

Dynamics of Fatty Acids in Pikeperch (Sander lucioperca) Larvae and Juveniles during Early Rearing and Weaning in a Commercial RAS—Implications for Dietary Refinement

 ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Hatching and Rearing of Pikeperch in a Commercial Recirculating Aquaculture System (RAS)
2.2. Feed and Feeding Regime
2.3. Determination of Fish Larvae Mass
2.4. Sampling and Fatty Acid Analyses
2.5. Statistical Analyses
3. Results
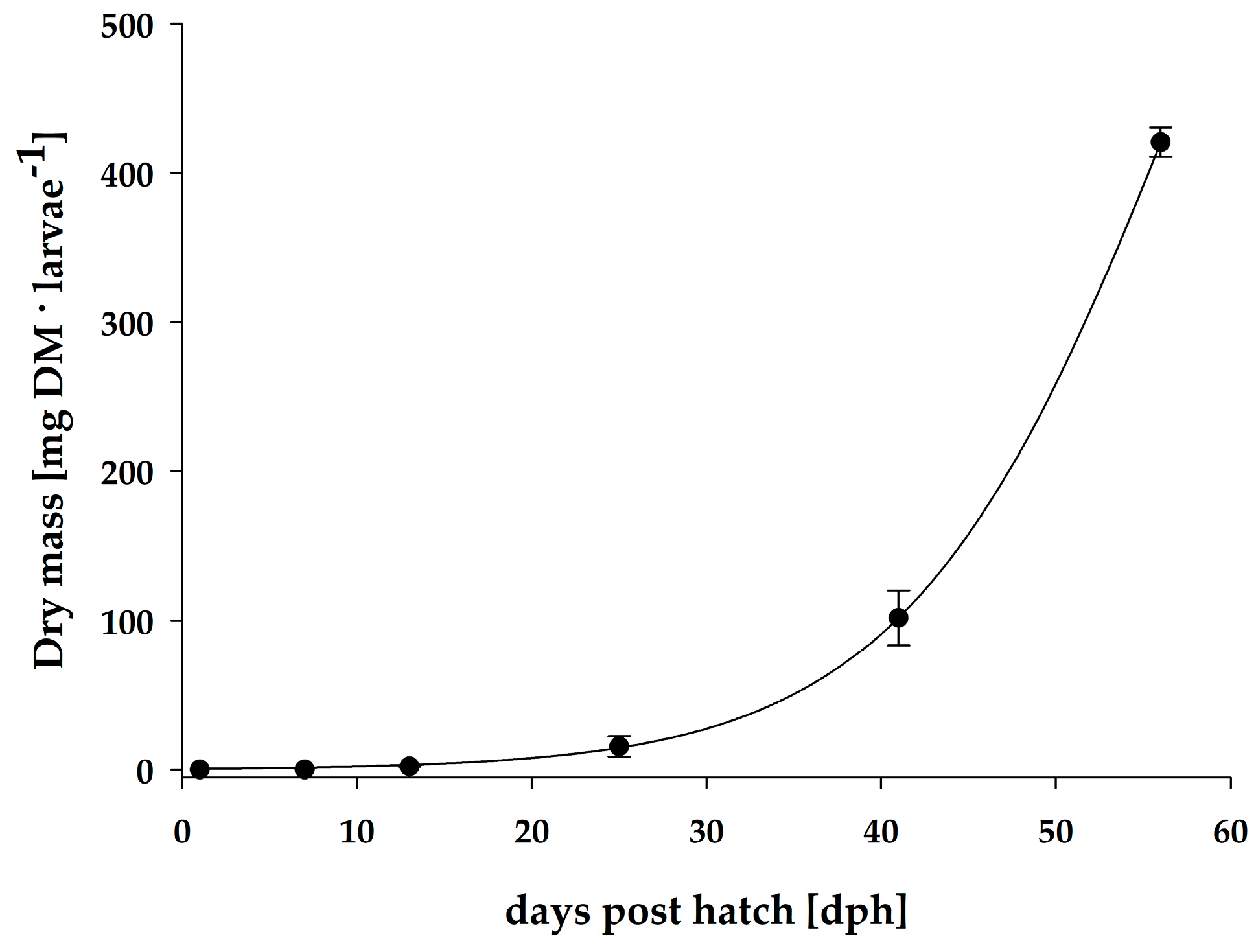
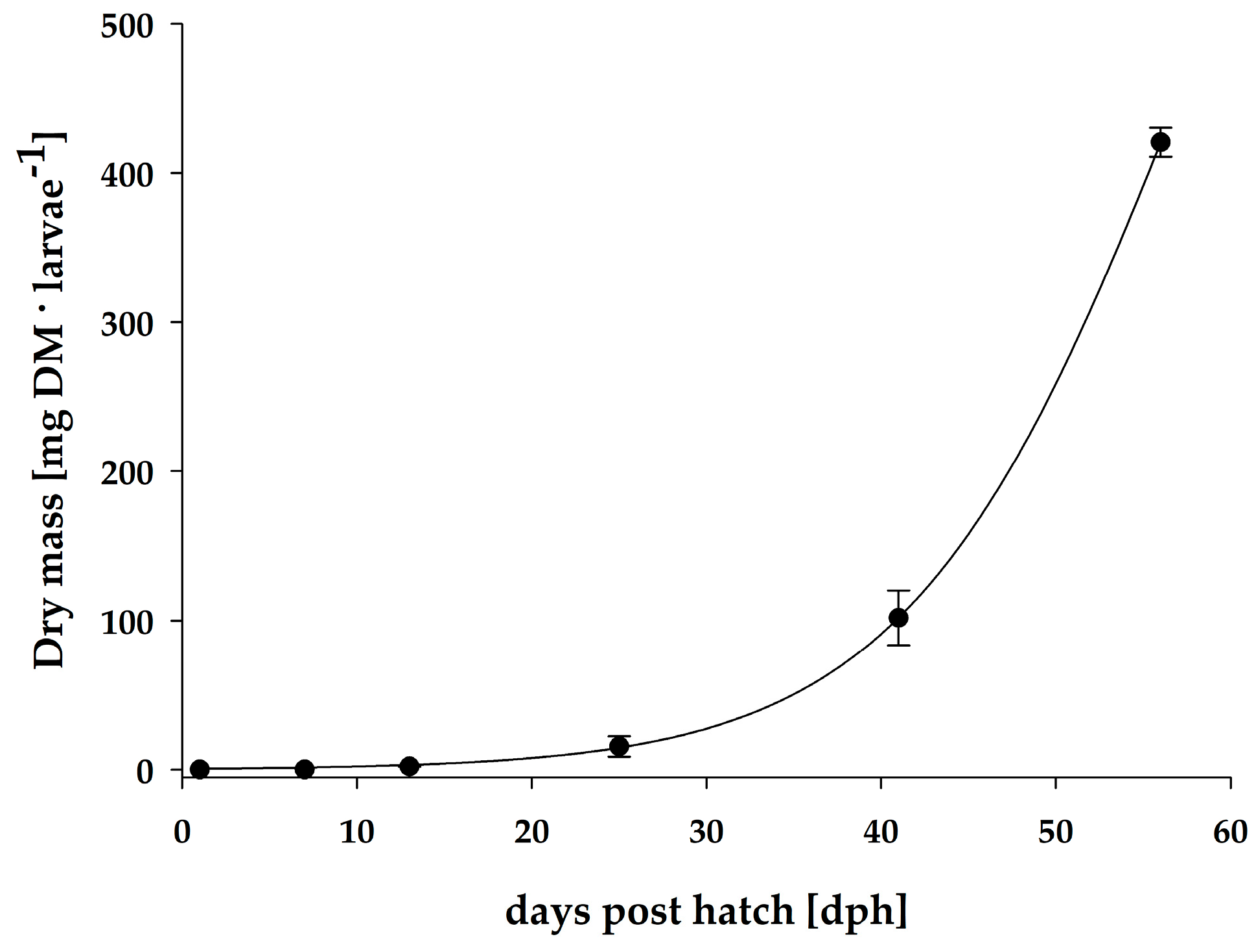
3.1. Growth
3.2. Fatty Acid Composition of Pikeperch Larvae
4. Discussion
4.1. Effect of Feeding on the Growth of Pikeperch Larvae and Juveniles
4.2. Effect of Feed on Pikeperch Fatty Acid Composition
4.2.1. Period 1–7 dph
4.2.2. Period 7–13 dph
4.2.3. Period 13–25 dph
4.2.4. Period from dph 25 Onward
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Boisneau, P.; Mennesson-Boisneau, C. Inland commercial fisheries management in France. Fish. Manag. Ecol. 2001, 8, 303–310. [Google Scholar] [CrossRef]
- Wedekind, H.; Hilge, V.; Steffens, W. Present status, and social and economic significance of inland fisheries in Germany. Fish. Manag. Ecol. 2001, 8, 405–414. [Google Scholar] [CrossRef]
- Schulz, C.; Huber, M.; Ogunji, J.; Rennert, B. Effects of varying dietary protein to lipid ratios on growth performance and body composition of juvenile pike perch (Sander lucioperca). Aquac. Nutr. 2008, 14, 166–173. [Google Scholar] [CrossRef]
