
Molecular Analysis of Two Endemic Squalius Species: Evidence for Intergeneric Introgression among Cyprinids and Conservation Issues

Abstract
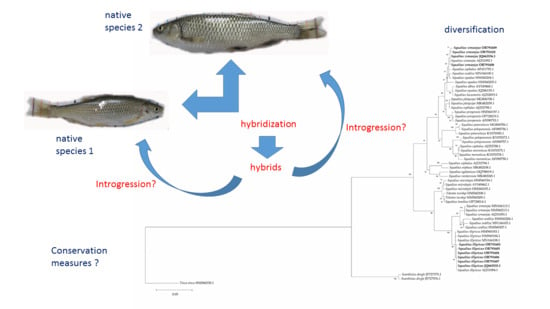
:
1. Introduction
2. Materials and Methods
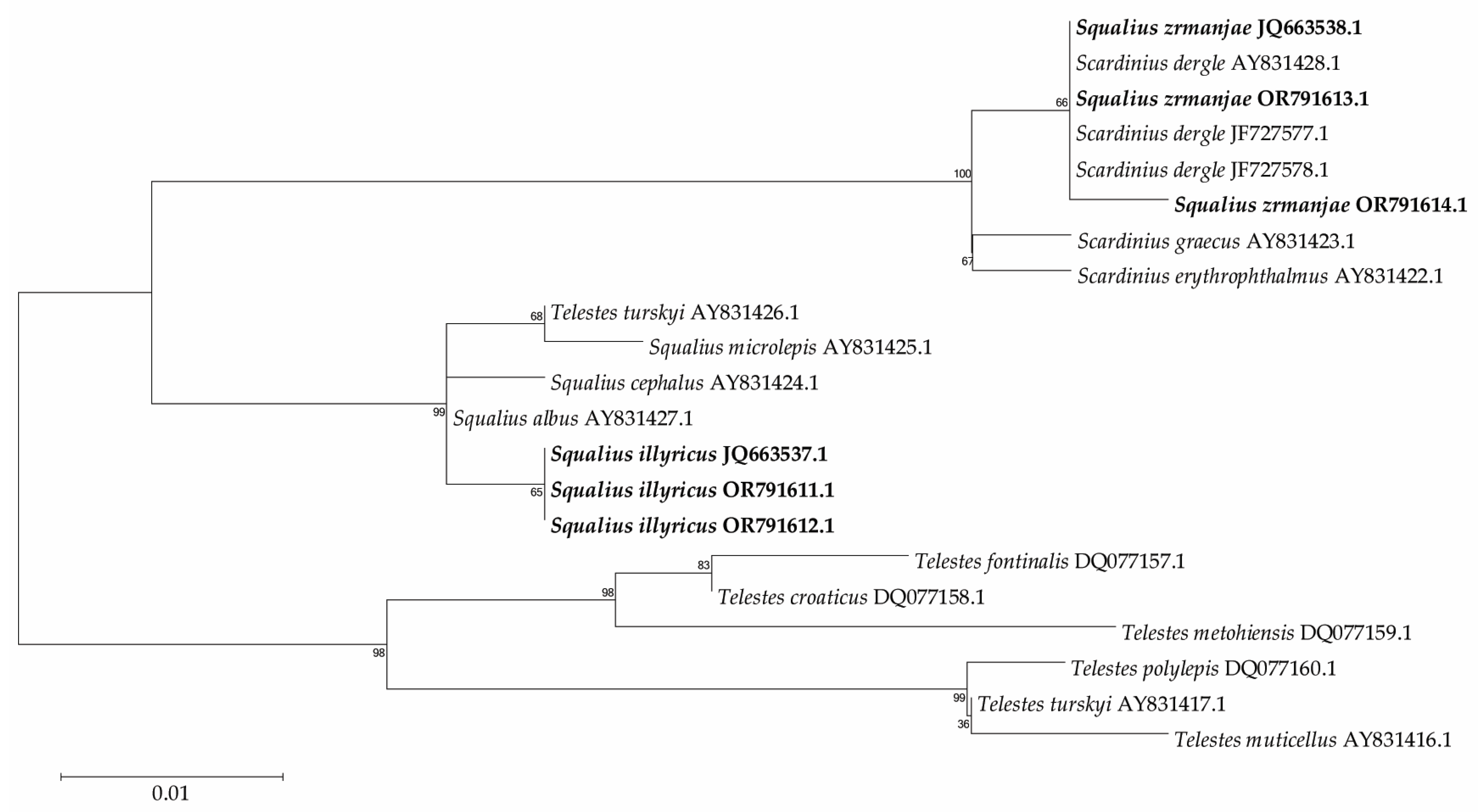
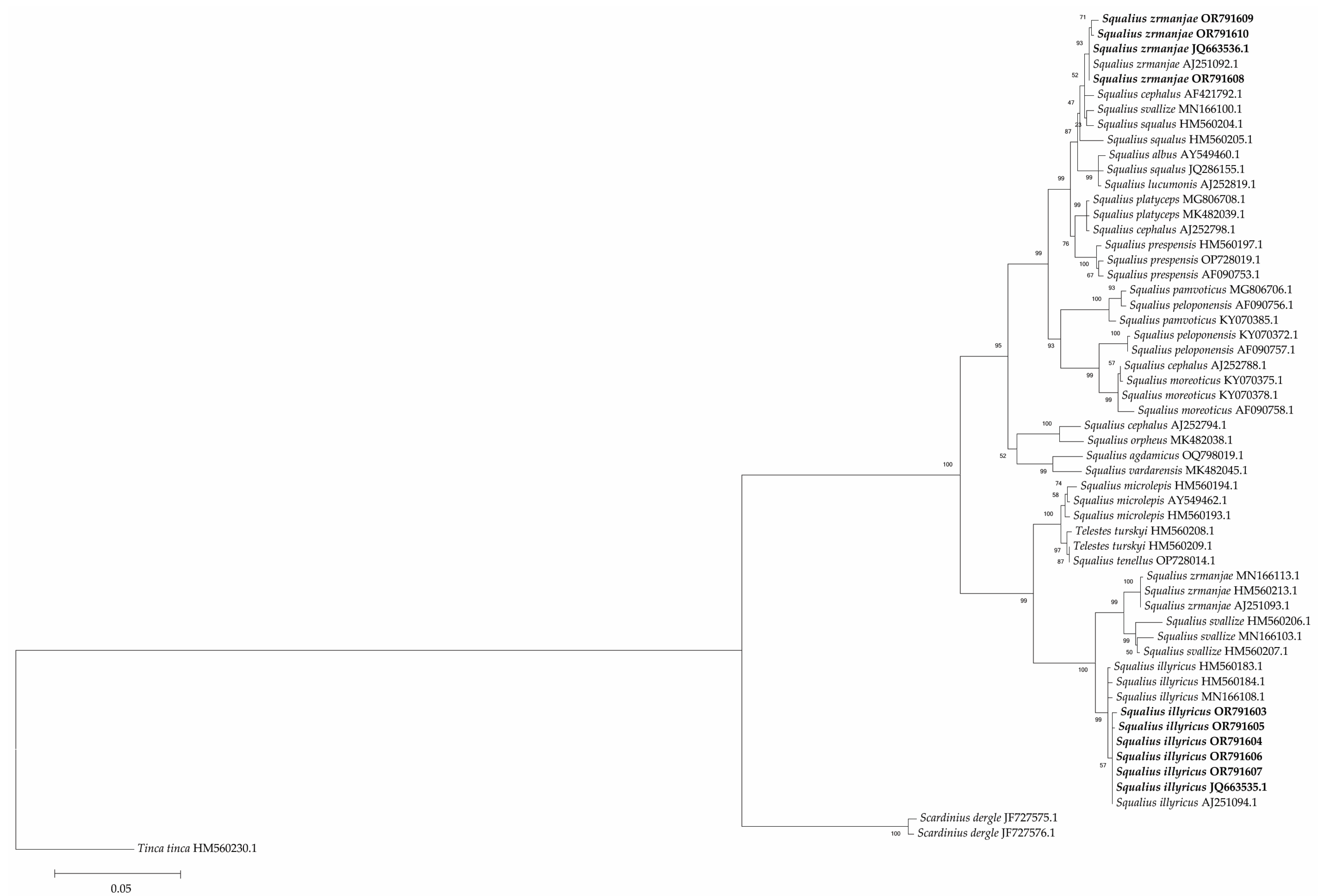
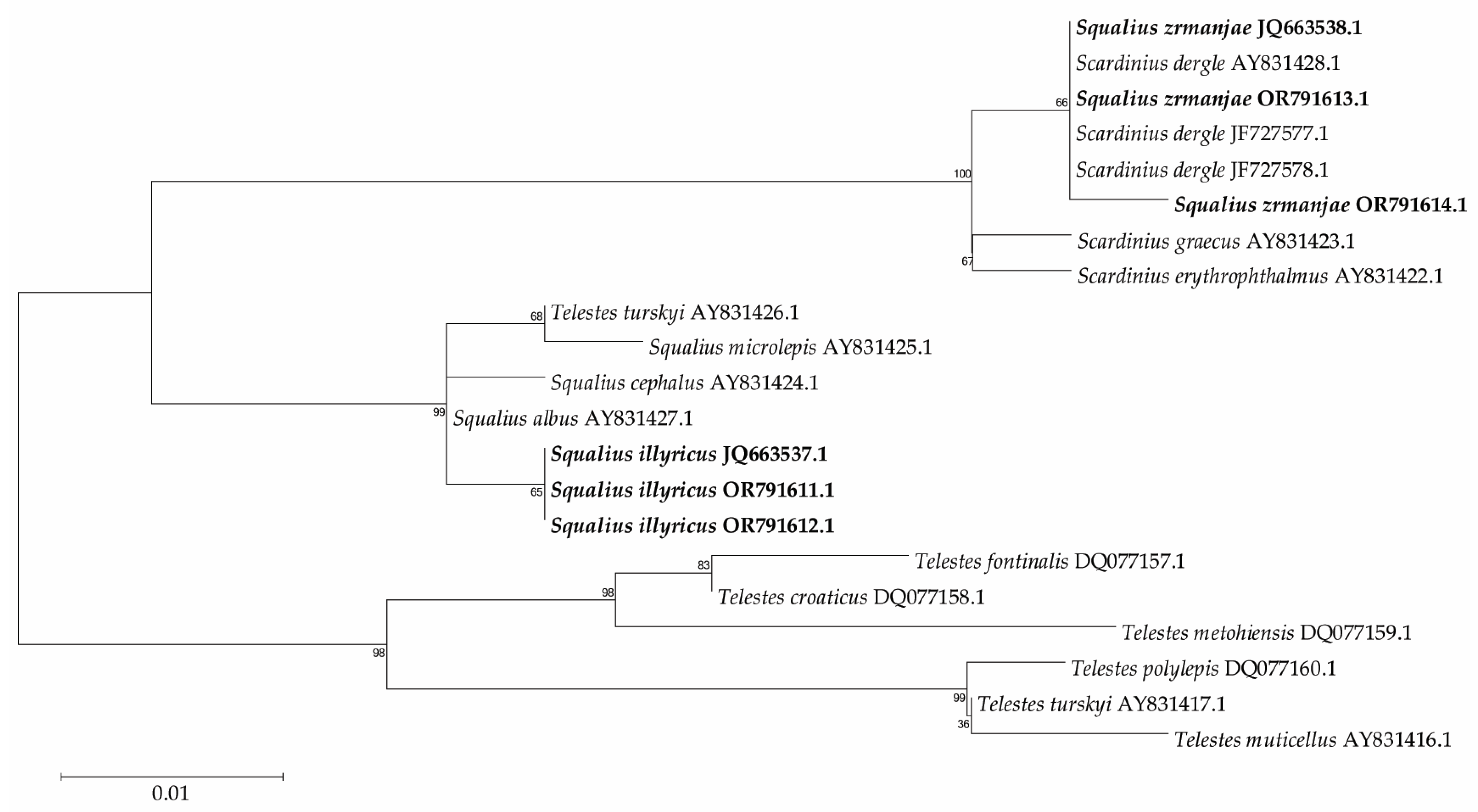
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kottelat, M.; Freyhof, J. Handbook of European Freshwater Fishes; Kottelat: Cornol, Switzerland; Freyhof: Berlin, Germany, 2007; ISBN 978-2-8399-0298-4. [Google Scholar]
- Ćaleta, M.; Buj, I.; Mrakovčić, M.; Mustafić, P.; Zanella, D.; Marčić, Z.; Duplić, A.; Mihinjač, T.; Katavić, I. Endemic Fishes of Croatia; Agencija za zaštitu okoliša: Zagreb, Croatia, 2015; ISBN 978-953-7582-14-2. [Google Scholar]
- Mrakovčić, M.; Brigić, A.; Buj, I.; Ćaleta, M.; Mustafić, P.; Zanella, D. Red Book of Freshwater Fish of Croatia; Ministry of Culture, State Institute for Nature Protection: Zagreb, Croatia, 2006; ISBN 953-7169-21-9. [Google Scholar]
- Ćaleta, M.; Marčić, Z.; Buj, I.; Zanella, D.; Mustafić, P.; Duplić, A.; Horvatić, S. A Review of Extant Croatian Freshwater Fish and Lampreys—Annotated list and distribution. Ribar. Croat. J. Fish. 2019, 77, 137–234. [Google Scholar] [CrossRef]
- Doadrio, J.; Carmona, J.A. Genetic divergence in Greek populations of the genus Leuciscus and its evolutionary biogeographical implications. J. Fish. Biol. 1998, 53, 591–613. [Google Scholar] [CrossRef]
- Durand, J.D.; Bianco, P.G.; Laroche, J.; Gilles, A. Insight into the origin of the endemic Mediterranean ichthyofauna: Phylogeography of the Chondrostoma genus (Teleostei, Cyprinidae). J. Hered. 2003, 94, 315–328. [Google Scholar] [CrossRef] [PubMed]
- Ketmaier, V.; Bianco, P.G.; Cobolli, M.; De Matthaeis, E. Genetic differentiation and biogeography in southern European populations of the genus Scardinius (Pisces, Cyprinidae) based on allozyme data. Zool. Scr. 2003, 32, 13–22. [Google Scholar] [CrossRef]
