Abstract
This report aims to characterize and reuse fish head powder (FHP) as a potential carbon/nitrogen source for microbial fermentation to produce prodigiosin (PG), and to evaluate its novel anti-fungal effect via an experimental and docking study. FHP has high protein (30.25%) and lipid (5.12%) content and a diversity of elements, which suggests that FHP is a potential substrate for microbial fermentation. Serratia marcescens TNU01 was screened as the most effective strain to produce PG from FHP. Further experiments indicated that the combination of FHP and crude chitin at the ratio of 9/1 and used at 1.5% as a C/N source for fermentation significantly enhanced PG production. Supplementing a minor amount of 0.05% Ca3(PO4)2, 0.03% K2SO4 also positively affects PG yield production. The fermentation using a bioreactor system (type of 14 L) achieved the highest PG yield production (6.4 mg/mL) in a short cultivation time (10 h) in comparison with the highest PG yield (5.8 mg/mL) obtained after 36 h of fermentation in a flask. In the anti-fungal evaluation of various plant pathogen fungal strains, PG demonstrated efficacy against Fusarium solani F04. The molecular docking results also showed that PG effectively interacted with targeting protein 3QPC of F. solani with good binding energy (DS—9.2 kcal/mol) and an acceptable RMSD value (0.94 Å). In this study, we provide a novel application of FHP for the eco-friendly production of PG, which can potentially used for the management of Fusarium solani.
Keywords:
fishery by-products; fish heads; microbial fermentation; prodigiosin; Fusarium solani; fungicide Key Contribution:
This work is the first to report the reuse of fish heads for producing prodigiosin with a high-level yield at a large scale via microbial fermentation using a bioreactor system. The novel anti-Fusarium solani was also characterized via experiments and molecular docking.
1. Introduction
Biopigment from microorganisms is a natural metabolite source that has attracted much interest in research due to possessing a variety of valuable bioactivities with promising applications in many fields [1,2,3,4,5,6,7]. Notably, microbial pigments (MPs) also have many advantages compared to other sources, such as plant or synthetic, regarding non-toxic properties, easily controlled parameters in production toward cost-effectiveness, not affecting the environment, and enhancing productivity [1,8]. Among those, bacteria, yeasts, fungi, and microalgae are abundant sources for producing MPs [3]. Producing MPs with a low cost and less environmental impact using a fermented medium containing byproducts/waste has recently emerged as an effective strategy [8]. Some common sources of byproducts/waste used in MP production include agriculture, fishery, and industrial processing. In this study, fish by-products will be utilized for fermentation in MP production.
In the fish processing industry, large amounts of fish discards (FDs) are created, accounting for 20–80%, including heads (9–12%), muscle trimmings (15–20%), bones (9–15%), viscera (12–18%), scales (5%), skin and fins (1–3%) [9]. Worldwide, an estimated 130 million tons of FDs, including fish heads, are produced annually, approximating 75% of total fish output [10]. Notably, this abundant discard remains rich in nutrients such as protein, minerals, lipids, and vitamins [11]. Thus, it is also recycled as low-value items such as animal food and fertilizer or discarded into the environment [12]. To avoid fish waste and reduce environmental pollution and simultaneously upgrade FDs, fish heads were used in this work as a source of nutrients for the potential biosynthesis of pigments via microbial fermentation. Numerous MPs were identified, such as prodigiosin, phenazine, carotenoids, canthaxanthin, astaxanthin, violacein, bacteriochlorophylls, etc. [1,13]. This study tends to produce prodigiosin (PG)—a red pigment from fish head powder (FHP) via S. marcescens fermentation.
PG production has attracted much attention in research due to possessing multi-value bioactivities, which have promising applications in various fields, including agriculture, medicine, the environment, colorants, and dyes [8,14,15,16,17,18,19,20]. The emerging trend in PG production is green biosynthesis via fermentation using various byproducts/wastes [8]. This strategy also offers advantages in enhancing PG yield and shortening fermentation time compared to other sources [8,21]. According to this trend, this work will utilize FHP as the novel substrate for fermentation. Additionally, scientific works rarely involve an analysis of the nutritional components of the input material in the cultural medium [22]. This scientific information is also vital for the effective fermenting process. Thus, this study will also exploit this new aspect. Moreover, another novelty in this work is exploring PG’s unexplored bioactivity via experimental tests and computational simulation. The steps of this study are summarized in Figure 1.
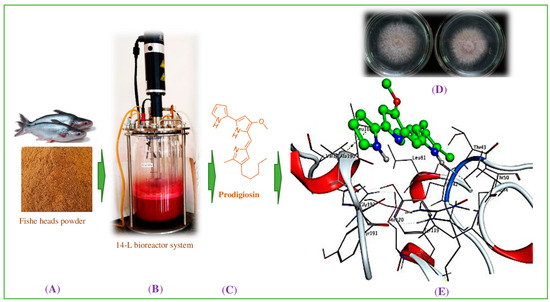
Figure 1.
The scheme of this study. The fishery processing by-product, fish head powder, (A) was fermented via TUN01 strain for the production of prodigiosin (B). Prodigiosin was purified (C) and its novel antifungal effects were determined via in vitro experiments (D). Insights into the interaction mechanism were obtained via molecular docking (E).
2. Materials and Methods
2.1. Materials
The fish (Pangasius bocourti) heads were obtained from Tan Long Seafood Co., Ltd. (Group 27, An Hai Hamlet, Loc An Ward, Dat Do District, Ba Ria-Vung Tau, Vietnam). In general, the fishes were washed and their heads were separated and considered as by-products. The fish heads were dried at 80–85 °C then ground to powder and used for the testing.
The four Serratia marcescens strains (TNU01, TNU02, CC17, and TKU011) were sourced from our earlier work [23]. Some plant-pathogen fungal strains provided by the Institute of Biotechnology and Environment, Tay Nguyen University, Buon Ma Thuot, Vietnam were used for the anti-fungal assay. Silicagel of Geduran® Si 60 (size: 0.040–0.063 mm) was purchased from Merck Sigma Chemical Co. (St. Louis City, MO, USA). All the common chemical agents and solvents were used at the highest grade.