- Steenfeldt, S. Culture Methods of Pikeperch Early Life Stages. In Biology and Culture of Percid Fishes: Principles and Practices, 1st ed.; Kestemont, P., Dabrowski, K., Summerfelt, R.C., Eds.; Springer: Dordrecht, The Netherlands, 2015; pp. 295–312. [Google Scholar]
- Summerfelt, R.C. Intensive Culture of Walleye Fry. In Walleye Culture Manual; Summerfelt, R.C., Ed.; NCRAC Culture Series 101; Iowa State University: Ames, IA, USA, 1996. [Google Scholar]
- Ljunggren, L.; Staffan, F.; Falk, S.; Linden, B.; Mendes, J. Weaning of juvenile pikeperch, Stizostedion lucioperca L., and perch, Perca fluviatilis L., to formulated feed. Aquac. Res. 2003, 34, 281–287. [Google Scholar] [CrossRef]
- Kestemont, P.; Xueliang, X.; Hamza, N.; Maboudou, J.; Imorou Toko, I. Effect of weaning age and diet on pikeperch larviculture. Aquaculture 2007, 264, 197–204. [Google Scholar] [CrossRef]
- Ostaszewska, T.; Dabrowski, K.; Czuminska, K.; Olech, W.; Olejniczak, M. Rearing of pikeperch larvae using formulated diets-first success with starter feeds. Aquac. Res. 2005, 36, 1167–1176. [Google Scholar] [CrossRef]
- Abi-Ayad, S.-M.E.-A.; Boutiba, Z.; Mélard, C.; Kestemont, P. Dynamics of total body fatty acids during early ontogeny of pikeperch (Sander lucioperca) larvae. Fish Physiol. Biochem. 2004, 30, 129–136. [Google Scholar] [CrossRef]
- Lund, I.; Steenfeldt, S.J. The effects of dietary long-chain essential fatty acids on growth and stress tolerance in pikeperch larvae (Sander lucioperca L.). Aquac. Nutr. 2011, 17, 191–199. [Google Scholar] [CrossRef]
- Hamza, N.; Kestemont, P.; Khemis, I.B.; Mhetli, M.; Cahu, C. Effect of different sources and levels of dietary phospholipids on performances and fatty acid composition of pikeperch (Sander lucioperca) larvae. Aquac. Nutr. 2012, 18, 249–257. [Google Scholar] [CrossRef]