- Milošević, D.; Winkler, K.A.; Marić, D.; Weiss, S. Genotypic and phenotypic evaluation of Rutilus spp. from Skadar, Ohrid and Prespa lakes supports revision of endemic as well as taxonomic status of several taxa. J. Fish. Biol. 2011, 79, 1094–1110. [Google Scholar] [CrossRef] [PubMed]
- Valić, D.; Vardić Smrzlić, I.; Kapetanović, D.; Teskeredžić, Z.; Pleše, B.; Teseredžić, E. Identification, phylogenetic relationships and a new maximum size of two rudd populations (Scardinius, Cyprinidae) from the Adriatic Sea drainage, Croatia. Biologia 2013, 68, 539–545. [Google Scholar] [CrossRef]
- Tan, M.; Armbruster, J.W. Phylogenetic classification of extant genera of fishes of the order Cypriniformes (Teleostei: Ostariophysi). Zootaxa 2018, 4476, 6–39. [Google Scholar] [CrossRef]
- Schönhuth, S.; Vukić, J.; Šanda, R.; Yang, L.; Mayden, R.L. Phylogenetic relationships and classification of the Holarctic family Leuciscidae (Cypriniformes: Cyprinoidei). Mol. Phylogenet. Evol. 2018, 127, 781–799. [Google Scholar] [CrossRef]
- Avise, J.C. Molecular Markers, Natural History and Evolution; Chapman & Hall: New York, NY, USA, 1994. [Google Scholar]
- Zardoya, R.; Meyer, A. Phylogenetic performance of mitochondrial protein-coding genes in resolving relationships among vertebrates. Mol. Biol. Evol. 1996, 13, 933–942. [Google Scholar] [CrossRef]
- Briolay, J.; Galtier, N.; Brito, R.M.; Bouvet, Y. Molecular phylogeny of Cyprinidae inferred from cytochrome b DNA sequences. Mol. Phylogenet. Evol. 1998, 9, 100–108. [Google Scholar] [CrossRef]
- Mallet, J. Hybridization as an invasion of the genome. Trends Ecol. Evol. 2005, 20, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Durand, J.D.; Ünlü, E.; Doadrio, I.; Pipoyan, S.; Templeton, A.R. Origin, radiation, dispersion and allopatric hybridization in the chub Leuciscus cephalus. Proc. Royal Soc. B 2000, 267, 1687–1697. [Google Scholar] [CrossRef] [PubMed]
- Scribner, K.T.; Page, K.S.; Bartron, M. Hybridization in freshwater fishes: A review of case studies and cytonuclear methods of biological inference. Rev. Fish. Biol. Fish. 2001, 10, 293–323. [Google Scholar] [CrossRef]
