Abstract
Vibrio alginolyticus, Vibrio chlorea, and Enterococcus cloacae were the main bacterial pathogens investigated during recurrent episodes of mortalities among farmed flathead grey mullet (Mugil cephalus) in the Bahar El-Baqar area, Sharqia governorate, Egypt, during the summer season in 2021. Nevertheless, the contamination of water with heavy metals was evaluated. In this study, a total number of sixty moribund and freshly dead Mugil cephalus were examined clinically and upon post-mortem. Then, the bacterial isolates were subjected to bacteriological phenotypic examination, 16S rRNA gene sequencing, and antibiotic sensitivity testing. Histopathological screening and the immunity response of naturally infected M. cephalus were also implemented. In areas where bacterial strains were recovered in a high percentage, water contamination with Cu, Fe, and Ni was higher than the permissible limit. The results showed a high similarity (99–100%) to Vibrio and E. cloacae strains using phylogenetic analysis of 16S rRNA. Most isolates exhibited sensitivity to florfenicol, ciprofloxacin, and norfloxacin, while they were resistant to erythromycin and amoxicillin. Hemorrhage, oedema, hemosiderosis, and melanomacrophage activation in most affected tissues of infected M. cephalus were detected by light microscopy. Nevertheless, higher levels of AST and ALT, as well as serum inflammatory markers, CRP, and lysozyme activity, were shown in infected fish. There were higher MDA levels, along with a reduction of GSH, SOD, and catalase, in the gills and brain of infected M. cephalus, compared to the control fish. Our obtained data confirmed the high prevalence of Vibrio serovars and E. cloacae in infected grey mullets, describing their histopathological alterations, higher inflammatory response, and oxidative stress impact. These results could be useful indicators for understanding the pathogenesis of these pathogens in correlation with water parameter deterioration. Thus, the emergence of Vibrio and E. cloacae species in this study threatens the brackish water aquaculture industry in Egypt and poses a public health concern.
Keywords:
Vibrio alginoticus; Enterococcus cloacae; Mugil cephalus; heavy metals; 16S RNA sequencing; histopathology; immune response Key Contribution:
Pathogenic Vibrio and E. cloacae strains were identified at molecular levels in the infected M. cephalus that were associated with higher levels of heavy metals in the affected fish farms.
1. Introduction
In the last few decades, flathead grey mullet (Mugil cephalus) has been one of the most significant candidate fish species in Egyptian aquaculture with a total global production of 250,000 tons in 2018 [1]. Mullets are the next most farmed fish species after Nile tilapia in brackish water, in which tilapia, carp, and mullets are raised together in polyculture earthen pond environments [2,3]. Moreover, grey mullet is the preferred choice of consumers due to its taste, texture, fewer spines, and good nutritive value [3,4].
In Egypt, aquatic ecology is frequently vulnerable to accidental contamination by agrochemical and sewage disposal in addition to the deterioration of water parameters. All of these stress factors have emphasized the outbreaks of bacterial infection among fish species [5,6], particularly flathead grey mullet [7]. Thus, the knowledge base of hazardous bacterial co-infections together with environmental stressors that alter fish health is crucial for investigating effective preventive strategies [8]. In general, concurrent mixed microbial infections have caused serious impacts on the affected fish [9].
Vibriosis is the most threatening bacterial disease facing coastal and estuarine environments [10]. Vibrio species are halophilic Gram-negative, facultative anaerobes, and motile rod bacteria. These pathogens are plentiful in aquatic environments, where they are free-living in water or in association with aquatic fauna, considering opportunistic pathogens in many aquatic animals, such as marine fishes, crustaceans, and bivalves [11,12]. In addition, many Vibrio species have been implicated as causes of human foodborne illnesses as V. cholerae, V. parahaemolyticus, V. vulnificus, and V. alginolyticus are associated with gastroenteritis, severe dehydration, and primary septicemia due to the consumption of contaminated drinking water and undercooked or raw sea food [13,14]. Moreover, the relationship between Vibrio spp. abundance and impairment of environmental factors has been documented in numerous studies and which is strongly correlated with high salinity (30–35 ppt), high temperature (>28 °C), and increased ammonia levels [6,15,16]. These factors suppress the immunity and promote the susceptibility of fish to vibriosis [17,18]. Given these obstacles, Vibrio infection represents a dreadful economic loss in aquaculture and so awakes public concern.
Vibrio alginolyticus has caused mass mortalities among many marine fish farms throughout the Mediterranean area with severe economic losses in aquaculture worldwide [11,19,20,21]. V. alginolyticus has been described as cpathogenic bacteria for turbot (Scophthalmus maximus L.) [22], sea bream (Sparus aurata L.) [23,24], grouper (Epinephelus malabaricus) [25], white leg shrimp (Litopenaeus vannamei) [26], grey mullet (Mugil capito) [27], and M. cephalus [18].
