Macroparasite Communities with Special Attention to Invasive Helminths in European Eels Anguilla anguilla from Freshwaters and Brackish Lagoons of a Mediterranean Island

 ,
,  ,
,
Abstract
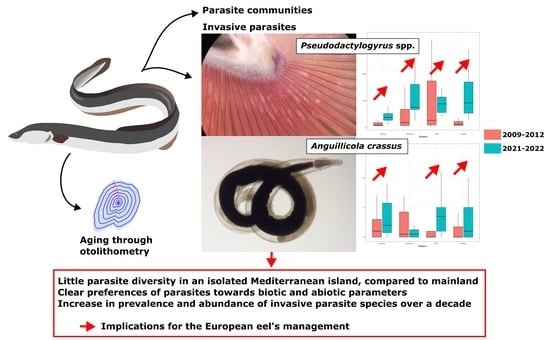
1. Introduction
2. Material and Methods
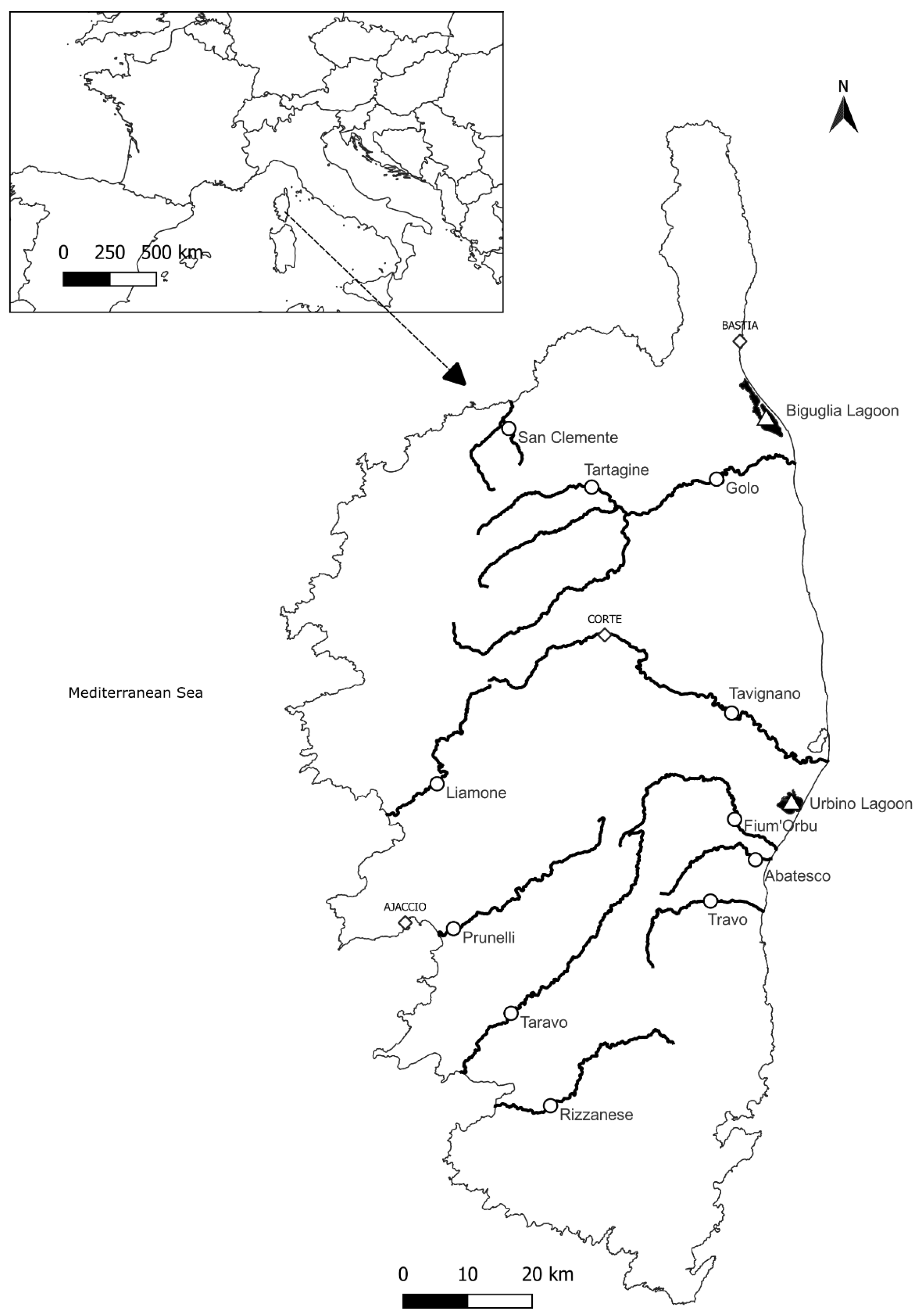
2.1. Sample Collection and Study Site
2.2. Morphometry and Parasitological Examination
2.3. Otolithometry
2.4. Statistical Analysis
3. Results
3.1. Parasite Diversity in European Eels in Corsica
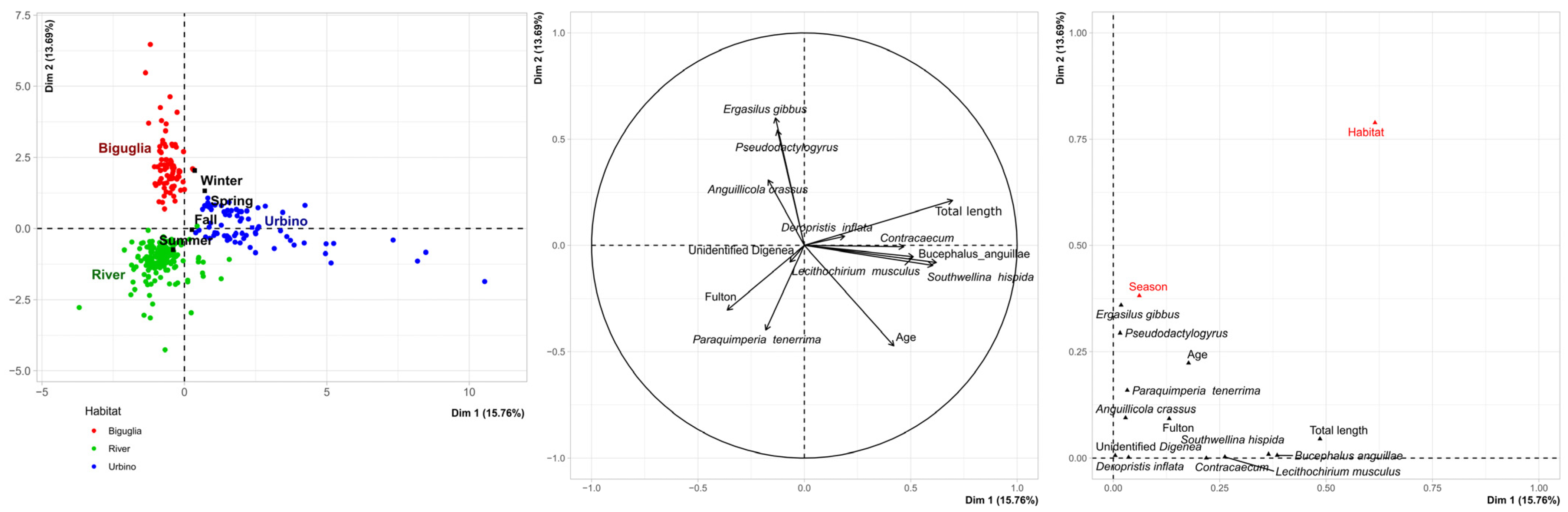
3.2. Analysis of Parasite Community of European Eels in Corsica
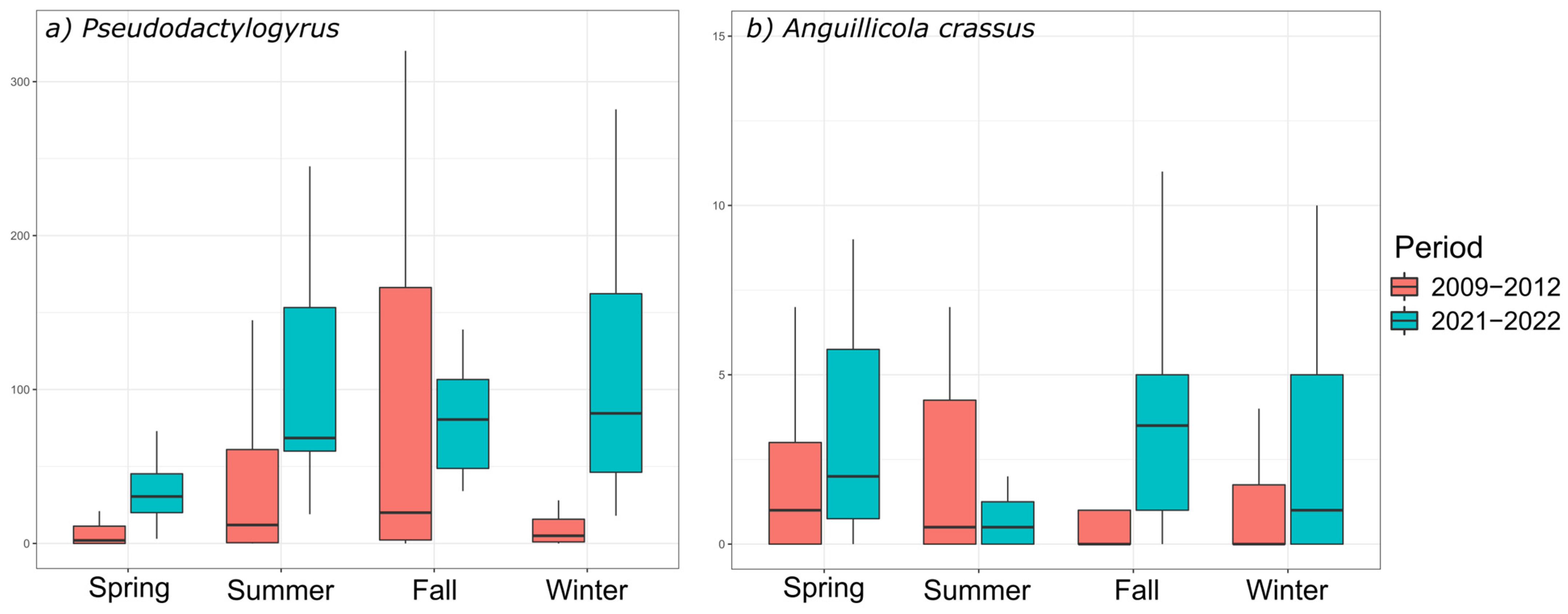
3.3. Seasonal Variations of Parasite Communities in Coastal Lagoons
3.4. Influence of Silvering Stage and Age on Parasite Communities in Brackish and Freshwater Sites
4. Discussion
4.1. Low Parasite Diversity in Mediterranean Islands
4.2. Habitat Specificity of A. anguilla Parasites
4.2.1. Parasites in Eels from Brackish Coastal Lagoons
4.2.2. Parasites in Eels from Freshwater Sites
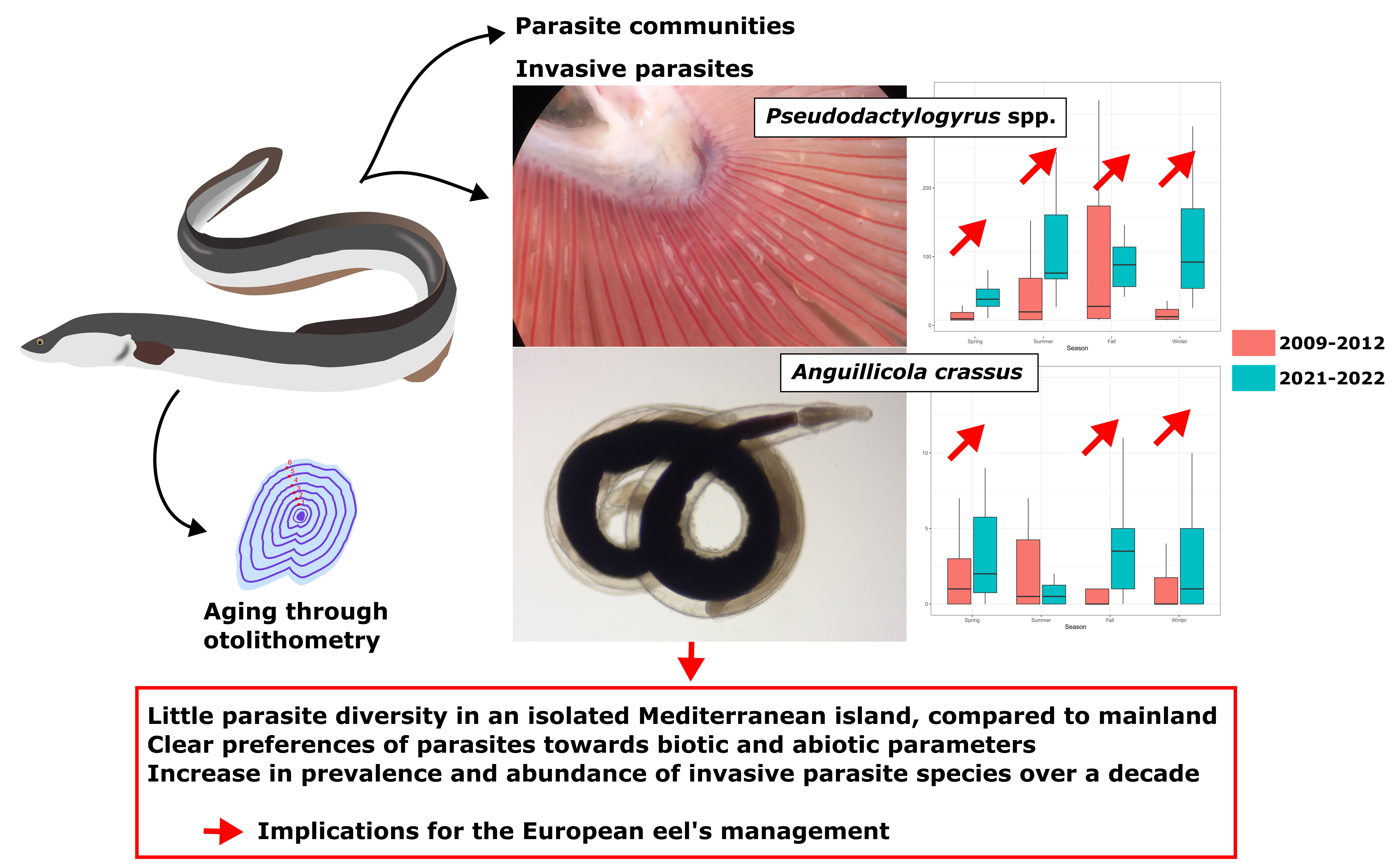
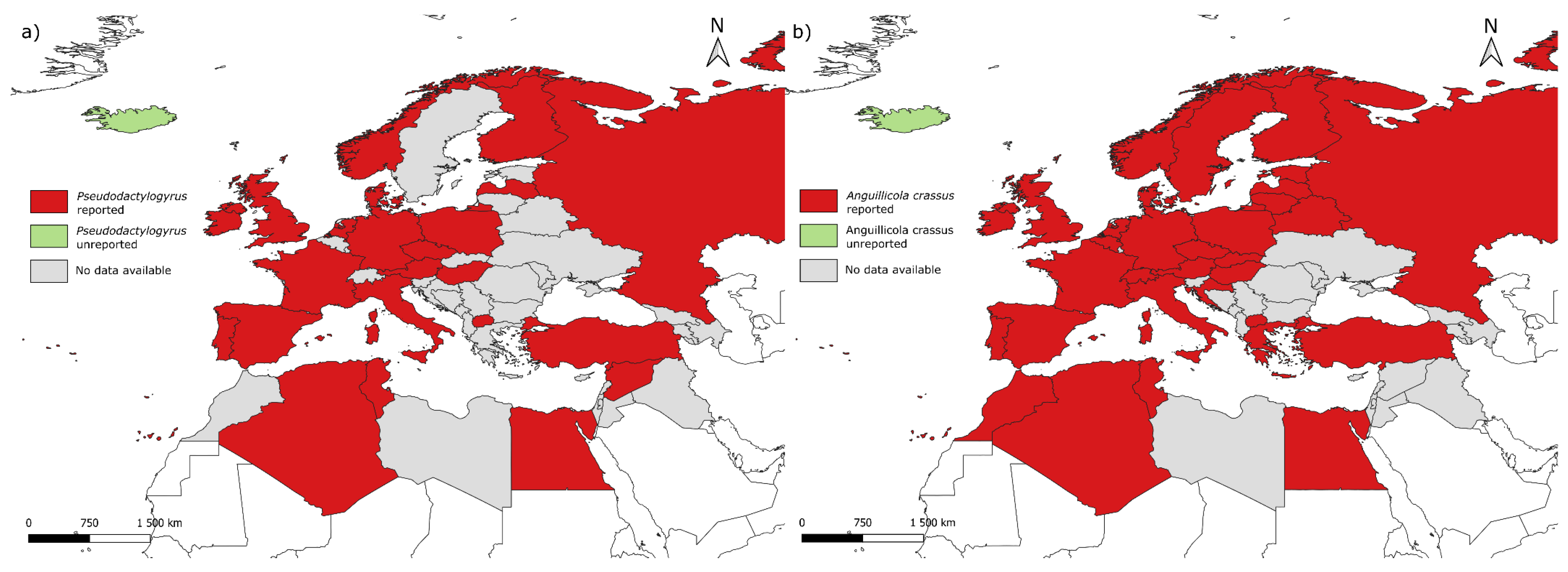
4.3. Invasive Species
4.3.1. Pseudodactylogyrus anguillae and P. bini
4.3.2. Anguillicola crassus
4.3.3. Implications for the Management of the European Eel
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tesch, T. The Eel, 5th ed.; John Wiley & Sons: Oxford, UK, 2003; ISBN 978-1-4051-7343-8. [Google Scholar]
- Jacoby, D.; Gollock, M. Anguilla anguilla. In The IUCN Red List of Threatened Species; ICUN: Gland, Switzerland, 2014; p. e.T60344A45833138. [Google Scholar] [CrossRef]
- Pike, C.; Crook, V.; Gollock, M. IUCN Red List of Threatened Species: Anguilla anguilla. In IUCN Red List of Threatened Species; ICUN: Gland, Switzerland, 2020. [Google Scholar] [CrossRef]
- EC. Council Regulation (EC) No 1100/2007 of 18 September 2007 Establishing Measures for the Recovery of the Stock of European Eel; European Council: Brussels, Belgium, 2007; Volume 248. [Google Scholar]
- Drouineau, H.; Durif, C.; Castonguay, M.; Mateo, M.; Rochard, E.; Verreault, G.; Yokouchi, K.; Lambert, P. Freshwater Eels: A Symbol of the Effects of Global Change. Fish Fish. 2018, 19, 903–930. [Google Scholar] [CrossRef]
- Taraschewski, H. Hosts and Parasites as Aliens. J. Helminthol. 2006, 80, 99–128. [Google Scholar] [CrossRef]
- Dunn, A.M. Chapter 7 Parasites and Biological Invasions. In Advances in Parasitology; Elsevier: Amsterdam, The Netherlands, 2009; Volume 68, pp. 161–184. ISBN 978-0-12-374787-7. [Google Scholar]
- Daszak, P.; Cunningham, A.A.; Hyatt, A.D. Emerging Infectious Diseases of Wildlife—Threats to Biodiversity and Human Health. Science 2000, 287, 443–449. [Google Scholar] [CrossRef] [PubMed]
- Jakob, E.; Hanel, R.; Klimpel, S.; Zumholz, K. Salinity Dependence of Parasite Infestation in the European Eel Anguilla anguilla in Northern Germany. ICES J. Mar. Sci. 2009, 66, 358–366. [Google Scholar] [CrossRef]
- Ternengo, S.; Levron, C.; Desideri, F.; Marchand, B. Parasite Communities in European Eels Anguilla anguilla (Pisces, Teleostei) from a Corsican Coastal Pond. Vie Milieu 2005, 55, 1–6. [Google Scholar]
- Filippi, J.-J.; Quilichini, Y.; Foata, J.; Marchand, B. Influence of Site, Season, Silvering Stage, and Length on the Parasites of the European Eel Anguilla anguilla in Two Mediterranean Coastal Lagoons of the Island of Corsica, France Using Indicator Species Method. Parasitol. Res. 2013, 112, 2959–2969. [Google Scholar] [CrossRef] [PubMed]
- Culurgioni, J.; Sabatini, A.; De Murtas, R.; Mattiucci, S.; Figus, V. Helminth Parasites of Fish and Shellfish from the Santa Gilla Lagoon in Southern Sardinia, Italy. J. Helminthol. 2014, 88, 489–498. [Google Scholar] [CrossRef]
- Culurgioni, J.; Figus, V.; Cabiddu, S.; De Murtas, R.; Cau, A.; Sabatini, A. Larval Helminth Parasites of Fishes and Shellfishes from Santa Gilla Lagoon (Sardinia, Western Mediterranean), and Their Use as Bioecological Indicators. Estuaries Coasts 2015, 38, 1505–1519. [Google Scholar] [CrossRef]
- Filippi, J.-J. Étude Parasitologique de Anguilla anguilla dans Deux Lagunes de Corse et Etude Ultrastructurale du Tégument de Trois Digènes Parasites de Cette Anguille. Ph.D. Thesis, Université de Corse Pasquale Paoli, Corte, France, 2013. [Google Scholar]
- Kennedy, C.R. Metapopulation and Community Dynamics of Helminth Parasites of Eels Anguilla anguilla in the River Exe System. Parasitology 2001, 122, 689–698. [Google Scholar] [CrossRef]
- Aguilar, A.; Álvarez, M.F.; Leiro, J.M.; Sanmartín, M.L. Parasite Populations of the European Eel (Anguilla anguilla L.) in the Rivers Ulla and Tea (Galicia, Northwest Spain). Aquaculture 2005, 249, 85–94. [Google Scholar] [CrossRef]
- Bracamonte, S.E.; Knopf, K.; Monaghan, M.T. Encapsulation of Anguillicola crassus Reduces the Abundance of Adult Parasite Stages in the European Eel (Anguilla anguilla). J. Fish Dis. 2021, 44, 771–782. [Google Scholar] [CrossRef]
- Fazio, G.; Sasal, P.; Lecomte-Finiger, R.; Da Silva, C.; Fumet, B.; Moné, H. Macroparasite Communities in European Eels, Anguilla anguilla, from French Mediterranean Lagoons, with Special Reference to the Invasive Species Anguillicola crassus and Pseudodactylogyrus spp. Knowl. Manag. Aquat. Ecosyst. 2008, 390–391, 6. [Google Scholar] [CrossRef]
