3.2. Relationships and Taxonomy of Main PAC Lineages
The phylogenetic analyses supported species of PAC from RDB belonging to four lineages (
Figure 2): 1—
Trichomycterus brigadeirensis Costa, Katz and Vilardo sp. nov., from the Serra do Brigadeiro and below described, supported as sister to a clade comprising all other species of PAC; 2—
Trichomycterus caratinguensis Costa, Katz and Vilardo sp. nov., from the Rio Caratinga drainage and below described, weakly supported as sister to the
T. goeldii complex; 3—the
Trichomycterus goeldii complex as delimited in Costa et al. [
12]; 4—the
Trichomycterus jacupiranga as delimited in Costa et al. [
12]. No species from RDB belong to the
Trichomycterus puriventris and
Trichomycterus travassosi complexes.
Trichomycterus brigadeirensis Costa, Katz and Vilardo sp. nov.
LSID:urn:lsid:zoobank.org:act:4E1ADC38-014E-4A3F-B8DE-AD09935F3996
Holotype. UFRJ 13480, 78.6 mm SL; Brazil: Minas Gerais State: Araponga Municipality: Ribeirão Félix, upper Rio Casca drainage, Rio Doce basin, Serra do Brigadeiro, 20°41′31″ S 42°29′52″ W, altitude about 1,100 m; A. M. Katz and P. J. Vilardo, 25 September 2022.
Paratypes. UFRJ 13239, 10 ex., 26.8–65.7 mm SL; UFRJ 13655, 4 ex. (C&S), 33.1–47.1 mm SL; UFRJ 13169, 4 ex. (DNA), 30.3–36.0 mm SL; CICCAA 07772, 2 ex., 45.1–45.8 mm SL; all collected with holotype.
Diagnosis. Trichomycterus brigadeirensis differs from all other congeners of PAC using the following unique combination of character states: 19–21 dorsal procurrent caudal-fin rays (vs. 13–15 in Trichomycterus saquarema Costa, Katz, Vilardo, and Amorim, 2022; 16–18 in T. auroguttatus, Trichomycterus caudofasciatus Alencar and Costa, 2004, Trichomycterus goeldii Boulenger, 1896, T. mimosensis Barbosa, 2013, and T. vinnulus; and 22–28 in Trichomycterus altipombensis Costa, Katz, Vilardo, and Mattos, 2022, T. astromycterus, Trichomycterus espinhacensis Costa and Katz sp. nov., Trichomycterus macrophthalmus Barbosa and Costa, 2012, and T. travassosi); 14–16 ventral procurrent caudal-fin rays (vs. 8–13 in T. altipombensis, T. astromycterus, T. auroguttatus, T. goeldii, Trichomycterus jacupiranga Wosiacki and Oyakawa, 2005, T. saquarema, T. travassosi, and T. vinnulus); 28–33 premaxillary teeth (vs. 39–60 in T. auroguttatus, Trichomycterus caratinguensis Costa, Katz, and Vilardo sp. nov., Trichomycterus castelensis Costa, Katz, and Vilardo sp. nov., T. goeldii, T. macrophthalmus, T. mimosensis, Trichomycterus puriventris Barbosa, and Costa, 2012, T. saquarema, and T. vinnulus); 8 pectoral-fin rays (vs. 7 in Trichomycterus gasparinii Barbosa, 2013 and T. pantherinus); 36–38 vertebrae (vs. 33–35 in T. astromycterus, T. jacupiranga, T. mimosensis, and T. travassosi); 12–13 ribs (vs. 10–11 in T. alternatus, T. altipombensis, T. astromycterus, T. auroguttatus, T. caudofasciatus, T. espinhacensis Costa and Katz sp. nov., T. jacupiranga, T. macrophthalmus, T. travassosi, and T. vinnulus; and 14 in T. caratinguensis Costa, Katz, and Vilardo sp. nov.); the dorsal-fin origin at a vertical between the centrum of the 18th and 20th vertebra (vs. between the centrum of the 16th and 17th vertebra in T. altipombensis, T. auroguttatus, T. caratinguensis Costa, Katz, and Vilardo sp. nov., T. caudofasciatus, T. espinhacensis Costa and Katz sp. nov., T. macrophthalmus, and T. travassosi, and between the centrum of the 14th and 15th vertebra in T. astromycterus); pointed mandibular teeth (vs. incisor-like in T. astromycterus and T. jacupiranga); the presence of the anterior segment of the infraorbital series (vs. absence in T. gasparinii and T. pantherinus); and presence of a broad dark brown stripe between the humeral region and the caudal-fin base (vs. absence in T. alternatus, T. altipombensis, T. astromycterus, T. auroguttatus, T. caudofasciatus, T. espinhacensis Costa and Katz sp. nov., T. gasparinii, T. goeldii, T. macrophthalmus, T. mimosensis, T. travassosi, and T. vinnulus). Trichomycterus brigadeirensis may also be distinguished from all other species of PAC using its colour pattern, in which LDPZ 1 and 2 are represented using brown round spots longitudinally in close proximity, weakly distinguishable from colour ground (vs. LDPZ 1 and 2 marks highly contrasting with lighter colour ground).
Description. General morphology: Morphometric data are in
Table 1. The dorsal surface of the head is plain between orbits, the eye slightly projected dorsally. Maxillary barbel posteriorly reaching pectoral-fin base, rictal barbel reaching between the middle and posterior portion of an interopercular patch of odontodes, nasal barbel reaching the area just anterior to opercle or barely touching it. Jaw teeth pointed. Premaxillary teeth 28–33, slightly curved, arranged in irregular rows. Dentary teeth 32–39, slightly curved backwards, irregularly arranged. Opercular odontodes 13–17, interopercular odontodes 35–37. Odontodes pointed, arranged in irregular rows. Anterior infraorbital canal present. Single median s6. Dorsal and anal fins are subtriangular, anterior margin straight, posterior margin slightly convex. Total dorsal-fin rays 12 (iii + II + 7), total anal-fin rays 9 or 10 (ii–iii + II + 5). Anal-fin origin at vertical through posterior portion of dorsal-fin base, at base of 6th bifid dorsal-fin ray. Pectoral-fin filament length is about 25–40% of pectoral-fin length without filament. Total pectoral-fin rays 8 (I + 7). Pelvic fin posteriorly reaching vertical just posterior to middle dorsal-fin base. Pelvic-fin bases are medially separated by interspace about three-fourths pelvic-fin base. Total pelvic-fin rays 5 (I + 4). Caudal fin sub-truncate, upper and lower corners rounded. Total principal caudal-fin rays 13 (I + 11 + I), total dorsal procurrent rays 19–21 (xviii–xx + I), total ventral procurrent rays 14–16 (xiii–xv + I). Anus and urogenital papilla at vertical through area just anterior to middle of dorsal-fin base.
