

The Composition and Diversity of Intestinal Microbes at Different Growth Stages of Giant Freshwater Prawns (Macrobrachium rosenbergii)


Abstract
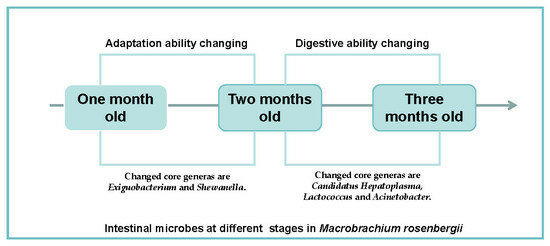
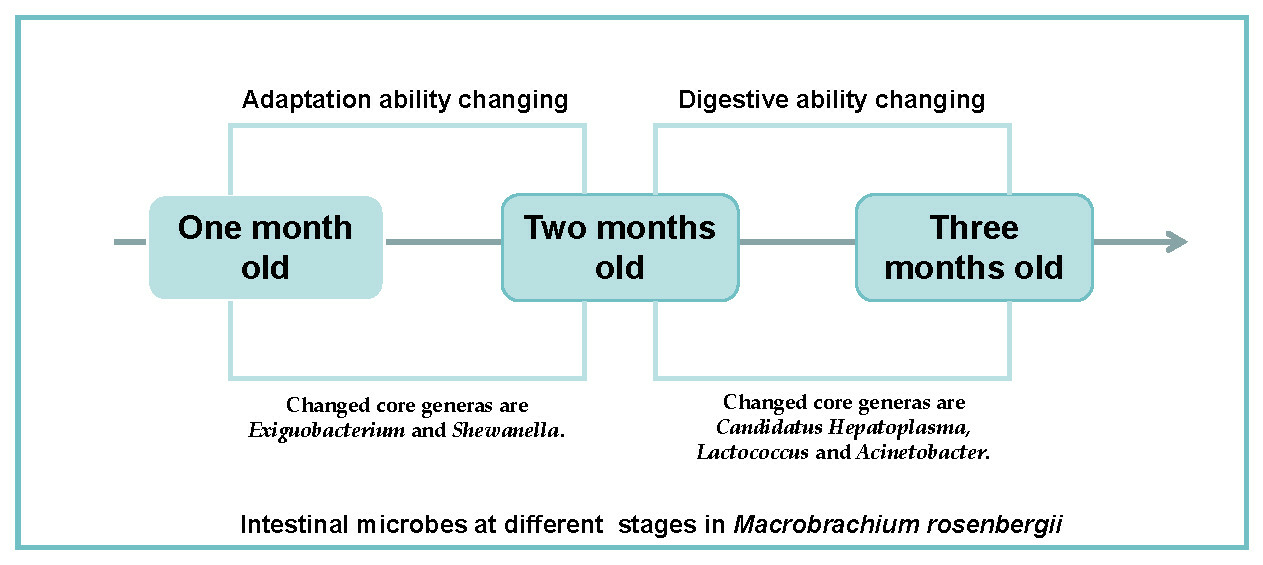
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Genomic DNA (gDNA) Extraction and 16S rDNA Sequencing
2.3. Data Analysis
2.4. Phylogenetic Distance and Community Distribution
2.5. Statistical Analysis
3. Results
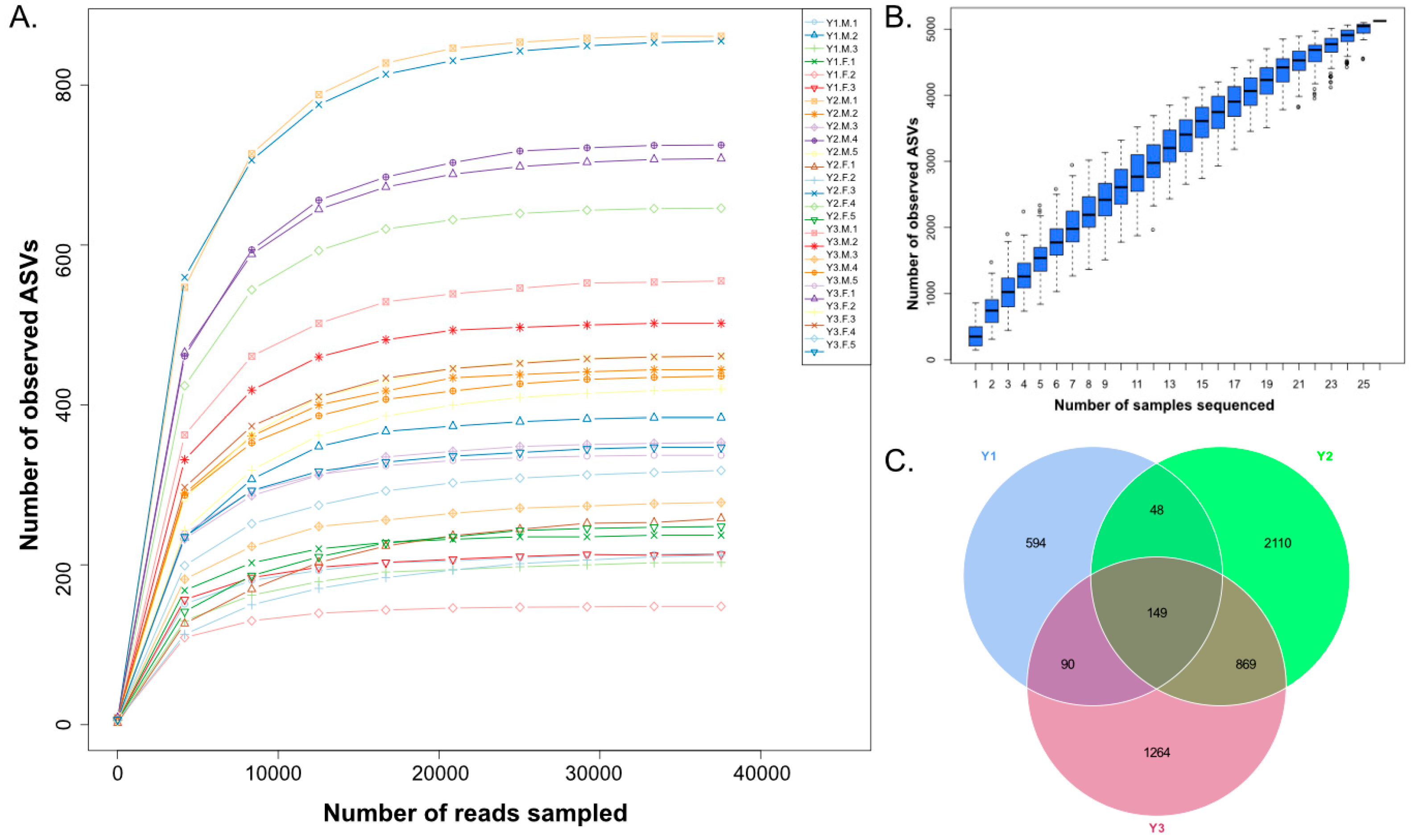
3.1. OTU Analysis and Alpha Diversity
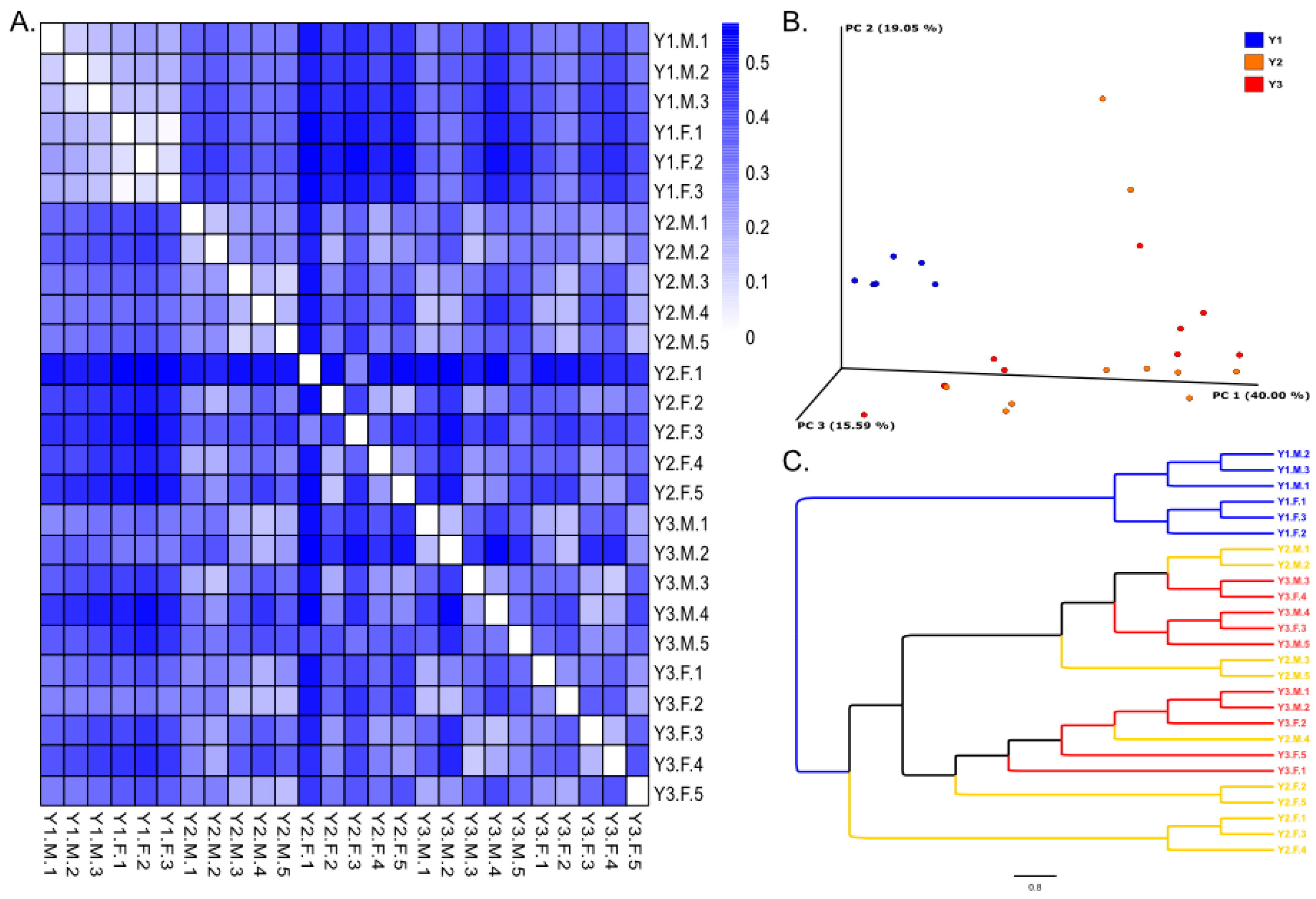
3.2. Beta Diversity
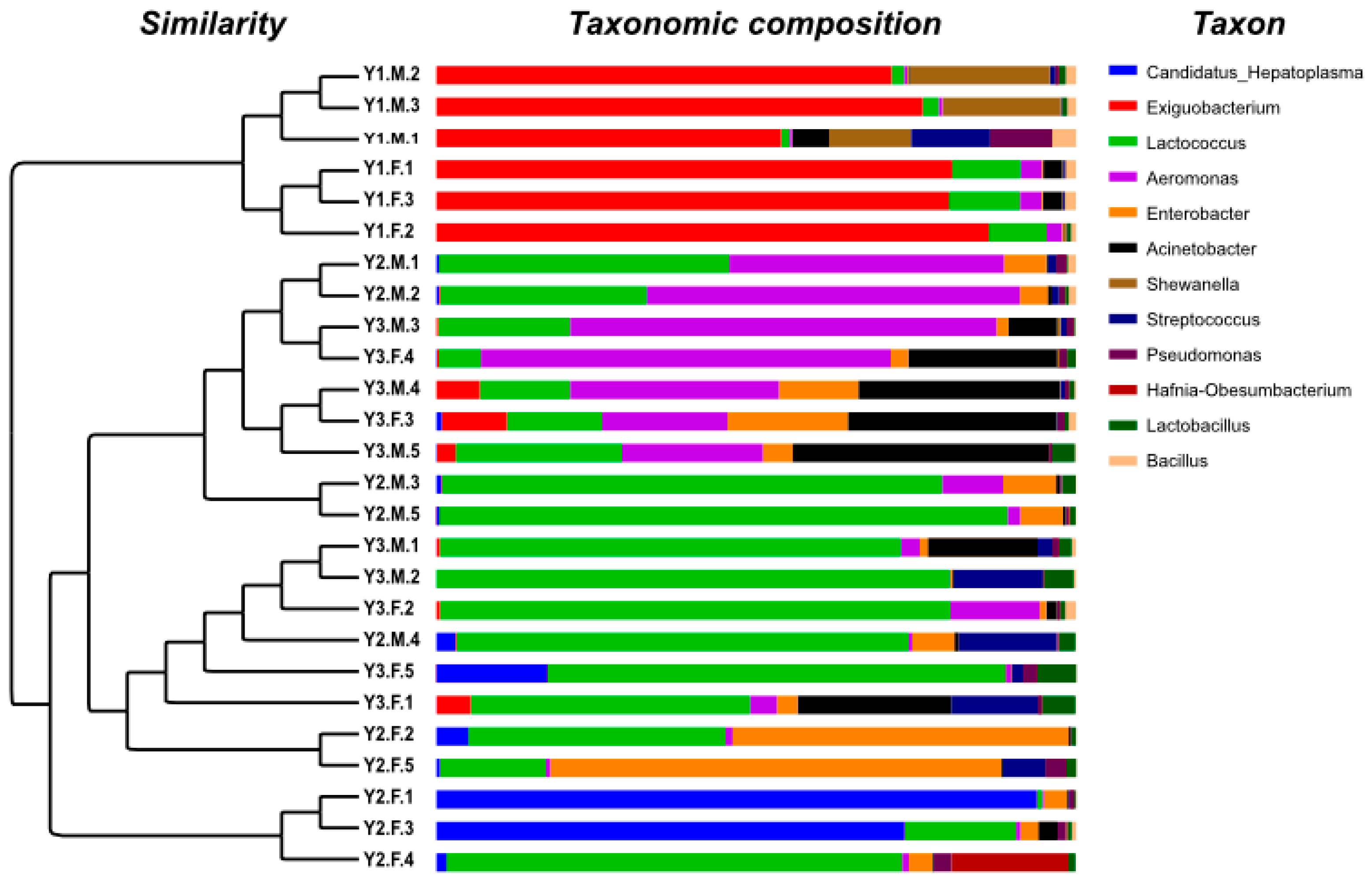
3.3. Taxonomic Composition
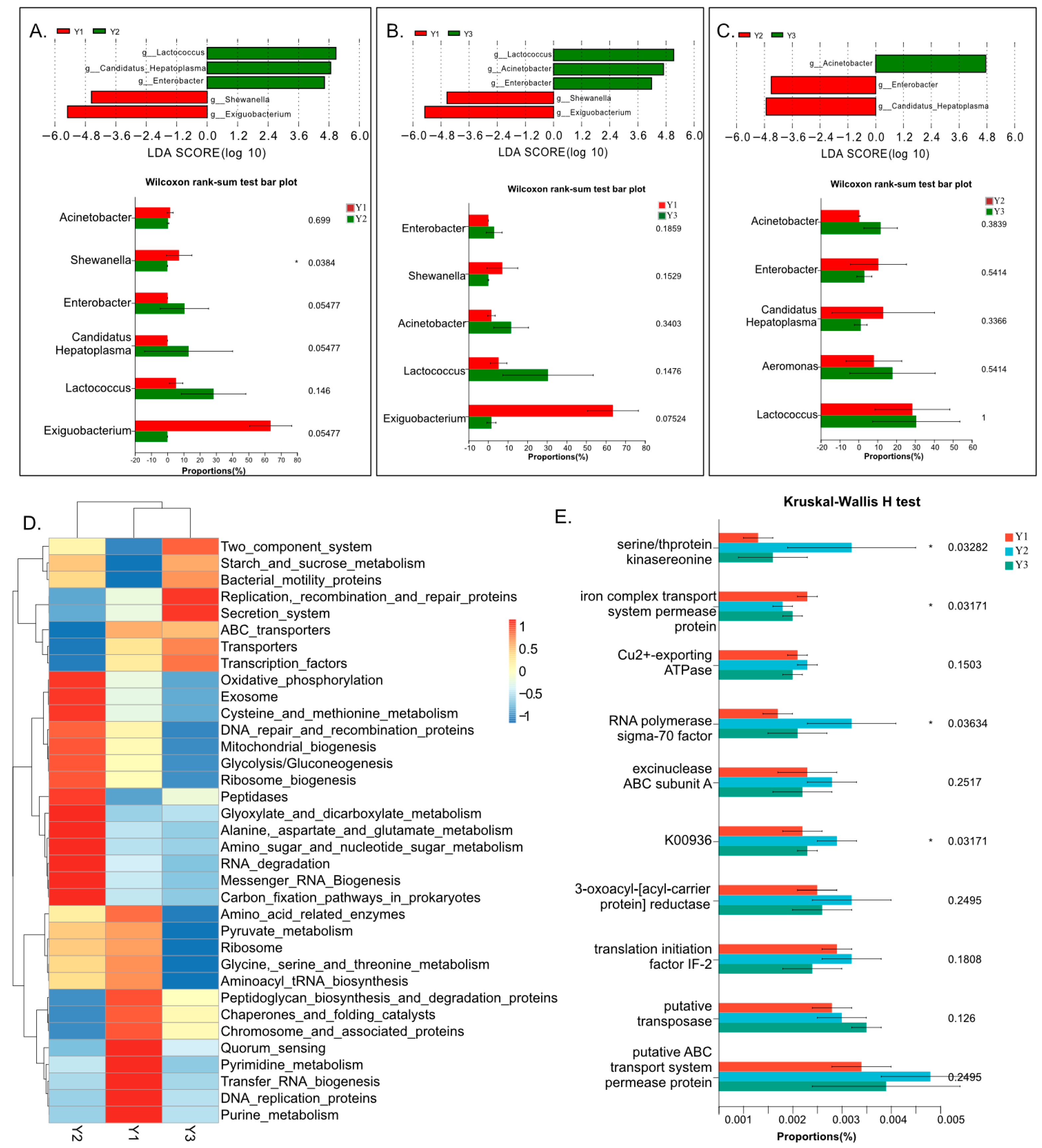
3.4. Variance Analysis and Function Prediction
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hameed, A.S.; Yoganandhan, K.; Sri Widada, J.; Bonami, J.R. Experimental transmission and tissue tropism of Macrobrachium rosenbergii nodavirus (MrNV) and its associated extra small virus (XSV). Dis. Aquat. Org. 2004, 62, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Hossain, F.E.; Ramim, A.M.; Rahman, M.S.; Faruque, A.; Naser, M.N.; Karim, M.M. Identification of bacterial agents causing mortality in postlarvae of giant freshwater prawn (Macrobrachium rosenbergii) in south-west coastal districts of bangladesh. Aquac. Res. 2017, 48, 3545–3555. [Google Scholar] [CrossRef]
- Bruyn, M.D.; Nugroho, E.; Hossain, M.M.; Wilson, J.C.; Mather, P.B. Phylogeographic evidence for the existence of an ancient biogeographic barrier: The isthmus of Kra seaway. Heredity 2005, 94, 370–378. [Google Scholar] [CrossRef] [PubMed]
- Kogut, M.H.; Lee, A.; Santin, E. Microbiome and pathogen interaction with the immune system. Poult. Sci. 2020, 99, 1906–1913. [Google Scholar] [CrossRef]
- Yatsunenko, T.; Rey, F.E.; Manary, M.J.; Trehan, I.; Dominguez-Bello, M.G.; Contreras, M.; Magris, M.; Hidalgo, G.; Baldassano, R.N.; Anokhin, A.P.; et al. Human gut microbiome viewed across age and geography. Nature 2012, 486, 222–227. [Google Scholar] [CrossRef]