Naturally, fish have a mutualistic relationship with V. cholera as this bacterium secretes proteases and chitinases, with which they digest food (e.g., chitin and protein) in the fish gut [13,14,28]. Previous studies have reported the isolation of V. cholerae from the internal organs of diseased ayu (Plecoglossus altivelis) and guppy fish (Poecilia reticulate) in Japan and Iran, respectively [29,30,31], as well as from Nile tilapia (Oreochromis niloticus) that were cultured in floating cages in Thailand [32]. In addition, V. cholera was identified from the intestine of Sarotherodon galilaeus and M. cephalus [33]. In addition, an intraperitoneal injection of V. cholerae into common carp and rainbow trout caused severe mortalities [34].
Enterobacter cloacae (E. cloacae) is an enteric Gram-negative bacterium belonging to the family Enterobacteriaceae and widely distributed in nature. This opportunistic bacterium is found in the fecal matter of humans and animals, water, soil, plants, insects, and dairy products. Enterobacter species have a resistance to many antibiotics, such as colistin, by expressing the mobile colistin resistance-1 (mcr-1) gene to modify the lipid A component of lipopolysaccharide; subsequently, many human bacterial diseases are becoming incurable. Moreover, foodborne diseases are widely caused by Enterobacteriaceae, which is highly spread among various aquatic systems to be colonized in humans, causing urinary tract infections. In addition, the abuse of antibiotics in the aquatic environment has led to the emergence of new bacterial Enterobacter strains [35,36,37,38,39,40].
Enterobacter strains are not the normal flora in the intestinal tract of fish and, hence, their presence in the infected fish may be related to polluted waters and the feeding behavior of fish [41,42]. Previous studies have assayed the isolation of various Enterobacter strains from brown bull head catfish (Ictalurus nebulosus, LeSueur) [43], dolphin fish (Coryphaena hippurus L.) [44], grey mullet Mugil cephalus [41,45], ornamental fish, Etroplus maculatus [42], and Nile tilapia (Oreochromis niloticus) [46]. Moreover, recent studies documented that E. cloacae is a new-borne bacterial pathogen causing severe deaths to farmed crayfish [47].
Analysis of 16S rRNA is the most precise and prompt diagnostic technique for different bacterial pathogens [48,49], such as Vibrio species [50] and Enterobacter species [41]. In addition, histopathological and immunological alterations, as well as oxidative stress impact, are adopted to be useful indicators for understanding Vibrio species pathogenesis [10].
In this study, we investigated the main pathogenic bacterial agents accompanied by poor water quality as a cause of summer mortalities in flathead grey mullets that were farmed in the Bahr El-Baqar area, Egypt. We initially performed a molecular characterization of bacterial isolates using 16S rRNA as a target marker followed by sequencing and phylogenetic analyses. Moreover, the immune response as well as histopathological changes in M. cephalus were evaluated in response to natural co-infection by Vibrio spp. and E. cloacae.
2. Materials and Methods
2.1. Study Area
Cases of mortality (approximately 40–50 fish/day) were reported in flathead grey mullet, M. cephalus (350 and 500 gm) during the late summer season of 2021 on 3 fish farms in the Bahr El-Baqar area, Sharqia governorate, Egypt (Figure 1). Mullet fish were cultured in 3 different polyculture earthen ponds (70, 15, and 25 acres) at an approximate density of 500–1000 per acre in an integrated system with Mugil capito (M. capito) (3000/acre) and Nile tilapia (15,000–20,000/acre). The aquatic ecosystem was contaminated by agrochemical and sewage drainage. No paddlewheel aerators were used in the farm. Between 40–50 estimated fish deaths were recorded daily per each pond for 15 successive days, with mortality rates peaking on these days and then a gradual decrease in mortalities was noticed after treatment trials.
Figure 1.
Satellite map from Google Earth showing different fish farms located at El-Baqar area, Sharqia governorate, Egypt. (https://www.google.com/maps/@30.9269852,32.1553382,4806m/data=!3m1!1e3!5m1!1e1?entry=ttu, accessed on 22 June 2023).
2.2. Fish Sampling and Clinical Examination
At the peak time of mortalities, a total number of 60 moribund M. cephalus were collected by repeated dip-netting from 3 earthen ponds (20 fish/farm) for further diagnostic procedures and classified as a diseased group. Recently moribund fish were kept in tanks with aerators and then transferred to the laboratory of fish disease and management in the Faculty of Veterinary Medicine, Mansoura University. The clinical signs and post-mortem (PM) lesions were examined according to previously described methods [51]. The procedures for the hygienic disposal of dead fish were performed. Ten apparently healthy M. cephalus specimens were collected from the other earthen ponds of the same farms (ten fish per farm) and examined, to be named as the control group. In addition, water samples were collected from the control and diseased ponds.
Fish were euthanized with clove oil at 60 mgL−1 [52]. Blood samples were then collected from both groups from the caudal vessel using a 5 mL heparinized syringe and a 24 GA needle; serum was separated by centrifugation at 3000 rpm for 10 min at 4 °C, removed and stored in 1.5 mL microcentrifuge tubes at −80 °C for subsequent analysis of some biochemical and immunity parameters. Then, fish were carefully dissected and internal organs were used for bacterial isolation. Gills and brain were excised and stored in cold phosphate buffer saline PBS (pH 7.5) at −20 °C for the determination of oxidative enzymes. Finally, gills, buccal cavity, and internal organs were stored in formalin for histopathological examination.