- Salzburger, W.; Brandstätter, A.; Gilles, A.; Parson, W.; Hempel, M.; Sturmbauer, C.; Meyer, A. Phylogeography of the vairone (Leuciscus souffia, Risso 1826) in central Europe. Mol. Ecol. 2003, 12, 2371–2386. [Google Scholar] [CrossRef] [PubMed]
- Yakovlev, V.N.; Slyn’ko, Y.V.; Grechanov, I.G.; Krysanov, E.Y. Distant hybridization in fish. J. Ichthyol. 2000, 40, 298–311. [Google Scholar]
- Hayden, B.; Pulcini, D.; Kelly-Quinn, M.; O’Grady, M.; Caffrey, J.; McGrath, A.; Mariani, S. Hybridisation between two cyprinid fishes in a novel habitat: Genetics, morphology and life-history traits. BMC Evol. Biol. 2010, 10, 169. [Google Scholar] [CrossRef]
- Ramoejane, M.; Gouws, G.; Swartz, E.R.; Sidlauskas, B.L.; Weyl, O.L.F. Molecular and morphological evidence reveals hybridisation between two endemic cyprinid fishes. J. Fish. Biol. 2020, 96, 1234–1250. [Google Scholar] [CrossRef]
- Freyhof, J.; Lieckfeldt, D.; Bogutskaya, N.G.; Pitra, C.; Ludwig, A. Phylogenetic position of the Dalmatian genus Phoxinellus and description of the newly proposed genus Delminichthys (Teleostei: Cyprinidae). Mol. Phylogenet. Evol. 2006, 38, 416–425. [Google Scholar] [CrossRef]
- Lieckfeldt, D.; Hett, A.K.; Ludwig, A.; Freyhof, J. Detection, characterization and utility of a new highly variable nuclear marker region in several species of cyprinid fishes (Cyprinidae). Eur. J. Wildl. Res. 2006, 52, 63–65. [Google Scholar] [CrossRef]
- Vadas, R.L., Jr.; Orth, D.J. Species associations and habitat use of stream fishes: The effects of unaggregated-data analysis. J. Freshw. Ecol. 1997, 12, 27–37. [Google Scholar] [CrossRef]
- Stauffer, J.R., Jr.; Hocutt, C.H.; Denoncourt, R.F. Status and distribution of the hybrid Nocomis micropogon X Rhinichthys cataractae, with a discussion of hybridization as a viable mode of vertebrate speciation. Am. Midl. Nat. 1979, 101, 355–365. [Google Scholar] [CrossRef]
- Goodfellow, W.L., Jr.; Hocutt, C.H.; Morgan, R.P., II; Stauffer, J.R., Jr. Biochemical assessment of the taxonomic status of “Rhinichthys bowersi” (Pisces: Cyprinidae). Copeia 1984, 3, 652–659. [Google Scholar] [CrossRef]