- Lund, I.; Skov, P.V.; Hansen, B.W. Dietary supplementation of essential fatty acids in larval pikeperch (Sander lucioperca); short and long term effects on stress tolerance and metabolic physiology. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2012, 162, 340–348. [Google Scholar] [CrossRef]
- Lund, I.; Höglund, E.; Ebbesson, L.O.E.; Skov, P.V. Dietary LC-PUFA deficiency early in ontogeny induces behavioural changes in pike perch (Sander lucioperca) larvae and fry. Aquaculture 2014, 432, 453–461. [Google Scholar] [CrossRef]
- Kestemont, P.; Henrotte, E. Nutritional Requirements and Feeding of Broodstock and Early Life Stages of Eurasian Perch and Pikeperch. In Biology and Culture of Percid Fishes: Principles and Practices, 1st ed.; Kestemont, P., Dabrowski, K., Summerfelt, R.C., Eds.; Springer: Dordrecht, The Netherlands, 2015; pp. 539–564. [Google Scholar]
- Heming, T.A.; Buddington, R.K. Yolk absorption in embryonic and larval fishes. In Fish Physiology: Vol. XI. The Physiology of Developing Fish Part A: Eggs and Larvae; Hoar, W.S., Randall, D.J., Eds.; Academic Press: London, UK; New York, NY, USA, 1988. [Google Scholar]
- Palm, H.W.; Bischoff, A.A.; Wranik, C.; Augustin, C.; Kubitz, M. Endbericht “Entwicklung Eines Zooplankton-Reaktors zur Unterstützung der Fischlarvenaufzucht Relevanter Zielfischarten in Mecklenburg-Vorpommern (MV)”. Available online: https://www.aquakultur-mv.de/static/AQUA/Dokumente/Forschen/Zooreaktor_Endbericht_2015_Finalversion.pdf (accessed on 27 June 2023).