- Martínez-Carrasco, C.; Serrano, E.; de Ybáñez, R.R.; Peñalver, J.; Antonio García, J.; García-Ayala, A.; Morand, S.; Muñoz, P. The European Eel—The Swim Bladder–Nematode System Provides a New View of the Invasion Paradox. Parasitol. Res. 2011, 108, 1501–1506. [Google Scholar] [CrossRef]
- Giari, L.; Ruehle, B.; Fano, E.A.; Castaldelli, G.; Poulin, R. Temporal Dynamics of Species Associations in the Parasite Community of European Eels, Anguilla anguilla, from a Coastal Lagoon. Int. J. Parasitol. Parasites Wildl. 2020, 12, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Panfili, J.; Ximénès, M.C. Évaluation de l’âge et de la croissance de l’anguille européenne (Anguilla anguilla L.) en milieu continental: Méthodologies, validation, application en Méditerranée et comparaisons en Europe. Bull. Fr. Pêche Piscic. 1994, 335, 43–66. [Google Scholar] [CrossRef]
- Daverat, F.; Tomas, J. Tactics and Demographic Attributes in the European Eel Anguilla anguilla in the Gironde Watershed, SW France. Mar. Ecol. Prog. Ser. 2006, 307, 247–257. [Google Scholar] [CrossRef]
- Daverat, F.; Beaulaton, L.; Poole, R.; Lambert, P.; Wickström, H.; Andersson, J.; Aprahamian, M.; Hizem, B.; Elie, P.; Yalçın-Özdilek, S.; et al. One Century of Eel Growth: Changes and Implications. Ecol. Freshw. Fish 2012, 21, 325–336. [Google Scholar] [CrossRef]
- Panfili, J.; Ximénès, M.-C.; Crivelli, A.J. Sources of Variation in Growth of the European Eel (Anguilla anguilla) Estimated from Otoliths. Can. J. Fish. Aquat. Sci. 1994, 51, 506–515. [Google Scholar] [CrossRef]
- Golovin, P.P. Monogeneans of Eel during Its Culture Using Heated Water. In Investigations of Monogenoidea in the USSR; USSR Academy of Sciences: Leningrad, Russia, 1977; pp. 144–150. [Google Scholar]
- Koops, H.; Hartmann, F. Anguillicola-Infestations in Germany and in German Eel Imports. J. Appl. Ichthyol. 1989, 5, 41–45. [Google Scholar] [CrossRef]
- Køie, M. Swimbladder Nematodes (Anguillicola spp.) and Gill Monogeneans (Pseudodactylogyrus spp.) Parasitic on the European Eel (Anguilla anguilla). ICES J. Mar. Sci. 1991, 47, 391–398. [Google Scholar] [CrossRef]
- van Banning, P.; Haenen, O.L.M. Effects of the Swimbladder Nematode Anguillicola crassus in Wild and Farmed Eel, Anguilla anguilla. In Pathology in Marine Science; Perkins, F.O., Cheng, T.C., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 317–330. ISBN 978-0-12-550755-4. [Google Scholar]
- Kennedy, C.R. The Pathogenic Helminth Parasites of Eels. J. Fish Dis. 2007, 30, 319–334. [Google Scholar] [CrossRef] [PubMed]
- Palstra, A.P.; Heppener, D.F.M.; van Ginneken, V.J.T.; Székely, C.; van den Thillart, G.E.E.J.M. Swimming Performance of Silver Eels Is Severely Impaired by the Swim-Bladder Parasite Anguillicola crassus. J. Exp. Mar. Biol. Ecol. 2007, 352, 244–256. [Google Scholar] [CrossRef]
- Buchmann, K. Pseudodactylogyrus anguillae and Pseudodactylogyrus bini. In Fish Parasites: Pathobiology and Protection; CABI: Wallingford, UK, 2012; pp. 209–224. [Google Scholar]
- Pergent-Martini, C.; Fernandez, C.; Agostini, S.; Pergent, G. Les Etangs de Corse, Bibliographie—Synthèse; Contrat Equipe Ecosystèmes Littoraux; Université de Corse/Office de l’Environnement de la Corse & IFREMER: Corte, France, 1997; p. 269. [Google Scholar]
- Pasqualini, V.; Derolez, V.; Garrido, M.; Orsoni, V.; Baldi, Y.; Etourneau, S.; Leoni, V.; Rébillout, P.; Laugier, T.; Souchu, P.; et al. Spatiotemporal Dynamics of Submerged Macrophyte Status and Watershed Exploitation in a Mediterranean Coastal Lagoon: Understanding Critical Factors in Ecosystem Degradation and Restoration. Ecol. Eng. 2017, 102, 1–14. [Google Scholar] [CrossRef]
- Ligorini, V.; Malet, N.; Garrido, M.; Derolez, V.; Amand, M.; Bec, B.; Cecchi, P.; Pasqualini, V. Phytoplankton Dynamics and Bloom Events in Oligotrophic Mediterranean Lagoons: Seasonal Patterns but Hazardous Trends. Hydrobiologia 2022, 849, 2353–2375. [Google Scholar] [CrossRef]
- Orsoni, V.; Souchu, P.; Sauzade, D. Caractérisation de L’état D’eutrophisation des Trois Principaux Etangs Corses (Biguglia, Diana et Urbino), et Proposition de Renforcement de Leur Surveillance. Rapport Final; Ifremer: Brest, France, 2001; p. 186. [Google Scholar]
- Durif, C.; Guibert, A.; Elie, P. Morphological Discrimination of the Silvering Stages of the European Eel. In Eels at the Edge: Science, Status, and Conservation Concerns; American Fisheries Society Symposium Publication: Bethesda, MD, USA, 2009. [Google Scholar]
- Panfili, J.; de Pontual, H.; Troadec, H.; Wright, P.J. Manual of Fish Sclerochronology; Ifremer-IRD coedition; Ifremer: Brest, France, 2002. [Google Scholar]
- ICES. Workshop on Age Reading of European and American Eel (WKAREA); ICES: Bordeaux, France, 2009; p. 66. [Google Scholar]
- Bush, A.O.; Lafferty, K.D.; Lotz, J.M.; Shostak, A.W. Parasitology Meets Ecology on Its Own Terms: Margolis et al. Revisited. J. Parasitol. 1997, 83, 575–583. [Google Scholar] [CrossRef]
- Vaissie, P.; Monge, A.; Husson, F. Factoshiny: Perform Factorial Analysis from “FactoMineR” with a Shiny Application; R Package Version 2.4.; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Mouillot, D.; Culioli, J.-M.; Chi, T.D. Indicator Species Analysis as a Test of Non-Random Distribution of Species in the Context of Marine Protected Areas. Environ. Conserv. 2002, 29, 385–390. [Google Scholar] [CrossRef]
- Dufrêne, M.; Legendre, P. Species Assemblages and Indicator Species: The Need for a Flexible Asymmetrical Approach. Ecol. Monogr. 1997, 67, 345–366. [Google Scholar] [CrossRef]