- Stauffer, J.R., Jr.; Hocutt, C.H.; Mayden, R.L. Pararhinichthys, a new monotypic genus of minnows (Teleostei: Cyprinidae) of hybrid origin from eastern North America. Ichthyol. Explor. Freshw. 1997, 7, 327–336. [Google Scholar]
- Allendorf, F.W.; Leary, R.F.; Spruell, P.; Wenburg, J.P. The problems with hybrids: Setting conservation guidelines. Trends Ecol. Evol. 2001, 16, 613–622. [Google Scholar] [CrossRef]
- Filipović Marijić, V.; Kapetanović, D.; Dragun, Z.; Valić, D.; Krasnići, N.; Redžović, Z.; Grgić, I.; Žunić, J.; Kružlicová, D.; Nemeček, P.; et al. Influence of technological and municipal wastewaters on vulnerable karst riverine system, Krka River in Croatia. Environ. Sci. Pollut. Res. 2018, 25, 4715–4727. [Google Scholar] [CrossRef] [PubMed]
- Kerovec, M.; Marguš, D.; Mrakovčić, M.; Kučinić, M. A review of research into the fauna of the Krka National Park. In Book of Papers, Symposium River Krka and National Park “Krka”: Natural and Cultural Heritage, Protection and Sustainable Development; Marguš, D., Ed.; Javna ustanova “Nacionalni park Krka”: Šibenik, Croatia, 2005; pp. 49–61. ISBN 978-953-7406-01-6. [Google Scholar]
- Rätz, H.-J.; Lloret, J. Variation in fish condition between Atlantic cod (Gadus morhua) stocks, the effect on their productivity and management implications. Fish. Res. 2003, 60, 369–380. [Google Scholar] [CrossRef]
- Durand, J.D.; Persat, H.; Bouvet, Y. Phylogeography and postglacial dispersion of the chub (Leuciscus cephalus), in Europe. Mol. Ecol. 1999, 8, 989–997. [Google Scholar] [CrossRef]
- Freyhof, J.; Lieckfeldt, D.; Pitra, C.; Ludwig, A. Molecules and morphology: Evidence for introgression of mitochondrial DNA in Dalmatian cyprinids. Mol. Phylogenet Evol. 2005, 37, 347–354. [Google Scholar] [CrossRef]
- Perea, S.; Vukić, J.; Šanda, R.; Doadrio, I. Ancient mitochondrial capture as factor promoting mitonuclear discordance in freshwater fishes: A case study in the genus Squalius (Actinopterygii, Cyprinidae) in Greece. PLoS ONE 2016, 11, e0166292. [Google Scholar] [CrossRef]