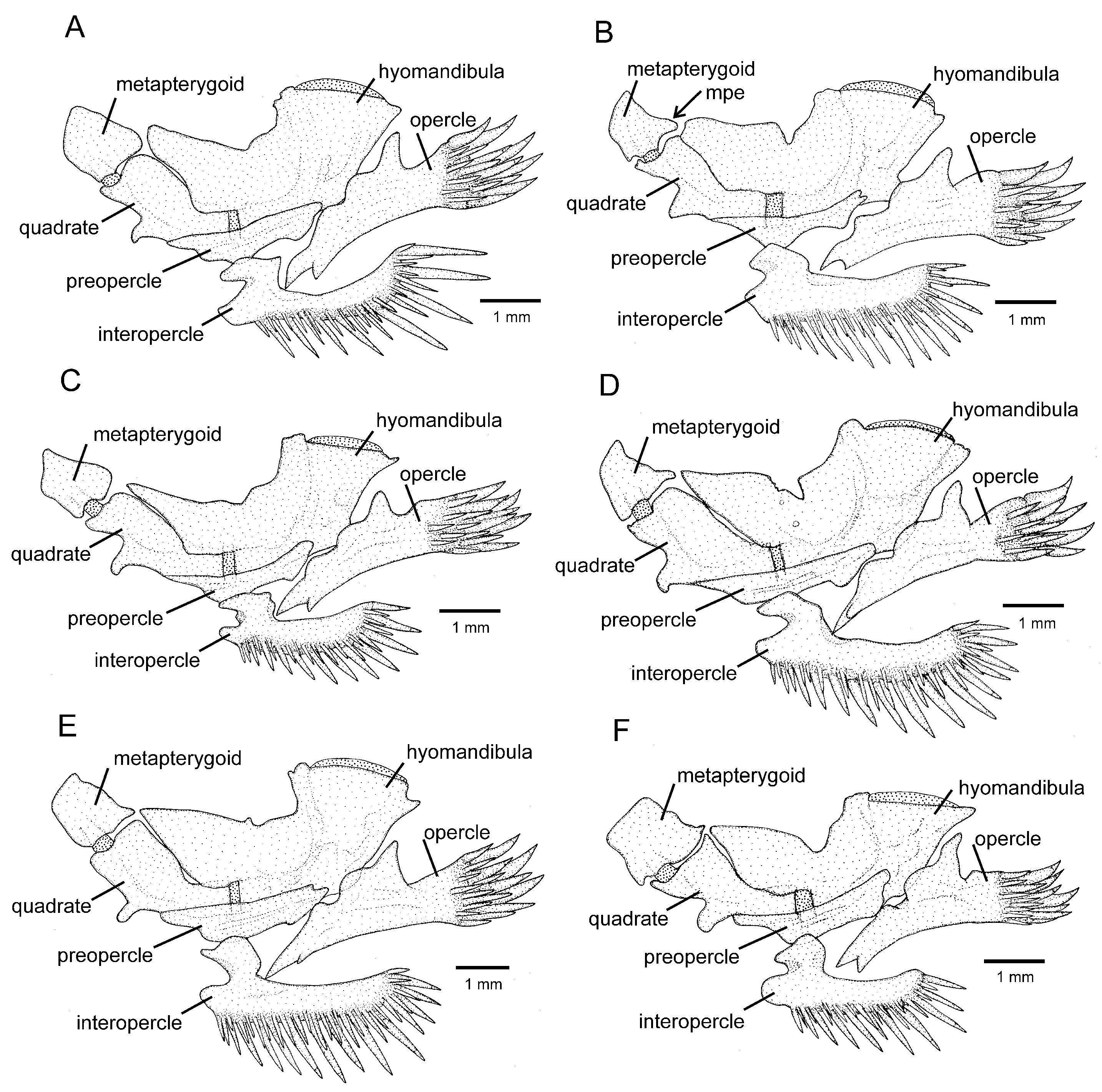
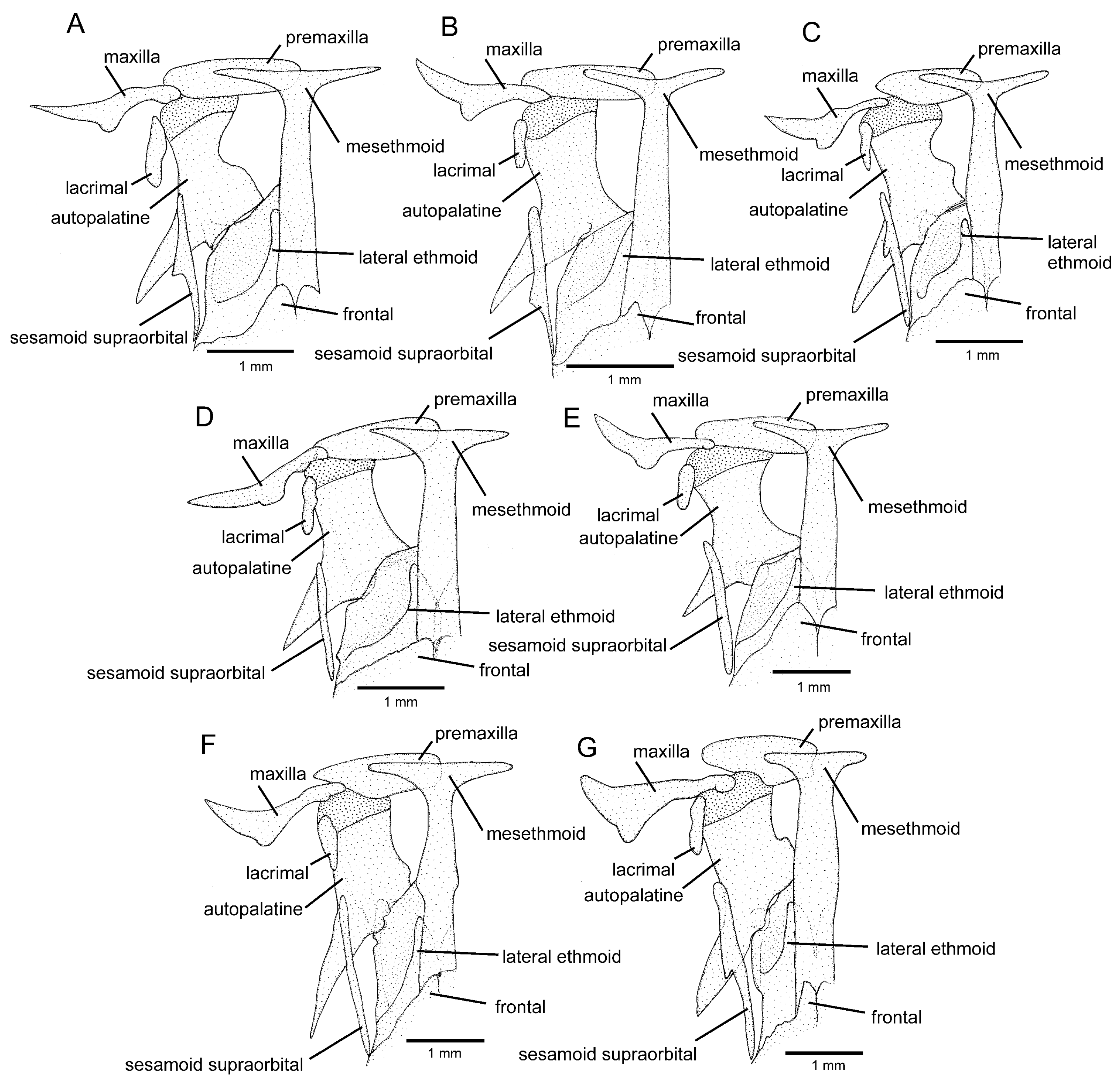
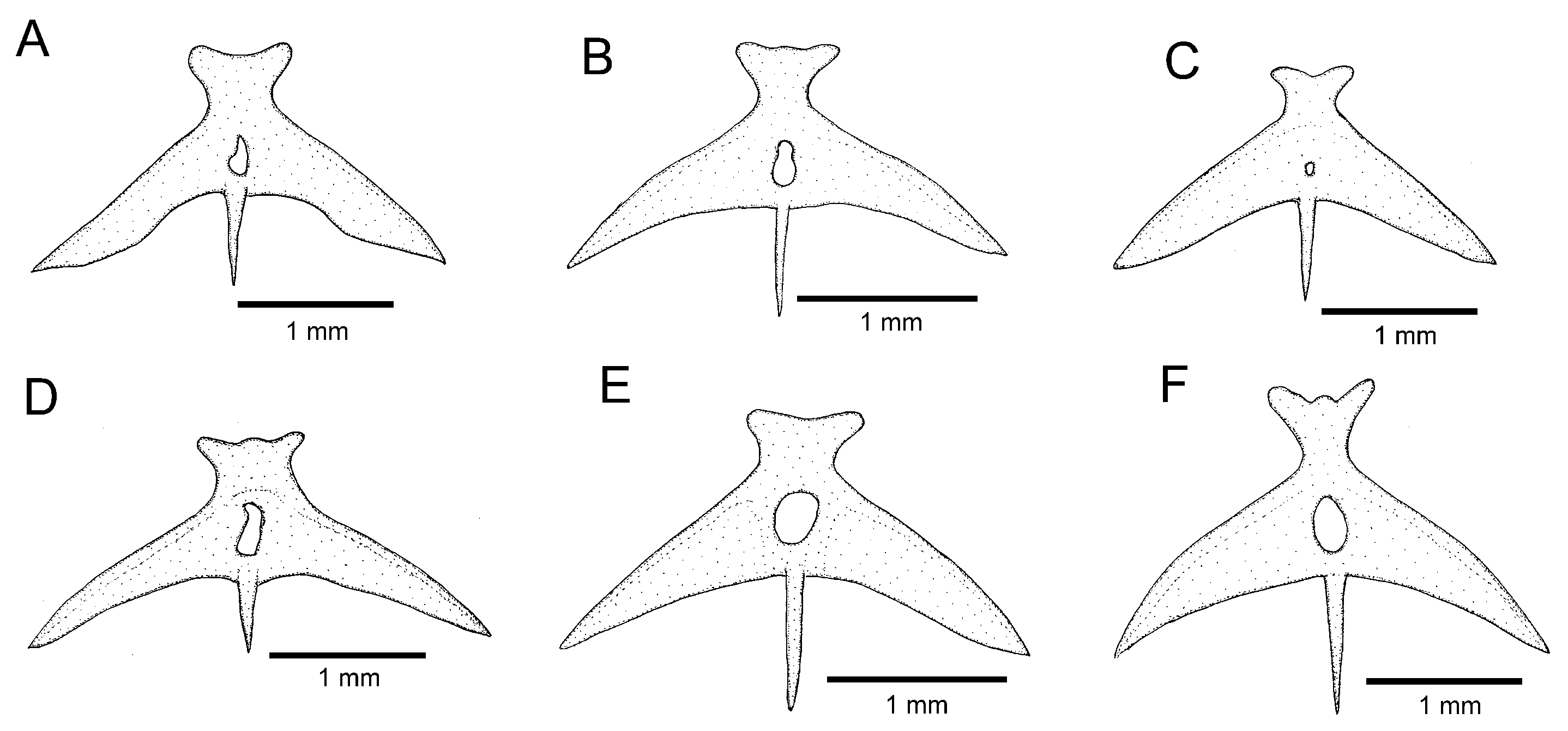
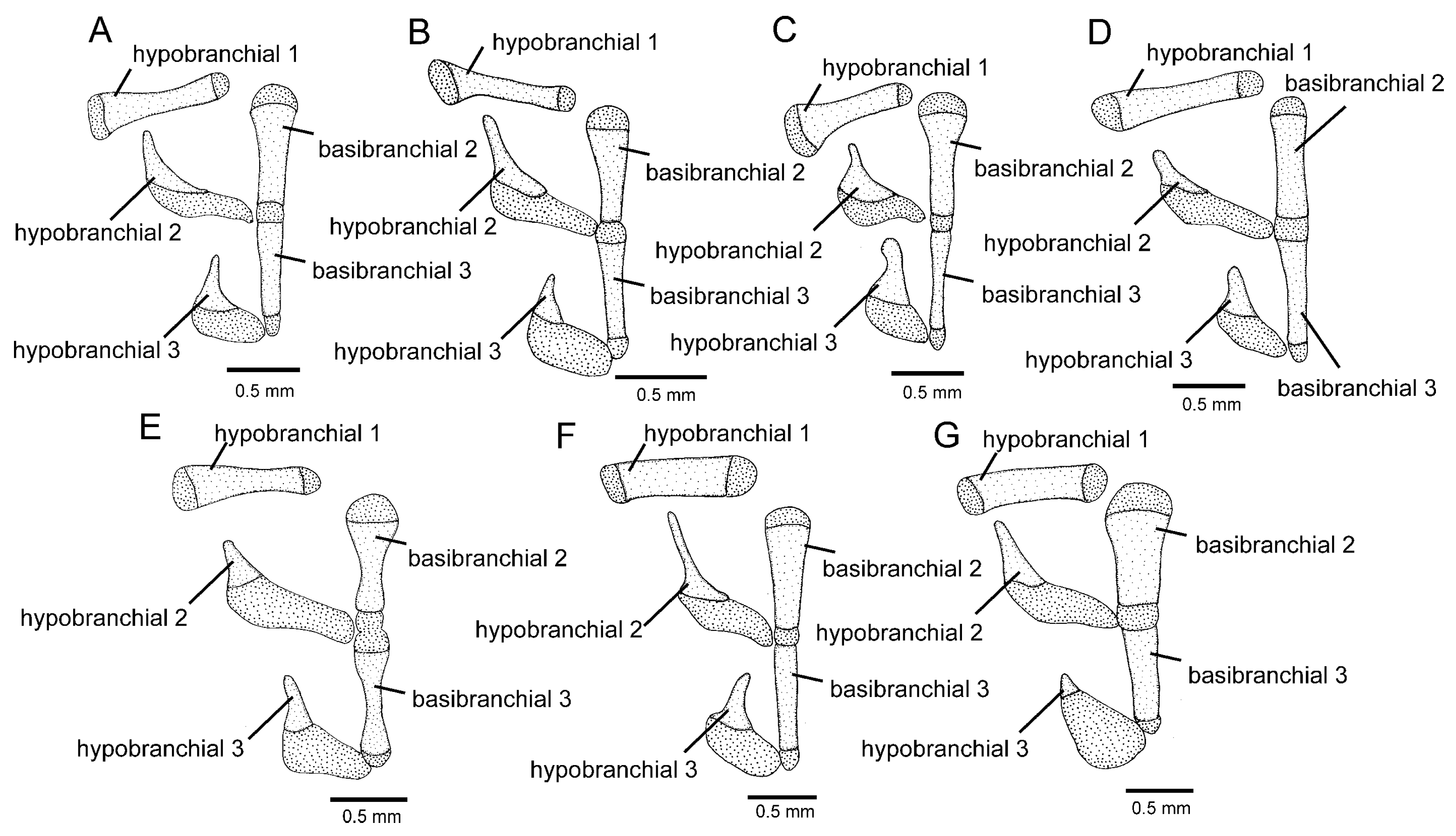
Osteology (
Figure 3A,
Figure 5A,
Figure 6A and
Figure 7A): Anterior margin of mesethmoid slightly concave, mesethmoid cornu rod-shaped. Lacrimal broad and flat, nearly elliptical; sesamoid supraorbital narrow and flat, with lateral expansion; sesamoid supraorbital length about two times lacrimal length. The premaxilla is sub-rectangular in the dorsal view. Maxilla boomerang-shaped, slender, slightly longer than premaxilla, with minute anterior expansion on subproximal region, posterior process well-developed. Autopalatine sub-rectangular in dorsal view when excluding posterolateral process, with pronounced lateral constriction posteriorly followed by weak sinuosity, shortest width about one-third of autopalatine length, lateral margin slightly convex. The postero-lateral process of autopalatine is subtriangular and long; its length is about three-fourths of autopalatine length. Metapterygoid subtrapezoidal, slightly deeper than long. Quadrate robust, dorsoposterior outgrowth in separated by minute interspace from hyomandibular outgrowth. Hyomandibula long, anterior outgrow shallow. The opercle is moderately elongated, the depth of the opercular odontode patch about four-fifths of the dorsal articular facet of the hyomandibula, and the dorsal process of the opercle is short and blunt. Interopercle long, about four-fifths of hyomandibula length. Preopercle compact, without ventral expansion. Basibranchials 2 and 3 elongate without distinctive constrictions; basibranchial 2 is an anterior region slightly wider than the posterior one, and basibranchial is rod-shaped. Hypobranchial 1 is a long, distal extremity slightly broader than the proximal extremity. Hypobranchials 2 and 3 are subtriangular, about equal in length; the basibranchial 2 is anterolaterally directed; the basibranchial 3 