2.3. Water Quality Measures
The physicochemical characteristics of the pond water were investigated through the collection of water samples from the control and diseased ponds at a depth of 50 cm by a column sampler in sanitary plastic bottles then stored according to typical methods described by [53]. A water thermometer (Yellow Springs Comp., Ohio-model 33) was used to evaluate the water temperatures. Levels of dissolved oxygen (DO) were measured using an oximeter model DO200A EcoSense® (YSI, Brannum Lane, Yellow Springs, OH, USA, EcoSense® products) which was used to measure the percentage of water salinity. A portable Digital PH meter was used to measure the pH value (EcoSense®pH100A Meter). The kits (Palintest, Water Analysis Technology, Golden, CO, USA) were used for the evaluation of the levels of un-ionized ammonia (NH3) and nitrate (NO3). In addition, the analysis of the heavy metals in the water (mgL−1) was carried out using the flame atomic absorption spectrophotometer (AAS) technique [54].
2.4. Bacterial Strains Isolation and Identification
Bacteriological samples were collected aseptically from the kidney, liver, and spleen of diseased M. cephalus, then cultured onto tryptic soya agar plates (TSA; Oxoid, Basingstoke, Hampshire, UK) and thiosulphate citrate bile salts sucrose (TCBS, Difco TM, Becton and Dickinson, ND, USA). Cultures were incubated for 30 ± 2 °C for 24 h. Then, a single distinct colony was carefully chosen from each plate, sub-cultured on the TSA and TCBS plates, and re-incubated under the same conditions. Subsequently, purified bacterial cultures were preliminarily recognized using Gram’s stain, motility, oxidase, and catalase tests using standard biochemical classification techniques API®20-NE, according to the manufacturer’s instructions (BioMérieux, Marcy-l’Étoile, France) [49,55].
2.5. Molecular Identification of Bacterial Isolates
2.5.1. Bacterial DNA Extraction
The QIAamp DNA Mini kit (Qiagen, Hilden, Germany, GmbH, Catalog no. 51304) was used for DNA extraction from the biochemically recognized V. alginolyticus, V. cholera, and E. cloacae isolates, according to the manufacturer’s protocol. Briefly, 200 µL of the sample suspension was incubated with 10 µL of proteinase K and 200 µL of lysis buffer at 56 °C for 10 min. After incubation, 200 µL of 100% ethanol was added to the lysate, which was then washed and centrifuged. The nucleic acid was then eluted with 100 µL of elution buffer and stored at −20 °C for further use in the PCR steps [6].
2.5.2. 16S rRNA PCR, Sequencing, and Phylogenetic Analysis
One universal 16S rRNA primer (5′AGAGTTTGATCMTGGCTCA-3′) and (3′TACGGYTACCTTGTTACGACTT-5′) was used for the molecular identification of V. alginolyticus, V. cholera, and E. cloacae, which amplified a 1485 bp fragment [56]. PCR amplification was performed in a 25 µL reaction mixture containing 12.5 µL of Emerald Amp Max PCR Master Mix (Takara, Kusatsu, Japan), 1 µL of each primer (20 pmol; Metabion, Planegg, Germany), 4.5 µL of distilled water, and 6 µL of DNA template. The reaction was performed in an Applied Biosystems 2720 thermal cycler (Applied Biosystems, Foster City, CA, USA). Amplification started with an initial denaturation step at 94 °C for 5 min, followed by 35 cycles of 94 °C for 30 s, 56 °C for 45 s, and 72 °C for 45 s, and a final extension step was adjusted at 72 °C for 10 min. Next, 15 µL of the amplified PCR products was analyzed by electrophoresis on 1.5% agarose gel (AppliChem, Darmstadt, Germany, GmbH) using gradients of 5 V cm−1, and a 100 bp DNA Ladder (Fermentas, Thermo Scientific, Bremen, Germany) was used to determine the fragment sizes. The bands were visualized by ultraviolet (UV) light and photographed using the agel documentation system (Alpha Innotech, San Leandro, CA, USA; Biometra, Gottingen, Germany).
The QIAquick PCR Product extraction kits (Qiagen, Valencia, Spain) were used to purify PCR products. Then, Bigdye Terminator V3.1 cycle sequencing kits (Perkin Elmer Applied Biosystems, Foster City, CA, USA) were utilized for the sequencing reaction. The genome DNA sequences were obtained using the Applied Biosystems 3130 × Genetic Analyzer (HITACHI, Chiyoda City, Japan). The 16S rRNA sequences were evaluated for their homology using the BLASTn program (http://www.ncbi.nlm.nih.gov/Blast, accessed on 29 May 2023). Multiple alignment through Muscle tool and maximum likelihood phylogenetic analysis were conducted based on the 16S rRNA sequence cluster that positioned our bacterial isolates in a very close proximity to the reference strains of V. alginolyticus, V. cholera, and E. cloacae using the Mega software version 7.0 [57].