- Roberts, D.W. Labdsv: Ordination and Multivariate Analysis for Ecology; R Package Version 2.0-1; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Barker, D.E.; Marcogliese, D.J.; Cone, D.K. On the Distribution and Abundance of Eel Parasites in Nova Scotia: Local versus Regional Patterns. J. Parasitol. 1996, 82, 697–701. [Google Scholar] [CrossRef]
- Kristmundsson, Á.; Helgason, S. Parasite Communities of Eels Anguilla anguilla in Freshwater and Marine Habitats in Iceland in Comparison with Other Parasite Communities of Eels in Europe. Folia Parasitol. 2007, 54, 141. [Google Scholar] [CrossRef]
- Kennedy, C.R. Introductions, Spread and Colonization of New Localities by Fish Helminth and Crustacean Parasites in the British Isles: A Perspective and Appraisal. J. Fish Biol. 1993, 43, 287–301. [Google Scholar] [CrossRef]
- Conneely, J.J.; McCarthy, T.K. Ecological Factors Influencing the Composition of the Parasite Fauna of the European Eel, Anguilla anguilla (L.), in Ireland. J. Fish Biol. 1986, 28, 207–219. [Google Scholar] [CrossRef]
- Sures, B.; Knopf, K.; Würtz, J.; Hirt, J. Richness and Diversity of Parasite Communities in European Eels Anguilla anguilla of the River Rhine, Germany, with Special Reference to Helminth Parasites. Parasitology 1999, 119, 323–330. [Google Scholar] [CrossRef] [PubMed]
- Sures, B.; Streit, B. Eel Parasite Diversity and Intermediate Host Abundance in the River Rhine, Germany. Parasitology 2001, 123, 185–191. [Google Scholar] [CrossRef]
- Dezfuli, B.S.; Giari, L.; Castaldelli, G.; Lanzoni, M.; Rossi, R.; Lorenzoni, M.; Kennedy, C.R. Temporal and Spatial Changes in the Composition and Structure of Helminth Component Communities in European Eels Anguilla anguilla in an Adriatic Coastal Lagoon and Some Freshwaters in Italy. Parasitol. Res. 2014, 113, 113–120. [Google Scholar] [CrossRef]
- Di Cave, D.; Berrilli, F.; De Liberato, C.; Orecchia, P.; Kennedy, C.R. Helminth Communities in Eels Anguilla anguilla from Adriatic Coastal Lagoons in Italy. J. Helminthol. 2001, 75, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Hermida, M.; Saraiva, A.; Cruz, C. Metazoan Parasite Community of a European Eel (Anguilla anguilla) Population from an Estuary in Portugal. Bull. Eur. Assoc. Fish Pathol. 2008, 28, 35–40. [Google Scholar]
- Outeiral, S.; Alvarez, F.; Paniagua, E.; Leiro, L.; Sanmartín Durán, M. Digenean Parasites of the European Eel from Estuaries in North-West Spain. Helminthologia 2001, 38, 85–92. [Google Scholar]
- Jousson, O.; Bartoli, P.; Pawlowski, J. Molecular Identification of Developmental Stages in Opecoelidae (Digenea). Int. J. Parasitol. 1999, 29, 1853–1858. [Google Scholar] [CrossRef]
- Levron, C.; Ternengo, S.; Marchand, B. Ultrastructure of Spermiogenesis and the Spermatozoon of Helicometra fasciata (Digenea, Opecoelidae). Parasitol. Res. 2000, 86, 301–310. [Google Scholar] [CrossRef]
- Dezfuli, B.S.; Lui, A.; Squerzanti, S.; Lorenzoni, M.; Shinn, A.P. Confirmation of the Hosts Involved in the Life Cycle of an Acanthocephalan Parasite of Anguilla anguilla (L.) from Lake Piediluco and Its Effect on the Reproductive Potential of Its Amphipod Intermediate Host. Parasitol. Res. 2012, 110, 2137–2143. [Google Scholar] [CrossRef]
- Kennedy, C.R.; Berrilli, F.; Di Cave, D.; De Liberato, C.; Orecchia, P. Composition and Diversity of Helminth Communities in Eels Anguilla anguilla in the River Tiber: Long-Term Changes and Comparison with Insular Europe. J. Helminthol. 1998, 72, 301–306. [Google Scholar] [CrossRef] [PubMed]
- Norton, J.; Lewis, J.W.; Rollinson, D. Parasite Infracommunity Diversity in Eels: A Reflection of Local Component Community Diversity. Parasitology 2003, 127, 475–482. [Google Scholar] [CrossRef]
- Kennedy, C.R.; Moriarty, C. Long-Term Stability in the Richness and Structure of Helminth Communities in Eels, Anguilla anguilla, in Lough Derg, River Shannon, Ireland. J. Helminthol. 2002, 76, 315–322. [Google Scholar] [CrossRef]
- MacArthur, R.H. Patterns of Species Diversity. Biol. Rev. 1965, 40, 510–533. [Google Scholar] [CrossRef]
- Whittaker, R.J.; Fernández-Palacios, J.M. Island Biogeography: Ecology, Evolution, and Conservation, 2nd ed.; Oxford Biology; Oxford University Press: Oxford, UK; New York, NY, USA, 2007; ISBN 978-0-19-856612-0. [Google Scholar]
- Køie, M. Parasites in European Eel Anguilla anguilla (L.) from Danish Freshwater, Brackishand Marine Localities. Ophelia 1988, 29, 93–118. [Google Scholar] [CrossRef]
- Copley, L.; McCarthy, T.K. The First Record of the Monogenean Gill Fluke Pseudodactylogyrus bini (Kikuchi 1929) in Ireland, with Observations on Other Ectoparasites of River Erne Eels. Ir. Nat. J. 2001, 26, 405–413. [Google Scholar]
- Nabil, L.; Abderrafik, M.; Boudjema, S. Biodiversité Des Parasites Chez Anguilla anguilla Linnaeus, 1758 Dans Le Parc National d’El Kala-Algérie. Eur. J. Sci. Res. 2009, 25, 300–309. [Google Scholar]
- Tahri, M.; Bensaad-Bendjedid, L.; Dahel, A.; Djebbari, N.; Nouara, N.; Bensouilah, M. Site Specificity-Not Everything Is Everywhere-Case of Gill Ectoparasites of European Eel Anguilla anguilla (Linnaeus, 1758) (Park National of El Kala, Algeria). Cah. Biol. Mar. 2018, 59, 71–78. [Google Scholar] [CrossRef]
- Raibaut, A.; Altunel, F.N. Redescription de Ergasilus gibbus, Nordmann, 1832, Copépode Parasite Branchial de l’anguille et Remarques Sur Sa Répartition Géographique. Bull. Société Sci. Nat. Tunis. 1976, 11, 75–80. [Google Scholar]