- Ackman, R.G. Characteristics of the fatty acid composition and biochemistry of some fresh-water fish oils and lipids in comparison with marine oils and lipids. Comp. Biochem. Physiol. 1967, 22, 907–922. [Google Scholar] [CrossRef]
- Henderson, R.J.; Tocher, D.R. The lipid composition and biochemistry of freshwater fish. Prog. Lipid Res. 1987, 26, 281–347. [Google Scholar] [PubMed]
- Sargent, J.R.; Henderson, R.J.; Tocher, D.R. The Lipids. In Fish Nutrition, 2nd ed.; Halver, J.E., Ed.; Academic Press: San Diego, CA, USA, 1989; pp. 154–219. [Google Scholar]
- Fink, P. Invasion of quality: High amounts of essential fatty acids in the invasive Ponto-Caspian mysid Limnomysis benedeni. J. Plankton Res. 2013, 35, 907–913. [Google Scholar] [CrossRef]
- Kattner, G.; Fricke, H.S.G. Simple gas-liquid chromatography method for simultaneous determination of fatty acids and alcohols in wax esters of marine organisms. J. Chromatogr. 1986, 361, 263–268. [Google Scholar] [CrossRef]
- Colchen, T.; Gisbert, E.; Kraus, D.; Pasquet, A.; Fontaine, P. Improving pikeperch larviculture by combining environmental, feeding and population factors. Aquac. Rep. 2020, 17, 100337. [Google Scholar] [CrossRef]
- Péter, G.; Lukic, J.; Alvestad, R.; Horváth, Z.; Nagy, Z.; Rónyai, A.; Bársony, P.; Ljubobratovic, U. Nursing of Pike-Perch (Sander lucioperca) in Recirculating Aquaculture System (RAS) Provides Growth Advantage in Juvenile Growth Phase. Animals 2023, 13, 347. [Google Scholar] [CrossRef]
- Ballesteros-Redondo, L.; Palm, H.W.; Bährs, H.; Wacker, A.; Bischoff, A.A. Pikeperch larviculture (Sander lucioperca [L., 1758]) with Brachionus plicatilis (Mueller, 1786) (Rotifera) and Apocyclops panamensis (Marsh, 1913) (Copepoda). J. World Aquac. Soc. 2023, 54, 1026–1039. [Google Scholar] [CrossRef]
- Bischoff, A.A.; Kubitz, M.; Wranik, C.M.; Ballesteros-Redondo, L.; Fink, P.; Palm, H.W. The Effect of Brachionus calyciflorus (Rotifera) on Larviculture and Fatty Acid Composition of Pikeperch (Sander lucioperca (L.)) Cultured under Pseudo-GreenWater Conditions. Sustainability 2022, 14, 6607. [Google Scholar] [CrossRef]
- Bischoff, A.A.; Kubitz, M.; Wranik, C.M.; Pfefferkorn, H.; Augustin, C.B.; Hagen, W.; Palm, H.W. Fatty acid utilization of pikeperch (Sander lucioperca (Linnaeus, 1758)) larvae under starvation conditions during early development. Bull. Fish Biol. 2018, 17, 59–73. [Google Scholar]
