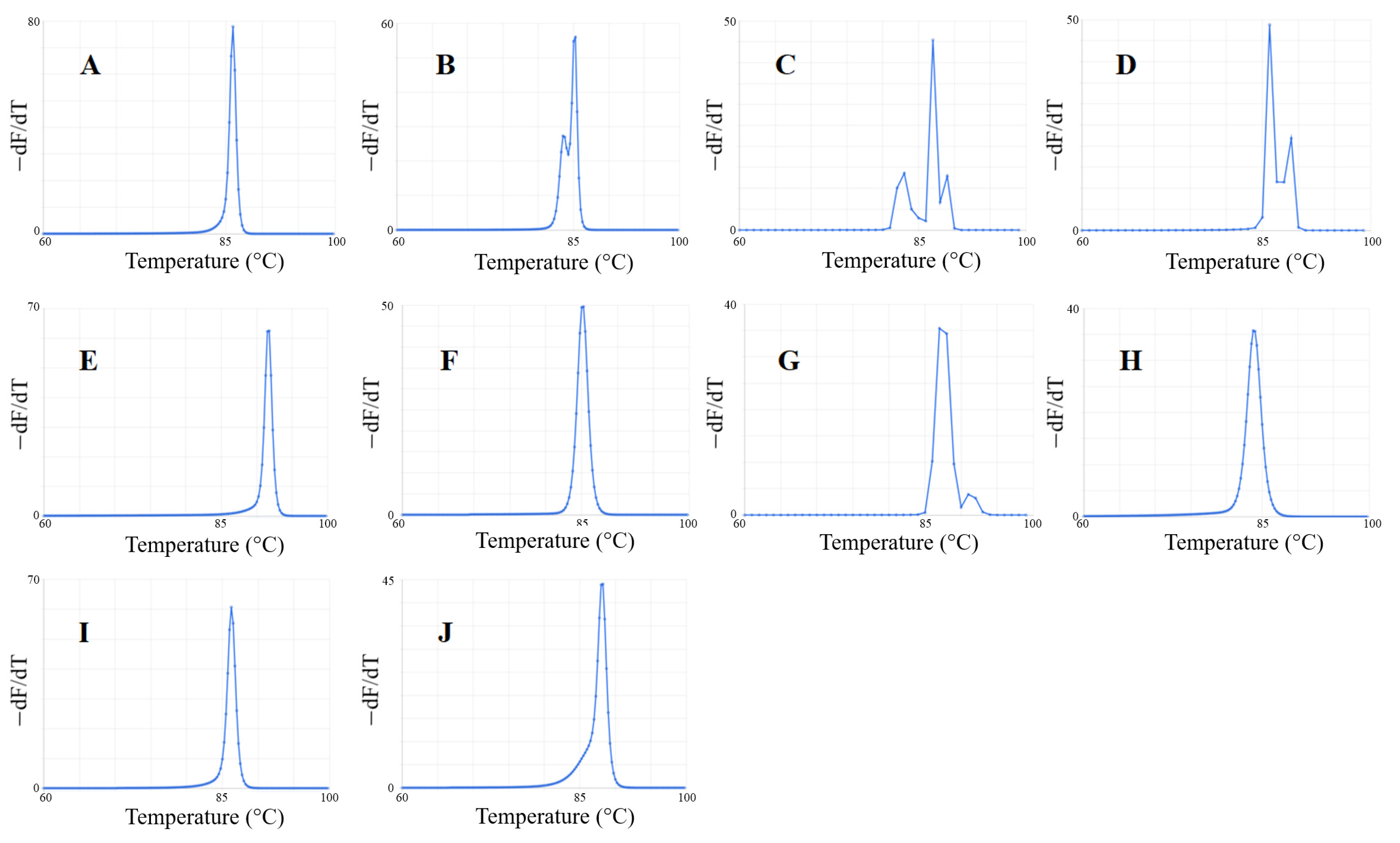
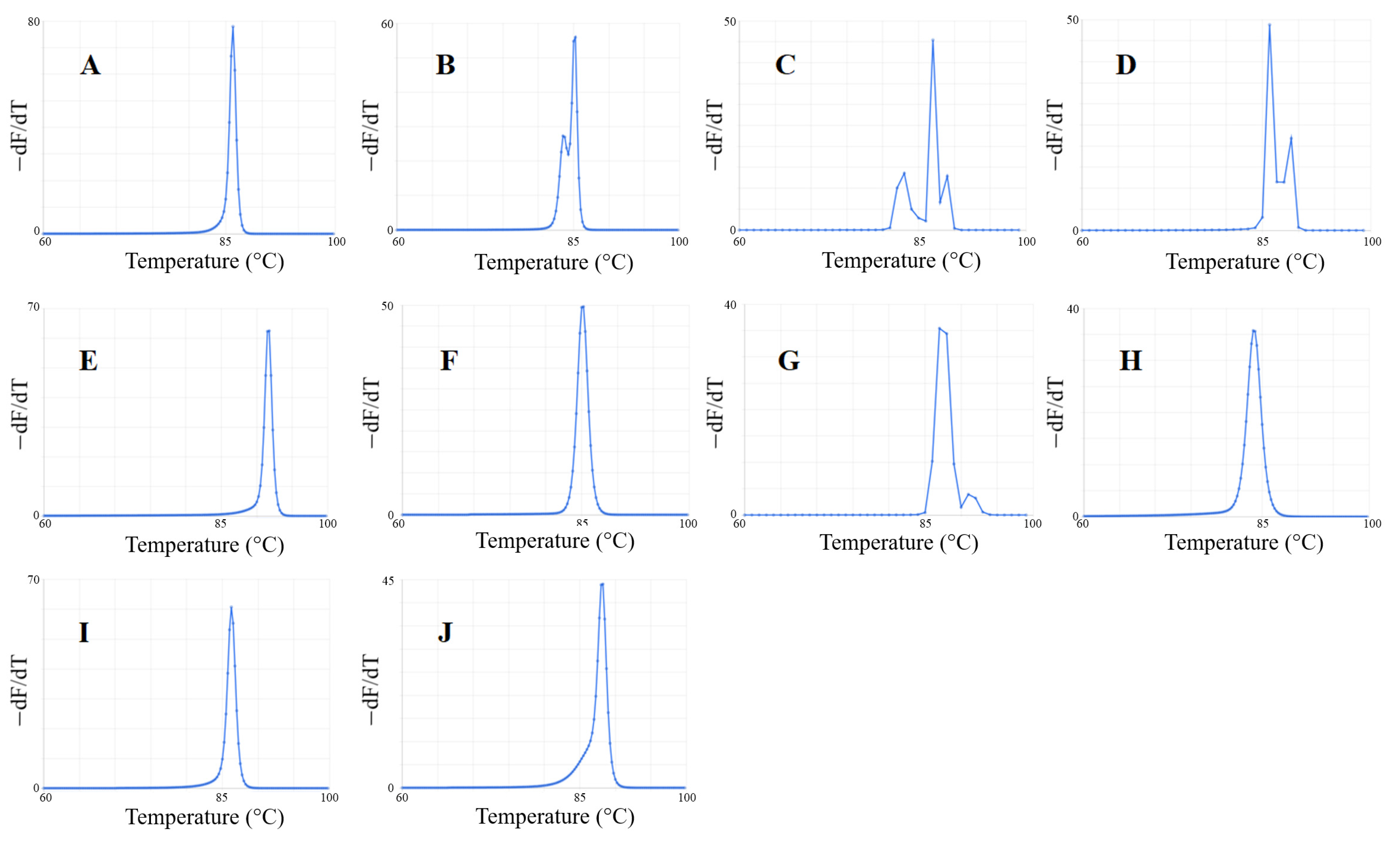
2.2.2. Predicted Melt Curves
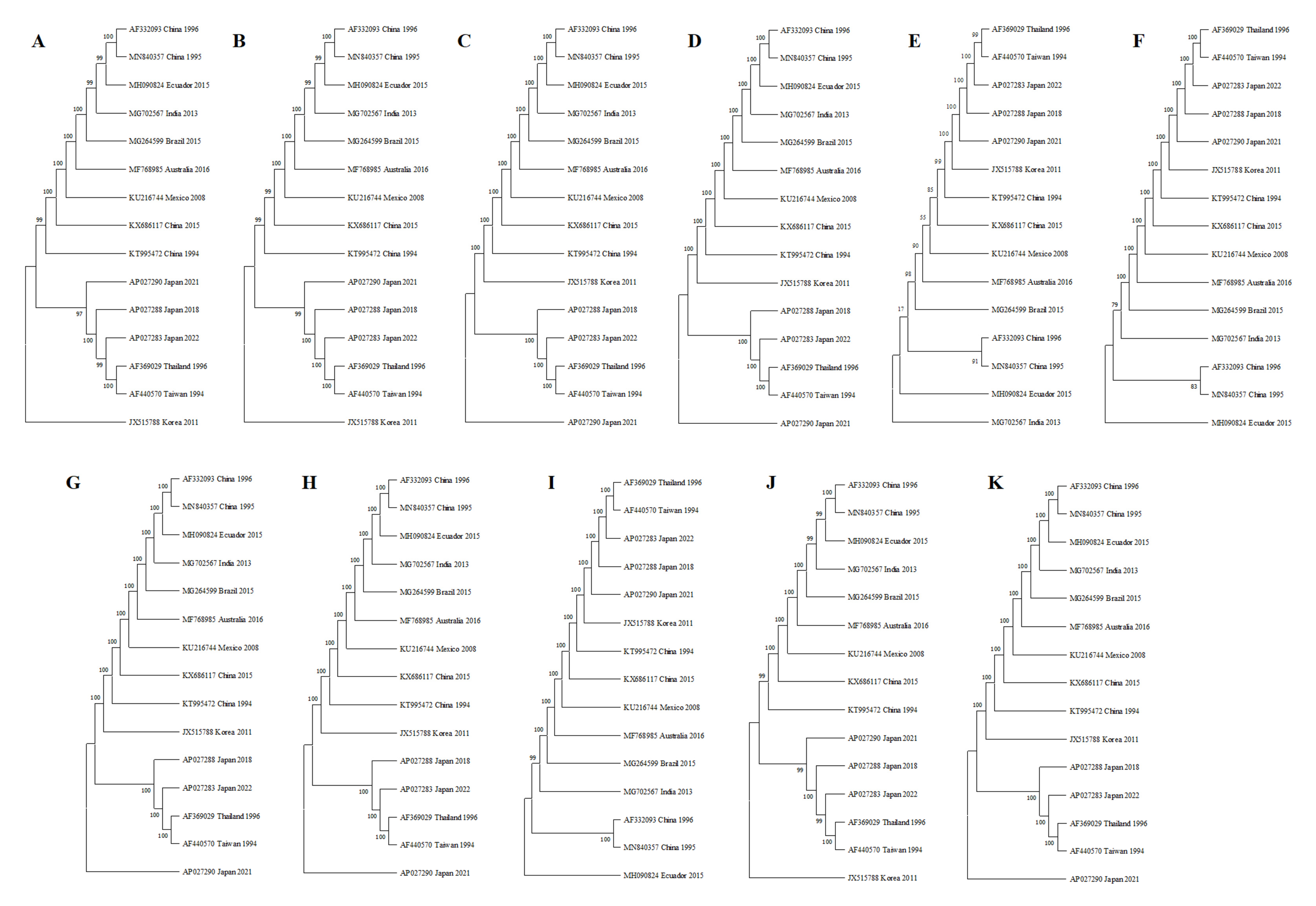
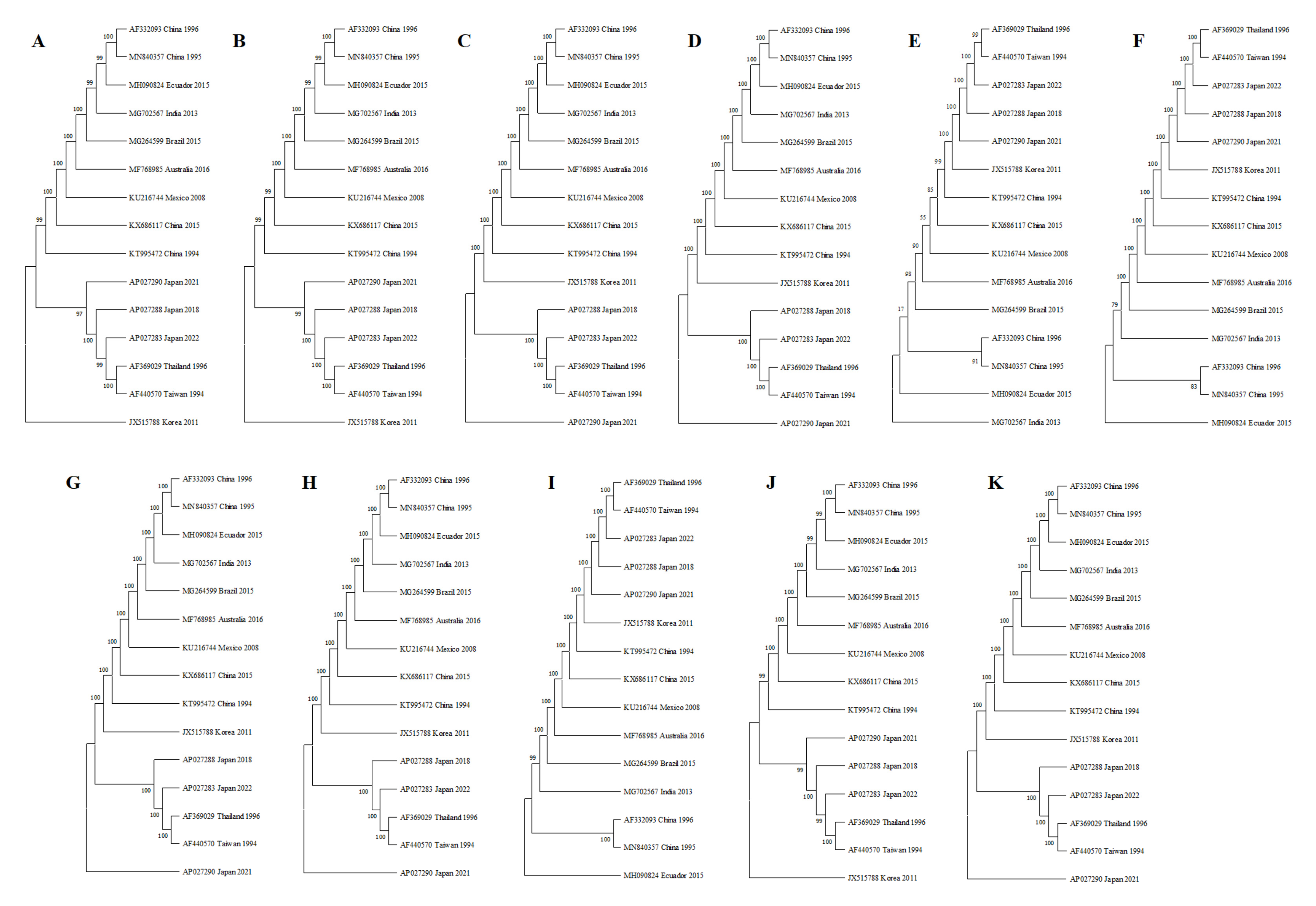
2.2.3. Phylogenetic Analysis of the Amplicons
2.2.4. Physicochemical Parameters
3. Results
3.1. Primer Mapping
3.2. Primer Specificity
3.3. Predicted Melt Curves of Amplicons
3.4. Phylogenetic Analysis of the Amplicons
3.5. Physicochemical Properties of Primers
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dey, B.K.; Dugassa, G.H.; Hinzano, S.M.; Bossier, P. Causative agent, diagnosis, and management of white spot disease in shrimp: A review. Rev. Aquacult. 2020, 12, 822–865. [Google Scholar] [CrossRef]
- Zhang, X.; Hu, L.H.; Song, D.W.; Hu, Y.; Chen, J. Evaluation on prevention and treatment of cuminaldehyde in culture of shrimp against white spot syndrome virus. Aquaculture 2023, 562, 738760. [Google Scholar] [CrossRef]
- Stentiford, G.D.; Neil, D.M.; Peeler, E.J.; Shields, J.D.; Small, H.J.; Flegel, T.W.; Vlak, J.M.; Jones, B.; Morado, F.; Moss, S.; et al. Disease will limit future food supply from the global crustacean fishery and aquaculture sectors. J. Invertebr. Pathol. 2012, 110, 141–157. [Google Scholar] [CrossRef] [PubMed]
- Flegel, T.W. Historic emergence, impact, and current status of shrimp pathogens in Asia. J. Invertebr. Pathol. 2012, 110, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Millard, R.S.; Ellis, R.P.; Bateman, K.S.; Bickley, L.K.; Tyler, C.R.; van Aerle, R.; Santos, E.M. How do abiotic environmental conditions influence shrimp susceptibility to disease? A critical analysis focused on White Spot Disease. J. Invertebr. Pathol. 2021, 186, 107369. [Google Scholar] [CrossRef] [PubMed]
- van Hulten, M.C.; Witteveldt, J.; Peters, S.; Kloosterboer, N.; Tarchini, R.; Fiers, M.; Sandbrink, H.; Lankhorst, R.K.; Vlak, J.M. The white spot syndrome virus DNA genome sequence. Virology 2001, 286, 7–22. [Google Scholar] [CrossRef]
- Yang, F.; He, J.; Lin, X.; Li, Q.; Pan, D.; Zhang, X.; Xu, X. Complete genome sequence of the shrimp white spot bacilliform virus. J. Virol. 2001, 75, 11811–11820. [Google Scholar] [CrossRef] [PubMed]
- Leu, J.H.; Yang, F.; Zhang, X.; Xu, X.; Kou, G.H.; Lo, C.F. Whispovirus. In Lesser-Known Large dsDNA Viruses; Van Etten, J.L., Ed.; Springer: Berlin/Heidelberg, Germany, 2009; pp. 197–227. [Google Scholar]
- Rozenberg, A.; Brand, P.; Rivera, N.; Leese, F.; Schubart, C.D. Characterization of fossilized relatives of the White Spot Syndrome Virus in genomes of decapod crustaceans. BMC Evol. Biol. 2015, 15, 142. [Google Scholar] [CrossRef]
- Desrina; Prayitno, S.B.; Verdegem, M.C.J.; Verreth, J.A.J.; Vlak, J.M. White spot syndrome virus host range and impact on transmission. Rev. Aquacult. 2022, 14, 1843–1860. [Google Scholar] [CrossRef]