is anteriorly directed. Parurohyal robust, lateral process relatively elongate, pointed; parurohyal head well-developed, with prominent anterolateral paired process; middle foramen elliptical; posterior process short, about half length of distance between anterior margin of parurohyal and anterior insertion of posterior process. Branchiostegal rays 6 or 7. Vertebrae 36–38. Ribs 12–13. Dorsal-fin origin at vertical between centrum of 18th and 20th vertebra; anal-fin origin at vertical between centrum of 22nd and 23rd vertebra. Two dorsal hypural plates, corresponding to hypurals 4 + 5 and 3, respectively; single ventral hypural plate corresponding to hypurals 1 and 2 and parhypural.
Colouration in alcohol (
Figure 4): The ground colour of the dorsum, dorsal portion of flank and head side is yellowish brown, becoming lighter on the ventral part of flank and head side. LDPZ 1 and 2 are represented by brown round spots longitudinally in close proximity, weakly distinguishable from colour ground. Dark brown chromatophores concentrated on a vertical zone on the posterior-most part of the caudal peduncle. LDPZ 3 is represented by a broad dark brown stripe between the humeral region and caudal-fin base, sometimes with short interruptions on its posterior portion. LDPZ 4 is represented by dark brown melanophores more concentrated on the posterior half of the flank, often coalesced, forming irregularly shaped marks. The dorsal and lateral surfaces of the head have a great concentration of dark brown spots, making it difficult to delimitate infraorbital, interopercular, and supraopercular marks. Venter white. Nasal barbel dark brown, maxillary and rictal barbels light grey. Fins hyaline, with small black spots on the basal portion of the pectoral and unpaired fins. In specimens below about 40 mm SL, LDPZ 1–4 are represented by small black spots, and LDPZ 3 often forms a narrow black longitudinal line.
Colouration in life: Similar to colouration in alcohol.