- Sargent, J.R.; Tocher, D.R.; Bell, J.G. The Lipids. In Fish Nutrition, 3rd ed.; Halver, J.E., Ed.; Academic Press: Amsterdam, The Netherlands, 2002; pp. 181–257. [Google Scholar]
- Henrotte, E.; Overton, J.L.; Kestemont, P. Effects of dietary n-3 and n-6 fatty acid levels on egg and larval quality of Eurasian perch. Cybium 2008, 32, 271–272. [Google Scholar]
- Tocher, D.R. Metabolism and Functions of Lipids and Fatty Acids in Teleost Fish. Rev. Fish. Sci. 2003, 11, 107–184. [Google Scholar] [CrossRef]
- Sargent, J.R.; Bell, J.G.; Bell, M.V.; Henderson, R.J.; Tocher, D.R. Requirement criteria for essential fatty acids. J. Appl. Ichthyol. 1995, 11, 183–198. [Google Scholar] [CrossRef]
- Mourente, G. Accumulation of DHA (docosahexaenoic acid; 22:6n-3) in larval and juvenile fish brain. In The Big Fish Bang; Institute of Marine Research: Bergen, Norway, 2003; pp. 239–248. [Google Scholar]
- Henrotte, E.; Mandiki, R.S.N.M.; Prudencio, A.T.; Vandecan, M.; Mélard, C.; Kestemont, P. Egg and larval quality, and egg fatty acid composition of Eurasian perch breeders (Perca fluviatilis) fed different dietary DHA/EPA/AA ratios. Aquac. Res. 2010, 41, 53–61. [Google Scholar] [CrossRef]
- Schulz, C.; Knaus, U.; Wirth, M.; Rennert, B. Effects of varying dietary fatty acid profile on growth performance, fatty acid, body and tissue composition of juvenile pike perch (Sander lucioperca). Aquac. Nutr. 2005, 11, 403–413. [Google Scholar] [CrossRef]
- Imentai, A.; Raskovic, B.; Steinbach, C.; Rahimnejad, S.; Yanes-Roca, C.; Policar, T. Effects of first feeding regime on growth performance, survival rate and development of digestive system in pikeperch (Sander lucioperca) larvae. Aquaculture 2020, 529, 735636. [Google Scholar] [CrossRef]
- Imentai, A.; Gilannejad, N.; Martinez-Rodriguez, G.; Lopez, F.J.M.; Martinez, F.P.; Penka, T.; Dzyuba, V.; Dadras, H.; Policar, T. Effects of first feeding regime on gene expression and enzyme activity in pikeperch (Sander lucioperca) larvae. Front. Mar. Sci. 2022, 9, 864536. [Google Scholar] [CrossRef]
- Hamza, N.; Ostaszewska, T.; Kestemont, P. Development and Functionality of the Digestive System in Percid Fishes Early Life Stages. In Biology and Culture of Percid Fishes: Principles and Practices, 1st ed.; Kestemont, P., Dabrowski, K., Summerfelt, R.C., Eds.; Springer: Dordrecht, The Netherlands, 2015; pp. 239–264. [Google Scholar]