2.2. Methods
2.2.1. Analysis of Nutrient Ingredients of the Input Material, Fish Head Powder, for Fermentation
The content of some constituents of FHP was determined, including total protein [24], total lipids [25], reducing sugar [26], total dissolved sugar [27], and total mineral ash [24]. The mineral elements were detected using the generation 5 phenom pro and proX SEMs. The content of some specific minerals (potassium, magnesium, and calcium) was determined via an atomic absorption spectrophotometer (AA-7000). Phosphorus was determined via spectrophotometric methods [28].
2.2.2. Production of Prodigiosin from Fish Head Powder via Fermentation
Biosynthesis of PG by Different S. marcescens Strains Using FHP as a C/N Source
Several S. marcescens strains, including TNU01, TNU02, CC17, and TUK011, were conducted for fermentation to produce PG. The liquid culture medium containing 1% C/N source (FHP/casein ratio of 7/3), 0.025% Ca3(PO4)2, 0.02% K2SO4, and an initial pH of 7.0 was used for fermentation by these strains of S. marcescens (TNU01, TNU02, CC17, and TUK011). The cultivation was performed using 100-mL flasks with shaking speed of 150 rpm, for 48 h at a cultivation temperature of 27.5 °C. This fermentation condition was labeled as (*). The culture was centrifuged at 6000 rpm for 20 min, and the supernatant was recovered for the determination of PG content. The strain, TNU01, was found to have produced the highest PG yield; thus, it was selected for further tests.
The Effect of Free Protein and Chitin Supplemented in the Culture Broth on PG Production
The FHP was combined with casein at different ratios of 10/0, 9/1, 8/2, 6/4, and 5/5 and used at a concentration of 1.5% as a C/N source for fermentation. The salt composition, including 0.025% Ca3(PO4)2, and 0.02% K2SO4 and the initial pH set at 7.0, was used to prepare the culture medium. The fermentation condition was conducted following the above-described protocol (*).
For enhancing PG production, crude chitin (CC) was also added into the culture broth. The FHP/CC was checked in the ratios of 10/10/0, 9/1, 8/2, 6/4, and 5/5 and used as a C/N source at 1.5%. The salt compositions of culture medium and the fermentation conditions were described as in the above protocol (*). The FHP/CC ratio of 9/1 was selected for further study.
The Effect of Salts Composition on PG Production
Various salt kinds of sulfate (K2SO4, (NH4)2SO4, ZnSO4, and MgSO4) and phosphate (KH2PO4, K2HPO4, Ca3(PO4)2, NaH2PO4, and Na2HPO4) were utilized for the tests in this study. Ca3(PO4)2 and K2SO4 were found to be effective on PG yield. Thus, these salts were further investigated to find their suitable added concentration in the culture broth. Ca3(PO4)2 and K2SO4 were supplemented into the medium with various concentration ranges 0, 0.01, 0.025, 0.05, 0.075, 0.1, 0.125, and 0.15 and 0, 0.01, 0.02, 0.03, 0.04, 0.05, and 0.06, respectively.
Scale-Up of PG Biosynthesis Using a 14-L Bioreactor System
Six hundred ml of bacterial seed was pre-cultivated in three 500 mL flasks at 30 °C for 36 h and then all were injected into a bioreactor tube (type of 14 L) containing 5.4 L of culture broth containing 1.5% C/N (FHP/CC = 9/1), 0.05% Ca3(PO4)2, 0.03% K2SO4, with an initial pH of 7.0. The cultivation was performed at 27.5 °C, shaking at 150 rpm, and the dissolved oxygen content was set at 1.0 vvm. The fermentation was carried out in 12 h and the PG yield was determined every 2 h.
Quantification, Purification, and Confirmation of Red Pigment Compound as Prodigiosin
PG was quantified according to the protocol presented in the earlier work [23]. A solution mixture containing 4 mL of methanol, 0.5 mL of cultural supernatant, and 0.5 mL of KAl(SO4)2.12H2O 2% (w/v) was centrifuged at rotation of 1400 rpm for 5 min. Harvesting 1 mL of supernatant and mixing with 0.5 N HCl/methanol with a ratio of 1/4 (v/v). The optical density of the final mixture was measured at a wavelength of 535 nm, and PG content was determined via a standard equation of PG. The red pigment compound was purified from the fermented culture broth using a liquid–liquid extraction method and purified via column chromatography through an earlier published method described in detail in [22]. This purified pigment was confirmed as PG via UV/vis absorption analysis, and its purity was determined via high-performance liquid chromatography (HPLC) and compared to the standard compound. The sample was separated in a C18 column with a mobile phase of MeOH/H2O (70/30 v/v) that was adjusted to a pH of 3.0 using 10 mM ammonium acetate. The condition for HPLC analysis was set up as a flow rate of 0.8 mL/min, 3 μL of injected samples (samples solvated in MeOH at 1 mg/mL), and a detection wavelength of 535 nm at column temperature 30 °C for 20 min.
2.2.3. Anti-Fungal Assays
Antifungal activity was determined via the inhibition against the mycelial growth of fungal strains and the fungal spore-germination inhibition. DMSO was used for dissolving PG, and this PG solution was used for testing antifungal activity at various concentrations.