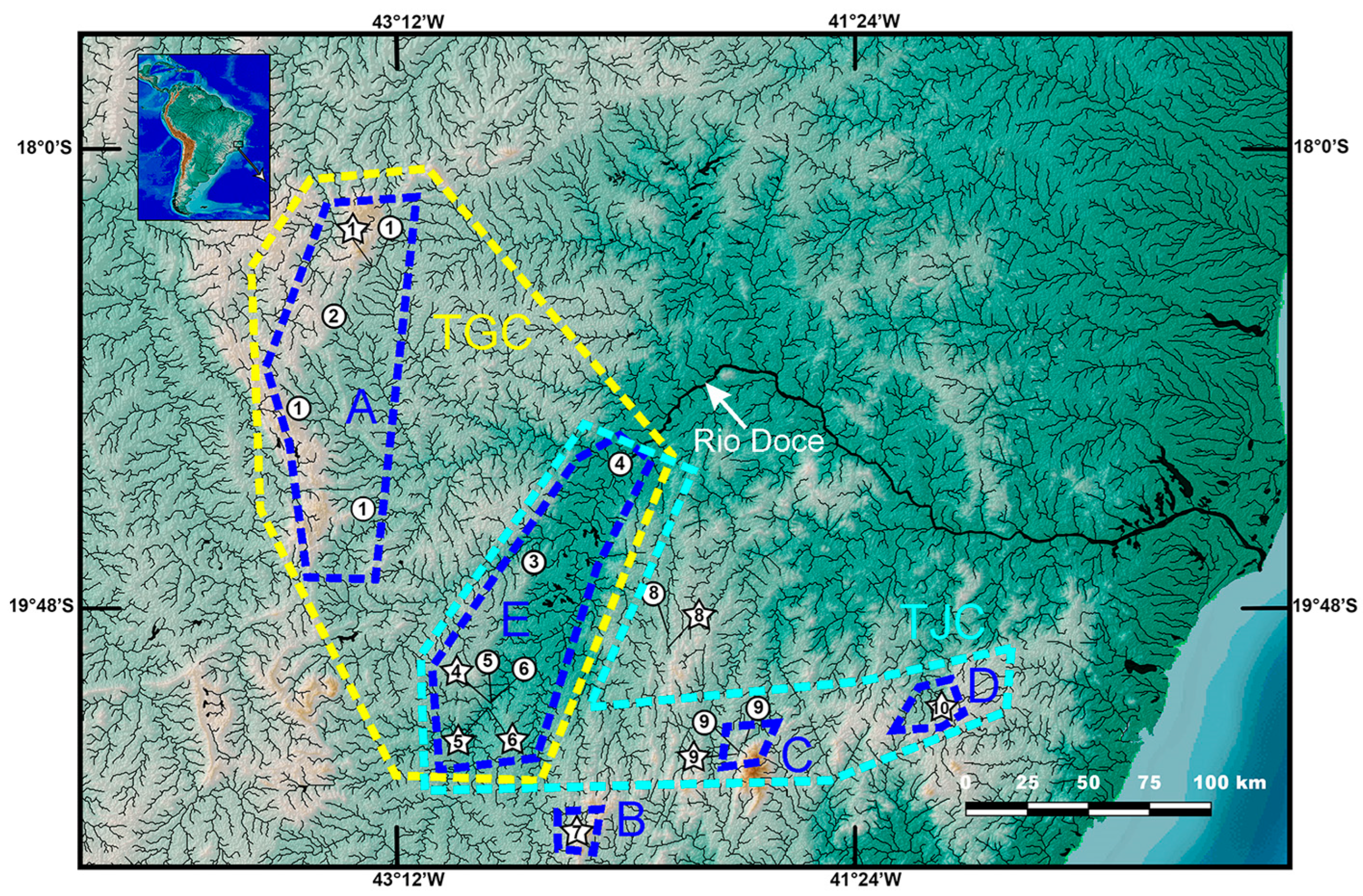
Distribution and habitat. Trichomycterus brigadeirensis is known only from the type locality, the Ribeirão Félix, a fast-flowing stream in the upper Rio Casca drainage, Rio Doce basin, Serra do Brigadeiro, southeastern Brazil (
Figure 8). Specimens were collected at an altitude of about 1100 m, where the Ribeirão Félix is about 1.5–3.0 m wide and about 0.3–1.2 m deep, with clear water, bottom comprising sand and small pebbles about 10–15 cm of diameter, and secondary vegetation on stream margins. Specimens were found under small pebbles.
Etymology. The name brigadeirensis refers to the occurrence of the new species in the Serra do Brigadeiro.
3.3. The Clade Trichomycterus caratinguensis Costa, Katz and Vilardo sp. nov. plus the T. goeldii Complex
A clade comprising
T. caratinguensis Costa, Katz and Vilardo sp. nov. and the
T. goeldii complex was weakly supported in phylogenetic analyses (
Figure 2) and not supported by any unique morphological character state. Therefore, the phylogenetic position of that new species is uncertain. The presence of a dark brown stripe between the humeral region and the caudal-fin base in
T. caratinguensis Costa, Katz and Vilardo sp. nov. (
Figure 9) and in both
T. brigadeirensis (
Figure 4), another species with basal phylogenetic position in PAC, would suggest that these species may be closest relatives. However, support values for the clade, including
T. caratinguensis Costa, Katz and Vilardo sp. Nov. and all other species of PAC except
T. brigadeirensis are high (
Figure 2), thus making the hypothesis of these two species being sistnr taxa improbable. On the other hand,
T. caratinguensis Costa, Katz and Vilardo sp. nov. possesses a small posterior expansion in the metapterygoid (
Figure 3B), a condition considered diagnostic for a clade including all species of PAC except those belonging to the
T. goeldii complex [
12]. Therefore, although a similar expansion may occur in other generic trichomycterine lineages, making polarization of this character uncertain, the hypothesis of
T. caratinguensis Costa, Katz and Vilardo sp. nov. to be sister to the well-supported clade comprising the
T. jacupiranga,
T. puriventris, and
T. travassosi complex cannot be discarded.
Trichomycterus caratinguensis Costa, Katz and Vilardo sp. nov.
LSID:urn:lsid:zoobank.org:act:CC19E25F-4730-4342-9413-26BEE380A23E
Trichomycterus alternatus non
T. alternatus (Eigenmann, 1917): figure 3H and 4A–C in Ref. [
6]: (misidentification).
Holotype. UFRJ 13815, 55.3 mm SL; Brazil: Minas Gerais State: Santa Bárbara do Leste Municipality: Córrego Tabuleiro II, a tributary of Rio Caratinga, Rio Doce basin, 19°55′53″ S 42°08′27″ W, about 680 m asl; A. M. Katz and P. J. Vilardo, 19 May 2023.
Paratypes. All from Brazil: Minas Gerais State: Córrego Tabuleiro II, a tributary of Rio Caratinga, Rio Doce basin. Santa Bárbara do Leste Municipality: UFRJ 13816, 3 ex., 30.7–40.6 mm SL (C&S); UFRJ 13817, 4 ex., 23.4–28.9 mm SL; collected with holotype. Santa Rita de Minas Municipality: UFRJ 13238, 11 ex., 26.5–33.7 mm SL; CICCAA 07773, 4 ex., 29.4–32.3 mm SL; 19°54′06″ S 42°08′08″ W, altitude about 660 m asl; A. M. Katz and P. J. Vilardo, 22 September 2022.
Diagnosis. Trichomycterus caratinguensis is distinguished from all other species of PAC by having a unique colour pattern combining LDPZ 1–3 represented by dark brown parallel stripes, with the LDPZ 2 and 3 stripes slightly converging on the caudal peduncle end (
Figure 9; vs. never a similar colour pattern).
Trichomycterus caratinguensis also differs from other congeners of PAC using the following combination of character states: 18–20 dorsal procurrent caudal-fin rays (vs. 22–28 in
T. altigrandensis;
T. astromycterus,
T. espinhacensis Costa and Katz sp. nov.,
T. macrophthalmus, and
T. travassosi); eight pectoral-fin rays (vs. seven in
T. gasparinii and
T. pantherinus); S6 pore paired, pores separated by interspace (vs. single S6 median pore, sometimes two pores in close proximity in
T. alternatus,
T. altipombensis,
T. auroguttatus,
T. astromycterus,
T. brigadeirensis,
T. castelensis,
T. caudofasciatus,
T. goeldii,
T. jacupiranga,
T. macrophthalmus,
T. mimosensis, and
T. travassosi); presence of the anterior section of the infraorbital canal present (vs. absent in
T. gasparinii and
T. pantherinus); pointed mandibular teeth (vs. incisor-like in
T. astromycterus and
T. jacupiranga); 39–45 premaxillary teeth (vs. 25–34 in
Trichomycterus brigadeirensis Costa, Katz and Vilardo sp. nov.,
Trichomcyterus caparaoensis Costa, Barbosa and Katz sp. nov.,
T. caudofasciatus, and
T. travassosi); a long nasal barbel, its tip reaching the opercle (vs. moderate in length or short, tip-reaching area anterior to the opercle in
T. alternatus,
T. altipombensis,
T. castelensis,
T. macrophthalmus,
T. mimosensis,
T. saquarema,
T. travassosi, and anterior to orbit in
T. astromycterus); sesamoid supraorbital bone with a lateral expansion (vs. without lateral projections and processes in
T. caudofasciatus,
T. puriventris,
T. mimosensis,
T. caparaoensis Costa, Barbosa and Katz sp. nov.,
T. castelensis Costa, Katz and Vilardo sp. nov., and
T. vinnulus), and 14 ribs (vs. 9–12 in
T. alternatus,
T. altipombensis,
T. astromycterus,
T. auroguttatus,
T. caudofasciatus,
T. espinhacensis Costa and Katz, sp. nov.,
T. gasparini,
T. goeldii,
T. jacupiranga,
T. caudofasciatus,
T. mimosensis,
T. macrophthalmus,
T. pantherinus,
T. travassosi, and
T. vinnulus).
Description. General morphology: Morphometric data are in
Table 2. The dorsal surface of the head is nearly plain between orbits, with the eye not projected dorsally. Maxillary barbel posteriorly reaching area between an interopercular patch of odontodes and pectoral-fin base, rictal barbel reaching the middle portion of the interopercular patch of odontodes, nasal barbel reaching area just anterior to opercle. Jaw teeth pointed. Premaxillary teeth 39–45, slightly curved, arranged in irregular rows; dentary teeth 32–41, slightly curved backwards, irregularly arranged. Opercular odontodes 14–18, interopercular odontodes 34–40. Odontodes pointed, arranged in irregular rows. Anterior infraorbital canal present. s6 pore paired, nearer its symmetrical homologous pair than orbit. Dorsal and anal fins are subtriangular, margins slightly convex. Total dorsal-fin rays 11 or 12 (ii–iii + II + 7), total anal-fin rays 9 or 10 (ii–iii + II + 5). Anal-fin origin at vertical through dorsal-fin base posterior end. Pectoral-fin filament length is about 30% of pectoral-fin length without filament. Total pectoral-fin rays 8 (I + 7). Pelvic fin posteriorly reaching vertical through middle of dorsal-fin base. Pelvic-fin bases are medially separated by interspace about equal pelvic-fin ray width. Total pelvic-fin rays 5 (I + 4). Caudal fin truncate. Total principal caudal-fin rays 13 (I + 11 + I), total dorsal procurrent rays 18–20 (xvii–xix + I), total ventral procurrent rays 13 or 14 (xii–xiii + I). Anus and urogenital papilla at vertical through anterior half of dorsal-fin base.