- Holt, C.C.; Bass, D.; Stentiford, G.D.; van der Giezen, M. Understanding the role of the shrimp gut microbiome in health and disease. J. Invertebr. Pathol. 2021, 186, 107387. [Google Scholar] [CrossRef]
- Zheng, X.; Liu, B.; Wang, N.; Yang, J.; Zhou, Q.; Sun, C.; Zhao, Y. Low fish meal diet supplemented with probiotics ameliorates intestinal barrier and immunological function of Macrobrachium rosenbergii via the targeted modulation of gut microbes and derived secondary metabolites. Front. Immunol. 2022, 13, 1074399. [Google Scholar] [CrossRef]
- Ma, R.; Wang, Y.; Zhao, S.; Yin, M.; Fang, W. The composition of the microbial community associated with Macrobrachium rosenbergii zoeae varies throughout larval development. Fish Dis. 2020, 43, 413–421. [Google Scholar] [CrossRef]
- Liu, B.; Song, C.; Gao, Q.; Liu, B.; Zhou, Q.; Sun, C.; Zhang, H.; Liu, M.; Tadese, D.A. Maternal and environmental microbes dominate offspring microbial colonization in the giant freshwater prawn Macrobrachium rosenbergii. Sci. Total Environ. 2021, 790, 148062. [Google Scholar] [CrossRef]
- Cornejo-Granados, F.; Gallardo-Becerra, L.; Leonardo-Reza, M.; Ochoa-Romo, J.P.; Ochoa-Leyva, A. A meta-analysis reveals the environmental and host factors shaping the structure and function of the shrimp microbiota. Peer J. 2018, 6, e5382. [Google Scholar] [CrossRef]
- Huang, Z.; Li, X.; Wang, L.; Shao, Z. Changes in the intestinal bacterial community during the growth of white shrimp, Litopenaeus Vannamei. Aquac. Res. 2016, 47, 1737–1746. [Google Scholar] [CrossRef]
- Zheng, Y.; Yu, M.; Liu, J.; Qiao, Y.; Wang, L.; Li, Z.; Zhang, X.-H.; Yu, M. Bacterial community associated with healthy and diseased Pacific white shrimp (Litopenaeus vannamei) larvae and rearing water across different growth stages. Front. Microbiol. 2017, 8, 1362. [Google Scholar] [CrossRef] [PubMed]
- Hou, D.; Huang, Z.; Zeng, S.; Liu, J.; Wei, D.; Deng, X.; Weng, S.; He, Z.; He, J. Environmental factors shape water microbial community structure and function in shrimp cultural enclosure ecosystems. Front. Microbiol. 2017, 8, 2359. [Google Scholar] [CrossRef] [PubMed]