- -
- Mycelial growth inhibition assay: This assay was carried out according to the methods presented in previous reports [29,30] with modification. A plug of fungal mycelia of the growing strain was obtained, then placed in the center of a Petri dish containing PDA (potato D-glucose agar) medium. Then, 25 µL of sample solution was slowly sprayed onto this mycelial plug. Next, 25 µL of DMSO was used as the control experiment. All these Petri dishes were incubated at 28 °C. After 5 days of incubation, the diameter growth of fungal mycelium in the experiment dish (De, mm) and the control dish (Dc, mm) was measured for the calculation of the antifungal effect based on the equation below:Mycelial growth inhibition (%) = [Dc − De]/Dc × 100%
- -
- Inhibitory spore-germination assay: The pathogen fungal strains were cultivated on PDA medium at 28 °C for 7 days then suspended in sterile distilled water (5 mL/disk), and the suspension was then filtered through the sterile paper tissues to harvest spores. The spore density was counted using a counting chamber (Marienfeld, Lauda-Königshofen, Germany) and a CX31 upright microscope (Olympus, Tokyo, Japan). The inhibitory spore-germination activity was performed according to the protocol previously presented in the reports [31,32] with modifications. Fifty microliters of spore solution (containing around 105 fungal spores) were mixed with 25 µL of the sample solution in a 200-µL Eppendorf tube, then incubated at 28 °C. A round of 200 spores was observed using a CX31 upright microscope (Olympus, Japan), and the number of germinated spores (GS) was counted every 8 h. A spore with the length of its germinal tube reaching one-half of the spore diameter indicates germination [33]. The solution containing 50 µL of fungal spores and 25 µL of sterile distilled water only was used as the control group. The inhibitory spore-germination effect was determined according to the equation below:where CGS is the number of germinated spores in the control group, while EGS is the number of germinated spores in the experimental group. To clear the results, after 16 h of incubation, the solution in the Eppendorf tube was injected into the center of the PDA plate and inoculated for a further 2 and 4 d at 28 °C. Then, based on the diameter of fungal growth in the control disk (C, mm) and the experiment disk (E, mm), the activity was determined following the below equation:The inhibitory spore-germination activity (%) = [CGS − EGS]/CGS × 100%% Inhibitory activity = [C − E]/C × 100%
2.2.4. Docking Study Protocol
The molecular docking was performed following the method previously presented in [31,32,33]. The data structure of targeting protein 3QPC was obtained from Worldwide Protein Data Bank (accessed on 20 June 2023 at https://www.rcsb.org/structure/3qpc) to prepare its 3D structure using MOE-2015.10 software. The 3D structure of PG and Demethoxycurcumin (DC) were also prepared and optimized using MOE-2015.10. The ligands (PG and DC) were docked into the active binding sites on protein 3QPC using the same software. The output data such as RMSD, DS, types of interaction, amino acid composition, and the distances of linkages were harvested and analyzed.
3. Results and Discussion
3.1. Nutrient Composition of Input Material (Fish Head Powder) for Fermentation
Some major nutrient ingredients such as the protein, lipids, sugars, and salts of FHP were determined. As per the data summarized in Table 1, this by-product has a high total protein content (30.25%), a moderate range of total lipids (5.12%), and a minor amount of sugars, with 0.105% and 0.059% of total dissolved sugar and reducing sugar, respectively. Scanning Electron Microscopy (Thermal Fisher Scientific, FEI Electron Optics B.V., Achtseweg Noord 5, 5651GG Elindhoven, the Netherlands was applied for the detection of the mineral elements in the ash of the FHP (Figure 2). The FHP ash was a rich source of diverse mineral elements such as Ca, P, Na, Mg, Si, and Cl. Some major mineral elements were further identified, such as K, Mg, and Ca, via AA-7000 atomic absorption spectrophotometry and P via spectrophotometric methods. Of these, K, Mg, and Ca were found to be present in FHP ash with a significant content of 0.156, 2.239, and 2.158, respectively, while P was detected in a minor concentration (0.918 ppm) in the FHP ash. These experimental data suggest that FHP is a substrate rich in nutrients and salts, suitable for microbial fermentation.

Table 1.
Major nutrients in fish head powder.
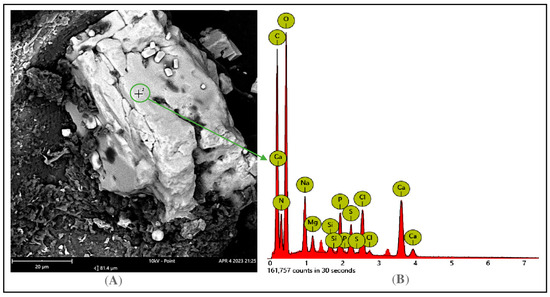
Figure 2.
Detection of mineral elements of FHP ash via SEM (Scanning Electron Microscopy) capture. The analyzed point on the surface of FHP ash (A) and its mineral elements was detected via EDX (Energy dispersive X-ray) spectra (B).
3.2. Establishment of Fermentation Process for PG Production on Small Scale and Scaling-Up in Bioreactor System
Screening PG-producing bacterial strain: Previous studies indicate that different substrates used for fermentation may require different bacterial strains for effective conversion [8,22,23,34]; as such, the tests for screening the suitable PG-producing strain were performed. Several S. marcescens strains (TNU01, TNU02, CC17, and TUK011) were utilized as a PG-producing strain for fermentation of FHP. As shown in Figure 3A, using TNU01 for fermentation resulted in the highest PG production (2.12 mg/mL), while other strains produced PG at a lower yield (1–1.32 mg/mL). Thus, the strain, TNU01, was suggested for our further tests. The current trend in PG production is using eco-friendly material for fermentation, and marine-derived by-products were also used for this purpose [8]. However, this is the first study of the potential application of fish head by-products for effective PG production through fermentation via S. marcescens.
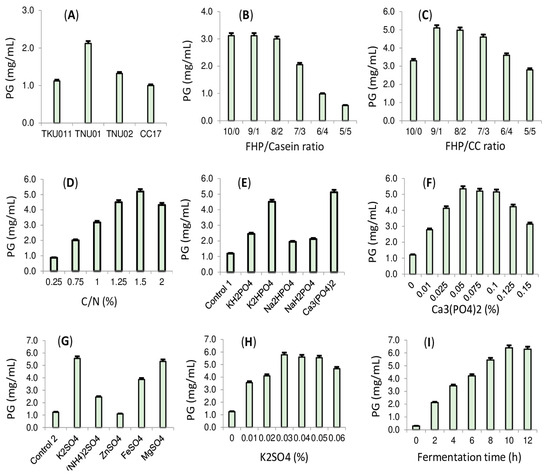
Figure 3.
The effect of different S. marcescens strains (A), FHP/casein ratio (B), FHP/CC ratio (C), the concentration of C/N source (D), different phosphate salts (E), Ca3(PO4)2 concentration (F), different sulfate salts (G), K2SO4 (H) on prodigiosin production. Scaling-up of PG production in a 14 L bioreactor system (I).