Osteology (
Figure 3B,
Figure 5B and
Figure 6B): Anterior margin of mesethmoid concave, mesethmoid cornu rod-shaped. Lacrimal and sesamoid supraorbital narrow and flat, sesamoid supraorbital length about three times lacrimal length, with weak lateral expansion. The premaxilla is sub-rectangular in the dorsal view. Maxilla is boomerang-shaped, slender, slightly longer than premaxilla, with a small posterior process. Autopalatine sub-rectangular in dorsal view when excluding posterolateral process, its shortest width about half autopalatine length, lateral and medial margins slightly convex. The postero-lateral process of autopalatine is subtriangular; its length is about two-thirds of autopalatine length. Metapterygoid subtriangular, slightly deeper than long. Quadrate robust, dorsoposterior outgrowth in contact with hyomandibular outgrowth. Hyomandibula long, anterior outgrow with a small concavity in its dorsal margin. The opercle is moderately elongated, the depth of the opercular odontode patch about four-fifths of the dorsal articular facet of the hyomandibula, and the dorsal process of the opercle is short and blunt. Interopercle long, about three-fourths of hyomandibula length. Preopercle compact, with small ventral expansion. Basibranchials 2 and 3 elongate without distinctive constrictions. Hypobranchial 1 is long, distal extremity slightly broader than the proximal extremity. Hypobranchials 2 and 3 are subtriangular and about equal in length. Parurohyal robust, lateral process relatively elongate, pointed, slightly curved; parurohyal head well-developed, with the prominent anterolateral paired process; middle foramen round, moderate; posterior process long, about four-fifths of the distance between anterior margin of parurohyal and anterior insertion of posterior process. Branchiostegal rays 7. Vertebrae 36 or 37. Ribs 14. Dorsal-fin origin at vertical through centrum of 16th or 17th vertebra; anal-fin origin at vertical through centrum of 21st or 22nd vertebra. One or two dorsal hypural plates, corresponding to hypurals 4 + 5 and 3, respectively; single ventral hypural plate corresponding to hypurals 1 and 2 and parhypural.
Colouration in alcohol (
Figure 9): The ground colour of the flank and head is light yellowish grey. LDPZ 1–3 represented by dark brown parallel stripes. LDPZ-4 is represented by dispersed dark brown chromatophores. LDPZ 2, 3 and 4 slightly converge on the caudal peduncle end. Venter white. The dorsal surface of the head is dark brown, and the lateral surface is pale grey with a great concentration of dark brown spots. Nasal and maxillary barbels light brown, rictal barbel light grey. Fins hyaline, with small black spots on the basal portion of pectoral, dorsal and caudal fins. Specimens below 40 mm SL LDPZ 1–3 are represented by a longitudinal row of horizontally elongated black spots.
Colouration in life: Similar to colouration in alcohol, but with a concentration of yellow pigment on fin bases and head side.
Distribution and habitat. Trichomycterus caratinguensis is known only from streams draining a plateau about 650–700 m asl in the Rio Caratinga drainage, Rio Doce basin (
Figure 8). The type locality is situated in a region where all the original vegetation cover has been lost, currently containing pastures, plantations and small urban centres, with rivers and streams showing strong signs of pollution. Where the species was found, Córrego Tabuleiro II is about 1.5–3.0 m wide, about 0.3 m deep, with a sandy bottom.
Etymology. The name caratinguensis is an allusion to this new species being presently only known from the Rio Caratinga drainage.
3.4. The Trichomycterus goeldii Complex
The
T. goeldii complex, as delimited in Costa et al. [
12], but also including a new species described below, is corroborated here. This clade has been diagnosed using three apomorphic features: 1—the presence of a long and robust postero-lateral process of the autopalatine (figure 2F in Ref. [
10]; figure 2A,B in Ref. [
12]); 2—a slightly folded maxilla (figure 2F in Ref. [
10]; figure 2A,B in Ref. [
12]); and 3—anterior cranial fontanel represented using a minute aperture (figure 9B in Ref. [
10]). Character states 1 and 2 are mostly observable in specimens above about 50 mm SL, in which the postero-lateral process of the autopalatine is broad and long, its length approximately equal or slightly longer than the autopalatine longitudinal length excluding the process and the anterior cartilage. Below this size, the anterior portion of the head is relatively shorter and bones of this region may have a different shape, including a slightly shorter and more slender postero-lateral process of the autopalatine and a less folded maxilla, making it difficult to evaluate the occurrence of character states 1 and 2 in species with only specimens below that size available for osteological examination. In addition, a long autopalatine postero-lateral process is also present in the more distantly related
T. vinnulus (
Figure 5F), here considered as homoplastic conditions. The two apomorphic conditions are here confirmed to occur in all species examined of the
T. goeldii. On the other hand, character state 3 is present in most specimens examined, but some variation precludes this condition as diagnostic for the clade.
This study corroborates two main lineages within the
T. goeldii complex. The first lineage includes nominal species and populations with a conspicuous level of morphological divergence but low molecular differentiation. This geographically widespread clade (hereafter the W-clade) includes
T. auroguttatus and
T. goeldii from the Rio Paraíba do Sul basin and populations with indeterminate taxonomical status from the upper Rio Grande drainage, upper Rio Paraná basin, and Rio Muriaé and Rio Pomba drainages of the lower Rio Paraíba do Sul basin. This clade also includes an unnamed taxon (
Figure 1) from the upper Rio Santo Antônio, RDB (
Figure 8), represented in our collections by only four small specimens (UFRJ 13103, 2 ex.; UFRJ 13121, 2 ex. C&S). Due to the small available specimen sample, this taxon is not described here. It is identified as
Trichomycterus sp. aff.
T. goeldii and is supported as a sister to
T. auroguttatus and
T. goeldii (
Figure 2).
The second lineage of the
T. goeldii complex is endemic to the RDB (hereafter RDB1-clade) and is highly supported by molecular data (
Figure 1), but no morphological apomorphy was found to be uniquely shared by species of this clade. The Doce-clade includes
T. alternatus and
T. astromycterus, two sympatric species supported as sister taxa, and
T. espinhacensis Costa and Katz sp. nov. from the Rio Santo Antônio drainage, which is supported as sister to all other species of the Doce-clade. This clade also includes an unnamed species (figure 8 in Ref. [
12]) previously identified as
T. alternatus [
12] from the lower Rio Piracicaba drainage (
Figure 8). Additional material is still necessary to provide a formal description of this taxon, which is here identified as
Trichomycterus sp. aff.
T. alternatus, supported as sister to the clade comprising
T. alternatus and
T. astromycterus (
Figure 2). Whereas
T. espinhacensis Costa and Katz sp. nov. occurs in streams draining the Serra do Espinhaço, in altitudes between about 680–715 m asl, the other taxa of the RDB1-clade are found in lower altitude places, between about 280 and 350 m asl, close to the main course of the Rio Doce (
Figure 8).
Trichomycterus espinhacensis Costa and Katz sp. nov.
LSID:urn:lsid:zoobank.org:act:1C174B5C-12E8-4BDF-AF24-8C7AF0EE668E
Holotype. UFRJ 13483, 46.8 mm SL; Brazil: Minas Gerais State: Santo Antônio do Imbé Municipality: Rio Preto, tributary of Rio Guanhães, Rio Santo Antônio drainage, Rio Doce basin, 18°27′49″ S 43°18′26″ W, altitude about 715 m; W. J. E. M. Costa et al., 16 August 2022.
Paratypes. All from Brazil: Minas Gerais State: Rio Santo Antônio drainage, Rio Doce basin. Santo Antônio do Imbé Municipality: UFRJ 13118, 11 ex., 32.7–51.2 mm SL; UFRJ 13484, 3 ex. (C&S), 33.1–44.0 mm SL; UFRJ 13105, 3 ex. (DNA), 26.6–33.2 mm SL; collected with holotype. UFRJ 13123, 7 ex., 27.7–46.4 mm SL; CICCAA 07774, 3 ex., 40.1–40.4 mm SL; Rio Guanhães, 18°27′40″ S 43°17′46″ W, altitude about 700 m asl; W. J. E. M. Costa et al., 17 August 2022. Conceição do Mato Dentro Municipality: UFRJ 13478, 56.1 mm SL; UFRJ 13119, 17 ex., 34.9–47.6 mm SL; UFRJ 13120, 3 ex. (C&S), 42.8–47.0 mm SL; UFRJ 13108, 2 ex. (DNA), 40.5–48.3 mm SL; Córrego da Guinda, 19°00’54″ S 43°35’07″ W, altitude about 690 m; W. J. E. M. et al., 10 August 2022. UFRJ 13135, 2 ex., 24.8–30.9 mm SL; Cachoeira Três Barras, 19°05’58″ S 43°29’13″ W, altitude about 695 m; W. J. E. M. et al., 10 August 2022.