- Buj, I.; Marčić, Z.; Čavlović, K.; Ćaleta, M.; Tutman, P.; Zanella, D.; Duplić, A.; Raguž, L.; Ivić, L.; Horvatić, S.; et al. Multilocus phylogenetic analysis helps to untangle the taxonomic puzzle of chubs (genus Squalius: Cypriniformes: Actinopteri) in the Adriatic basin of Croatia and Bosnia and Herzegovina. Zool. J. Linn. Soc. 2020, 189, 953–974. [Google Scholar] [CrossRef]
- Brito, R.M.; Briolay, J.; Galtier, N.; Bouvet, Y.; Coelho, M.M. Phylogenetic relationships within genus Leuciscus (Pisces, Cyprinidae) in Portuguese fresh waters, based on mitochondrial DNA cytochrome b sequences. Mol. Phylogenet Evol. 1997, 8, 435–442. [Google Scholar] [CrossRef] [PubMed]
- Cantatore, P.; Roberti, M.; Pesole, G.; Ludovico, A.; Milella, F.; Gadaleta, M.N.; Saccone, C. Evolutionary analysis of cytochrome b sequences in some perciformes: Evidence for a slower rate of evolution than in mammals. J. Mol. Evol. 1994, 39, 589–597. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed]
- Castresana, J. Selection of conserved blocks from multiple alignments for their use in phylogenetic analysis. Mol. Biol. Evol. 2000, 17, 540–552. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stechter, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Guindon, S.; Gascuel, O. A simple, fast, and accurate algorithm to estimate large phylogenies by maximum likelihood. Syst. Biol. 2003, 52, 696–704. [Google Scholar] [CrossRef] [PubMed]
- Posada, D.; Crandall, K.A. Modeltest: Testing the model of DNA substitution. Bioinformatics 1998, 14, 817–818. [Google Scholar] [CrossRef]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press: New York, NY, USA, 2000; ISBN 9780195135855. [Google Scholar]
- Perea, S.; Böhme, M.; Zupančić, P.; Freyhof, J.; Šanda, R.; Özuluğ, M.; Abdoli, A.; Doadrio, I. Phylogenetic relationships and biogeographical patterns in circum-Mediterranean subfamily Leuciscinae (Teleostei, Cyprinidae) inferred from both mitochondrial and nuclear data. BMC Evol. Biol. 2010, 10, 265. [Google Scholar] [CrossRef]