- Hammer, T.J.; McMillan, W.O.; Fierer, N. Metamorphosis of a butterfly-associated bacterial community. PLoS ONE 2014, 9, e86995. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Sun, X. Core gut microbiota of shrimp sunction as a regulator to maintain immune homeostasis in response to WSSV infection. Microbiol. Spectr. 2022, 10, e0246521. [Google Scholar]
- Muhammad, A.; Fang, Y.; Hou, Y.; Shi, Z. The gut entomotype of red palm weevil Rhynchophorus ferrugineus Olivier (Coleoptera: Dryophthoridae) and their rffect on host nutrition metabolism. Front. Microbiol. 2017, 8, 2291. [Google Scholar] [CrossRef]
- Oliveira, R.A.; Ng, K.M.; Correia, M.B.; Cabral, V.; Shi, H.; Sonnenburg, J.L.; Huang, K.C.; Xavier, K.B. Klebsiella michiganensis transmission enhances resistance to Enterobacteriaceae gut invasion by nutrition competition. Nat. Microbiol. 2020, 5, 630–641. [Google Scholar] [CrossRef]
- Manfioletti, G.; Schneider, C. A new and fast method for preparing high quality lambda DNA suitable for sequencing. Nucleic Acids Res. 1988, 16, 2873–2884. [Google Scholar] [CrossRef]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef]
- Haas, B.J.; Gevers, D.; Earl, A.M.; Feldgarden, M.; Ward, D.V.; Giannoukos, G.; Ciulla, D.; Tabbaa, D.; Highlander, S.K.; Sodergren, E.; et al. Chimeric 16S rRNA sequence formation and detection in Sanger and 454-pyrosequenced PCR amplicons. Genome Res. 2011, 21, 494–504. [Google Scholar] [CrossRef]
- Li, M.; Shao, D.; Zhou, J.; Gu, J.; Qin, J.; Chen, W.; Wei, W. Signatures within esophageal microbiota with progression of esophageal squamous cell carcinoma. Chin. J. Cancer Res. 2020, 32, 755–767. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Gaughan, S.; Chang, Q.; Chen, H.; Lu, G.; Wang, X.; Xu, L.; Zhu, L.; Jiang, J. Age-related changes in the gut microbiota of the Chinese giant salamander (Andrias davidianus). Microbiologyopen 2019, 8, e00778. [Google Scholar] [CrossRef]
- Zhang, C.; Zhang, J.; Fan, W.; Huang, M.; Liu, M. Effects of dietary Lactobacillus delbrueckii on growth performance, body composition, digestive and absorptive capacity, and gene expression of common carp (Cyprinus carpio Huanghe var). Aquac. Nutr. 2019, 25, 166–175. [Google Scholar] [CrossRef]