- Sánchez-Paz, A. White spot syndrome virus: An overview on an emergent concern. Vet. Res. 2010, 41, 43. [Google Scholar] [CrossRef]
- Poulos, B.T.; Pantoja, C.R.; Bradley-Dunlop, D.; Aguilar, J.; Lightner, D.V. Development and application of monoclonal antibodies for the detection of white spot syndrome virus of penaeid shrimp. Dis. Aquat. Organ. 2001, 47, 13–23. [Google Scholar] [CrossRef]
- Murugan, V.; Sankaran, K. Bacterial lipid modification of ICP11 and a new ELISA system applicable for WSSV infection detection. Mar. Biotechnol. 2018, 20, 375–384. [Google Scholar] [CrossRef]
- Anil, T.M.; Shankar, K.M.; Mohan, C.V. Monoclonal antibodies developed for sensitive detection and comparison of white spot syndrome virus isolates in India. Dis. Aquat. Organ. 2001, 51, 67–75. [Google Scholar] [CrossRef]
- Liu, W.; Wang, Y.T.; Tian, D.S.; Yin, Z.C.; Kwang, J. Detection of white spot syndrome virus (WSSV) of shrimp by means of monoclonal antibodies (MAbs) specific to an envelope protein (28 kDa). Dis. Aquat. Organ. 2002, 49, 11–18. [Google Scholar] [CrossRef]
- You, Z.; Nadala, E.C.; Yang, J.; van Hulten, M.C.; Loh, P.C. Production of polyclonal antiserum specific to the 27.5 kDa envelope protein of white spot syndrome virus. Dis. Aquat. Organ. 2002, 51, 77–80. [Google Scholar] [CrossRef]
- Sithigorngul, P.; Rukpratanporn, S.; Chaivisuthangkura, P.; Sridulyakul, P.; Longyant, S. Simultaneous and rapid detection of white spot syndrome virus and yellow head virus infection in shrimp with a dual immunochromatographic strip test. J. Virol. Methods 2011, 173, 85–91. [Google Scholar] [CrossRef]
- Durand, S.V.; Lightner, D.V. Quantitative real time PCR for the measurement of white spot syndrome virus in shrimp. J. Fish Dis. 2002, 25, 381–389. [Google Scholar] [CrossRef]
- Fouzi, M.; Shariff, M.; Omar, A.R.; Yusoff, F.M.; Tan, S.W. TaqMan real-time PCR assay for relative quantification of white spot syndrome virus infection in Penaeus monodon Fabricius exposed to ammonia. J. Fish Dis. 2010, 33, 931–938. [Google Scholar] [CrossRef]
- Kim; Kim; Sohn; Sim; Park; Heo; Lee; Lee; Jun; Jang. Development of a polymerase chain reaction (PCR) procedure for the detection of baculovirus associated with white spot syndrome (WSBV) in penaeid shrimp. J. Fish Dis. 1998, 21, 11–17. [Google Scholar] [CrossRef]
- Leal, C.A.G.; Carvalho, A.F.; Leite, R.C.; Figueiredo, H.C.P. Development of duplex real-time PCR for the detection of WSSV and PstDV1 in cultivated shrimp. BMC Vet. Res. 2014, 10, 150. [Google Scholar] [CrossRef]
- Lo, C.F.; Ho, C.H.; Peng, S.E.; Chen, C.H.; Hsu, H.C.; Chiu, Y.L.; Chang, C.F.; Liu, K.F.; Su, M.S.; Wang, C.H.; et al. White spot syndrome baculovirus (WSBV) detected in cultured and captured shrimp, crabs, and other arthropods. Dis. Aquat. Organ. 1996, 27, 215–225. [Google Scholar] [CrossRef]