Additional material (non-type specimens). UFRJ 13479, 49.8 mm SL; UFRJ 13122, 9 ex., 32.7–44.2 mm SL; UFRJ 13117, 3 ex. (C&S), 39.2–47.8 mm SL; UFRJ 13104, 5 ex. (DNA), 24.1–35.0 mm SL; Brazil: Minas Gerais State: Itambé do Mato Dentro: Rio Preto do Itambé, just below Cachoeira do Lúcio, Rio Santo Antônio drainage, Rio Doce basin, 19°24’50″ S 43°19’49″ W, altitude about 680 m; W. J. E. M. et al., 11 August 2022.
Diagnosis. Trichomycterus espinhacensis is distinguished from all other congeners of the
T. goeldii complex, except
T. astromycterus, by having 23–26 dorsal procurrent caudal-fin rays (vs. 17–19).
Trichomycterus espinhacensis differs from
T. astromycterus by having: a relatively long and thin mesethmoid cornu (
Figure 5C; vs. short and robust,
Figure 5G; see also figure 4 in Ref. [17l, 2020]); a relatively shorter maxilla, slightly longer than the premaxilla, without a distal widening (
Figure 5C; vs. maxilla about one and half time longer than the premaxilla, with distinctive distal widening,
Figure 5G; see also figure 3 in Ref. [
17]); well-developed third hypobranchial ossification (
Figure 7C; vs. rudimentary,
Figure 7G); 35 or 36 vertebrae (vs. 33 or 34); the dorsal-fin origin at a vertical through the centrum of 17th vertebra (vs. 14th or 15th vertebra); a longer nasal barbel, its tip posteriorly reaching the opercle or the area just anterior to it (vs. reaching the area anterior to the orbit); and caudal fin subtruncate (vs. emarginate).
Trichomycterus espinhacensis is also distinguished from all other species of the
T. goeldii complex by having a rudimentary foramen on the parurohyal (
Figure 6C; vs. well-developed) and a narrow, rod-shaped basibranchial 3 (vs.
Figure 7C; vs. never rod-shaped, figure 3 in Ref. [
12]).
Trichomycterus espinhacensis also differs from
T. auroguttatus and
T. goeldii by having 36–39 premaxillary teeth (vs. 42–45), absence of a robust comma-shaped osseous core in the autopalatine articular shell for the lateral ethmoid (vs. presence; figure 2A,B in Ref. [
12]), and a relatively narrow and long second hypobranchial (
Figure 7C; vs. broad and short, figure 3A in Ref. [
12]); and from
T. alternatus by having a row of variably shaped small pale brown spots on LDPZ 3 (vs. row of black round spots, figure 2 in Ref. [
6]).
Description. General morphology: Morphometric data are in
Table 3. The dorsal surface of the head between orbits is plain to slightly concave, with the eye slightly projected dorsally. Maxillary barbel posteriorly reaching pectoral-fin base or area just anterior to it, rictal barbel reaching area between the middle and posterior part of an interopercular patch of odontodes, the tip of nasal barbel reaching opercle or area just anterior to it. Jaw teeth are pointed, slightly curved, irregularly arranged; premaxillary teeth 37–39; dentary teeth 29–33. Opercular odontodes 15–17, interopercular odontodes 30–33. Odontodes pointed, arranged in irregular rows Branchiostegal rays 7. s6 pore nearer to its symmetrical homologous s6 pore than to orbit. Dorsal and anal fins are subtriangular, anterior margin about straight, posterior margin slightly convex. Total dorsal-fin rays 11 or 12 (iii + II + 6–7), total anal-fin rays 10 (iii + II + 5). Anal-fin origin at vertical through the posterior part of dorsal-fin base, through the base of 5th or 6th branched dorsal-fin ray. Dorsal-fin origin at vertical through centrum of 17th vertebra; anal-fin origin at vertical through centrum of 21st vertebra. Pectoral-fin filament length is about 10–25% of pectoral-fin length without filament. Total pectoral-fin rays 8 (I + 7). Pelvic fin posteriorly reaching vertical at middle dorsal-fin base. Pelvic-fin bases are medially separated by small interspace. Total pelvic-fin rays 5 (I + 4). Caudal fin subtruncate, upper and lower corners rounded. Total principal caudal-fin rays 13 (I + 11 + I), total dorsal procurrent rays 24–26 (xxiii–xxv + I), total ventral procurrent rays 14 or 15 (xiii–xiv + I). Anus and urogenital papilla at vertical through dorsal-fin origin or immediately anterior to it.
Osteology (
Figure 3C,
Figure 5C and
Figure 6C): Anterior margin of mesethmoid slightly concave, mesethmoid cornu rod-shaped. Lacrimal and sesamoid supraorbital narrow, rod-like, sesamoid supraorbital length about two times and half lacrimal length. The premaxilla is sub-rectangular in dorsal view, slightly tapering laterally. Maxilla boomerang-shaped, slender, longer than premaxilla, slightly folded in its main axis, with small projection on anteromedian margin and small posterior process. Autopalatine sub-rectangular in dorsal view when excluding posterolateral process, its shortest width about half autopalatine length, lateral margin about straight, medial margin sinuous, with deep concave. In the postero-lateral process of autopalatine subtriangular, its length is about equal to autopalatine length without anterior cartilage. Metapterygoid subtrapezoidal, deeper than long. Quadrate robust, dorsoposterior outgrowth in close proximity with hyomandibular outgrowth. Hyomandibula long, anterior outgrow with concave dorsal margin. The opercle is moderately elongated, the depth of the opercular odontode patch about three-fourths of the dorsal articular facet of the hyomandibula, and the dorsal process of the opercle is short and blunt. Interopercle long, about three-fourths of hyomandibula length. Preopercle compact, without ventral expansion. Basibranchials 2 and 3 elongate without distinctive middle constrictions; basibranchial 3 is rod-shaped. Hypobranchial 1 is long, distal extremity broader than proximal extremity. Hypobranchials 2 and 3 are subtriangular, about equal in length, and anterolaterally directed. Parurohyal robust, lateral process relatively elongate, pointed, slightly curved; parurohyal head well-developed, narrow, with prominent anterolateral paired process; middle foramen minute; posterior process moderate in length, about four-fifths distance between anterior margin of parurohyal and anterior insertion of posterior process. Vertebrae 35 or 36. Ribs 10 or 11. Two dorsal hypural plates, corresponding to hypurals 4 + 5 and 3, respectively; single ventral hypural plate corresponding to hypurals 1 and 2 and parhypural.
Colouration in alcohol (
Figure 10): The ground colour of the dorsum, flank and head side is light brownish yellow on the dorsal part, gradually becoming lighter ventrally. Flank and dorsum with variably shaped small pale brown spots, their diameter approximately equal, slightly larger or slightly smaller than opercular patch of odontodes. LDPZ 1 spots sometimes longitudinally coalesced to form a stripe, sometimes transversely coalesced with LDPZ 2 spots. LDPZ 3 usually has round spots forming regular series, sometimes irregularly arranged and irregularly shaped. LDPZ 4 with paler spots. Dark chromatophores are scattered over the dorsal portion of the head and dorsal part of the trunk. Head with small light brown dots. Venter white. Nasal and maxillary barbels light brown, rictal barbel light grey. Fins hyaline, with dark brown chromatophores on the basal portion.
Colouration in life: Similar to colouration in alcohol.
Distribution and habitat. Trichomycterus espinhacensis is known from the upper course of tributaries of the Rio Santo Antônio drainage, middle Rio Doce basin, Serra do Espinhaço, in altitudes between about 680 and 715 m asl (
Figure 8). This species was found to be associated with fine gravel bottom of clear water and fast-flowing streams. Specimens were collected above the bottom gravel.
Etymology. The name espinhacensis is an allusion to the occurrence of this new species in the Serra do Espinhaço, an important biodiversity centre of south-eastern Brazil.