2.6. Antibiogram Testing
The antibiotic susceptibility of the pathogenic bacterial strains was carried out using the disc diffusion method of Muller–Hinton Agar (Oxoid, Hampshire, UK) [58,59] supplemented with 1.5% (w/v) sodium chloride. After 24 h of incubation at 30 ± 2 °C, the organisms were classified as sensitive, intermediate, or resistant in accordance with the instructions of the Clinical and Laboratory Standards Institute guidelines [60] through measuring the diameters of the inhibition halos surrounding the discs. The selected antimicrobial discs were [ciprofloxacin (CIP, 5 μg), amoxicillin (AML, 25 μg), norfloxacin (NOR, 10 μg), doxycycline (DO, 30 μg), erythromycin (E, 15 μg), and florfenicol (F, 30 μg)], which were obtained from Bioanalyse® (Ankara, Turkey).
2.7. Serum Biochemical and Immune Markers
The serum activities of alanine aminotransferase (ALT, Catalog No.; MBS038444), (MyBioSource Co., San Diego, CA, USA), and aspartate aminotransferase (AST, Catalog No.; EK12276) (Biotrend Co., MD, USA), as well as levels of total protein (Catalog No.; MBS9917835) (MyBioSource Co., San Diego, CA, USA) and albumin (Catalog No.; SB-028-500) (Stanbio Laboratory, Boerne, TX, USA), were estimated spectrophotometrically according to the standard protocol of their specific pamphlets using a spectrophotometer (Lambda EZ201; Perkin Elmer, Waltham, MA, USA) [61,62].
The serum levels of immunoglobulin M (IgM, Catalog No.; CSB-E12045Fh, complement (C3, Catalog No.; MBS281020)) and C-reactive protein (CRP, Catalog No.; MBS016586) were assayed using the kits obtained from Cusabio Co. (Houston, TX, USA) and MyBioSource Co. (San Diego, CA, USA), respectively, based on the manufacturer’s instructions [63,64,65].
Serum lysozyme activity was measured according to the previously described method [66] based on the lysis of Micrococcus lysodeikticus (Sigma Co., Ronkonkoma, NY, USA), with some modifications. The serum and M. lysodeikticus suspension (0.2 mg/mL in 0.05 M PBS, pH 6.2) mixture was reacted at 25 °C for 5 min. The optical density was then measured at 1 min intervals for 5 min at 540 nm (5010, Photometer, BM Co., Herne, Germany). The concentration of serum lysozyme was calculated using a calibration curve constructed using different dilutions of lyophilized chicken egg-white lysozyme (Sigma Co., Ronkonkoma, NY, USA).
2.8. Oxidative Stress/Antioxidant Parameters
Parts of gills and brain tissues from apparently healthy and diseased stripped mullet were washed three times using cold NaCl solution (0.9%) and homogenized in cold PBS (pH 7.5). Later, the homogenates were cold centrifuged for about 15 min at 3000 rpm and the supernatants were carefully collected in a clean tube to be used in the evaluation of antioxidant and oxidative stress parameters [67]. The homogenate protein level was determined using the protocols described [68].
The gills and brain levels of malondialdehyde (MDA, catalog No.; MD 25 29), glutathione (GSH, catalog No.; No. GR 25 11), catalase (catalog No.; CA 25 17), and superoxide dismutase (SOD, catalog No.; SD 25 21) were estimated spectrophotometrically using commercial test kits obtained from Biodiagnostics company (Cairo, Egypt).
2.9. Histopathological Examination
The brain, liver, spleen, and gills were dissected from the control and diseased M. cephalus and fixed in 10% neutral buffered formalin for 24 h. The dissected organs were placed in tissue cassettes, processed, and embedded in paraffin wax. Embedded samples were sliced into 5 µm sections by a microtome. The sections were stained using haematoxylin and eosin [69]. The stained slides were examined under a light microscope (Olympus CX 31, Tokyo, Japan).
2.10. Statistical Analysis
Data were expressed as mean ± standard error (SE). The differences among all parameters were applied via the SPSS software program (version 20, (IBM Corporation, SPSS Statistics, Chicago, IL, USA) using independent samples t-test to determine the differences between groups.
3. Results
3.1. Clinical Signs and Necropsy Findings
The diseased M. cephalus samples were anorexic and exhibited external generalized erythematic signs, such as severe hemorrhages on the opercula, buccal cavity, and on the base of fins with eroded fins, as well as an inflamed swollen vent was noticed (Figure 2). On necropsy, the PM lesions revealed congested gills. In addition, the abdominal cavity was loaded with copious amounts of bloody ascetic fluids with a distended gall bladder. Congested kidney tissue with surface hemorrhages over the brain, liver, and intestinal mucosa were noticed (Figure 3).
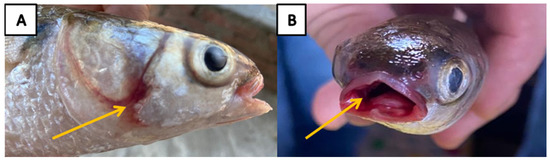
Figure 2.
(A) Naturally diseased cultured flathead grey mullet (Mugil cephalus) showed opercular hemorrhage (yellow arrow). (B) Buccal cavity hemorrhage (yellow arrow) was also observed.