- Dhert, P.; Lavens, P.; Duray, M.; Sorgeloos, P. Improved larval survival at metamorphosis of Asian seabass (Lates calcarifer) using ω3-HUFA-enriched live food. Aquaculture 1990, 90, 63–74. [Google Scholar] [CrossRef]

{kind=link}
| Days Post Hatch | Fish Larval Feeds |
|---|---|
| 0–3 | no exogenous feed |
| 4–7 | Micro Artemia (AF430) |
| 8 | Micro Artemia and Artemia (GSL) |
| 9–10 | Micro Artemia, Artemia and microdiets (O.range Start, INVE) |
| 11–20 | Artemia and microdiets |
| 21–56 | microdiets |
| Fatty Acids | 4–10 dph | 8–20 dph | 9–56 dph |
|---|---|---|---|
| Micro Artemia | Artemia spp. | microdiet | |
| 14:0 | 2.3 ± 0.4 | 0.6 ± 0.4 | 1.3 ± 0.4 |
| 16:0 | 18.8 ± 1.3 | 13.7 ± 0.7 | 11.6 ± 1.5 |
| 18:0 | 6.1 ± 0.4 | 6.2 ± 0.4 | 5.2 ± 0.5 |
| ∑SFA | 28.0 ± 2.2 | 20.4 ± 0.8 | 18.0 ± 2.3 |
| 16:1n-7 | 17.9 ± 1.4 | 3.0 ± 0.1 | 2.1 ± 0.3 |
| 18:1n-9 | 25.0 ± 1.5 | 23.7 ± 1.5 | 8.3 ± 1.0 |
| 18:1n-7 | 17.2 ± 1.0 | 7.8 ± 0.5 | 1.8 ± 0.2 |
| 20:1n-9 | 0.7 ± 0.0 | 0.6 ± 0.0 | 3.1 ± 0.4 |
| 22:1n-11 | 0.0 ± 0.0 | 0.0 ± 0.0 | 3.0 ± 0.3 |
| ∑MUFA | 62.4 ± 4.0 | 35.9 ± 2.1 | 18.6 ± 2.2 |
| 16:3n-4 | 11.3 ± 0.8 | 1.2 ± 0.1 | 0.3 ± 0.0 |
| 18:2n-6 | 4.9 ± 0.4 | 7.5 ± 0.6 | 8.2 ± 1.1 |
| 18:2n-4 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 |
| 18:3n-3 | 1.8 ± 0.2 | 34.4 ± 3.5 | 1.1 ± 0.2 |
| 18:4n-3 | 0.9 ± 0.1 | 5.5 ± 0.6 | 0.8 ± 0.1 |
| 20:4n-6 | 4.4 ± 0.5 | 0.8 ± 0.1 | 0.5 ± 0.1 |
| 20:5n-3 | 15.4 ± 1.8 | 1.9 ± 0.3 | 5.2 ± 0.9 |
| 22:6n-3 | 0.0 ± 0.0 | 0.0 ± 0.0 | 9.0 ± 1.8 |
| ∑PUFA | 41.9 ± 4.0 | 54.1 ± 5.3 | 27.2 ± 4.4 |
| ∑TFAs | 132.2 ± 10.1 | 110.4 ± 8.0 | 63.8 ± 8.3 |
| ∑C18 PUFA | 8.5 ± 0.7 | 48.0 ± 4.7 | 10.1 ± 1.3 |
| ∑C20 + 22 HUFA | 19.8 ± 2.3 | 4.5 ± 0.6 | 16.8 ± 3.1 |
| ∑n-3 | 18.1 ± 2.1 | 43.6 ± 4.57 | 17.55 ± 3.24 |
| ∑n-6 | 10.2 ± 0.9 | 9.0 ± 0.7 | 8.6 ± 1.1 |
| n-3/n-6 | 1.8:1.0 | 4.9:1.0 | 2.0:1.0 |
| DHA/EPA | 0.0:15.4 | 0.0:1.9 | 1.7:1.0 |
| EPA/ARA | 3.5:1.0 | 2.3:1.0 | 11.5:1.0 |
| 18:3n-3/18:2n-6 | 1.0:2.8 | 4.6:1.0 | 1.0:7.4 |
| SFA/MUFA | 1.0:2.2 | 1.0:1.8 | 1.0:1.0 |
| SFA/PUFA | 1.0:1.5 | 1.0:2.7 | 1.0:1.5 |
| MUFA/PUFA | 1.5:1.0 | 1.0:1.5 | 1.0:1.5 |
| Pikeperch of a Commercial Recirculating Aquaculture System (RAS) | |||||