- Mendoza-Cano, F.; Sánchez-Paz, A. Development and validation of a quantitative real-time polymerase chain assay for universal detection of the White Spot Syndrome Virus in marine crustaceans. Virol. J. 2013, 10, 186. [Google Scholar] [CrossRef] [PubMed]
- Natividad, K.D.T.; Nomura, N.; Matsumura, M. Detection of White spot syndrome virus DNA in pond soil using a 2-step nested PCR. J. Virol. Methods 2008, 149, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.; Zhang, Y.; Sha, X.; Wang, J.; Li, J.; Dong, P.; Liang, X. Highly sensitive detection of low-abundance White Spot Syndrome Virus by a pre-amplification PCR method. J. Microbiol. Biotechnol. 2017, 27, 471–479. [Google Scholar] [CrossRef] [PubMed]
- Caipang, C.M.A.; Sibonga, M.F.J.; Geduspan, J.S.; Amar, M.J.A. An optimized loop-mediated isothermal amplification (LAMP) assay for the detection of white spot syndrome virus (WSSV) among cultured shrimps in the Philippines. J. Anim. Plant Sci. 2012, 22, 927–932. [Google Scholar]
- Chou, P.H.; Lin, Y.C.; Teng, P.H.; Chen, C.L.; Lee, P.Y. Real-time target-specific detection of loop-mediated isothermal amplification for white spot syndrome virus using fluorescence energy transfer-based probes. J. Virol. Methods 2011, 173, 67–74. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Xu, H. Development of a multiplex loop-mediated isothermal amplification (mLAMP) method for the simultaneous detection of white spot syndrome virus and infectious hypodermal and hematopoietic necrosis virus in penaeid shrimp. Aquaculture 2011, 311, 94–99. [Google Scholar] [CrossRef]
- Jaroenram, W.; Kiatpathomchai, W.; Flegel, T.W. Rapid and sensitive detection of white spot syndrome virus by loop-mediated isothermal amplification combined with a lateral flow dipstick. Mol. Cel. Probes 2009, 23, 65–70. [Google Scholar] [CrossRef]
- Kono, T.; Savan, R.; Sakai, M.; Itami, T. Detection of white spot syndrome virus in shrimp by loop-mediated isothermal amplification. J. Virol. Methods 2004, 115, 59–65. [Google Scholar] [CrossRef]
- Bai, Y.; He, L.; Sun, M.; Zhou, X.; Xu, Z. Dark-field visual counting of white spot syndrome virus using gold nanoparticle probe. Aquaculture 2023, 562, 738797. [Google Scholar] [CrossRef]
- Etedali, P.; Behbahani, M.; Mohabatkar, H.; Dini, G. Field-usable aptamer-gold nanoparticles-based colorimetric sensor for rapid detection of white spot syndrome virus in shrimp. Aquaculture 2022, 548, 737628. [Google Scholar] [CrossRef]
- WOAH. Infection with White Spot Syndrome Virus. In Manual of Diagnostic Tests for Aquatic Animals; World Organisation for Animal Health: Paris, France, 2023; Chapter 2.2.8; pp. 1–17. Available online: https://www.woah.org/fileadmin/Home/eng/Health_standards/aahm/current/2.2.08_WSSV.pdf (accessed on 15 May 2023).