- Zardoya, R.; Doadrio, I. Molecular evidence of the evolutionary and biogeographical patterns of European cyprinids. J. Mol. Evol. 1999, 49, 227–237. [Google Scholar] [CrossRef]
- Rhymer, J.M.; Simberloff, D. Extinction by hybridization and introgression. Annu. Rev. Ecol. Syst. 1996, 27, 83–109. [Google Scholar] [CrossRef]
- Sousa-Santos, C.; Collares-Pereira, M.J.; Almada, V.C. Evidence of extensive mitochondrial introgression with nearly complete substitution of the typical Squalius pyrenaicus-like mtDNA of the Squalius alburnoides complex (Cyprinidae) in an independent Iberian drainage. J. Fish. Biol. 2006, 68, 292–301. [Google Scholar] [CrossRef]
- Tancioni, L.; Russo, T.; Cataudella, S.; Milana, V.; Hett, A.K.; Corsi, E.; Rossi, A.R. Testing species delimitations in four Italian sympatric leuciscine fishes in the Tiber River: A combined morphological and molecular approach. PLoS ONE 2013, 8, e60392. [Google Scholar] [CrossRef] [PubMed]
- Ünver, B.; Erk’akan, F. A natural hybrid of Leuciscus cephalus (L.) and Chalcalburnus chalcoides (Güldenstädt) (Osteichthyes: Cyprinidae) from Lake Tödürge (Sivas, Turkey). J. Fish. Biol. 2005, 66, 899–910. [Google Scholar] [CrossRef]
- Robalo, J.I.; Sousa-Santos, C.; Levy, A.; Almada, V.C. Molecular insights on the taxonomic position of the paternal ancestor of the Squalius alburnoides hybridogenetic complex. Mol. Phylogenet. Evol. 2006, 97, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Sousa-Santos, C.; Collares-Pereira, M.J.; Almada, V. Reading the history of a hybrid fish complex from its molecular record. Mol. Phylogenet. Evol. 2007, 45, 981–996. [Google Scholar] [CrossRef] [PubMed]
- Aboim, M.A.; Mavárez, J.; Bernatchez, L.; Coelho, M.M. Introgressive hybridization between two Iberian endemic cyprinid fish: A comparison between two independent hybrid zones. J. Evol. Biol. 2010, 23, 817–828. [Google Scholar] [CrossRef] [PubMed]
- Allen, B.E.; Anderson, M.L.; Mee, J.A.; Coombs, M.; Rogers, S.M. Role of genetic background in the introgressive hybridization of rainbow trout (Oncorhynchus mykiss) with westslope cutthroat trout (O. clarkia lewisi). Conserv. Genet. 2016, 17, 521–531. [Google Scholar] [CrossRef]