Evaluating the effect of casein and chitin supplemented into the culture medium on PG capacity biosynthesis: In order to obtain a higher PG yield via S. marcescens fermentation, the supplementation of protein and carbohydrates is required [23,34]. Casein was found to be the most suitable free protein source added to culture medium for fermentation in our previous works [23,34]; thus, this protein was combined with FHP at various ratios and then used for fermentation. The results (Figure 3B) indicate that the FHP/casein at the ratios of 10/0, 9/1, and 8/2 are suitable for fermentation to produce PG, with a high yield of 3–3.1 mg/mL. For the eco-friendly bioproduction of this pigment, FFP was chosen as the sole C/N source for fermentation; as such, no further casein was required to add into the medium. Among various carbohydrates, a-chitin was found to enhance PG production [23]; thus, the crude a-chitin (CC) was added into the medium to investigate its effect. As presented in Figure 3C, the combination of FHP and CC at the ratio of 9/1 and 8/2 significantly enhanced PG yield (4.6–5.1 mg/mL) compared to other FHP/CC ratios (lower than 3.3 mg/mL). Of these, the FHP/CC ratio of 9/1 resulted in the highest PG yield (5.1 mg/mL). Thus, FHP was combined with CC at this ratio for fermentation as a C/N source at various concentrations, and the most suitable concentration of C/N was found to be 1.5%.
Evaluating the composition of the salts supplemented into the culture broth on PG production: Phosphate and sulfate salts were evidenced showing a significant enhancement of PG production via S. marcescens fermentation. In the present study, various salt kinds were tested. Ca3(PO4)2 was screened as a good source of phosphate salts (Figure 3E), and the most suitable concentration added into the culture medium was found to be 0.05% in various tested concentrations of 0–0.15% (Figure 3F). Among the various sulfates, K2SO4 was found to be the most effective at enhancing PG yield (Figure 3G), and its suitable supplemented concentration was found to be 0.03% in different tested concentrations of 0–0.06% (Figure 3H). Overall, the novel cultured medium for cost-effective PG biosynthesis was investigated and contained 1.5% C/N (FHP/CC = 9/1), 0.05% Ca3(PO4)2, and 0.03% K2SO4.
Scale-up of PG biosynthesis using a 14-L bioreactor system: For the massive production of PG, the above-designed culture medium was fermented by S. marcescens TNU01 using a bioreactor system. As summarized in Figure 3I, PG production in a bioreactor system achieved the highest yield (6.4 mg/mL) in a shorter cultivation time (10 h) compared to fermenting in a 100 mL flask (PG yield of 5.8 mg/mL and fermentation time of 36 h). In several previous reports summarized in our overview work [8], PG was produced in the range of 3450–6886 mg/L with a cultivation time of from 8–10 h, while other studies have reported Pg production of from 50–595 mg/L in the longer fermentation time of over 20 h. Compared to these, the current study demonstrated the potential PG yield of 6400 mg/L within a shorter fermentation time on a large scale, using a newly designed media formulation mainly based on the fishery processing by-product, FHP.
3.3. Extraction, Isolation, and Confirmation of Red Pigment Compound as Pure Prodigiosin
The crude pigment collected from the fermented broth in the bioreactor was used for extraction, purification, and identification, using the protocol previously presented in [23]. PG was previously biosynthesized, isolated, and identified from cassava-based industrial wastes via Serratia marcescens TNU01. To reconfirm the red pigment compound as prodigiosin biosynthesized in this work via the same bacterial strain, TNU01, some rapid methods including UV/vis absorption (Figure 4) and HPLC (Figure 5) were employed. This purified pigment showed the highest absorption spectroscopy at the wavelength of 535 nm (Figure 4), which is the specific absorption of prodigiosin [23,34].
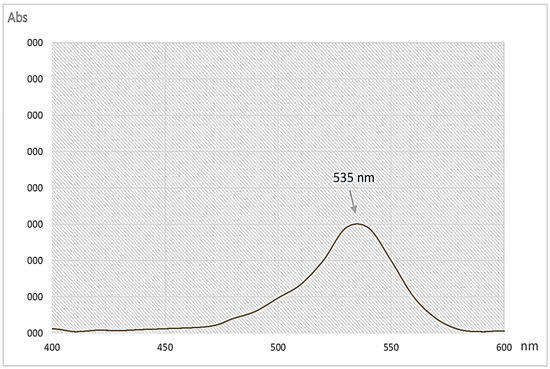
Figure 4.
UV/vis spectrum of purified prodigiosin produced via Serratia marcescens TNU01 in this work.
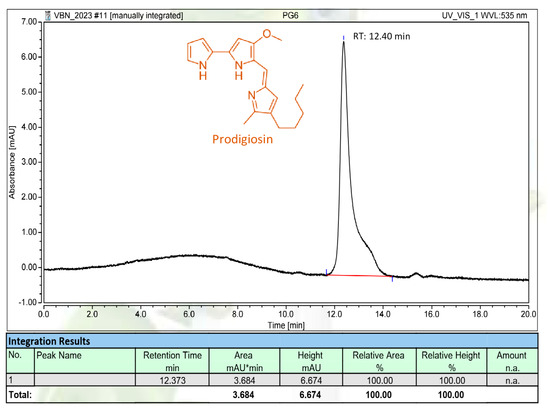
Figure 5.
The HPLC fingerprint of purified PG produced via Serratia marcescens TNU01 in this work. RT: retention time.
For further confirmation, the PG produced and isolated in this work was subjected to HPLC analysis. As illustrated in Figure 5, the PG compound appeared as a main and single peak in its HPLC fingerprinting at 12.40 min of retention time, which is approximately similar to those of the PG analyzed in our earlier reports with the retention time of 12.368–12.425 min [23,34]. The HPLC fingerprinting also indicated that the PG synthesized in this work was purified to a high grade; as such, it was qualified for use in the following experiments.