- Claydon, K.; Cullen, B.; Owens, L. OIE white spot syndrome virus PCR gives false-positive results in Cherax quadricarinatus. Dis. Aquat. Organ. 2004, 62, 265–268. [Google Scholar] [CrossRef] [PubMed]
- De Filippis, I.; de Azevedo, A.C.; de Oliveira Lima, I.; Ramos, N.F.L.; de Andrade, C.F.; de Almeida, A.E. Accurate, fast and cost-effective simultaneous detection of bacterial meningitis by qualitative PCR with high-resolution melting. BioTechniques 2023, 74, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Bustin, S.; Huggett, J. qPCR primer design revisited. Biomol. Detect. Quantif. 2017, 14, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Kalendar, R.; Khassenov, B.; Ramankulov, Y.; Samuilova, O.; Ivanov, K.I. FastPCR: An in silico tool for fast primer and probe design and advanced sequence analysis. Genomics 2017, 109, 312–319. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.H.; Lo, C.F.; Leu, J.H.; Chou, C.M.; Yeh, P.Y.; Chou, H.Y.; Tung, M.C.; Chang, C.F.; Su, M.S.; Kou, G.H. Purification and genomic analysis of baculovirus associated with white spot syndrome (WSBV) of Penaeus monodon. Dis. Aquat. Organ. 1995, 23, 239–242. [Google Scholar] [CrossRef]
- Davis, M.W.; Jorgensen, E.M. ApE, A plasmid editor: A freely available DNA manipulation and visualization program. Front. Bioinformatics 2022, 2, 818619. [Google Scholar] [CrossRef]
- Nunan, L.M.; Lightner, D.V. Optimized PCR assay for detection of white spot syndrome virus (WSSV). J. Virol. Methods 2011, 171, 318–321. [Google Scholar] [CrossRef]
- Jang, I.K.; Meng, X.H.; Seo, H.C.; Cho, Y.R.; Kim, B.R.; Ayyaru, G.; Kim, J.S. A TaqMan real-time PCR assay for quantifying white spot syndrome virus (WSSV) infections in wild broodstock and hatchery-reared postlarvae of fleshy shrimp, Fenneropenaeus chinensis. Aquaculture 2009, 287, 40–45. [Google Scholar] [CrossRef]
- Tang, K.F.J.; Lightner, D.V. Quantification of white spot syndrome virus DNA through a competitive polymerase chain reaction. Aquaculture 2000, 189, 11–21. [Google Scholar] [CrossRef]
- Kimura, T.; Yamano, K.; Nakano, H.; Momoyama, K.; Hiraoka, M.; Inouye, K. Detection of Penaeid Rod-shaped DNA Virus (PRDV) by PCR. Fish Pathol. 1996, 31, 93–98. [Google Scholar] [CrossRef]
- Yuan, L.; Zhang, X.; Chang, M.; Jia, C.; Hemmingsen, S.M.; Dai, H. A new fluorescent quantitative PCR-based in vitro neutralization assay for white spot syndrome virus. J. Virol. Methods 2007, 146, 96–103. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F.; Quan, H. A new method for quantifying white spot syndrome virus: Experimental challenge dose using TaqMan real-time PCR assay. J. Virol. Methods 2012, 184, 121–124. [Google Scholar] [CrossRef]