- Castex, M.; Leclercq, E.; Lemaire, P.; Chim, L. Dietary probiotic Pediococcus acidilactici MA18/5M improves the growth, feed performance and antioxidant status of penaeid shrimp Litopenaeus stylirostris: A Growth-Ration-Size Approach. Animals 2021, 11, 3451. [Google Scholar] [CrossRef]
- Amenyogbe, E.; Luo, J.; Fu, W.J.; Abarike, E.D.; Wang, Z.L.; Huang, J.S.; Ayisi, C.L.; Chen, G. Effects of autochthonous strains mixture on gut microbiota and metabolic profile in cobia (Rachycentron canadum). Sci. Rep. 2022, 12, 17410. [Google Scholar] [CrossRef] [PubMed]
- Cao, Q.; Najnine, F.; Han, H.; Wu, B.; Cai, J. BALOs Improved gut microbiota health in postlarval shrimp (Litopenaeus vannamei) after being subjected to salinity reduction treatment. Front. Microbiol. 2020, 11, 1296. [Google Scholar] [CrossRef]
- Guo, Y.; Zhang, L.; Liang, Y.; Li, P.; Zhang, T.; Meng, F.; Liu, B.; Zhang, H.; Fu, W.; Wang, W.; et al. Effects of dietary yeast culture on health status in digestive tract of juvenile Pacific white shrimp Litopenaeus Vannamei. Fish Shellfish Immunol. Rep. 2022, 3, 100065. [Google Scholar]
- Zeng, S.; Khoruamkid, S.; Kongpakdee, W.; Wei, D.; Yu, L.; Wang, H.; Deng, Z.; Weng, S.; Huang, Z.; He, J.; et al. Dissimilarity of microbial diversity of pond water, shrimp intestine and sediment in aquamimicry system. AMB Express 2020, 10, 180. [Google Scholar] [CrossRef]
- Patil, P.K.; Vinay, T.N.; Ghate, S.D.; Baskaran, V.; Avunje, S. 16 S rRNA gene diversity and gut microbial composition of the Indian white shrimp (Penaeus indicus). Antonie Van Leeuwenhoek 2021, 114, 2019–2031. [Google Scholar] [CrossRef]
- Chen, H.; Liu, F.; Ouyang, M.; Zhou, H.; Lou, B. Differences in intestinal microbial composition between red claw crayfish (Cherax quadricarinatus) and red swamp crayfish (Procambarus clarkii) cultured in pond. Fishes 2022, 7, 241. [Google Scholar] [CrossRef]