|---|---|---|---|---|---|
| Days post hatch | Dry mass (mg·Ind−1) | Fulton’s condition index | SGR (%·day−1) | Total fatty acids (µg·mg−1 DM) | Total fatty acids (µg·Ind−1) |
| 1 | 0.1 ± 0.0 | 0.138 | - | 97.8 ± 7.8 | 13.0 ± 0.7 |
| 7 | 0.3 ± 0.0 | 0.085 | 10.9 | 90.9 ± 4.3 | 23.0 ± 4.3 * |
| 13 | 2.2 ± 0.3 | 0.118 | 36.3 | 95.0 ± 2.9 | 212.7 ± 46.4 * |
| 25 | 15.6 ± 6.8 | # n.d. | 16.3 | 39.8 ± 7.0 | 59.1 ± 14.6 * |
| 41 | 101.8 ± 18.4 | # n.d. | 11.7 | 85.5 ± 22.0 | 425.8 ± 239.0 |
| 56 | 420.5 ± 9.7 | # n.d. | 9.5 | 81.4 ± 20.6 | 473.2 ± 144.6 |
| Fatty Acids | dph 1 | dph 7 | dph 13 | dph 25 | dph 41 | dph 56 |
|---|---|---|---|---|---|---|
| 16:0 | 7.9 ± 0.7 a | 13.0 ± 0.6 b | 13.1 ± 0.6 b | 8.4 ± 1.2 a | 16.9 ± 4.3 b | 14.6 ± 3.5 b |
| 18:0 | 2.8 ± 0.3 a | 5.5 ± 0.3 b | 6.6 ± 0.4 c | 4.1 ± 0.7 b | 5.3 ± 0.8 b | 3.6 ± 0.6 a |
| ∑SFA | 11.3 ± 1.0 a | 20.0 ± 0.9 b | 21.0 ± 1.0 b | 12.8 ± 1.9 a | 24.1 ± 5.8 b | 20.1 ± 4.8 b |
| 16:1n-9 | 2.2 ± 0.2 a | 1.2 ± 0.1 b | 1.3 ± 0.1 b | 0.4 ± 0.1 c | 0.4 ± 0.1 c | 0.4 ± 0.2 c |
| 16:1n-7 | 4.4 ± 0.3 a | 9.0 ± 0.4 b | 5.4 ± 0.2 c | 1.0 ± 0.2 d | 3.2 ± 1.2 a | 3.9 ± 1.0 ac |
| 18:1n-9 | 18.6 ± 1.4 ab | 16.7 ± 0.7 a | 20.3 ± 0.7 b | 6.5 ± 1.1 c | 16.0 ± 4.6 a | 19.8 ± 5.1 ab |
| 18:1n-7 | 4.7 ± 0.4 a | 8.9 ± 0.3 b | 8.4 ± 0.5 b | 1.9 ± 0.3 c | 2.8 ± 0.8 c | 2.7 ± 0.7 c |
| ∑MUFA | 30.4 ± 2.3 a | 36.6 ± 1.7 b | 35.9 ± 1.2 b | 10.3 ± 1.7 c | 26.2 ± 7.7 a | 31.3 ± 8.2 a |
| 16:3n-4 | 1.7 ± 0.2 a | 4.9 ± 0.2 b | 3.6 ± 0.1 c | 0.5 ± 0.1 d | 0.0 ± 0.0 d | 0.2 ± 0.1 d |
| 18:2n-6 | 10.9 ± 0.9 a | 4.4 ± 0.2 b | 5.1 ± 0.2 c | 3.3 ± 0.5 d | 11.9 ± 3.7 a | 12.1 ± 3.2 a |
| 18:3n-3 | 7.7 ± 0.6 a | 2.0 ± 0.1 b | 12.8 ± 0.8 c | 1.4 ± 0.3 d | 1.7 ± 0.6 d | 1.8 ± 0.5 d |
| 20:4n-6 | 4.5 ± 0.4 a | 4.0 ± 0.2 a | 2.4 ± 0.1 b | 1.0 ± 0.2 c | 0.7 ± 0.1 d | 0.6 ± 0.1 d |
| 20:5n-3 | 4.2 ± 0.3 a | 8.5 ± 0.5 b | 5.6 ± 0.4 c | 2.2 ± 0.5 d | 4.7 ± 1.0 a | 3.7 ± 1.0 a |
| 22:6n-3 | 16.3 ± 1.3 a | 5.8 ± 0.5 b | 1.2 ± 0.1 c | 6.5 ± 1.5 b | 13.4 ± 2.6 a | 8.5 ± 2.3 b |
| ∑PUFA | 56.1 ± 4.6 a | 34.2 ± 1.7 b | 38.0 ± 1.5 b | 16.7 ± 3.4 c | 35.2 ± 8.6 b | 30.0 ± 7.8 b |
| ∑TFAs | 97.8 ± 7.8 a | 90.9 ± 4.3 a | 95.0 ± 2.9 a | 39.8 ± 7.0 b | 85.5 ± 22.0 a | 81.4 ± 20.6 a |
| ∑C18 PUFA | 22.7 ± 2.0 a | 7.9 ± 0.4 b | 21.3 ± 1.1 a | 5.1 ± 0.8 b | 14.5 ± 4.6 c | 15.0 ± 4.0 c |