3.5. The Trichomycterus jacupiranga Complex
The
T. jacupiranga complex as delimited in Costa et al. [
12], but also including a new species from the Serra do Caparaó below described, a new species from the Serra do Castelo below described, and
T. gasparinii, not included in the previous analysis, was corroborated maximum support values. According to the phylogenetic analysis (
Figure 2), the
T. jacupiranga complex is sister to a clade containing the
T. puriventris and
T. travassosi complexes; the
T. jacupiranga contains two subclades, one endemic to RDB (hereafter RDB2-clade), comprising
T. vinnulus, a new species from the Serra do Caparaó, and a new species from the Serra do Castelo, and one endemic to smaller coastal basins (hereafter the C-clade), comprising
T. jacupiranga and
T. gasparinii.
The RDB2-clade is morphologically supported by the presence of a hypertrophied foramen of the parurohyal (
Figure 6D–F), instead of being a smaller aperture as in other taxa of the
T. jacupiranga complex and other congeners (
Figure 6A–C), except species of a clade supported in Vilardo et al. [
14] including
Trichomycterus melanopygius Reis, Santos, Britto, Volpi and Pinna, 2020 and closely related species (see discussion below). Species of the RDB2-clade also share the presence of a peculiar morphology of the metapterygoid, in which the anterior region is slightly folded, resulting in an irregular quadrilateral shape (
Figure 3D–F), thus contrasting with the subtriangular metapterygoid present in other taxa of the
T. jacupiranga complex and the great majority of congeners (Figure 3 in Ref. [
10]). However, both character states relative to the metapterygoid (i.e., irregular quadrilateral and subtriangular shapes) occur in different lineages of
Psammocambeva, making it difficult to establish character state polarization without ambiguities. Species of the RDB2-clade also differ from species of the C-clade by the absence of a distinctive lateral process in the sesamoid supraorbital (
Figure 5D–F), which is present in both
T. gasparinii and
T. jacupiranga, and most other species congeners of PAC (
Figure 5A–C,G). The phylogenetic analysis supported the new species from the Serra do Castelo as sister to a clade including
T. vinnulus and the new species from the Serra do Caparaó (
Figure 2), but no morphological apomorphic condition uniquely shared by these two last species was found.
Trichomycterus caparaoensis Costa, Barbosa and Katz sp. nov.
Holotype. UFRJ 6006, 53.4 mm SL; Brazil: Minas Gerais State: Martins Soares Municipality: Córrego Feio, tributary of Rio José Pedro, Rio Manhuaçu drainage, Rio Doce basin, 20°23′37″ S 41°51′31″ W, altitude about 1015 m; W. J. E. M. Costa, B. B. Costa and C. P. Bove 24 December 2002.
Paratypes. All from Brazil: Minas Gerais State: Rio Manhuaçu drainage, Rio Doce basin. Martins Soares Municipality: UFRJ 5676, 8 ex., 34.8–47.6 mm SL; UFRJ 7881, 2 ex., 42.8–47.0 mm SL; UFRJ 5682, 4 ex. (C&S), 40.5–48.3 mm SL; CICCAA 07775, 2 ex., 43.7–46.0 mm SL; collected with holotype. UFRJ 7044, 43.0 mm SL; Brazil: Minas Gerais State: Martins Soares Municipality: Poço das Crianças, Rio José Pedro, Rio Manhuaçu drainage, Rio Doce basin, 20°22′28″ S 41°51′32″ W, about 875 m asl; W. J. E. M. Costa, B. B. Costa and C. P. Bove, 24 December 2002. Iuna Municipality: UFRJ 13172, 2 ex. (DNA), 30.7–37.0 mm SL; Rio José Pedro, 20°22′09″ S 41°51′28″ W, about 800 m asl; A. M. Katz and P. Vilardo, 22 September 2022.
Diagnosis. Trichomycterus caparaoensis differs from all congeners of the
T. jacupiranga complex by having the dorsal surface of the head between orbits slightly convex (vs. plain to slightly concave).
Trichomycterus caparaoensis is distinguished from other species of the RDB2-clade by a colour pattern of LDPZ 3 in specimens above about 45 mm SL, consisting of a dark brown stripe interrupted in its posterior portion (vs. a row of dark brown to black spots).
Trichomycterus caparaoensis is also distinguished from
T. vinnulus, its hypothesised sister species, by having: a longer pectoral-fin filament (filament length about 30–50% of the pectoral fin, vs. about 10–20%); a relatively long interopercular patch of odontodes, longer than the largest horizontal length of the hyomandibular anterior outgrown (
Figure 3D; vs. shorter,
Figure 3F), more ribs (13 vs. 11); the postero-lateral process of the autopalatine postero-laterally directed, its length about two thirds of the autopalatine length (
Figure 5D; vs. posteriorly directed, its length about equal autopalatine length,
Figure 5F); the second hypobranchial short, about so long as the third hypobranchial (
Figure 7D; vs. long, about twice longer than the third hypobranchial,
Figure 7F); and the posterior process of the parurohyal short, about half the distance between the anterior margin of the parurohyal and the anterior insertion of the posterior process (
Figure 6D; vs. about equal or slightly shorter,
Figure 6F).
Trichomycterus caparaoensis is also distinguished from
T. castelensis and
T. jacupiranga by having paired S6 pore (vs. a single S6 median pore) and fewer opercular odontodes (11–15 vs. 17–19); from
T. gasparinii by the presence of the anterior section of the infraorbital canal (vs. absence) and eight pectoral-fin rays (vs. seven); and from
T. jacupiranga by having pointed mandibular teeth (vs. incisor-like).
Description. General morphology: Morphometric data are in
Table 4. The dorsal surface of the head between orbits is slightly convex, with the eye not projected dorsally. Maxillary barbel posteriorly reaching pectoral-fin base, rictal barbel reaching posterior part of an interopercular patch of odontodes, the tip of nasal barbel reaching opercle. Jaw teeth pointed. Premaxillary teeth 25–35, slightly curved, arranged in irregular rows; dentary teeth 30–41, slightly curved backwards, irregularly arranged. Opercular odontodes 9–13, interopercular odontodes 31–42. Odontodes pointed, arranged in irregular rows. Anterior infraorbital canal present. s6 pore nearer to its symmetrical homologous s6 pore than to orbit. Dorsal and anal fins are subtriangular, anterior margin about straight, posterior margin slightly convex. Total dorsal-fin rays 12 (iii + II + 7), total anal-fin rays 10 (iii + II + 5); anal-fin origin at vertical through the posterior part of dorsal-fin base. Pectoral-fin filament length is about 30–50% of pectoral-fin length without filament. Total pectoral-fin rays 8 (I + 7). Pelvic fin posteriorly reaching vertical just posterior to middle dorsal-fin base. Pelvic-fin bases are medially separated by a small interspace, about half-length pelvic-fin base. Total pelvic-fin rays 5 (I + 4). Caudal fin subtruncate, upper and lower corners rounded. Total principal caudal-fin rays 13 (I + 11 + I), total dorsal procurrent rays 16–18 (xv–xvii + I), total ventral procurrent rays 13 or 14 (xii–xiii + I). Anus and urogenital papilla at vertical through anterior portion of dorsal-fin base.
Osteology (
Figure 3D,
Figure 5D,
Figure 6D and
Figure 7D): Anterior margin of mesethmoid nearly straight, mesethmoid cornu subtriangular in dorsal view. Lacrimal and sesamoid supraorbital narrow, rod-like, sesamoid supraorbital length about two times lacrimal length. The premaxilla is sub-rectangular in dorsal view, slightly tapering laterally. Maxilla is boomerang-shaped, slender, slightly longer than premaxilla, with a minute posterior process. Autopalatine sub-rectangular in dorsal view when excluding posterolateral process, its shortest width about half autopalatine length, lateral margin about straight, medial margin slightly concave. The postero-lateral process of autopalatine is subtriangular; its length is about two-thirds of autopalatine length. Metapterygoid subtrapezoidal, slightly longer than deep. Quadrate robust, dorsoposterior outgrowth in contact with hyomandibular outgrowth. Hyomandibula long, anterior outgrow shallow, with a steep concavity in the middle portion of dorsal margin. The opercle is moderately elongated; the depth of the opercular odontode patch is about two-thirds of the dorsal articular facet of the hyomandibula; the dorsal process of the opercle is short and blunt. Opercular odontodes 13–15, odontodes pointed, arranged in irregular transverse rows. Interopercle long, about three-fourths of hyomandibula length. Interopercular odontodes 31–42, odontodes pointed, arranged in irregular longitudinal rows. Preopercle compact, without ventral expansion. Basibranchials 2 and 3 elongate without distinctive constrictions. Hypobranchial 1 long, distal extremity slightly broader than proximal extremity. Hypobranchials 2 and 3 are subtriangular and about equal in length. Parurohyal robust, lateral process relatively elongate, pointed, slightly curved; parurohyal head well-developed, with prominent anterolateral paired process; middle foramen oval; posterior process short, about half distance between anterior margin of parurohyal and anterior insertion of posterior process. Branchiostegal rays 7. Vertebrae 36 or 37. Ribs 13. Dorsal-fin origin at vertical through centrum of 19th or 20th vertebra; anal-fin origin at vertical through centrum of 23rd or 24th vertebra. Two dorsal hypural plates, corresponding to hypurals 4 + 5 and 3, respectively; single ventral hypural plate corresponding to hypurals 1 and 2 and parhypural.
Colouration in alcohol (
Figure 11): The ground colour of the dorsum, dorsal portion of flank and head side is pale brownish yellow, becoming lighter on the ventral part of flank and head side. LDPZ 1 and 2 are represented by pale brown round spots longitudinally in close proximity, sometimes coalesced. LDPZ 3 is represented by a dark brown stripe interrupted in its posterior portion. In some specimens below about 45 mm SL and all specimens below about 35 mm SL, LDPZ 3 is represented by separate dark brown blotches. LDPZ 4 is represented by dark brown melanophores more concentrated on the posterior half of the flank, forming small rounded marks with diffuse limits. Venter white. Odontode patches are light grey. Barbels are light grey. Unpaired fins are hyaline, with dark brown chromatophores concentrated on rays and a vertical series of brown spots on the caudal-fin base, sometimes forming diffuse bars. Paired fins are hyaline, with brown dots on the pectoral-fin base.