- Kasana, R.C.; Pandey, C.B. Exiguobacterium: An overview of a versatile genus with potential in industry and agriculture. Crit. Rev. Biotechnol. 2018, 38, 141–156. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.K.; Raichand, R.; Kaur, I.; Kaur, C.; Pareek, S.; Mayilraj, S. Exiguobacterium himgiriensis sp. nov. a novel member of the genus Exiguobacterium, isolated from the Indian Himalayas. Antonie Van Leeuwenhoek 2013, 103, 789–796. [Google Scholar] [CrossRef] [PubMed]
- Raichand, R.; Pareek, S.; Singh, N.K.; Mayilraj, S. Exiguobacterium aquaticum sp. nov., a member of the genus Exiguobacterium. Int. J. Syst. Evol. Microbiol. 2012, 62, 2150–2155. [Google Scholar] [CrossRef]
- Zhang, D.; Zhu, Z.; Li, Y.; Li, X.; Guan, Z.; Zheng, J. Comparative genomics of Exiguobacterium reveals what makes a cosmopolitan bacterium. Msystems 2021, 6, e0038321. [Google Scholar] [CrossRef] [PubMed]
- Cong, M.; Jiang, Q.; Xu, X.; Huang, L.; Su, Y.; Yan, Q. The complete genome sequence of Exiguobacterium arabatum W-01 reveals potential probiotic functions. MicrobiologyOpen 2017, 6, e496. [Google Scholar] [CrossRef]
- Cai, M.; Deng, H.; Sun, H.; Si, W.; Li, X.; Hu, J.; Huang, M.; Fan, W. Changes of intestinal microbiota in the giant salamander (Andrias davidianus) during growth based on high-throughput sequencing. Front. Microbiol. 2023, 14, 1052824. [Google Scholar] [CrossRef]
- Ling, F.; Steinel, N.; Weber, J.; Ma, L.; Smith, C.; Correa, D.; Zhu, B.; Bolnick, D.; Wang, G. The gut microbiota response to helminth infection depends on host sex and genotype. ISME J. 2020, 14, 1141–1153. [Google Scholar] [CrossRef]
- Meziti, A.; Mente, E.; Kormas, K.A. Gut bacteria associated with different diets in reared Nephrops norvegicus. Syst. Appl. Microbiol. 2012, 35, 473–482. [Google Scholar] [CrossRef]
- Fraune, S.; Zimmer, M. Host-specificity of environmentally transmitted Mycoplasma-like isopod symbionts. Environ. Microbiol. 2008, 10, 2497–2504. [Google Scholar] [CrossRef]
- Ooi, M.C.; Goulden, E.F.; Smith, G.G.; Nowak, B.F.; Bridle, A.R. Developmental and gut-related changes to microbiomes of the cultured juvenile spiny lobster Panulirus ornatus. FEMS Microbiol. Ecol. 2017, 93, 1–10. [Google Scholar] [CrossRef]