- Magalhaes, I.S.; Ornelas-Garcia, C.P.; Leal-Cardin, M.; Ramirez, T.; Barluenga, M. Untangling the evolutionary history of a highly polymorphic species: Introgressive hybridization and high genetic structure in the desert cichlid fish Herichthys minckleyi. Mol. Ecol. 2015, 24, 4505–4520. [Google Scholar] [CrossRef]
- Desvignes, T.; Le François, N.R.; Goetz, L.C.; Smith, S.S.; Shusdock, K.A.; Parker, S.K.; Postlethwait, J.H.; William Detrich, H., III. Intergeneric hybrids inform reproductive isolating barriers in the Antarctic icefish radiation. Sci. Rep. 2019, 9, 5989. [Google Scholar] [CrossRef]
- Kwan, Y.-S.; Ko, M.-H.; Jeon, Y.-S.; Kim, H.-J.; Won, Y.-J. Bidirectional mitochondrial introgression between Korean cobitid fish mediated by hybridogenetic hybrids. Ecol. Evol. 2019, 9, 1244–1254. [Google Scholar] [CrossRef]
- Ketmaier, V.; Bianco, P.G.; Cobolli, M.; Krivokapić, M.; Canigla, R.; De Matthaeis, E. Molecular phylogeny of two lineages of Leuciscinae cyprinids (Telestes and Scardinius) from the peri-Mediterranean area based on cytochrome b data. Mol. Phylogenet. Evol. 2004, 32, 1061–1071. [Google Scholar] [CrossRef] [PubMed]
- Bianco, P.G. Potential role of the paleohistory of the Mediterranean and Paratethys basins on the early dispersal of Euro-Mediterranean freshwater fishes. Ichthyol. Explor. Freshw. 1990, 1, 167–184. [Google Scholar]
- Doadrio, I. Origen y evolución de la ictiofauna continental Española. In Atlas y Libro Rojo de los Peces Espańoles; Doadrio, I., Ed.; DGCONA-CSIC: Madrid, Spain, 2001; pp. 20–34. ISBN 8480143134. [Google Scholar]
- Sabolić, M.; Buj, I.; Ćaleta, M.; Marčić, Z. Morphological diversity and relationships of Scardinius dergle and Scardinius plotizza populations (Actinopteri; Cypriniformes) from Croatia and Bosnia and Herzegovina. J. Cent. Eur. Agric. 2021, 22, 713–734. [Google Scholar] [CrossRef]



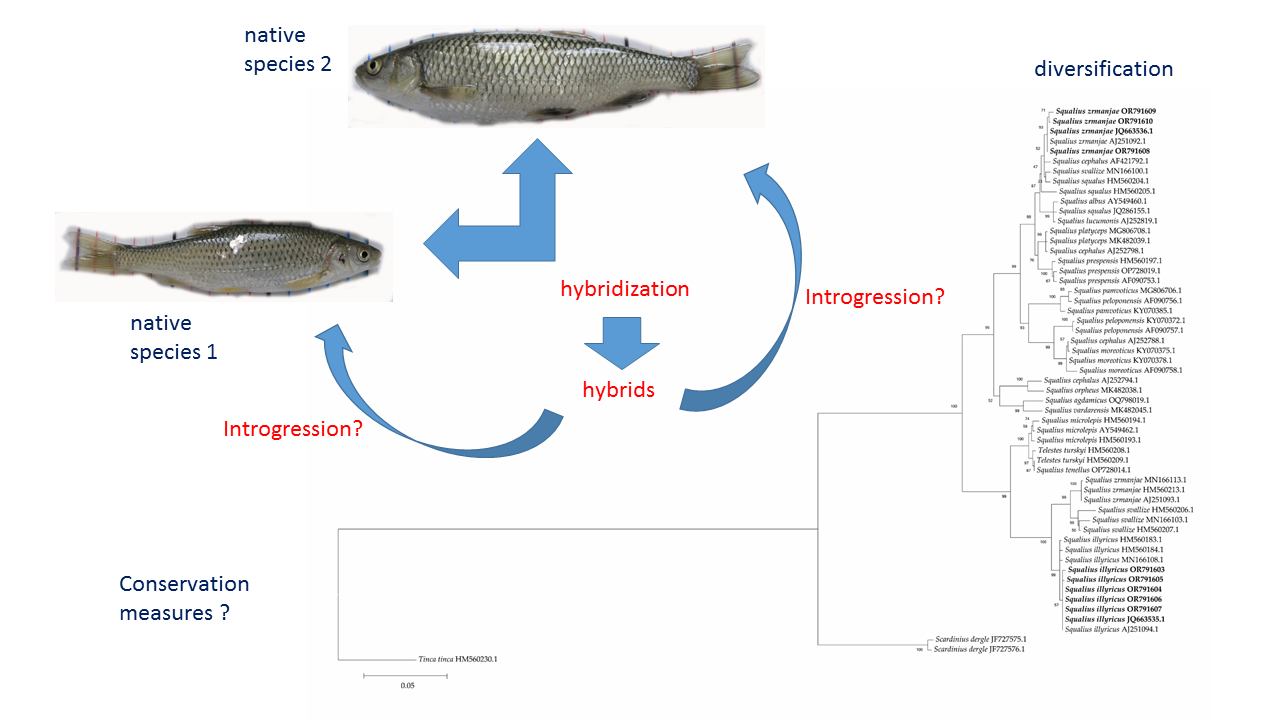{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
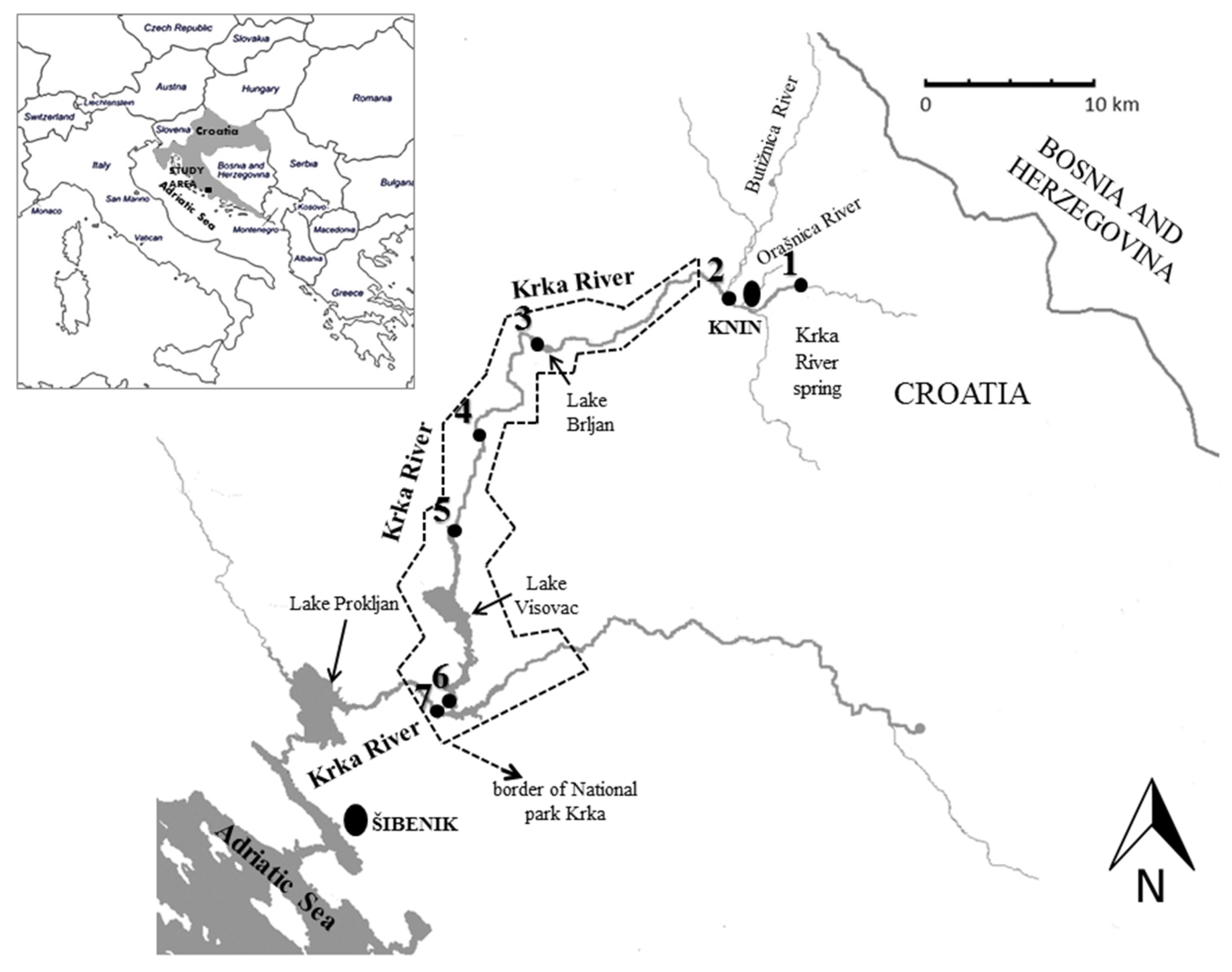