- Sivakumar, S.; Vimal, S.; Abdul Majeed, S.; Santhosh Kumar, S.; Taju, G.; Madan, N.; Rajkumar, T.; Thamizhvanan, S.; Shamsudheen, K.V.; Scaria, V.; et al. A new strain of white spot syndrome virus affecting Litopenaeus vannamei in Indian shrimp farms. J. Fish Dis. 2018, 41, 1129–1146. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Coulouris, G.; Zaretskaya, I.; Cutcutache, I.; Rozen, S.; Madden, T.L. Primer-BLAST: A tool to design target-specific primers for polymerase chain reaction. BMC Bioinform. 2012, 13, 134. [Google Scholar] [CrossRef] [PubMed]
- Dwight, Z.; Palais, R.; Wittwer, C.T. uMELT: Prediction of high-resolution melting curves and dynamic melting profiles of PCR products in a rich web application. Bioinformatics 2011, 27, 1019–1020. [Google Scholar] [CrossRef]
- SantaLucia, J. A unified view of polymer, dumbbell, and oligonucleotide DNA nearest-neighbor thermodynamics. Proc. Natl. Acad. Sci. USA 1998, 95, 1460–1465. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Sela, I.; Ashkenazy, H.; Katoh, K.; Pupko, T. GUIDANCE2: Accurate detection of unreliable alignment regions accounting for the uncertainty of multiple parameters. Nucleic Acids Res. 2015, 43, W7–W14. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Hasegawa, M.; Kishino, H.; Yano, T. Dating of the human-ape splitting by a molecular clock of mitochondrial DNA. J. Mol. Evol. 1985, 22, 160–174. [Google Scholar] [CrossRef] [PubMed]
- Jukes, T.H.; Cantor, C.R. Evolution of protein molecules. In Mammalian Protein Metabolism; Munro, H.N., Ed.; Academic Press: New York, NY, USA, 1969; pp. 21–132. [Google Scholar]
- Tamura, K. Estimation of the number of nucleotide substitutions when there are strong transition-transversion and G+C-content biases. Mol. Biol. Evol. 1992, 9, 678–687. [Google Scholar] [CrossRef] [PubMed]
- Kibbe, W.A. OligoCalc: An online oligonucleotide properties calculator. Nucleic Acids Res. 2007, 35, W43–W46. [Google Scholar] [CrossRef] [PubMed]
- Dang, L.T.; Koyama, T.; Shitara, A.; Kondo, H.; Aoki, T.; Hirono, I. Involvement of WSSV–shrimp homologs in WSSV infectivity in kuruma shrimp: Marsupenaeus japonicus. Antivir. Res. 2010, 88, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Hasson, K.W.; Fan, Y.; Reisinger, T.; Venuti, J.; Varner, P.W. White-spot syndrome virus (WSSV) introduction into the Gulf of Mexico and Texas freshwater systems through imported, frozen bait-shrimp. Dis. Aquat. Organ. 2006, 71, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H. Alignment of BLAST high-scoring segment pairs based on the longest increasing subsequence algorithm. Bioinformatics 2003, 19, 1391–1396. [Google Scholar] [CrossRef]
- Boyle, B.; Dallaire, N.; MacKay, J. Evaluation of the impact of single nucleotide polymorphisms and primer mismatches on quantitative PCR. BMC Biotechnol. 2009, 9, 75. [Google Scholar] [CrossRef]
- De Long, S.K.; Kinney, K.A.; Kirisits, M.J. qPCR assays to quantify genes and gene expression associated with microbial perchlorate reduction. J. Microbiol. Methods 2010, 83, 270–274. [Google Scholar] [CrossRef]