- Sui, J.; Luan, S.; Yang, G.; Xia, Z.; Luo, K.; Tang, Q.; Lu, X.; Meng, X.; Kong, J. Genetic parameters and selection response for the harvest body weight of the giant freshwater prawn (Macrobrachium rosenbergii) in a breeding program in China. PLoS ONE 2019, 14, e0218379. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.L.; Liu, Z.Y.; Jin, Y.M.; Liu, Z.X.; Zhang, B.Y.; Yuan, Z.H.; Ye, J.D.; Sun, Y.Z. Preventive and reparative functions of host-associated probiotics against soybean meal induced growth, immune suppression and gut injury in Japanese seabass (Lateolabraxjaponicus). Fish Shellfish Immunol. 2022, 128, 651–663. [Google Scholar] [CrossRef] [PubMed]
- Amoah, K.; Huang, Q.C.; Tan, B.P.; Zhang, S.; Chi, S.Y.; Yang, Q.H.; Liu, H.Y.; Dong, X.H. Dietary supplementation of probiotic Bacillus coagulans ATCC 7050, improves the growth performance, intestinal morphology, microflora, immune response, and disease confrontation of Pacific white shrimp, Litopenaeus Vannamei. Fish Shellfish Immunol. 2019, 87, 796–808. [Google Scholar] [CrossRef]
- Won, S.; Hamidoghli, A.; Choi, W.; Bae, J.; Jang, W.J.; Lee, S.; Bai, S.C. Evaluation of potential probiotics Bacillus subtilis WB60, Pediococcus pentosaceus, and Lactococcus lactis on growth performance, immune response, gut histology and immune-related genes in whiteleg shrimp, Litopenaeus Vannamei. Microorg. 2020, 8, 281. [Google Scholar] [CrossRef] [PubMed]
- Adel, M.; El-Sayed, A.M.; Yeganeh, S.; Dadar, M.; Giri, S.S. Effect of potential probiotic Lactococcus lactis Subsp. lactis on growth performance, intestinal microbiota, digestive enzyme activities, and disease resistance of Litopenaeus vannamei. Probiotics Antimicrob. Proteins 2017, 9, 150–156. [Google Scholar] [CrossRef] [PubMed]
- Dou, Y.; Wen, M.; Shen, H.; Zhang, S.; Jiang, G.; Qiao, Y.; Cheng, J.; Cao, X.; Wan, X.; Sun, X. Intestinal microbiota differences in Litopenaeus vannamei shrimp between greenhouse and aquaponic rearing. Life 2023, 13, 525. [Google Scholar] [CrossRef] [PubMed]
- Elizarov, S.M.; Mironov, V.A.; Danilenko, V.N. Dynamics of serine/threonine protein kinase activity during the growth of the wild-type Streptomyces avermitilis strain and its chloramphenicol-resistant mutant. Microbiology 2000, 69, 281–286. [Google Scholar] [CrossRef]