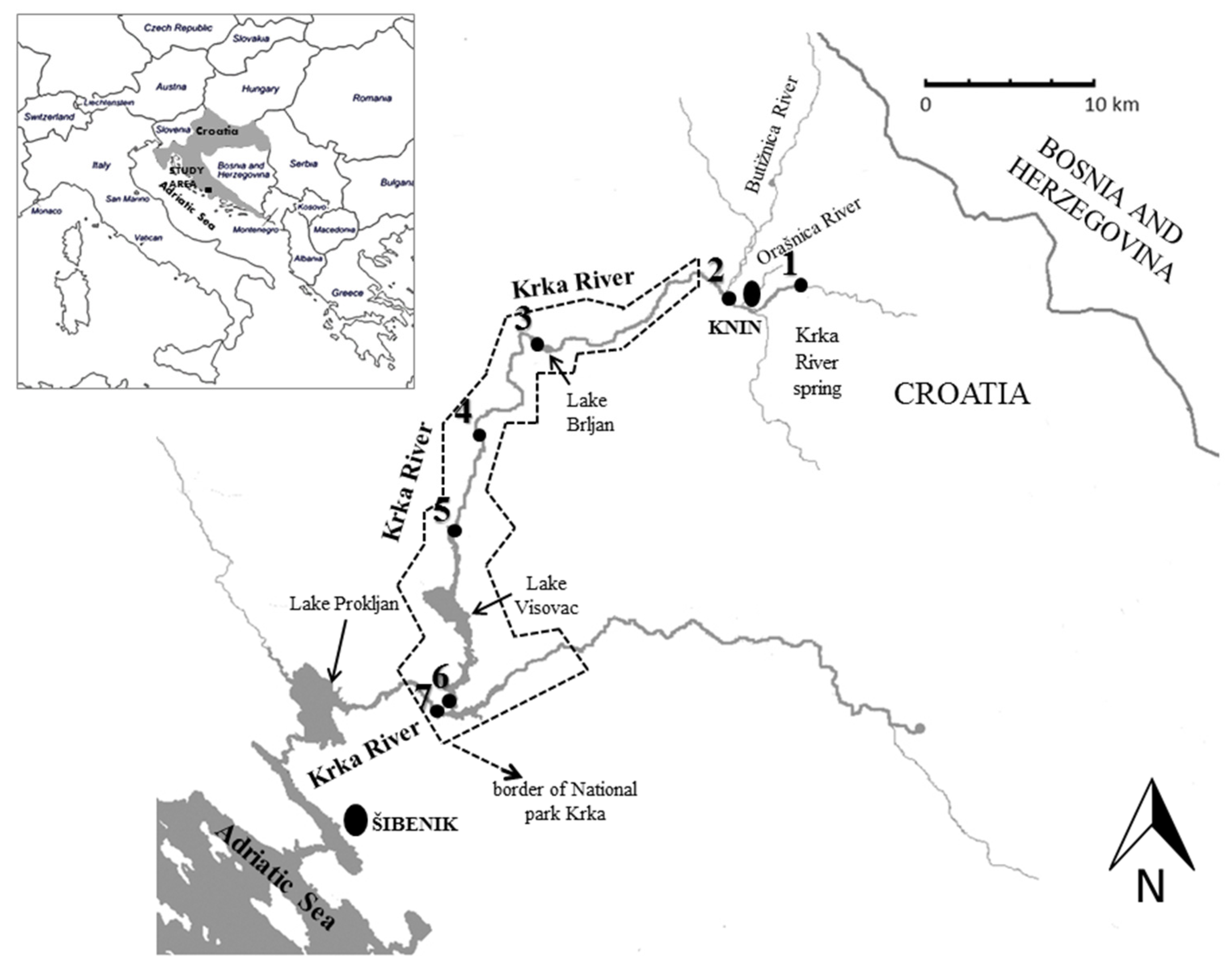
| No. | Name of the Sampling Site | Coordinates |
|---|---|---|
| 1. | Krka River spring | 44°02.563′ N, 16°14.412′ E |
| 2. | Krka River upstream of the Butižnica River tributary | 44°02.295′ N, 16°10.347′ E |
| 3. | Lake Brljan | 44°00.343′ N, 16°02.444′ E |
| 4. | “Krka” monastery | 43°57.538′ N, 15°59.833′ E |
| 5. | Roški Slap waterfall | 43°54.140′ N, 15°58.815′ E |
| 6. | Visovac Lake upstream from waterfall Skradinski Buk | 43°48.349′ N, 15°58.693′ E |
| 7. | Skradinski Buk waterfall | 43°48.426′ N, 15°58.110′ E |
| Morphological Characters | Scardinius | Squalius |
|---|---|---|
| Body shape | Compressed | Cylindrical |
| Shape of the posterior anal-fin margin | Concave | Convex |
| Mouth position | Superior | Terminal |
| Pharyngeal teeth | Slightly serrated | Smooth |
| Branched anal rays | 10 | 8–9½ * |
| Gill rakers | 10–13 | 12–16 ** |
| Lateral line scales | 38–41 | 46–54 or 44–49* |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valić, D.; Mirković, M.K.; Besendorfer, V.; Teskeredžić, E. Molecular Analysis of Two Endemic Squalius Species: Evidence for Intergeneric Introgression among Cyprinids and Conservation Issues. Fishes 2024, 9, 4. https://doi.org/10.3390/fishes9010004
Valić D, Mirković MK, Besendorfer V, Teskeredžić E. Molecular Analysis of Two Endemic Squalius Species: Evidence for Intergeneric Introgression among Cyprinids and Conservation Issues. Fishes. 2024; 9(1):4. https://doi.org/10.3390/fishes9010004
Chicago/Turabian StyleValić, Damir, Matej Kristan Mirković, Višnja Besendorfer, and Emin Teskeredžić. 2024. "Molecular Analysis of Two Endemic Squalius Species: Evidence for Intergeneric Introgression among Cyprinids and Conservation Issues" Fishes 9, no. 1: 4. https://doi.org/10.3390/fishes9010004
APA StyleValić, D., Mirković, M. K., Besendorfer, V., & Teskeredžić, E. (2024). Molecular Analysis of Two Endemic Squalius Species: Evidence for Intergeneric Introgression among Cyprinids and Conservation Issues. Fishes, 9(1), 4. https://doi.org/10.3390/fishes9010004