3.4. Evaluation of Potential Anti-Fungal Effect of PG against Plant-Pathogenic Fungi
For the evaluating the anti-fungal effect of PG, some plant-pathogenic fungal strains, including F. solani F04, G. butleri F07, P.mangiferae F08, F. oxysporum F10, and F. incarnatum F15, collected in the central highlands of Vietnam [29], were conducted for testing the mycelial growth inhibition. The results are presented in Table 2 and Figure 6. Of the tested fungal strains, the PG showed a positive inhibition against F. solani F04 and F. oxysporum F10, with a mycelial growth inhibition value of 37.08% and 19.69%, respectively, while the PG showed a weak effect (3.05–8.55%) against G. butleri F07, P.mangiferae F08, and F. incarnatum F15.
Table 2.
Anti-Fungal Effect of PG Against Plant-Pathogenic Fungi.

Figure 6.
Anti-fungal effect of PG against plant-pathogenic fungi, including F. solani F04, G. butleri F07, P. mangiferae F08, F. oxysporum F10, and F. incarnatum F15. C and E were symbolized for control and experiment.
PG has been widely investigated for its potential application in many fields. According to 2022 recent report of Nguyen et al. [8], PG showed potential for managing numerous organisms harmful to various crops. This metabolite was found to have a potential inhibitory effect on several plant-damaging nematodes [22,35,36] and insects [37,38]. PG was also found to be an effective anti-fungal agent on various plant-pathogenic strains [8]. PG was reported to show potent inhibition against various strains of Fusarium genus such as F. oxysporum, F. roseum, F. ventricosum, F. moniliforme [8,39,40,41]. However, few studies concerning the anti-F. solani activity have been conducted so far [8]. This is the first study to report PG’s potential inhibitory effect on the mycelial growth of the pathogen fungi, F. solani F04, originating from coffee roots.
Though PG inhibited F. solani F04 to a moderate extent (37.08%), it was clearly observed that the amount of F04 mycelia in the treated Petri dish was much lower than that in the control Petri dish (Figure 6). The inhibitory spore germination activity was also accessed, and PG demonstrated a high inhibitory effect on the spore germination of F. solani F04, with an inhibitory activity value of more than 50% after 8 and 16 h of prodigiosin treatment (Figure 7). To further elucidate the potent anti-F. solani effect of PG, the solution of F. solani F04 spores treated with PG was injected into the center of the PDA plate and inoculated for a further 2 and 4 d at 28 °C; then the fungal growth was examined. As presented in Figure 7, the mycelium grown from F. solani F04 spores treated with PG seems to have a much lower resistance. PG demonstrated potent inhibition (68.5–80%) at 2 days of incubation, and PG can inhibit the mycelia growth at 2 days of incubation (Figure 8). These newly recorded data suggest that PG is an effective anti-fungal agent for managing F. solani. However, further works accessing the effect of PG in a greenhouse and the field should be conducted to confirm its suitability for use as an anti-F. solani product.

Figure 7.
The inhibitory activity on spore germination after 8 h and 16 h of prodigiosin treatment and the inhibitory ability on mycelial growth developed from spores treated with prodigiosin after 2 days and 4 days of incubation.

Figure 8.
The inhibition ability of mycelial growth developed from F. solani F04 spores treated with prodigiosin at various concentrations. The experiment was recorded after 4 days of incubation. The diameter of fungal growth in the control disk (C, mm) and the experiment disk were measured for the calculation.
3.5. The Molecular Action Mechanism of Anti-Fungal Compound PG via Docking Study
The potential anti-fungal effect of PG on F. solani is newly reported; as such, the possible mechanism of action of this active compound against F. solani is still unknown. For assessing the mechanism of PG in anti-fungal activity, we conducted the assay of molecular mechanism action via a virtual screening study using the protein, 3QPC, targeting anti-F. solani in this work [42]. The 3D structure of 3QPC was produced by MOE (Figure 9A), and the binding sites (BSs) were found via the function of the site finder in MOE. Based on the output data of MOE, four possible BSs were determined (Figure 9B). The sizes and residues of these BSs are summarized in Table A1 in the Appendix A. In a docking study, a ligand may interact with various possible binding sites on a protein, and only the BS with the most potential was chosen for detailed description. The two ligands, including prodigiosin (PG) and demethoxycurcumin (DC), a reported 3QPC inhibitor [42], were selected for docking performance analysis. Of these, PG was found to interact the most effectively with 3QPC at BS4, while DC was found to bind to 3QPC at BS3 with a high affiliation. The computational data are recorded and summarized in Table 3.

Figure 9.
The 3D structures of protein 3QPC (A) and the binding sites (BS) on 3QPC (B).
Table 3.
The summarized data of docking study of ligands prodigiosin (PG) and demethoxycurcumin (DC) binding with the protein, 3QPC, targeting anti-F. solani.
In docking analysis, the two parameters, RMSD and DS, have been widely used to determine the successful interaction (RMSD ≤ 2.0Å) and the effective binding (DS ≤ −3.20 kcal/mol) of a ligand toward its targeting protein [43,44]. As shown in Table 3, the ligands, PG and DC, interacted with protein 3QPC, with low RMSD values of 0.94 and 1.60, indicating that these interactions are significant and eligible for further investigation. Based on the DS energy data, both ligands showed their potent binding ability toward the binding sites of protein 3QPC (DS values in the range of −8.9 to −9.2). The experimental data (in vitro anti-fungal assay) and these docking study results suggest that PG may be a potential candidate for managing the pathogenic fungi, F. solani. The above docking results indicate that the anti-F. solani activity of PG may possiblybe due to the high molecular interaction of PG toward the protein, 3QPC.
To understand the interactions, details of simultaneous interactions and the linkages were also examined (Figure A1 in the Appendix A). Ligand PG bound to the protein, 3QPC, at the BS4 via the interaction with Thr50 and Ala190 to generate two linkages. Of these, PG interacted with Thr50 to form an H-donor linkage with a distance of 3.07 Å and an energy binding value of −2.5 kcal/mol, while this ligand interacted with Ala190 to create a pi-H linkage with a distance of 3.9 Å and an energy binding value of −0.9 kcal/mol. Ligand DC was found to bind to 3QPC at the BS3 by interacting with three prominent amino acids, Asp139, Asp132, and Arg138, thus generating three 2 H-donor linkages and 1 pi-H linkage with the distance/energy binding values of 3.08 Å/-−2.0 kcal/mol, 3.51 Å/−0.6 kcal/mol, and 4.21 Å/−0.9 kcal/mol, respectively.