- Richardson, L.A.; Reed, B.J.; Charette, J.M.; Freed, E.F.; Fredrickson, E.K.; Locke, M.N.; Baserga, S.J.; Gardner, R.G. A conserved deubiquitinating enzyme controls cell growth by regulating RNA polymerase I stability. Cell Rep. 2012, 2, 372–385. [Google Scholar] [CrossRef] [PubMed]
- Deng, Z.; Zeng, S.; Zhou, R.; Hou, D.; Bao, S.; Zhang, L.; Hou, Q.; Li, X.; Weng, S.; He, J.; et al. Phage-prokaryote coexistence strategy mediates microbial community diversity in the intestine and sediment microhabitats of shrimp culture pond ecosystem. Front. Microbiol. 2022, 13, 1011342. [Google Scholar] [CrossRef] [PubMed]
- Interaminense, J.A.; Vogeley, J.L.; Gouveia, C.K.; Portela, R.S.; Oliveira, J.P.; Silva, S.M.B.C.; Coimbra, M.R.M.; Peixoto, S.M.; Soares, R.B.; Buarque, D.S.; et al. Effects of dietary Bacillus subtilis and Shewanella algae in expression profile of immune-related genes from hemolymph of Litopenaeus vannamei challenged with Vibrio parahaemolyticus. Fish Shellfish Immunol. 2019, 86, 253–259. [Google Scholar] [CrossRef]
- Natnan, M.E.; Low, C.F.; Chong, C.M.; Bunawan, H.; Baharum, S.N. Oleic acid as potential immunostimulant in metabolism pathways of hybrid grouper fingerlings (Epinephelus fuscoguttatus × Epinephelus lanceolatus) infected with Vibrio vulnificus. Sci. Rep. 2023, 13, 12830. [Google Scholar] [CrossRef] [PubMed]
- Peters, T.J. Intestinal peptidases. Gut 1970, 11, 720–725. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Tassanakajon, A.; Rimphanitchayakit, V.; Visetnan, S.; Amparyup, P.; Somboonwiwat, K.; Charoensapsri, W.; Tang, S. Shrimp humoral responses against pathogens: Antimicrobial peptides and melanization. Dev. Comp. Immunol. 2018, 80, 81–93. [Google Scholar] [CrossRef] [PubMed]
- Schubiger, C.B.; Orfe, L.H.; Sudheesh, P.S.; Cain, K.D.; Shah, D.H.; Call, D.R. Entericidin is required for a probiotic treatment (Enterobacter sp. strain C6-6) to protect trout from cold-water disease challenge. Appl. Environ. Microbiol. 2015, 81, 658–665. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diversity Indices | Y1 | Y2 | Y3 | Wilcoxon Test |