- Kennedy, C.R.; Di Cave, D.; Berrilli, F.; Orecchia, P. Composition and Structure of Helminth Communities in Eels Anguilla anguilla from Italian Coastal Lagoons. J. Helminthol. 1997, 71, 35–40. [Google Scholar] [CrossRef]
- Maíllo, P.A.; Vich, M.A.; Salvadó, H.; Marqués, A.; Gracia, M.P. Parasites of Anguilla anguilla (L.) from Three Coastal Lagoons of the River Ebro Delta (Western Mediterranean). Acta Parasitol. 2005, 50, 156–160. [Google Scholar]
- Mayo-Hernández, E.; Peñalver, J.; García-Ayala, A.; Serrano, E.; Muñoz, P.; Ybáñez, R.R. de Richness and Diversity of Helminth Species in Eels from a Hypersaline Coastal Lagoon, Mar Menor, South-East Spain. J. Helminthol. 2015, 89, 345–351. [Google Scholar] [CrossRef] [PubMed]
- Gargouri-Ben Abdallah, L.; Maamouri, F. Life cycle of Bucephalus anguillae Spakulova, Macko, Berrilli & Dezfulli, 2002 (Digenea, Buccephalidae), a parasite of Anguilla anguilla (L.). Syst. Parasitol. 2002, 53, 207–217. [Google Scholar]
- Roché, B. Atlas des Poissons D’eau de Douce de Corse; Diren de Corse: Bastia, France, 2001. [Google Scholar]
- Cable, R.M.; Hunninen, A.V. Studies on Deropristis inflata (Molin), Its Life History and Affinities to Trematodes of the Family Acanthocolpidae. Biol. Bull. 1942, 82, 292–312. [Google Scholar] [CrossRef]
- Gibson, D.I.; Bray, R.A. The Hemiuridae (Digenea) of Fishes from the North-East Atlantic. Bull. Br. Mus. Nat. Hist. 1986, 51, 1–125. [Google Scholar] [CrossRef]
- Dezfuli, B.S.; Volponi, S.; Beltrami, I.; Poulin, R. Intra-and Interspecific Density-Dependent Effects on Growth in Helminth Parasites of the Cormorant, Phalacrocorax carbo sinensis. Parasitology 2002, 124, 537–544. [Google Scholar] [CrossRef]
- García-Varela, M.; Aznar, F.J.; Rodríguez, R.P.; de León, G.P.-P. Genetic and Morphological Characterization of Southwellina hispida Van Cleave, 1925 (Acanthocephala: Polymorphidae), a Parasite of Fish-Eating Birds. Comp. Parasitol. 2012, 79, 192–201. [Google Scholar] [CrossRef]
- Moravec, F. Parasitic Nematodes of Freshwater Fishes of Europe, 1st ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1994; ISBN 978-0-7923-2172-9. [Google Scholar]
- Lefebvre, F.; Contournet, P.; Priour, F.; Soulas, O.; Crivelli, A.J. Spatial and Temporal Variation in Anguillicola crassus Counts: Results of a 4 Year Survey of Eels in Mediterranean Lagoons. Dis. Aquat. Org. 2002, 50, 181–188. [Google Scholar] [CrossRef] [PubMed]
- McGladdery, S.E. Anisakis simplex (Nematoda: Anisakidae) Infection of the Musculature and Body Cavity of Atlantic Herring (Clupea harengus harengus). Can. J. Fish. Aquat. Sci. 1986, 43, 1312–1317. [Google Scholar] [CrossRef]
- Shamsi, S.; Suthar, J. A revised method of examining fish for infection with zoonotic nematode larvae. Int. J. Food Microbiol. 2016, 227, 13–16. [Google Scholar] [CrossRef]
- Shamsi, S.; Turner, A.; Wassens, S. Description and genetic characterization of a new Contracaecum larval type (Nematoda: Anisakidae) from Australia. J. Helminthol. 2017, 92, 216–222. [Google Scholar] [CrossRef] [PubMed]
- Norton, J.; Rollinson, D.; Lewis, J.W. Patterns of Infracommunity Species Richness in Eels, Anguilla anguilla. J. Helminthol. 2004, 78, 141–146. [Google Scholar] [CrossRef]
- Saraiva, A.; Antao, A.; Cruz, C. Comparative Study of Parasite Communities in European Eel Anguilla anguilla from Rivers of Northern Portugal. Helminthologia 2005, 42, 99. [Google Scholar]
- Shears, J.A.; Kennedy, C.R. The Life Cycle of Paraquimperia tenerrima: A Parasite of the European Eel Anguilla anguilla. J. Helminthol. 2005, 79, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Buchmann, K.; Mellergaard, S.; Køie, M. Pseudodactylogyrus Infections in Eel—A Review. Dis. Aquat. Org. 1987, 3, 51–57. [Google Scholar] [CrossRef]
- Kolar, C.S.; Lodge, D.M. Progress in Invasion Biology: Predicting Invaders. Trends Ecol. Evol. 2001, 16, 199–204. [Google Scholar] [CrossRef]
- Egusa, S. Notes on the Culture of the European Eel (Anguilla anguilla L.) in Japanese Eel-Farming Ponds. Rapp. Proc.-Verbaux Reun. Cons. Int. L’explor. Mer 1979, 174, 51–58. [Google Scholar]
- Lambert, A.; Le Brun, N.; Pariselle, A. Occurrence in France of Pseudodactylogyrus anguilla (Yin et Sproston, 1948) Gussev, 1965 (Monogenea, Monopisthocotylea), a gill parasite of Anguilla anguilla in fresh water. Ann. Parasitol. Hum. Comp. 1985, 60, 91–92. [Google Scholar]
- Hayward, C.J.; Iwashita, M.; Crane, J.S.; Ogawa, K. First Report of the Invasive Eel Pest Pseudodactylogyrus bini in North America and in Wild American Eels. Dis. Aquat. Org. 2001, 44, 53–60. [Google Scholar] [CrossRef]
- Truter, M.; Hadfield, K.A.; Weyl, O.L.F.; Smit, N.J. Pseudodactylogyrus anguillae (Yin & Sproston, 1948) from the Giant Mottled Eel Anguilla marmorata Quoy & Gaimard, 1824, in the Phongolo River, South Africa: An Invader on the African Continent. Parasitol. Res. 2021, 120, 1247–1268. [Google Scholar] [CrossRef]
- Kennedy, C.R.; Nie, P.; Kaspers, J.; Paulisse, J. Are Eels (Anguilla anguilla L.) Planktonic Feeders? Evidence from Parasite Communities. J. Fish Biol. 1992, 41, 567–580. [Google Scholar] [CrossRef]
- Skorikova, B.; Scholz, T.; Moravec, F. Spreading of Introduced Monogeneans Pseudodactylogyrus anguillae and P. bini among Eel Populations in the Czech Republic. Folia Parasitol. 1996, 43, 155–156. [Google Scholar]