- Ledeker, B.M.; De Long, S.K. The effect of multiple primer-template mismatches on quantitative PCR accuracy and development of a multi-primer set assay for accurate quantification of pcrA gene sequence variants. J. Microbiol. Methods 2013, 94, 224–231. [Google Scholar] [CrossRef]
- Hongoh, Y.; Yuzawa, H.; Ohkuma, M.; Kudo, T. Evaluation of primers and PCR conditions for the analysis of 16S rRNA genes from a natural environment. FEMS Microbiol. Lett. 2003, 221, 299–304. [Google Scholar] [CrossRef]
- Tsai, J.M.; Wang, H.C.; Leu, J.H.; Wang, A.H.; Zhuang, Y.; Walker, P.J.; Kou, G.H.; Lo, C.F. Identification of the nucleocapsid, tegument, and envelope proteins of the shrimp white spot syndrome virus virion. J. Virol. 2006, 80, 3021–3029. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.; Xu, L.; Yang, F. Proteomic analysis of the major envelope and nucleocapsid proteins of white spot syndrome virus. J. Virol. 2006, 80, 10615–10623. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Xu, L.; Li, H.; Qi, Y.P.; Yang, F. Four major envelope proteins of white spot syndrome virus bind to form a complex. J. Virol. 2009, 83, 4709–4712. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Li, F.; Han, Y.; Xu, L.; Yang, F. VP24 is a chitin-binding protein involved in White Spot Syndrome Virus infection. J. Virol. 2015, 90, 842–850. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.S.; Liu, W.J.; Lee, C.C.; Chou, T.L.; Lee, Y.T.; Wu, T.S.; Huang, J.Y.; Huang, W.T.; Lee, T.L.; Kou, G.H.; et al. A 3D model of the membrane protein complex formed by the White Spot Syndrome Virus structural proteins. PLoS ONE 2010, 5, e10718. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.J.; Tang, S.L.; Chang, Y.C.; Wang, H.C.; Ng, T.H.; Garmann, R.F.; Chen, Y.W.; Huang, J.Y.; Kumar, R.; Chang, S.H.; et al. Multiple nucleocapsid structural forms of shrimp White Spot Syndrome Virus suggests a novel viral morphogenetic pathway. Int. J. Mol. Sci. 2023, 24, 7525. [Google Scholar] [CrossRef] [PubMed]
- Green, S.J.; Venkatramanan, R.; Naqib, A. Deconstructing the polymerase chain reaction: Understanding and correcting bias associated with primer degeneracies and primer-template mismatches. PLoS ONE 2015, 10, e0128122. [Google Scholar] [CrossRef] [PubMed]
- Whiley, D.M.; Sloots, T.P. Sequence variation in primer targets affects the accuracy of viral quantitative PCR. J. Clin. Virol. 2005, 34, 104–107. [Google Scholar] [CrossRef]
- Robalino, J.; Payne, C.; Parnell, P.; Shepard, E.; Grimes, A.C.; Metz, A.; Prior, S.; Witteveldt, J.; Vlak, J.M.; Gross, P.S.; et al. Inactivation of White Spot Syndrome Virus (WSSV) by normal rabbit serum: Implications for the role of the envelope protein VP28 in WSSV infection of shrimp. Virus Res. 2006, 118, 55–61. [Google Scholar] [CrossRef]