|---|---|---|---|---|
| Chao1 | 234.37 ± 79.27 | 508.59 ± 15.54 | 438.11 ± 128.8 | Y2, Y3 vs. Y1 with p < 0.01 |
| Shannon | 3.87 ± 0.47 | 4.29 ± 0.09 | 4.52 ± 1.12 | Pairwise test with p > 0.05 |
| Simpson | 0.86 ± 0.04 | 0.79 ± 0.01 | 0.8 ± 0.14 | Pairwise test with p > 0.05 |
| Dominance | 0.14 ± 0.04 | 0.21 ± 0.01 | 0.2 ± 0.14 | Pairwise test with p > 0.05 |
| Good’s coverage | 1.00 ± 0.00 | 1.00 ± 0.00 | 1.00 ± 0.00 | Pairwise test with p > 0.05 |
| Observed ASVs | 233.17 ± 79.63 | 506.3 ± 14.6 | 436.2 ± 129.1 | Y2, Y3 vs. Y1 with p < 0.01 |
| Phyla Name | Y1 | Y2 | Y3 |
|---|---|---|---|
| Firmicutes | 79.24 ± 3.96% | 47.84 ± 6.87% | 47.3 ± 8.83% |
| Proteobacteria | 17.09 ± 3.4% | 44.22 ± 6.84% | 46.66 ± 9.23% |
| Actinobacteriota | 2.01 ± 0.54% | 1.83 ± 0.75% | 1.52 ± 0.44% |
| Acidobacteriota | 0.04 ± 0.04% * | 1.66 ± 0.65% | 0.36 ± 0.1% * |
| Bacteroidota | 0.64 ± 0.18% * | 1.24 ± 0.43% | 1.45 ± 0.38% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, H.; Ren, J.; Xu, X.; Lou, B.; Zhang, D. The Composition and Diversity of Intestinal Microbes at Different Growth Stages of Giant Freshwater Prawns (Macrobrachium rosenbergii). Fishes 2023, 8, 473. https://doi.org/10.3390/fishes8090473
Xu H, Ren J, Xu X, Lou B, Zhang D. The Composition and Diversity of Intestinal Microbes at Different Growth Stages of Giant Freshwater Prawns (Macrobrachium rosenbergii). Fishes. 2023; 8(9):473. https://doi.org/10.3390/fishes8090473
Chicago/Turabian StyleXu, Hongxiang, Jindong Ren, Xiaojun Xu, Bao Lou, and Demin Zhang. 2023. "The Composition and Diversity of Intestinal Microbes at Different Growth Stages of Giant Freshwater Prawns (Macrobrachium rosenbergii)" Fishes 8, no. 9: 473. https://doi.org/10.3390/fishes8090473
APA StyleXu, H., Ren, J., Xu, X., Lou, B., & Zhang, D. (2023). The Composition and Diversity of Intestinal Microbes at Different Growth Stages of Giant Freshwater Prawns (Macrobrachium rosenbergii). Fishes, 8(9), 473. https://doi.org/10.3390/fishes8090473