4. Conclusions
Fish head powder (FHP) has been investigated as a suitable substrate for the eco-friendly bioproduction of PG. The novel cultured medium for the biosynthesis of PG with a high-level yield via S. marcescens TNU01 fermentation was found to contain 1.5% C/N source (FHP/CC = 9/1), 0.05% Ca3PO4, and 0.03% K2SO4. The PG yield was significantly enhanced up to 6.4 mg/mL in a short cultivation time of 10 h when a 14 L bioreactor system was used for conducting fermentation. The pure PG compound was found to be a novel and potent anti-fungal compound against F. solani F04 via in vitro assays. The molecular docking data indicate that PG possibly inhibited F. solani via effective binding to the protein, 3QPC, targeting anti-F. solani. The results of this work suggest the suitability of FHP for PG production at a high level and its potential use for the management of the pathogenic fungi, F. solani.
Overall, this study provides some highlights and novel findings as below:
- ▪
- Fish head powder (FHP) was found to be rich in protein (30.25%), lipids (5.12%), and a diversity of elements in its ash.
- ▪
- This work was the first to report the reuse of FHP for producing prodigiosin with a high yield at a large scale via microbial fermentation.
- ▪
- The utilization of a 14 L bioreactor system for fermentation achieved a higher PG yield production (6.4 mg/mL) in a shorter cultivation time (10 h) compared to those fermented in a flask.
- ▪
- PG demonstrated novel efficiency against the pathogenic fungi, F. solani F04, originating from coffee roots.
- ▪
- The molecular docking data indicate that PG possibly inhibited F. solani via effective binding to the protein, 3QPC, targeting anti-F. solani.
Author Contributions
Conceptualization, V.B.N., and S.-L.W.; methodology, T.H.N. (Thi Hanh Nguyen), and V.B.N.; software, V.B.N. and T.Q.P.; validation, S.-L.W., V.B.N., and A.D.N.; formal analysis, V.B.N., S.-L.W., A.D.N., M.D.D., and V.A.N.; investigation, T.H.N. (Thi Hanh Nguyen), T.H.N. (Thi Huyen Nguyen), T.H.T.T.; resources, V.B.N.; data curation, S.-L.W., V.B.N., and A.D.N.; writing—original draft preparation, V.B.N. and T.H.N. (Thi Hanh Nguyen); writing—review and editing, S.-L.W., V.B.N. and A.D.N.; supervision, V.B.N. and S.-L.W.; project administration, V.B.N. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by a grant from the Ministry of Education and Training, Vietnam (B2022-TTN-06), and was supported in part by National Science and Technology Council, Taiwan (NSTC 111-2320-B-032-001).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All the data for this article can be found in the article.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A
Table A1.
The sizes and residues of the four binding sites on protein 3QPC.
Table A1.
The sizes and residues of the four binding sites on protein 3QPC.
| SITE | SIZE | Residues |
|---|---|---|
| 1 | 22 | 1:(ASP111 ALA112 THR113 LEU114 ASP139 LYS140 ILE141 ALA142 ARG166 VAL210 ARG211) |
| 2 | 20 | 1:(ASP132 LEU133 ASP134 SER135 ARG138 ASP139 ASN161 PRO163) |
| 3 | 16 | 1:(THR19 ASP21 ILE24 ASN47 GLN71 GLY72 VAL73 GLY74 GLY75 GLU97 LEU101) |
| 4 | 16 | 1:(GLY41 SER42 THR43 GLU44 THR50 LEU81 TYR119 MIR120 VAL184 HIS188 LEU189) |
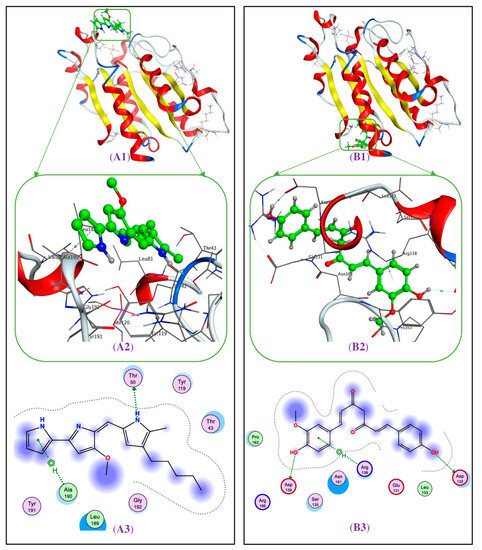
Figure A1.
The interaction of prodigiosin (A1) and demethoxycurcumin (B1) with protein 3QPC, targeting anti-F. solani at the BS4 and BS3, respectively. The 3D (A2) and 2D (A3) structures of ligand, prodigiosin, at the BS4 of protein 3QPC. The 3D (B2) and 2D (B3) structures of ligand demethoxycurcumin at the BS3 of protein 3QPC.