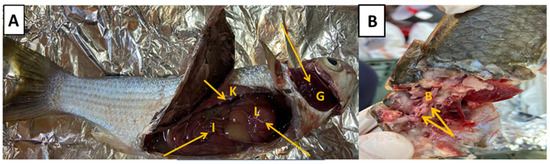
Figure 3.
(A) Naturally infected grey mullet (Mugil cephalus) showed congested gills (G), hemorrhage of intestinal mucosa (I), congested kidney (K), and surface hemorrhages over liver (L) (yellow arrows). (B) Severe hemorrhages over the brain (B) were revealed (yellow arrow).
3.2. Water Physicochemical Properties
The quality parameters of the pond water collected from the Bahr El-Baqar fish farms during the mass mortalities of M. cephalus are summarized in Table 1. The results showed a decrease in dissolved oxygen (4 mgL−1) and elevated levels of un-ionized ammonia (0.90 mgL−1) and nitrite (0.07 mgL−1) were detected. In addition, the measured levels of heavy metals were 0.64, 1.9, 0.8, 0.09, and 0.40 mgL−1 for copper (Cu), iron (Fe), nickel (Ni), cadmium (Cd), and lead (pb), respectively. These levels are over the recommended permissible levels [70].

Table 1.
The physicochemical properties of pond water in Bahr El-Baqar fish farms during mortalities.
3.3. Phenotypic Identification of Bacterial Pathogens
The strains of V. alginolyticus, V. cholera, and E. cloacae were isolated and identified from the diseased fish following phenotypic characterization, as illustrated in Table 2. V. alginolyticus was isolated from 50% (30/60) of the sampled M. cephalus. Meanwhile, V. cholerae was isolated from 25% (15/60) of the sampled fish and E. cloacae was successfully isolated from 10% (6/60). In addition, 15% of the fish harbored a mixed infection of two or three isolates which were identified.
Table 2.
The physicochemical properties of pond water in Bahr El-Baqar fish farms during mortalities.
3.4. Molecular Identification of Bacterial Pathogens
PCR successfully amplified the 1485 bp band of the universal 16S rRNA primers of Gram-negative bacteria (Figure 4). One sequence of V. alginolyticus was deposited in the GenBank (Accession No. OM459825) and two sequences of V. cholerae were deposited in the GenBank (Accession No. OM185314 and OM841535). One sequence of E. cloacae was also deposited in the GenBank (Accession No. OM914983). BLAST analysis of the nucleotide sequence of the 16S rRNA genes from V. alginolyticus in this study shared 94.16% to 100% identity with other published sequences of V. alginolyticus obtained from the GenBank database (Figure 5). Similarly, the 16S rRNA genes of V. chlorea sequenced in this study shared 99.44% to 99.87% identity with other published sequences of V. chlorea obtained from the GenBank database (Figure 4). In addition, the sequence of E. cloacae in our study shared 99.16% to 100% identity with other published sequences of E. cloacae obtained from the GenBank database (Figure 6). The phylogenetic analysis of the amplified 16S rRNA sequences from V. alginolyticus, V. cholera, and E. cloacae were grouped together with their relevant sequences in the GenBank database, respectively (Figure 5 and Figure 6).
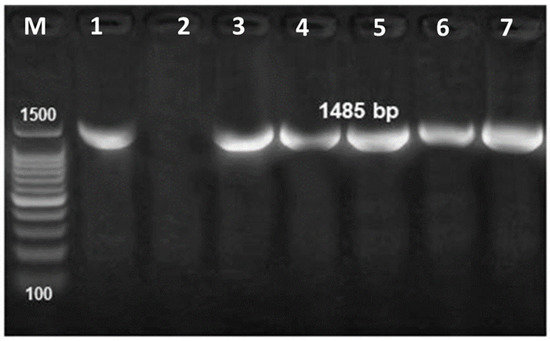
Figure 4.
PCR amplification of the universal 16S rRNA primers of Gram-negative bacteria at 1485 bp. M: 1000–1500 bp DNA size marker, Lane 1: control positive and Lane 2: control negative, Lane 3–7: bacterial strains obtained from naturally infected flathead grey mullet.

Figure 5.
Maximum likelihood phylogenetic tree showing the relationships between the Vibrio strains (three strains) isolated from diseased grey mullet, Mugil cephalus, and other reference strains of Vibrio spp. based on the 16S rRNA sequences of Vibrio spp. The numbers above the branches are the percentages determined in a bootstrap analysis (1000 replicates).

Figure 6.
Maximum likelihood phylogenetic tree showing the relationships between the E. cloacae strain isolated from diseased grey mullet, Mugil cephalus, and other reference strains of E. cloacae spp. based on the 16S rRNA sequences. The numbers above the branches are the percentages determined in a bootstrap analysis (1000 replicates).
3.5. Susceptibility of Pathogenic Strains to Antibiotics
The Vibrio strains were sensitive to norfloxacin, florfenicol, and ciprofloxacin, whereas they were resistant to amoxicillin. They also exhibited a moderate sensitivity to erythromycin and doxycycline. The E. cloacae strain was sensitive to doxycycline, while was exhibited an absolute resistance to erythromycin, as well as a moderate sensitivity being exhibited to ciprofloxacin and amoxicillin (Table 3).