- Caillot, C.; Morand, S.; Muller-Graf, C.M.; Faliex, E.; Marchand, B. Parasites of Dicentrarchus labrax, Anguilla anguilla, and Mugil cephalus from a Pond in Corsica, France. J. Helminthol. Soc. Wash. 1999, 66, 95–98. [Google Scholar]
- Buchmann, K. Microhabitats of Monogenean Gill Parasites on European Eel (Anguilla anguilla). Folia Parasitol. 1989, 36, 321–329. [Google Scholar]
- Nie, P.; Kennedy, C.R. Occurrence and Seasonal Dynamics of Pseudodactylogyrus anguillae (Yin & Sproston) (Monogenea) in Eel, Anguilla anguilla (L.), in England. J. Fish Biol. 1991, 39, 897–900. [Google Scholar] [CrossRef]
- Matejusová, I.; Simková, A.; Sasal, P.; Gelnar, M. Microhabitat Distribution of Pseudodactylogyrus anguillae and Pseudodactylogyrus bini among and within Gill Arches of the European Eel (Anguilla anguilla L.). Parasitol. Res. 2003, 89, 290–296. [Google Scholar] [CrossRef]
- Koie, M. Parasites in Eels, Anguilla (L.), from Eutrophic Lake Esrum (Denmark). Acta Parasitol. Pol. 1988, 33, 89–100. [Google Scholar]
- Moser, M.L.; Patrick, W.S.; Crutchfield, J.U., Jr. Infection of American Eels, Anguilla rostrata, by an Introduced Nematode Parasite, Anguillicola crassus, in North Carolina. Copeia 2001, 2001, 848–853. [Google Scholar] [CrossRef]
- Schneebauer, G.; Hanel, R.; Pelster, B. Anguillicola crassus Impairs the Silvering-Related Enhancements of the ROS Defense Capacity in Swimbladder Tissue of the European Eel (Anguilla anguilla). J. Comp. Physiol. B 2016, 186, 867–877. [Google Scholar] [CrossRef]
- Currie, H.A.L.; Martin, N.F.; Garcia, G.E.; Davis, F.M.; Kemp, P.S. A Mechanical Approach to Understanding the Impact of the Nematode Anguillicoloides crassus on the European Eel Swimbladder. J. Exp. Biol. 2020, 223, jeb219808. [Google Scholar] [CrossRef]
- Muñoz, P.; Peñalver, J.; Ruiz de Ybañez, R.; Garcia, J. Influence of Adult Anguillicoloides crassus Load in European Eels Swimbladder on Macrophage Response. Fish Shellfish Immunol. 2015, 42, 221–224. [Google Scholar] [CrossRef]
- Gollock, M.J.; Kennedy, C.R.; Brown, J.A. European Eels, Anguilla anguilla (L.), Infected with Anguillicola crassus Exhibit a More Pronounced Stress Response to Severe Hypoxia than Uninfected Eels. J. Fish Biol. 2005, 28, 429–436. [Google Scholar] [CrossRef]
- Kirk, R.S. The Impact of Anguillicola crassus on European Eels. Fish. Manag. Ecol. 2003, 10, 385–394. [Google Scholar] [CrossRef]
- Moravec, F.; Nagasawa, K.; Miyakawa, M. First Record of Ostracods as Natural Intermediate Hosts of Anguillicola crassus, a Pathogenic Swimbladder Parasite of Eels Anguilla spp. Dis. Aquat. Org. 2005, 66, 171–173. [Google Scholar] [CrossRef] [PubMed]
- Tahri, M. Étude comparative de l’infestation de l’anguille européenne Anguilla anguilla par Anguillicoloïdes crassus dans deux biotopes de salinité différente. Hydroécol. Appl. 2023, 23, 1. [Google Scholar] [CrossRef]
- Nielsen, M.E. Infection Status of the Swimbladder Worm, Anguillicola crassus in Silver Stage European Eel, Anguilla anguilla, from Three Different Habitats in Danish Waters. J. Appl. Ichthyol. 1997, 13, 195–196. [Google Scholar] [CrossRef]
- Cakic, P.; Stojanovski, S.; Kulišić, Z.; Hristovski, N.; Lenhardt, M. Occurrence of Anguillicola crassus (Nematoda: Dracunculoidea) in Eels of Lake Ohrid, Macedonia. Acta Vet. 2002, 52, 163–168. [Google Scholar] [CrossRef]
- Schabuss, M.; Konecny, R.; Belpaire, C.; Schiemer, F. Endoparasitic Helminths of the European Eel, Anguilla anguilla, from Four Disconnected Meanders from the Rivers Leie and Scheldt in Western Flanders, Belgium. Folia Parasitol. 1997, 44, 12–18. [Google Scholar]
- Audenaert, V.; Huyse, T.; Goemans, G.; Belpaire, C.; Volckaert, F.a.M. Spatio-Temporal Dynamics of the Parasitic Nematode Anguillicola crassus in Flanders, Belgium. Dis. Aquat. Org. 2003, 56, 223–233. [Google Scholar] [CrossRef]
- Schabuss, M.; Kennedy, C.R.; Konecny, R.; Grillitsch, B.; Schiemer, F.; Herzig, A. Long-Term Investigation of the Composition and Richness of Intestinal Helminth Communities in the Stocked Population of Eel, Anguilla anguilla, in Neusiedler See, Austria. Parasitology 2005, 130, 185–194. [Google Scholar] [CrossRef]
- Hoglund, J.; Andersson, J. Prevalence and Abundance of Anguillicola crassus in the European Eel (Anguilla anguilla) at a Thermal Discharge Site on the Swedish Coast. J. Appl. Ichthyol. 1993, 9, 115–122. [Google Scholar] [CrossRef]
- Gargouri Ben Abdallah, L.; Maamouri, F. Spatio-Temporal Dynamics of the Nematode Anguillicola crassus in Northeast Tunisian Lagoons. Comptes Rendus Biol. 2006, 329, 785–789. [Google Scholar] [CrossRef]
- Habbechi, B.H.; Kraiem, M.M.; Élie, P. Study of European eel (Anguilla anguilla L., 1758) contamination by Anguillicoloides crassus in some Tunisian hydrosystems: Its impacts on growth parameters. Cybium 2012, 36, 417–433. [Google Scholar]
- Wielgoss, S.; Taraschewski, H.; Meyer, A.; Wirth, T. Population Structure of the Parasitic Nematode Anguillicola crassus, an Invader of Declining North Atlantic Eel Stocks. Mol. Ecol. 2008, 17, 3478–3495. [Google Scholar] [CrossRef] [PubMed]
- Benajiba, M.H.; Silan, P.; Marquès, A.; Bouix, G. Protozoaires et Métazoaires Parasites de l’anguille Anguilla anguilla, L. 1758: Structures Temporelles de Leurs Populations Dans Une Lagune Méditerranéenne. Ann. Sci. Nat. Zool. Biol. Anim. 1994, 15, 141–149. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| N | Total Length (mm) | Total Weight (g) | Age (a) | Fulton Condition Index | N I | N FII | N FIII | N FV | N MII | ||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Lagoon | Season | ||||||||||