| ∑C20 + 22 HUFA | 31.3 ± 2.6 a | 20.5 ± 1.1 b | 13.1 ± 0.7 c | 11.1 ± 2.5 c | 20.7 ± 4.0 b | 14.8 ± 3.9 c |
| ∑n-3 | 33.0 ± 2.5 a | 18.5 ± 1.1 b | 25.9 ± 1.4 c | 11.6 ± 2.6 d | 22.5 ± 4.8 bc | 16.5 ± 4.5 b |
| ∑n-6 | 19.4 ± 1.8 a | 9.6 ± 0.4 b | 8.5 ± 0.2 c | 4.4 ± 0.7 d | 12.6 ± 3.8 e | 13.3 ± 3.4 e |
| n-3/n-6 | 1.7:1.0 | 1.9:1.0 | 3.0:1.0 | 2.6:1.0 | 1.8:1.0 | 1.2:1.0 |
| DHA/EPA | 3.9:1.0 | 1.0:1.5 | 1.0:4.7 | 3.0:1.0 | 2.9:1.0 | 2.3:1.0 |
| EPA/ARA | 1.0:1.1 | 2.1:1.0 | 2.3:1.0 | 2.2:1.0 | 6.7:1.0 | 6.2:1.0 |
| 18:3n-3/18:2n-6 | 1.0:1.4 | 1.0:2.2 | 2.5:1.0 | 1.0:2.4 | 1.0:7.0 | 1.0:6.7 |
| SFA/MUFA | 1.0:2.7 | 1.0:1.8 | 1.0:1.7 | 1.2:1.0 | 1.0:1.1 | 1.0:1.6 |
| SFA/PUFA | 1.0:5.0 | 1.0:1.7 | 1.0:1.8 | 1.0:1.3 | 1.0:1.5 | 1.0:1.5 |
| MUFA/PUFA | 1.0:1.8 | 1.1:1.0 | 1.0:1.1 | 1.0:1.6 | 1.0:1.3 | 1.0:1.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bischoff, A.A.; Kubitz, M.; Ballesteros-Redondo, L.; Stüeken, M.; Rapp, T.; Fink, P.; Hagen, W.; Palm, H.W. Dynamics of Fatty Acids in Pikeperch (Sander lucioperca) Larvae and Juveniles during Early Rearing and Weaning in a Commercial RAS—Implications for Dietary Refinement. Fishes 2023, 8, 444. https://doi.org/10.3390/fishes8090444
Bischoff AA, Kubitz M, Ballesteros-Redondo L, Stüeken M, Rapp T, Fink P, Hagen W, Palm HW. Dynamics of Fatty Acids in Pikeperch (Sander lucioperca) Larvae and Juveniles during Early Rearing and Weaning in a Commercial RAS—Implications for Dietary Refinement. Fishes. 2023; 8(9):444. https://doi.org/10.3390/fishes8090444
Chicago/Turabian StyleBischoff, Adrian A., Melanie Kubitz, Laura Ballesteros-Redondo, Marcus Stüeken, Tobias Rapp, Patrick Fink, Wilhelm Hagen, and Harry Wilhelm Palm. 2023. "Dynamics of Fatty Acids in Pikeperch (Sander lucioperca) Larvae and Juveniles during Early Rearing and Weaning in a Commercial RAS—Implications for Dietary Refinement" Fishes 8, no. 9: 444. https://doi.org/10.3390/fishes8090444
APA StyleBischoff, A. A., Kubitz, M., Ballesteros-Redondo, L., Stüeken, M., Rapp, T., Fink, P., Hagen, W., & Palm, H. W. (2023). Dynamics of Fatty Acids in Pikeperch (Sander lucioperca) Larvae and Juveniles during Early Rearing and Weaning in a Commercial RAS—Implications for Dietary Refinement. Fishes, 8(9), 444. https://doi.org/10.3390/fishes8090444