- Sritunyalucksana, K.; Wannapapho, W.; Lo, C.F.; Flegel, T.W. PmRab7 is a VP28-binding protein involved in white spot syndrome virus infection in shrimp. J. Virol. 2006, 80, 10734–10742. [Google Scholar] [CrossRef]
- van Hulten, M.C.; Witteveldt, J.; Snippe, M.; Vlak, J.M. White spot syndrome virus envelope protein VP28 is involved in the systemic infection of shrimp. Virology 2001, 285, 228–233. [Google Scholar] [CrossRef] [PubMed]
- Yi, G.; Wang, Z.; Qi, Y.; Yao, L.; Qian, J.; Hu, L. Vp28 of shrimp white spot syndrome virus is involved in the attachment and penetration into shrimp cells. J. Biochem. Mol. Biol. 2004, 37, 726–734. [Google Scholar] [CrossRef] [PubMed]
- Durand, S.; Lightner, D.V.; Redman, R.M.; Bonami, J.R. Ultrastructure and morphogenesis of White Spot Syndrome Baculovirus (WSSV). Dis. Aquat. Organ. 1997, 29, 205–211. [Google Scholar] [CrossRef]
- Liu, Q.H.; Ma, C.Y.; Chen, W.B.; Zhang, X.L.; Liang, Y.; Dong, S.L.; Huang, J. White spot syndrome virus VP37 interacts with VP28 and VP26. Dis. Aquat. Organ. 2009, 85, 23–30. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence (5′–3′) | Reference | PCR | Size (bp) |
|---|---|---|---|---|
| 146F1 | ACTACTAACTTCAGCCTATCTAG | [22] | Nested * | 1447 |
| 146R1 | TAATGCGGGTGTAATGTTCTTACGA | |||
| 146F2 | GTAACTGCCCCTTCCATCTCCA | 941 | ||
| 146R2 | TACGGCAGCTGCTGCACCTTGT | |||
| 146F2 | GTAACTGCCCCTTCCATCTCCA | [40] | One-step | 941 |
| 146R2 | TACGGCAGCTGCTGCACCTTGT | |||
| WSS1011F | TGGTCCCGTCCTCATCTCAG | [18] | qPCR * | 69 |
| WSS1079R | GCTGCCTTGCCGGAAATTA | |||
| Probe | AGCCATGAAGAATGCCGTCTATCACACA | |||
| RT-WSSV-F154 | CCAGTTCAGAATCGGACGTT | [41] | qPCR | 154 |
| RT-WSSV-R154 | AAAGACGCCTACCCTGTTGA | |||
| RT-WSSV-TP154 Probe | TCCATAGTTCCTGGTTTGTAATGTGCCG | |||
| WSSV341F | TGGCTACATCTGCATTGCTC | [42] | One-step | 341 |
| WSSV341R | TAGAGACGTGGCTGGAGAGG | |||
| P1 | ATCATGGCTGCTTCACAGAC | [43] | Nested | 982 |
| P2 | GGCTGGAGAGGACAAGACAT | |||
| P3 | TCTTCATCAGATGCTACTGC | 570 | ||
| P4 | TAACGCTATCCAGTATCACG | |||
| FQ-P3 (up) | AAGCATCGTGGAGACTCTTGC | [44] | qPCR | 129 |
| FQ-P4 (down) | GAAGATTCGCCGCTCATACC | |||
| WSSV-RT1 | TTGGTTTCATGCCCGAGATT | [45] | qPCR | 154 (57) |
| WSSV-RT2 | CCTTGGTCAGCCCCTTGA | |||
| Probe | TGCTGCCGTCTCCAA | |||
| Forward | CCCACACAGACAATATCGAGAC | [46] | qPCR | 109 |
| Reverse | TCGCTGTCAAAGGACACATC | |||
| Probe TP109 | TTCCTGTGACTGCTGAGGTTGGAT | |||
| VP28-140Fw | AGGTGTGGAACAACACATCAAG | [23] | qPCR | 141 |
| VP28-140Rv | TGCCAACTTCATCCTCATCA |
| GenBank Accession Number | Geographic Location |
|---|---|
| AF332093 | China |
| AF369029 | Thailand |
| AF440570 | Taiwan |
| AP027283 | Japan |
| AP027288 | Japan |
| AP027290 | Japan |
| JX515788 | South Korea |
| KT995472 | China |
| KU216744 | Mexico |