| Biguglia | Spring 2021 | 20 | 386 ± 54 (313–525) | 117 ± 63 (54–315) | 6 ± 1 (4–7) | 0.19 ± 0.02 (0.15–0.24) | 18 | 2 | 0 | 0 | 0 |
| Summer 2021 | 20 | 409 ± 44 (355–517) | 126 ± 49 (80–287) | 6 ± 1 (5–7) | 0.18 ± 0.03 (0.09–0.21) | 14 | 3 | 0 | 0 | 3 | |
| Fall 2021 | 20 | 382 ± 29 (305–425) | 98 ± 26 (35–150) | 6 ± 1 (4–7) | 0.17 ± 0.02 (0.12–0.21) | 4 | 0 | 0 | 0 | 16 | |
| Winter 2021–2022 | 20 | 380 ± 23 (334–449) | 92 ± 18 (61–149) | 6 ± 1 (5–8) | 0.17 ± 0.01 (0.15–0.18) | 3 | 1 | 0 | 0 | 16 | |
| Urbino | Spring 2021 | 20 | 421 ± 41 (341–510) | 105 ± 29 (58–184) | 8 ± 2 (6–13) | 0.14 ± 0.05 (0.09–0.34) | 8 | 9 | 0 | 1 | 2 |
| Summer 2021 | 20 | 466 ± 121 (327–771) | 178 ± 159 (47–661) | 9 ± 3 (6–17) | 0.15 ± 0.02 (0.12–0.21) | 11 | 5 | 4 | 0 | 0 | |
| Fall 2021 | 20 | 432 ± 64 (344–597) | 149 ± 75 (59–379) | 8 ± 2 (6–14) | 0.17 ± 0.02 (0.14–0.22) | 10 | 7 | 0 | 0 | 3 | |
| Winter 2021–2022 | 20 | 379 ± 61 (307–580) | 94 ± 60 (40–319) | 8 ± 2 (5–14) | 0.16 ± 0.01 (0.13–0.19) | 15 | 1 | 0 | 1 | 3 | |
| River | |||||||||||
| Abatesco | 15 | 368 ± 83 (278–541) | 123 ± 86 (41–295) | 11.1 ± 2.8 (7–16) | 0.21 ± 0.03 (0.18–0.27) | 12 | 3 | 0 | 0 | 0 | |
| Fium’Orbo | 15 | 320 ± 77 (256–582) | 79 ± 86 (25–377) | 8.3 ± 2.7 (5–17) | 0.18 ± 0.03 (0.15–0.26) | 12 | 0 | 1 | 0 | 2 | |
| Golo | 11 | 3717 ± 87 (234–504) | 126 ± 81 (48–280) | 9.3 ± 3.5 (5–15) | 0.24 ± 0.12 (0.17–0.58) | 7 | 2 | 1 | 0 | 1 | |
| Liamone | 15 | 332 ± 47 (282–479) | 89 ± 54 (45–273) | 8.3 ± 1.6 (6–13) | 0.22 ± 0.03 (0.19–0.26) | 14 | 0 | 0 | 0 | 1 | |
| Prunelli | 15 | 370 ± 62 (304–493) | 90 ± 47 (46–194) | 8.5 ± 1.8 (6–12) | 0.17 ± 0.02 (0.13–0.2) | 11 | 2 | 1 | 0 | 1 | |
| Rizzanese | 15 | 309 ± 31 (266–363) | 53 ± 19 (31–88) | 6.5 ± 1.1 (5–9) | 0.17 ± 0.02 (0.14–0.23) | 15 | 0 | 0 | 0 | 0 | |
| San Clemente | 15 | 305 ± 17 (280–340) | 60 ± 13 (41–80) | 6.9 ± 1.0 (5–8) | 0.21 ± 0.04 (0.16–0.29) | 13 | 0 | 0 | 0 | 2 | |
| Taravo | 15 | 295 ± 78 (181–406) | 60 ± 46 (8–154) | 7.2 ± 2.5 (3–12) | 0.18 ± 0.03 (0.13–0.23) | 15 | 0 | 0 | 0 | 0 | |
| Tartagine | 14 | 386 ± 71 (266–512) | 117 ± 86 (29–321) | 7.4 ± 2.2 (3–11) | 0.18 ± 0.03 (0.15–0.24) | 11 | 1 | 1 | 0 | 1 | |
| Tavignano | 15 | 335 ± 59 (259–502) | 74 ± 47 (26–217) | 7.5 ± 1.6 (5–11) | 0.18 ± 0.02 (0.15–0.21) | 13 | 1 | 0 | 0 | 1 | |
| Travo | 15 | 309 ± 28 (259–367) | 59 ± 17 (28–93) | 9.1 ± 1.4 (7–12) | 0.20 ± 0.03 (0.16–0.24) | 11 | 0 | 0 | 0 | 4 |
| Biguglia | ||||||||||||
| Spring 2021 | Summer 2021 | Fall 2021 | Winter 2021–2022 | |||||||||
| N | 20 | 20 | 20 | 20 | ||||||||
| Shannon index | 0.71 ± 0.27 (0.32–1.47) | 0.6 ± 0.23 (0.06–1.17) | 0.47 ± 0.27 (0.05–1.17) | 0.47 ± 0.27 (0.03–0.93) | ||||||||
| Berger–Parker dominance index | 0.7 ± 0.15 (0.41–0.92) | 0.72 ± 0.14 (0.43–0.99) | 0.83 ± 0.13 (0.49–0.99) | 0.81 ± 0.15 (0.53–1) | ||||||||
| Prevalence | Mean abundance | Mean intensity | Prevalence | Mean abundance | Mean intensity | Prevalence | Mean abundance | Mean intensity | Prevalence | Mean abundance | Mean intensity | |
| Copepoda | ||||||||||||
| Ergasilus gibbus | 100 | 11.15 ± 6.75 | 11.15 ± 6.75 | 100 | 38.5 ± 32.13 | 38.5 ± 32.13 | 75 | 12.7 ± 15.2 | 16.93 ± 15.39 | 85 | 38.7 ± 49.86 | 33.76 ± 52.63 |
| Cestoda | ||||||||||||
| Bothriocephalus claviceps | 0 | - | - | 0 | - | - | 0 | - | - | 5 | 0.1 ± 0.45 | 2 ± - |
| Proteocephalus macrocephalus | 10 | 0.7 ± 2.7 | 7 ± 7.07 | 0 | - | - | 0 | - | - | 0 | - | - |
| Larvae indet. | 0 | - | - | 0 | - | - | 0 | - | - | 0 | - | - |
| Digenea | ||||||||||||
| Bucephalus anguillae | 0 | - | - | 0 | - | - | 0 | - | - | 0 | - | - |
| Deropristis inflata | 0 | - | - | 0 | - | - | 5 | 0.3 ± 1.34 | 6 ± - | 15 | 0.15 ± 0.37 | 1 ± 0 |
| Lecithochirium musculus | 0 | - | - | 0 | - | - | 0 | - | - | 0 | - | - |
| Monogenea | ||||||||||||
| Pseudodactylogyrus anguillae + P. bini | 100 | 35.85 ± 24.36 | 35.85 ± 24.36 | 100 | 126.65 ± 133.75 | 126.65 ± 133.75 | 100 | 161.5 ± 273.96 | 161.5 ± 273.96 | 100 | 131.25 ± 141.62 | 131.25 ± 141.62 |
| Acanthocephala | ||||||||||||
| Solearhynchus rhytidotes | 0 | - | - | 0 | - | - | 0 | - | - | 0 | - | - |
| Southwellina hispida (cystacanth) | 5 | 0.05 ± 0.22 | 1 ± - | 0 | - | - | 5 | 0.05 ± 0.22 | 1 ± - | 0 | - | - |
| Telosentis exiguus | 0 | - | - | 0 | - | - | 0 | - | - | 0 | - | - |
| Acanthocephala indet. | 0 | - | - | 0 | - | - | 0 | - | - | 0 | - | - |
| Nematoda | ||||||||||||
| Anguillicola crassus | 75 | 5.35 ± 9.52 | 7.13 ± 10.46 | 50 | 1.25 ± 1.94 | 2.5 ± 2.12 | 95 | 3.6 ± 2.72 | 3.79 ± 2.66 | 60 | 2.55 ± 2.12 | 4.25 ± 2.99 |
| Contracaecum sp. (encysted larvae) | 5 | 1.3 ± 5.81 | 26 ± - | 15 | 4.8 ± 14.63 | 32 ± 26.96 | 35 | 3.74 ± 10.83 | 11.83 ± 17.53 | 25 | 0.74 ± 1.79 | 3.5 ± 2.2 |
| Urbino | ||||||||||||
| Spring 2021 | Summer 2021 | Fall 2021 | Winter 2021–2022 | |||||||||
| N | 20 | 20 | 20 | 20 | ||||||||
| Shannon index | 0.26 ± 0.26 (0–0.69) | 0.43 ± 0.3 (0–0.86) | 0.62 ± 0.43 (0–1.78) | 0.55 ± 0.42 (0–1.18) | ||||||||
| Berger–Parker dominance index | 0.88 ± 0.15 (0.5–1) | 0.8 ± 0.16 (0.52–1) | 0.72 ± 0.21 (0.29–1) | 0.75 ± 0.2 (0.47–1) | ||||||||