Table 3.
Antibiogram profile of pathogenic Vibrio spp. and E. cloacae strains.
3.6. Serum Biochemical and Inflammatory Findings
As illustrated in Table 4, the obtained data demonstrated significantly higher serum activities of ALT (p < 0.01) and AST (p < 0.001) in the diseased stripped mullet than that of the controls. There were no significantly valuable differences in the serum levels of total protein, albumin, globulin, and C3 (p ≥ 0.05) between the groups.
Table 4.
Serum biochemical and immune markers in control and diseased M. cephalus.
To assess the mechanisms involved in the progression and damage induced by bacterial infection, some serum inflammatory markers were estimated in our study (Table 4). The serum CRP level and lysozyme activity (p < 0.01) were found to be highly elevated with a lower IgM level (p < 0.001) in the naturally infected flathead grey mullet compared to the control one.
3.7. Oxidative Stress/Antioxidant Status
The progression of infection caused the generation of an accentuated ROS and impairment of antioxidant molecules confirmed in our results by the higher MDA levels, along with a reduction of GSH, SOD, and catalase in both the gills and brain of the diseased flathead grey mullet compared to the control one (Table 5).
Table 5.
Gills and brain oxidative stress/antioxidant status of healthy and infected M. cephalus.
3.8. Histopathological Findings
The diseased and control M. cephalus were examined for histopathological changes during the peak of mortalities. The liver had diffuse necrosis with a cellular detachment that was accompanied by an eosinophilic cytoplasm and pyknotic to a complete loss of the nucleus. Furthermore, there was diffuse edema and hemorrhage admixed with hemosiderin, widely separated damaged hepatocytes, and congested blood vessels (Figure 7A–C). In the spleen, there was a multifocal splenic granuloma and necrosis infiltrated with numerous melanomacrophages that were surrounded by numerous lymphocytic aggregations and leukocytic infiltrations (Figure 7D–F). The gills had marked diffuse lamellar sloughing, necrosis with lamellar telangiectasis, and interlamellar necrotic debris with severe interlamellar hemorrhage (Figure 7A–C). The most characteristic findings of the fish brain are congested and dilated blood vessels, multifocal granuloma with few inflammatory cells, and numerous degenerated cells with focally leukocytic infiltrations (Figure 8D–F).
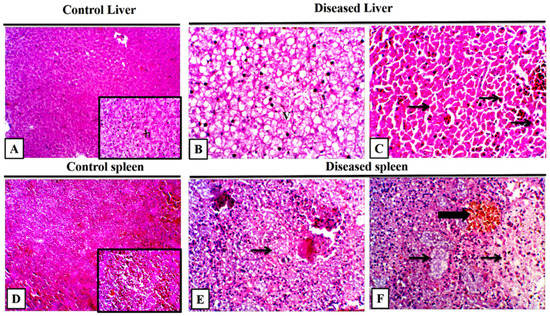
Figure 7.
Representative photomicrograph of grey mullet (Mugil cephalus) liver and spleen. (A) Control liver showed normal hepatic architecture (h) in low and high power, H and E, 100×, and 400×. (B) Diseased mullet liver showed a diffuse hepatic vacuolation (v), H and E, 400×. (C) Diseased liver showed a diffuse hepatic individualization with pyknotic nuclei (arrows), H and E, 400×. (D) Control spleen showed normal splenic architecture. (E,F) Diseased spleen showed a loss of normal architecture with lytic necrosis (thin arrows) and melanomacrophage aggregations (thick arrow), H and E, 400×.
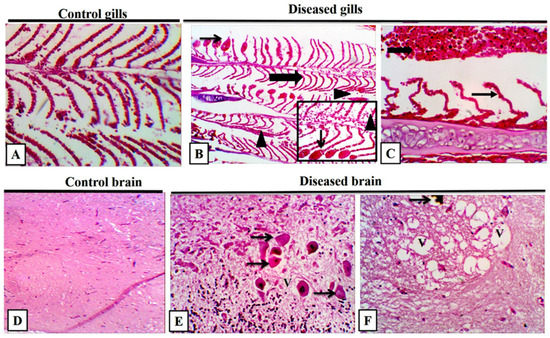
Figure 8.
Representative photomicrograph of grey mullet (Mugil cephalus) gills and brain. (A) Control gills showed a normal lamellar architecture with minimal sloughed epithelium. (B) Diseased gills showed clubbing (thick arrow) and telangiectasis of the gill lamellae (thin arrow) with detachment of gill epithelium (arrow heads), high power focus on telangiectasis (thin arrow), and interlamellar sloughed epithelium admixed with RBCs (arrow head). (C) Lamellar lifting (thin arrow) with extensive interlamellar hemorrhage (thick arrow) was revealed. H and E, 400×. (D) Control brain showed a normal histological appearance. (E) Diseased brain showed nerve cell bodies exhibiting pyknotic or complete loss of nuclei with shrunken cellular outline and hypereosinophilic cytoplasm (neuronal necrosis) (thin arrows) with moderate neuropil vacuolation H and E, 400×. (F) Neuronal parenchyma with clustered, focal variably sized, clear vacuoles (V), few vacuoles had brown pigment (thin arrow) H and E, 400×.