References
- Ramesh, C.H.; Vinithkumar, N.V.; Kirubagaran, R.; Venil, C.K.; Dufossé, L. Multifaceted applications of microbial pigments: Current knowledge, challenges and future directions for public health implications. Microorganisms 2019, 7, 186. [Google Scholar] [CrossRef] [PubMed]
- Sajjad, W.; Din, G.; Rafiq, M.; Iqbal, A.; Khan, S.; Zada, S.; Ali, B.; Kang, S. Pigment production by cold-adapted bacteria and fungi: Colorful tale of cryosphere with wide range applications. Extremophiles 2020, 24, 447–473. [Google Scholar] [CrossRef] [PubMed]
- Celedón, R.S.; Díaz, L.B. Natural pigments of bacterial origin and their possible biomedical applications. Microorganisms 2021, 9, 739. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi, M.; Ahangari, H.; Mousazadeh, S.; Hosseini, S. Microbial pigments as an alternative to synthetic dyes and food additives: A brief review of recent studies. Bioprocess Biosyst. Eng. 2021, 45, 1–12. [Google Scholar] [CrossRef]
- Ding, J.; Wu, B.; Chen, L. Application of marine microbial natural products in cosmetics. Front. Microbiol. 2022, 13, 892505. [Google Scholar] [CrossRef]
- Chatragadda, R.; Dufossé, L. Ecological and biotechnological aspects of pigmented microbes: A way forward in development of food and pharmaceutical grade pigments. Microorganisms 2021, 9, 637. [Google Scholar] [CrossRef]
- Ramesh, C.; Prasastha, V.R.; Venkatachalam, M.; Dufossé, L. Natural substrates and culture conditions to produce pigments from potential microbes in submerged fermentation. Fermentation 2022, 8, 460. [Google Scholar] [CrossRef]
- Nguyen, T.H.; Wang, S.L.; Nguyen, V.B. Recent advances in eco-friendly and scaling-up bioproduction of prodigiosin and its potential applications in agriculture. Agronomy 2022, 12, 3099. [Google Scholar] [CrossRef]
- Coppola, D.; Lauritano, C.; Esposito, F.P.; Riccio, G.; Rizzo, C. Fish waste: From problem to valuable resource. Mar. Drugs 2021, 19, 116. [Google Scholar] [CrossRef]
- Binsi, P.K. Overview of waste generation in fish and shellfish processing industry. In Protocols for the Production of High Value Secondary Products from Industrial Fish and Shellfish Processing; Bindu, J., Sreejith, S., Sarika, K., Eds.; Central Institute of Fisheries Technology: Cochin, India, 2018; pp. 18–27. [Google Scholar]
- Tia, Y.; Junianto. Utilization of Fish Head as Feed Flour. Glob. Sci. J. 2021, 9, 674–680. [Google Scholar]
- Islam, J.; Yap, E.E.S.; Krongpong, L.; Toppe, J.; Peñarubia, O.R. Fish Waste Management—An Assessment of the Potential Production and Utilization of Fish Silage in Bangladesh, Philippines and Thailand; FAO Fisheries and Aquaculture Circular No. 1216; FAO: Rome, Italy, 2021. [Google Scholar]
- Kaur, P.; Singh, S.; Ghoshal, G.; Ramamurthy, P.C.; Parihar, P.; Singh, J.; Singh, A. Valorization of agri-food industry waste for the production of microbial pigments: An eco-friendly approach. In Advances in Agricultural and Industrial Microbiology; Nayak, S.K., Baliyarsingh, B., Mannazzu, I., Singh, A., Mishra, B.B., Eds.; Springer: Singapore, 2022; pp. 137–166. [Google Scholar]
- Rafael, G.A.; Natalia, R.Z.; Carlos, C.Z.; Mario, E.B.; Enrique, H.V.; Lizeth, P.A.; Jesús, A.R.H.; María, A.M.P.; Juan, E.S.H.; Manuel, M.R.; et al. Recent advances in prodigiosin as a bioactive compound in nanocomposite applications. Molecules 2022, 27, 4982. [Google Scholar]
- Wang, S.L.; Nguyen, V.B.; Doan, C.T.; Tran, T.N.; Nguyen, M.T.; Nguyen, A.D. Production and potential applications of bioconversion of chitin and protein-containing fishery byproducts into prodigiosin: A review. Molecules 2020, 25, 2744. [Google Scholar] [CrossRef] [PubMed]
- Islan, G.A.; Rodenak, K.B.; Noacco, N.; Duran, N.; Castro, G.R. Prodigiosin: A promising biomolecule with many potential biomedical applications. Bioengineered 2022, 13, 14227–14258. [Google Scholar] [CrossRef]
- Mnif, S.; Jardak, M.; Bouizgarne, B.; Aifa, S. Prodigiosin from Serratia: Synthesis and potential applications. Asian Pac. J. Trop. Biomed. 2022, 12, 233–242. [Google Scholar] [CrossRef]
- Yip, C.H.; Yarkoni, O.; Ajioka, J.; Wan, K.L.; Nathan, S. Recent advancements in high-level synthesis of the promising clinical drug, prodigiosin. Appl. Microbiol. Biotechnol. 2019, 103, 1667–1680. [Google Scholar] [CrossRef]
- Tania, P.; Tarun, K.B.; Abhijit, M.; Onkar, N.T.; Muthuraj, M.; Biswanath, B. A comprehensive review on recent trends in production, purification, and applications of prodigiosin. Biomass Convers. Biorefin. 2022, 12, 1409–1431. [Google Scholar]
- Darshan, N.; Manonmani, H.K. Prodigiosin and its potential applications. J. Food Sci. Technol. 2015, 52, 5393–5407. [Google Scholar] [CrossRef] [PubMed]
- Han, R.; Xiang, R.; Li, J.; Wang, F.; Wang, C. High-level production of microbial prodigiosin: A review. J. Basic Microbiol. 2021, 61, 506–523. [Google Scholar] [CrossRef] [PubMed]
- Amera, E.; Ahmed, A.E.; Amr, M.M.; Yehia, O. Optimization of bioreactor cultivation parameters by Taguchi Orthogonal array design for enhanced prodigiosin production. Iran. J. Chem. Chem. Eng. 2020, 39, 319–330. [Google Scholar]
- Nguyen, V.B.; Chen, S.P.; Nguyen, T.H.; Nguyen, M.T.; Tran, T.T.T.; Doan, C.T.; Tran, T.N.; Nguyen, A.D.; Kuo, Y.H.; Wang, S.L. Novel efficient bioprocessing of marine chitins into active anticancer prodigiosin. Mar. Drugs 2020, 18, 15. [Google Scholar] [CrossRef]