| Prevalence | Mean abundance | Mean intensity | Prevalence | Mean abundance | Mean intensity | Prevalence | Mean abundance | Mean intensity | Prevalence | Mean abundance | Mean intensity | |
| Copepoda | ||||||||||||
| Ergasilus gibbus | 0 | - | - | 0 | - | - | 0 | - | - | 0 | - | - |
| Cestoda | ||||||||||||
| Bothriocephalus claviceps | 0 | - | - | 0 | - | - | 0 | - | - | 0 | - | - |
| Proteocephalus macrocephalus | 0 | - | - | 0 | - | - | 0 | - | - | 0 | - | - |
| Larvae indet. | 0 | - | - | 0 | - | - | 10 | 0.1 ± 0.31 | 1 ± 0 | 5 | 0.05 ± 0.22 | 1 ± - |
| Digenea | ||||||||||||
| Bucephalus anguillae | 55 | 14.25 ± 44.4 | 25.91 ± 58.43 | 50 | 23.05 ± 48.82 | 46.1 ± 62.06 | 45 | 6.25 ± 19.47 | 13.89 ± 27.95 | 44 | 4.19 ± 7.03 | 9.57 ± 7.96 |
| Deropristis inflata | 25 | 1.35 ± 4.25 | 5.4 ± 7.64 | 0 | - | - | 55 | 6.65 ± 17.85 | 12.09 ± 23.09 | 78 | 34.83 ± 88.41 | 44.79 ± 98.7 |
| Lecithochirium musculus | 15 | 0.15 ± 0.37 | 1 ± 0 | 35 | 4.05 ± 9.71 | 11.57 ± 14.05 | 50 | 10.5 ± 15.26 | 21 ± 15.71 | 68 | 3.94 ± 6.68 | 5.77 ± 7.45 |
| Monogenea | ||||||||||||
| Pseudodactylogyrus anguillae + P. bini | 0 | - | - | 0 | - | - | 0 | - | - | 0 | - | - |
| Acanthocephala | ||||||||||||
| Solearhynchus rhytidotes | 0 | - | - | 0 | - | - | 10 | 0.1 ± 0.31 | 1 ± 0 | 0 | - | - |
| Southwellina hispida (cystacanth) | 11 | 0.32 ± 1.16 | 3 ± 2.83 | 55 | 2.4 ± 4.51 | 4.36 ± 5.41 | 55 | 3.15 ± 5.21 | 5.73 ± 5.95 | 0 | - | - |
| Telosentis exiguus | 0 | - | - | 0 | - | - | 0 | - | - | 5 | 0.1 ± 0.45 | 2 ± - |
| Acanthocephala indet. | 0 | - | - | 5 | 0.05 ± 0.22 | 1 ± - | 15 | 0.35 ± 1.14 | 2.33 ± 2.31 | 0 | - | - |
| Nematoda | ||||||||||||
| Anguillicola crassus | 10 | 0.2 ± 0.7 | 2 ± 1.41 | 15 | 0.3 ± 0.8 | 2 ± 1 | 0 | - | - | 15 | 0.15 ± 0.37 | 1 ± 0 |
| Contracaecum sp. (encysted larvae) | 68 | 33.11 ± 52.44 | 48.38 ± 57.65 | 70 | 53.15 ± 137.43 | 75.93 ± 160.44 | 70 | 36.85 ± 68.12 | 52.64 ± 76.72 | 55 | 6.68 ± 12.15 | 12.7 ± 14.5 |
| Abatesco | Fium’Orbo | Golo | Liamone | |||||||||
| N | 15 | 15 | 11 | 15 | ||||||||
| Shannon index | 0.53 ± 0.39 (0–1.33) | 0.16 ± 0.25 (0–0.69) | 0.14 ± 0.22 (0–0.64) | 0.55 ± 0.38 (0–1.01) | ||||||||
| Berger–Parker dominance index | 0.7 ± 0.19 (0.33–1) | 0.91 ± 0.17 (0.5–1) | 0.94 ± 0.11 (0.67–1) | 0.76 ± 0.18 (0.5–1) | ||||||||
| Prevalence | Mean abundance | Mean intensity | Prevalence | Mean abundance | Mean intensity | Prevalence | Mean abundance | Mean intensity | Prevalence | Mean abundance | Mean intensity | |
| Copepoda | ||||||||||||
| Ergasilus gibbus | 6.67 | 0.27 ± 1.03 | 4 ± - | 0 | - | - | 0 | - | - | 0 | - | - |
| Cestoda | ||||||||||||
| Bothriocephalus claviceps | 0 | - | - | 0 | - | - | 0 | - | - | 13.33 | 0.13 ± 0.35 | 1 ± 0 |
| Digenea | ||||||||||||
| Unidentified Digenea | 0 | - | - | 0 | - | - | 0 | - | - | 0 | - | - |
| Monogenea | ||||||||||||
| Pseudodactylogyrus anguillae + P. bini | 66.67 | 4.13 ± 4.41 | 6.2 ± 5. 59 | 60 | 7.93 ± 17.58 | 13.22 ± 21.5 | 81.81 | 10 ± 13.51 | 12.22 ± 14.05 | 86.67 | 6.8 ± 10.51 | 7.85 ± 10.95 |
| Nematoda | ||||||||||||
| Anguillicola crassus | 53.33 | 2.67 ± 3.96 | 5 ± 4.24 | 53.33 | 1.33 ± 1.76 | 2.5 ± 1.69 | 36.36 | 1.18 ± 2.44 | 3.25 ± 3.3 | 53.33 | 1 ± 1.56 | 1.88 ± 1.73 |
| Contracaecum sp. (encysted larvae) | 0 | - | - | 6.67 | 0.47 ± 1.81 | 7 ± - | 7 | - | - | 6.67 | 0.33 ± 1.29 | 5 ± - |
| Paraquimperia tenerrima | 60 | 3.27 ± 4.03 | 5.44 ± 3.88 | 0 | - | - | 0 | - | - | 66.67 | 1.33 ± 1.8 | 2 ± 1.88 |
| Spinitectus inermis | 0 | - | - | 0 | - | - | 9.09 | 0.09 ± 0.3 | 1 ± - | 0 | - | - |
| Pseudocapillaria tomentosa | 0 | - | - | 0 | - | - | 0 | - | - | 0 | - | - |
| Prunelli | Rizzanese | San Clemente | Taravo | |||||||||
| N | 15 | 15 | 15 | 15 | ||||||||
| Shannon index | 0.19 ± 0.28 (0–0.69) | 0.37 ± 0.26 (0–0.69) | 0.34 ± 0.33 (0–0.74) | 0.48 ± 0.42 (0–1.1) | ||||||||
| Berger–Parker dominance index | 0.89 ± 0.17 (0.5–1) | 0.82 ± 0.18 (0.5–1) | 0.84 ± 0.17 (0.5–1) | 0.74 ± 0.24 (0.33–1) | ||||||||
| Prevalence | Mean abundance | Mean intensity | Prevalence | Mean abundance | Mean intensity | Prevalence | Mean abundance | Mean intensity | Prevalence | Mean abundance | Mean intensity | |
| Copepoda | ||||||||||||
| Ergasilus gibbus | 0 | - | - | 0 | - | - | 0 | - | - | 0 | - | - |
| Cestoda | ||||||||||||
| Bothriocephalus claviceps | 0 | - | - | 0 | - | - | 0 | - | - | 6.67 | 0.07 ± 0.26 | 1 ± - |
| Digenea | ||||||||||||
| Unidentified Digenea | 0 | - | - | 0 | - | - | 0 | - | - | 33.33 | 2 ± 6.39 | 6 ± 10.63 |
| Monogenea | ||||||||||||
| Pseudodactylogyrus anguillae + P. bini | 80 | 14.47 ± 36.87 | 18.08 ± 40.73 | 93.33 | 9.73 ± 10.54 | 10.43 ± 10.57 | 80 | 4.47 ± 4.55 | 5.58 ± 4.42 | 73.33 | 12.87 ± 19.26 | 17.55 ± 20.72 |
| Nematoda | ||||||||||||
| Anguillicola crassus | 40 | 0.93 ± 1.62 | 2.33 ± 1.86 | 66.67 | 1.73 ± 2.09 | 2.6 ± 2.07 | 53.33 | 1.8 ± 2.31 | 3.36 ± 2.13 | 26.67 | 0.6 ± 1.24 | 2.25 ± 1.5 |
| Contracaecum sp. (encysted larvae) | 0 | - | - | 0 | - | - | 0 | - | - | 6.67 | 0.07 ± 0.26 | 1 ± - |
| Paraquimperia tenerrima | 0 | - | - | 26.67 | 0.4 ± 0.74 | 1.5 ± 0.58 | 33.33 | 1 ± 1.92 | 3 ± 2.35 | 53.33 | 0.87 ± 1.19 | 1.63 ± 1.19 |
| Spinitectus inermis | 0 | - | - | 0 | - | - | 0 | - | - | 0 | - | - |
| Pseudocapillaria tomentosa | 6.67 | 0.27 ± 1.03 | 4 ± - | 0 | - | - | 0 | - | - | 0 | - | - |
| Tartagine | Tavignano | Travo | ||||||||||
| N | 14 | 15 | 15 | |||||||||
| Shannon index | 0 ± 0 (0–0) | 0.13 ± 0.24 (0–0.68) | 0.18 ± 0.32 (0–0.9) | |||||||||