4. Discussion
The opportunistic bacteria from the genus Vibrio are ranked number one among all the pathogenic agents which have caused mortalities in estuarine and marine fish farms [10,48]. Environmental stressors have been correlated with Vibrio spp. abundance that can be used as a predictive strategy to determine when these pathogens may be present [6,11,71]. Moreover, Vibrios outbreaks mainly occur during the summer season, as a result of the deterioration of water parameters such as the temperature, pH, DO, and salinity [72]. This pathogen load was also enhanced, particularly in developing countries that have disputed sanitary circumstances [73]. Given these constraints, the prevalence of bacterial infections has become a crucial problem faced via an M. cephalus culturing strategy [7].
In the current work, the most common generalized septicemic clinical signs and PM lesions were assigned in the infected striped mullet M. cephalus infected with Vibrio spp., reporting a sudden onset of mortalities during the summer season in many fish farms native to the Bahr El-Baqar region, Sharqia governorate. These findings were similarly reported in M. capito infected with V. alginoticus in the Wadi-mariut region, Borg El-Arab city, Alexandria governorate, Egypt [27]. In concordance with our results, many studies have reported the same typical signs of Vibrio infection in M. capito and M. cephalus [11,16,48,74]. A recent study showed septicemic lesions in diseased M. cephalus with V. parahaemolyticus in Lake Al-Manzala [6]. These results could be attributed to the detrimental effect of several virulence factors produced by Vibrio spp., such as siderophore anguibactin, iron-binding capacity, the adherence capacities to epithelial cells of V. alginoticus, and extracellular products (e.g., hemolysin, cytotoxin, lipase, protease, and phospholipase) [75,76]. These factors are responsible for invasive and proliferative pathogenicity in fish. Of particular concern, circulatory component disturbances and immune suppression are mediated by these extracellular products [10]. It has been documented that the Enterococcus species caused septicemic-like infections in Nile tilapia in Egypt, Thailand, and Bangladesh [77,78].
Our data represented decreased DO levels accompanied by elevated levels of NH3 and NO2 along with a high temperature and increased heavy metal levels (more particularly, Cu, Fe, Ni, Cd, and pb) in water. The above-mentioned substandard water quality results are positively associated with increased fish susceptibility to bacterial diseases which are consequent to the immune suppression of fish. It was noted that the continuous unhygienic disposal of agricultural and sewage pollutants in the water supply of the Bahr EL-Baqar drain usually led to a deterioration of the water quality parameters [79,80]. A potential correlation was observed between the Vibrio spp. infections in diseased M. cephalus which suffered from mortalities in numerous Egyptian fish ponds during the summer season and poor water characteristics including high levels of heavy metals [6,81]. In similar studies, a sharp increase in NH3, water salinity, and temperature with decreased dissolved oxygen and ability to scavenge iron from iron-binding proteins are the most likely triggering factors for the spread of V. alginoticus infections in coastal areas in China [82], and also in many Egyptian coastal sites [83], including Lake Al-Manzala’s fish farms [8,84]. Moreover, several reports assayed that abnormal water parameters are the pre-disposing factors for Enterococcosis in Egyptian fish farms [46,77]. Higher organic matter and un-ionized ammonia detected in the earthen ponds in the Kafr El sheikh governorate and supplied with agriculture drainage water increased the risk of infectious Enterococcus faecalis in Nile tilapia [46]. Nevertheless, a higher prevalence of Vibrio species in Mugil cephalus were isolated from Lake Qarun, and this could be related to the deterioration of water quality via the sewage and agriculture discharges of Lake Qarun [18]. Changes in the water quality parameters in highly polluted Abbassa farms with subsequent heavy metal bioaccumulation enhanced the bacterial infections in many farmed fish species [85].
Additionally, heavy metals pollution, such as Ni [85,86] and Cu [87], exhibited an immunosuppressive effect in many teleost fish. Iron, an excellent oxygen transporter, has been shown to stimulate the growth of Vibrio spp. by iron acquisition through chelation to host iron-binding proteins (e.g., transferrin and lactoferrin) [88,89]. Copper toxicity has caused respiratory stress from the coagulation of the mucus layer of the gills, prohibiting oxygen transport, which was accompanied by reduced phagocytic activity [90]. Elevated levels of Fe, Cu, and Ni were positively related with vibriosis in Nile tilapia and silver carp from fish farms in Lake Al-Manzala during the summer season [8]. In addition, a high percentage of Vibrio was isolated from farmed M. cephalus in many Egyptian provinces where water was contaminated with Cu and Fe [91]. Consequently, it is plausible that the levels of metals detected in our study could have contributed to the outbreaks of disease with the resulting immunosuppression in stripped mullet.