Colouration in life: Similar to colouration in alcohol, but with pale golden dots above LDPZ 3.
Distribution and habitat. Trichomycterus caparaoensis is known only from the upper Rio Manhuaçu tributaries draining the Serra do Caparaó (
Figure 8). The species was found in clear water, fast-flowing streams with forested margins, and with gravel, sand and rocks on the bottom, in altitudes between about 800 and 1015 m asl. Specimens were collected above the bottom gravel.
Etymology. The name caparaoensis is an allusion to this new species presently being known only from the Serra do Caparaó region.
Trichomycterus castelensis Costa, Katz and Vilardo sp. nov.
LSID:urn:lsid:zoobank.org:act:3CCC62D6-67EC-4C10-8388-442551EDBAE4
Holotype. UFRJ 13481, 63.3 mm SL; Brazil: Minas Gerais State: Afonso Claudio Municipality: small stream tributary to Rio Boa Sorte, Rio Guandu drainage, Rio Doce basin, 20°11′40″ S 41°03′42″ W, altitude about 630 m, Serra do Castelo; A. M. Katz and P. Vilardo, 20 September 2022.
Paratypes. UFRJ 13250, 17 ex., 21.9–57.1 mm SL; UFRJ 13482, 4 ex. (C&S), 33.6–51.1 mm SL; UFRJ 13251, 3 ex., 20.2–40.0 mm SL; UFRJ 13108, 1 ex. (DNA), 40.2 mm SL; CICCAA 07776, 3 ex., 40.6–49.3 mm SL; collected with holotype.
Diagnosis. Trichomycterus castelensis is distinguished from all other congeners of the
T. jacupiranga complex by the following combination of character states: LDPZ-3 being represented by a row of dark brown to black spots, decreasing in size posteriorly, their diameter usually greater or sometimes nearly equal to opercular odontode diameter, anterior-most spots sometimes longitudinally coalesced (vs. a dark brown stripe interrupted in its posterior portion in
T. caparaoensis); the dorsal surface of head between orbits slightly concave (vs. convex in
T. caparaoensis); a relatively long interopercular patch of odontodes, longer than the largest horizontal length of the hyomandibular anterior outgrown (
Figure 3E; vs. shorter in
T. vinnulus,
Figure 3F); 14 ribs (vs. 10 or 11 in
T. gasparinii,
T. jacupiranga, and
T. vinnulus); the postero-lateral process of the autopalatine postero-laterally directed, its length about two thirds or less of the autopalatine length (
Figure 5E; vs. posteriorly directed, its length about equal autopalatine length in
T. vinnulus,
Figure 5F); the second hypobranchial short, about so long as the third hypobranchial (
Figure 7E; vs. long, about twice longer than the third hypobranchial in
T. vinnulus,
Figure 7F); the posterior process of the parurohyal long, about eight tenths of the distance between the anterior margin of the parurohyal and the anterior insertion of the posterior process (
Figure 6E; vs. short, about half the distance in
T. castelensis,
Figure 6F); a single S6 median pore (vs. paired S6 pore in
T. caparaoensis and
T. gasparinii); opercular odontodes 17–19 (vs. 11–15 in
T. caparaoensis); anterior section of the infraorbital canal present (vs. absent in
T. gasparinii); eight pectoral-fin rays (vs. seven in
T. gasparinii); and pointed mandibular teeth (vs. incisor-like in
T. jacupiranga).
Trichomycterus castelensis is also distinguished from all other congeners of PAC by having a relatively shorter autopalatine postero-lateral process (
Figure 5E), its length about two-fifths of autopalatine length (vs. three-fifths or more).
Description. General morphology: Morphometric data are in
Table 5. The dorsal surface of the head between orbits is slightly concave, and the eye is slightly projected dorsally. Maxillary barbel posteriorly reaching pectoral-fin base, rictal barbel reaching a middle portion of an interopercular patch of odontodes, nasal barbel reaching the area just anterior to opercle. Jaw teeth pointed. Premaxillary teeth 39–42, slightly curved, arranged in irregular rows; dentary teeth 40–50, slightly curved backwards, irregularly arranged. Opercular odontodes 17–19, interopercular odontodes 36–40. Odontodes pointed, arranged in irregular rows.Anterior infraorbital canal present. Single median s6. Dorsal and anal fins are subtriangular, margins slightly convex. Total dorsal-fin rays 12 (iii + II + 7), total anal-fin rays 10 (iii + II + 5). Anal-fin origin at vertical through dorsal-fin base posterior end or just posterior to it. Pectoral-fin filament length is about 20–40% of pectoral-fin length without filament. Total pectoral-fin rays 8 (I + 7). Pelvic fin posteriorly reaching vertical just posterior to middle dorsal-fin base. Pelvic-fin bases are medially separated by interspace about three-fourths pelvic-fin base. Total pelvic-fin rays 5 (I + 4). Caudal fin subtruncate, upper and lower corners rounded. Total principal caudal-fin rays 13 (I + 11 + I), total dorsal procurrent rays 17–21 (xvi–xix + I), total ventral procurrent rays 13–15 (xii–xiv + I). Anus and urogenital papilla at vertical through area just anterior to middle of dorsal-fin base.
Osteology (
Figure 3E,
Figure 5E,
Figure 6E and
Figure 7E): Anterior margin of mesethmoid slightly concave, mesethmoid cornu rod-shaped. Lacrimal and sesamoid supraorbital narrow and flat, sesamoid supraorbital length about three times lacrimal length. The premaxilla is sub-rectangular in the dorsal view. Maxilla is boomerang-shaped, slender, slightly longer than premaxilla, with a minute posterior process. Autopalatine is sub-rectangular in the dorsal view when excluding the posterolateral process; its shortest width is about two-fifths autopalatine length, lateral margin slightly convex, and medial margin with pronounced concavity. The postero-lateral process of autopalatine is subtriangular and short, its length about two-fifths of autopalatine length. Metapterygoid subtrapezoidal, slightly deeper than long. Quadrate robust, dorsoposterior outgrowth in contact with hyomandibular outgrowth. Hyomandibula long, anterior outgrow shallow. The opercle is moderately elongated; the depth of the opercular odontode patch is about two-thirds of the dorsal articular facet of the hyomandibula, and the dorsal process of the opercle is short and blunt. Interopercle long, about three-fourths of hyomandibula length. Preopercle compact, without ventral expansion. Basibranchials 2 and 3 elongate, slightly constricted at the middle portion. Hypobranchial 1 long, distal extremity slightly broader than proximal extremity. Hypobranchials 2 and 3 are subtriangular and about equal in length. Parurohyal robust, lateral process relatively elongate, pointed, slightly curved; parurohyal head well-developed, with the prominent anterolateral paired process; middle foramen round, large; posterior process long, about eight-tenths of distance between anterior margin of parurohyal and anterior insertion of posterior process. Branchiostegal rays 6 or 7. Vertebrae 37. Ribs 14. Dorsal-fin origin at vertical through centrum of 20th vertebra; anal-fin origin at vertical through centrum of 24th vertebra. Two dorsal hypural plates, corresponding to hypurals 4 + 5 and 3, respectively; single ventral hypural plate corresponding to hypurals 1 and 2 and parhypural.
Colouration in alcohol (
Figure 12): The ground colour of the flank and dorsum is light brownish yellow, gradually becoming lighter ventrally, and the colour ground of the dorsum is light brown. LDPZ 1–4 comprising dark brown to black round spots. LDPZ 1 spots sometimes coalesced with neighbouring LDPZ 2 and 3 spots to form transverse bars. LDPZ 3 spots decreasing in size posteriorly, anterior-most spots sometimes longitudinally coalesced. LDPZ 4 is represented by dispersed dark brown chromatophores, not forming distinctive marks. Venter white. The dorsal surface of the head is brown, and the lateral surface is light brownish yellow with irregularly diffuse dark brown chromatophores, more concentrated above the opercle. Nasal barbel light brown, maxillary and rictal barbels light grey. Fins hyaline, with small black spots on the basal portion of the pectoral and unpaired fins. Adults and juveniles with similar colour patterns.
Colouration in life: Similar to colouration in alcohol, with minute patches of golden iridescence on the dorsum and dorsal part of the flank.
Distribution and habitat. Trichomycterus castelensis is known only from the type locality area, in a stream tributary to the Rio Boa Sorte, Rio Guandu drainage at the Serra do Castelo, about 630 m asl, Rio Doce basin (
Figure 8). The site where the species was collected is clear water and fast-flowing streams, about 1 m wide and 0.3 wide, with gravel, sand and rocks on the bottom and secondary vegetation on the banks. Specimens were collected below rocks.
Etymology. The name castelensis refers to the occurrence of the new species in the Serra do Castelo region.


 ,
, 












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}