| KX686117 | China |
| MF768985 | Australia |
| MG264599 | Brazil |
| MG702567 | India |
| MH090824 | Ecuador |
| MN840357 | China |
| Primer | Number of Blast Hits Analyzed | Filtered Hits | Mismatches | Number of Nonspecific Amplifications |
|---|---|---|---|---|
| 146F1-146R1 | 16,581 | 45 | No | 0 |
| 146F2-146R2 | 30,191 | 56 | No | 0 |
| WSS1011F-WSS1079R | 12,224 | 81 | Yes | 0 |
| RT-WSSV-F154-RT-WSSV-R154 | 9819 | 61 | Yes | 0 |
| WSSV341F-WSSV341R | 13,699 | 47 | No | 0 |
| P1-P2 | 21,128 | 47 | No | 0 |
| P3-P4 | 20,724 | 52 | Yes | 0 |
| FQ-P3-FQ-P4 | 9141 | 44 | No | 0 |
| WSSV-RT1-WSSV-RT2 | 7857 | 45 | Yes | 0 |
| F-W | 16,758 | 109 | Yes | 0 |
| VP28-140Fw-VP28-140Rv | 36,827 | 126 | No | 0 |
| Primer | GC (%) | Length (bp) | Tm (°C) | ∆G (Kcal/mol) | SD | HF | SA |
|---|---|---|---|---|---|---|---|
| 146F1 | 39 | 23 | 52.67 | −26.0 | - | - | - |
| 146R1 | 40 | 25 | 56.74 | −32.0 | - | - | - |
| 146F2 | 55 | 22 | 56.22 | −29.1 | - | - | - |
| 146R2 | 59 | 22 | 62.57 | −32.3 | ✓ | ✓ | ✓ |
| WSS1011F | 60 | 20 | 57.38 | −27.2 | - | - | - |
| WSS1079R | 53 | 19 | 56.04 | −25.5 | - | - | - |
| RT-WSSV-F154 | 50 | 20 | 54.82 | −26.2 | - | - | - |
| RT-WSSV-R154 | 50 | 20 | 55.16 | −26.3 | - | - | - |
| WSSV341F | 50 | 20 | 56.43 | −25.4 | - | - | - |
| WSSV341R | 60 | 20 | 58.46 | −27.2 | - | - | - |
| P1 | 50 | 20 | 54.94 | −25.2 | - | - | - |
| P2 | 55 | 20 | 55.21 | −25.8 | - | - | - |
| P3 | 45 | 20 | 53.29 | −23.8 | - | - | - |
| P4 | 45 | 20 | 52.75 | −24.6 | - | - | - |
| FQ-P3 (up) | 52 | 21 | 59.38 | −27.9 | - | - | ✓ |
| FQ-P4 (down) | 55 | 20 | 57.63 | −26.7 | - | - | - |
| WSSV-RT1 | 45 | 20 | 53.72 | −25.6 | - | - | - |
| WSSV-RT2 | 61 | 18 | 54.0 | −24.3 | - | - | - |
| Forward | 50 | 22 | 55.45 | −27.7 | - | - | ✓ |
| Reverse | 50 | 20 | 54.83 | −25.8 | - | - | - |
| VP28-140Fw | 45 | 22 | 53.39 | −27.4 | ✓ | - | - |
| VP28-140Rv | 45 | 20 | 52.25 | −24.7 | - | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sánchez-Paz, A.; Encinas-García, T.; Mendoza-Cano, F. In Silico Evaluation of the PCR Performance of Different Tests for Detection of WSSV. Fishes 2024, 9, 5. https://doi.org/10.3390/fishes9010005
Sánchez-Paz A, Encinas-García T, Mendoza-Cano F. In Silico Evaluation of the PCR Performance of Different Tests for Detection of WSSV. Fishes. 2024; 9(1):5. https://doi.org/10.3390/fishes9010005
Chicago/Turabian StyleSánchez-Paz, Arturo, Trinidad Encinas-García, and Fernando Mendoza-Cano. 2024. "In Silico Evaluation of the PCR Performance of Different Tests for Detection of WSSV" Fishes 9, no. 1: 5. https://doi.org/10.3390/fishes9010005
APA StyleSánchez-Paz, A., Encinas-García, T., & Mendoza-Cano, F. (2024). In Silico Evaluation of the PCR Performance of Different Tests for Detection of WSSV. Fishes, 9(1), 5. https://doi.org/10.3390/fishes9010005