| Berger–Parker dominance index | 1 ± 0 (1–1) | 0.94 ± 0.1 (0.76–1) | 0.87 ± 0.16 (0.62–1) | |||||||||
| Prevalence | Mean abundance | Mean intensity | Prevalence | Mean abundance | Mean intensity | Prevalence | Mean abundance | Mean intensity | ||||
| Copepoda | ||||||||||||
| Ergasilus gibbus | 0 | - | - | 0 | - | - | 0 | - | - | |||
| Cestoda | ||||||||||||
| Bothriocephalus claviceps | 0 | - | - | 0 | - | - | 0 | - | - | |||
| Digenea | ||||||||||||
| Unidentified Digenea | 0 | - | - | 0 | - | - | 0 | - | - | |||
| Monogenea | ||||||||||||
| Pseudodactylogyrus anguillae + P. bini | 35.71 | 29.57 ± 83.78 | 82.8 ± 131.53 | 80 | 11 ± 17.04 | 13.75 ± 18.12 | 40 | 2.6 ± 5.83 | 6.5 ± 8.04 | |||
| Nematoda | ||||||||||||
| Anguillicola crassus | 7.14 | 0.07 ± 0.27 | 1 ± - | 26.67 | 1.2 ± 3.34 | 4.5 ± 5.69 | 33.33 | 1.13 ± 1.77 | 3.4 ± 1.14 | |||
| Contracaecum sp. (encysted larvae) | 0 | - | - | 6.67 | 0.07 ± 0.26 | 1 ± - | 0 | - | - | |||
| Paraquimperia tenerrima | 0 | - | - | 0 | - | - | 20 | 0.93 ± 2.34 | 4.67 ± 3.51 | |||
| Spinitectus inermis | 0 | - | - | 0 | - | - | 0 | - | - | |||
| Pseudocapillaria tomentosa | 0 | - | - | 0 | - | - | 0 | - | - | |||
| Habitat | Biguglia Lagoon | Urbino Lagoon | Rivers |
|---|---|---|---|
| Number of eels | 78 | 72 | 159 |
| Monogenea | |||
| Pseudodactylogyrus anguillae + P. bini | 91.9–100.0.0 (91.9) **** | 0.0–0.0 (0.0) | 8.1–71.1 (5.8) |
| Digenea | |||
| Bucephalus anguillae | 0.0–0.0 (0.0) | 100.0.0–47.2 (47.2) **** | 0.0–0.0 (0.0) |
| Deropristis inflata | 1.1–5.1 (0.1) | 98.9–36.1 (35.7) **** | 0.0–0.0 (0.0) |
| Lecithochirium musculus | 0.0–0.0 (0.0) | 100.0.0–38.9 (38.9) **** | 0.0–0.0 (0.0) |
| Copepoda | |||
| Ergasilus gibbus | 99.9–89.7 (89.6) **** | 0.0–0.0 (0.0) | 0.1–0.6 (0.0) |
| Acanthocephala | |||
| Southwellina hispida | 1.6–2.5 (0.0) | 98.4–31.3 (30.7) **** | 0.0–0.0 (0.0) |
| Acanthocephala indet. | 0.0–0.0 (0.0) | 100.0.0–5.5 (5.5) ** | 0.0–0.0 (0.0) |
| Nematoda | |||
| Anguillicola crassus | 68.7–69.2 (47.6) **** | 3.7–9.7 (0.4) | 27.6–41.5 (11.5) |
| Contracaecum sp. | 7.0–17.9 (1.3) | 92.7–66.7 (61.8) **** | 0.2–2.5 (0.0) |
| Paraquimperia tenerrima | 0.0–0.0 (0.0) | 0.0-–0.0 (0.0) | 100.0.0–24.5 (24.5) **** |
| Biguglia | ||||
| Season | Spring | Summer | Fall | Winter |
| Number of eels | 20 | 20 | 19 | 19 |
| Copepoda | ||||
| Ergasilus gibbus | 12.0–100.0 (12.0) | 41.4–100.0 (41.4) * | 14.2–73.7 (10.5) | 32.4–84.2 (27.3) |
| Urbino | ||||
| Season | Spring | Summer | Fall | Winter |
| Number of eels | 18 | 20 | 20 | 14 |
| Digenea | ||||
| Deropristis inflata | 0.9–22.2 (0.2) | 0.0–0.0 (0.0) | 13.7–55.0 (7.5) | 85.4–78.6 (67.1) **** |
| Acanthocephala | ||||
| Southwellina hispida | 5.7–11.1 (0.6) | 40.8–55.0 (22.4) | 53.5–55.0 (29.4) * | 0.0–0.0 (0.0) |
| Biguglia | |||||
| Silvering Stage | I | FII | FIII | FV | MII |
| Number of eels | 38 | 6 | 34 | ||
| Cestoda | |||||
| Proteocephalus macrocephalus | 2.6–2.6 (0.0) | 97.4–16.7 (16.2) * | - | - | 0.0–0.0 (0.0) |
| Urbino | |||||
| Silvering stage | I | FII | FIII | FV | MII |
| Number of eels | 38 | 21 | 4 | 2 | 7 |
| Nematoda | |||||
| Contracaecum sp. | 6.6–50.0 (3.3) | 6.8–85.7 (5.8) | 66.5–100.0 (66.5) * | 7.6–100.0 (7.6) | 12.5–71.4 (8.9) |
| Urbino | |||
| Age | 4–6 a | 7–9 a | ≥10 a |
| Number of eels | 11 | 47 | 14 |
| Nematoda | |||
| Anguillicola crassus | 75.5–27.3 (20.6) * | 7.6–4.3 (0.3) | 16.9–14.3 (2.4) |
| Contracaecum sp. | 29.3–54.5 (16.0) | 15.2–59.6 (9.1) | 55.4–100.0 (55.4) * |
| Rivers | |||
| Age | 4–6 a | 7–9 a | ≥10 a |
| Number of eels | 30 | 97 | 30 |
| Monogenea | |||
| Pseudodactylogyrus anguillae + P. bini | 17.0–76.7 (13.1) | 22.6–67.0 (15.2) | 60.3–76.7 (46.3) * |
| Nematoda | |||
| Paraquimperia tenerrima | 13.7–23.3 (3.2) | 20.4–21.6 (4.4) | 65.9–36.7 (24.2) * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Esposito, A.; Filippi, J.-J.; Gerbaud, C.; Godeaux, Q.; Millot, R.; Agostini, P.-J.; Albertini, C.; Durieux, E.; Foata, J.; Quilichini, Y. Macroparasite Communities with Special Attention to Invasive Helminths in European Eels Anguilla anguilla from Freshwaters and Brackish Lagoons of a Mediterranean Island. Fishes 2023, 8, 375. https://doi.org/10.3390/fishes8070375
Esposito A, Filippi J-J, Gerbaud C, Godeaux Q, Millot R, Agostini P-J, Albertini C, Durieux E, Foata J, Quilichini Y. Macroparasite Communities with Special Attention to Invasive Helminths in European Eels Anguilla anguilla from Freshwaters and Brackish Lagoons of a Mediterranean Island. Fishes. 2023; 8(7):375. https://doi.org/10.3390/fishes8070375
Chicago/Turabian StyleEsposito, Anaïs, Jean-José Filippi, Charlotte Gerbaud, Quentin Godeaux, Rémi Millot, Paul-Jean Agostini, Camille Albertini, Eric Durieux, Joséphine Foata, and Yann Quilichini. 2023. "Macroparasite Communities with Special Attention to Invasive Helminths in European Eels Anguilla anguilla from Freshwaters and Brackish Lagoons of a Mediterranean Island" Fishes 8, no. 7: 375. https://doi.org/10.3390/fishes8070375
APA StyleEsposito, A., Filippi, J.-J., Gerbaud, C., Godeaux, Q., Millot, R., Agostini, P.-J., Albertini, C., Durieux, E., Foata, J., & Quilichini, Y. (2023). Macroparasite Communities with Special Attention to Invasive Helminths in European Eels Anguilla anguilla from Freshwaters and Brackish Lagoons of a Mediterranean Island. Fishes, 8(7), 375. https://doi.org/10.3390/fishes8070375