- Oko, J.O.; Abriba, C.; Audu, J.A.; Kutman, N.A.; Okeh, Q. Bacteriological and nutritional analysis of groundnut cake sold in an open market in Samaru, Zaria-Kaduna State. Int. J. Sci. Technol. Res. 2015, 4, 225–228. [Google Scholar]
- Señoráns, F.J.; Luna, P. Sample preparation techniques for the determination of fats in food. Compr. Sampl. Sample Preparat. 2012, 4, 203–211. [Google Scholar]
- Miller, G.L. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Shibli, S.; Siddique, F.; Raza, S.; Ahsan, Z.; Raza, I. Chemical composition and sensory analysis of peanut butter from indigenous peanut cultivars of Pakistan. Pak. J. Agric. Sci. 2019, 32, 159–169. [Google Scholar] [CrossRef]
- Nguyen, D.N.; Wang, S.L.; Nguyen, A.D.; Doan, M.D.; Tran, D.M.; Nguyen, T.H.; Ngo, V.A.; Doan, C.T.; Tran, T.N.; Do, V.C.; et al. Potential application of rhizobacteria isolated from the central highland of Vietnam as an effective biocontrol agent of robusta coffee nematodes and as a bio-fertilizer. Agronomy 2021, 11, 1887. [Google Scholar] [CrossRef]
- Lee, K.W.; Omar, D.; Cheng, G.L.E.; Nasehi, A.; Wong, M.Y. Characterization of phenazine and phenazine-1-carboxylic acid isolated from Pseudomonas aeruginosa UPMP3 and their antifungal activities against Ganoderma boninense. Pertanika J. Trop. Agric. Sci. 2018, 41, 1795–1809. [Google Scholar]
- Mamadalieva, N.Z.; Youssef, F.S.; Hussain, H.; Zengin, G.; Mollica, A.; Al Musayeib, N.M.; Ashour, M.L.; Westermann, B.; Wessjohann, L.A. Validation of the antioxidant and enzyme inhibitory potential of selected triterpenes using in vitro and in silico studies, and the evaluation of their ADMET properties. Molecules 2021, 26, 6331. [Google Scholar] [CrossRef]
- Mollica, A.; Zengin, G.; Durdagi, S.; Ekhteiari Salmas, R.; Macedonio, G.; Stefanucci, A.; Dimmito, M.P.; Novellino, E. Combinatorial peptide library screening for discovery of diverse α-glucosidase inhibitors using molecular dynamics simulations and binary QSAR models. J. Biomol. Str. Dynam. 2019, 37, 726–740. [Google Scholar] [CrossRef]
- Li, H.; He, Z.; Shen, Q.; Fan, W.; Tan, G.; Zou, Y.; Mei, Q.; Qian, Z. Rapid screening alpha-glucosidase inhibitors from polygoni vivipari rhizoma by multi-step matrix solid-phase dispersion, ultrafiltration and HPLC. Molecules 2021, 26, 6111. [Google Scholar] [CrossRef]
- Nguyen, V.B.; Wang, S.L.; Nguyen, A.D.; Phan, T.Q.; Techato, K.; Pradit, S. Bioproduction of prodigiosin from fishery processing waste shrimp heads and evaluation of its potential bioactivities. Fishes 2021, 6, 30. [Google Scholar] [CrossRef]
- Samer, S.H.; Hannah, U.C.B.; Andreas, S.K.; David, P.K.; Tim, M.W.; Thomas, C.; Jörg, P.; Florian, M.W.G.; Sylvia, A.S.S. Novel prodiginine derivatives demonstrate bioactivities on plants, nematodes, and fungi. Front. Plant Sci. 2020, 11, 579807. [Google Scholar]
- Rahul, S.; Patil, C.; Hemant, B.; Chandrakant, N.; Laxmikant, S.; Satish, P. Nematicidal activity of microbial pigment from Serratia marcescens. Nat. Prod. Res. 2014, 28, 1399–1404. [Google Scholar] [CrossRef] [PubMed]
- Patil, N.G.; Kadam, M.S.; Patil, V.R.; Chincholkar, S.B. Insecticidal properties of water diffusible prodigiosin produced by Serratia nematodiphila 213C. Curr. Trends Biotechnol. Pharm. 2013, 7, 773–781. [Google Scholar]
- Wei, H.; Zheng, R.; Liao, Y.; Fan, K.; Yang, Z.; Chen, T.; Zhang, N. Evaluating the biological potential of prodigiosin from Serratia marcescens KH-001 against Asian citrus psyllid. J. Econ. Entomol. 2021, 114, 1219–1225. [Google Scholar]
- Hiroshi, O.; Sato, Z.; Sato, M.; Koiso, Y.; Iwasaki, S.; Isaka, M. Identification of antibiotic red pigments of Serratia marcescens F-1-1, a biocontrol agent of damping-off of cucumber, and antimicrobial activity against other plant pathogens. Jap. J. Phytopathol. 1998, 64, 294–298. [Google Scholar]
- Sumathi, C.; Mohana, P.D.; Swarnalatha, S.; Dinesh, M.G.; Sekaran, G. Production of prodigiosin using tannery fleshing and evaluating its pharmacological effects. Sci. World J. 2014, 2014, 290327. [Google Scholar] [CrossRef]
- Parani, K.; Saha, B.K. Optimization of prodigiosin production from a strain of Serratia marcescens SRI and screening for antifungal activity. J. Biol. Control. 2008, 22, 73–79. [Google Scholar]
- Akter, J.; Islam, M.Z.; Takara, K.; Hossain, M.A.; Sano, A. Isolation and structural elucidation of antifungal compounds from Ryudai gold (Curcuma longa) against Fusarium solani sensu lato isolated from American manatee. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2019, 219, 87–94. [Google Scholar] [CrossRef]
- Ding, Y.; Fang, Y.; Moreno, J.; Ramanujam, J.; Jarrell, M.; Brylinski, M. Assessing the similarity of ligand binding conformations with the contact mode score. Comput. Biol. Chem. 2016, 64, 403–413. [Google Scholar] [CrossRef]
- Chandra, B.T.M.; Rajesh, S.S.; Bhaskar, B.V.; Devi, S.; Rammohan, A.; Sivaraman, T.; Rajendra, W. Molecular docking, molecular dynamics simulation, biological evaluation and 2D QSAR analysis of flavonoids from Syzygium alternifolium as potent anti-Helicobacter pylori agents. RSC Adv. 2017, 7, 18277–18292. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).