The insight analysis of the morpho-chemical characteristics for the majority of retrieved isolates coincided with the Vibrio sp. profiles as previously reported in diseased farmed M. cephalus [6,11,18,92]. Similarly, the phenotypic characteristics of the E. cloacae isolate in farmed M. cephalus corresponded with the description of E. cloacae in infected M. cephalus in the lagoon of Muttukadu, Chennai, India [41], and also with the E. cloacae strain from diseased crayfish (Procambarus clarkii) in China [47]. Biochemically identified isolates were further confirmed by the 16S rRNA PCR gene; however, this gene has a low distinguishing power to differentiate closely related Vibrio species that have nearly identical sequences [18,93].
The results of the phylogenetic analysis based on the 16S rRNA gene sequences of the bacterial isolates retrieved during the current study proposed that this gene is useful in allocating species to various branches of the Vibrionaceae family [94] and to the Enterobacteriaceae family [7]. The bacterial isolates identified in the present study shared nearly identical sequences in the 16S rRNA gene (99.5% to 100% identity). Subsequently, 16S rRNA gene sequencing analyses are highly discriminative as phylogenetic identifiers in ascertaining Vibrio and E. cloacae species.
The isolated Vibrio strains were highly sensitive to norfloxacin, florfenicol, and ciprofloxacin, whereas they were resistant to amoxicillin. These findings are in line with previous studies [83,95]. In addition, the E. cloacae strain was sensitive to doxycycline, while it exhibited absolute resistance to erythromycin, as well as an intermediate sensitivity being exhibited to ciprofloxacin and amoxicillin. Similar results were reported in virulent E. cloacae isolated from diseased crayfish, Procambarus clarkii [47]. Therefore, these selected antibiotics could be recommended for prevention during the peak season of disease manifestation.
Many fish species have shown changes in blood chemistry parameters during acute bacterial infections. The biomarkers ALT and AST are important indicators of liver dysfunction and injury [96], and their detection in blood is due to the release of these enzymes into the bloodstream by damaged hepatocytes [97]. Our data revealed significantly higher serum activities of ALT (p < 0.01) and AST (p < 0.001) with insignificant changes in the serum levels of the total protein, albumin, globulin, and C3 (p ≥ 0.05) of the diseased stripped mullet than that of the controls. Similarly, the ALT and AST activities were higher in the Aeromonas hydrophila and Pseudomonas fluorescens-infected Nile tilapia [98]. Furthermore, Rhamdia quelen infected with Aeromonas hydrophila had higher ALT levels than the control group [99]. Additionally, another previous study found that Vibrio vulnificus or Streptococcus iniae infection caused dramatically higher ALT and AST, with lower total protein and albumin levels in infected fish [100]. Unlike our findings, complement pathway activation has been recorded in M. amblycephala after infection with A. hydrophila, mandarin fish (Siniperca chuatsi) infected with Flavobacterium columnare, and soiny mullet (Liza haematocheila) in response to Streptococcus dysgalactiae infection [101,102,103].
IgM and lysozyme are involved in a fish’s innate and adaptive immune responses to bacterial infection [104]. C-reactive protein (CRP) is a cyclic serum protein that is released from the serum of many fish species during inflammation, making it a well-acknowledged biomarker of fish health status [105]. In our study, the serum CRP level and lysozyme activity were significantly elevated with a lower IgM level (p < 0.001) in the naturally infected flathead grey mullet compared to the control one. A previous report documented that the common carp was injected with Aeromonas hydrophila and Escherichia coli lipopolysaccharide [106]. In addition, CRP levels in rainbow trout serum have been shown to be high interacting with Aeromonas salmonicida-derived LPS, suggesting that a number of virulence factors are involved in causing a pro-inflammatory response to bacterial infection in fish [107]. Likewise in our results, lysozyme activity was considerably higher in the A. hydrophila and Photobacterium damselae-infected Nile tilapia [65]. On day 1, 3, 5, 14, and 21 after A. hydrophila infection, a similar reaction was observed in P. damselae subsp. piscicida (Phdp)-infected Solea senegalensis [108,109], as well as in blunt snout bream (Megalobrama amblycephala) [110]. Furthermore, in the liver of S. senegalensis infected with Phdp, the lysozyme gene was dramatically increased [111]. The high level of serum lysozyme in infected fish implies a natural protective mechanism in fish against infection [112].
Oxidative stress is defined as an imbalance in antioxidant–oxidant status, which can impact the redox status of cell compartments in a reversible manner, causing harm to biological cell components such as lipid peroxidation and protein carbonylation [113]. Overproduction of reactive oxygen species (ROS) and/or a weakening of the defense antioxidant system during bacterial infection can cause oxidative stress in fish [114]. Pseudomonas aeruginosa infection caused an elevation in the brain lipid peroxide (MDA) of silver catfish, which led to the emergence of clinical symptoms and disease pathogenesis [115]. Moreover, Providencia rettgeri infection induces oxidative damage in infected O. niloticus via increasing hepatic and renal MDA levels and reducing SOD and catalase activities [116]. Similarly, the SOD and GSH levels were diminished with higher ROS levels but were not affected in the liver and kidney of silver catfish (Rhamdia quelen) experimentally infected with Aeromonas caviae, and in the liver of sea trout (Salmo trutta L.) experimentally infected with A. hydrophila [114,117]. Our data demonstrated that Vibrio and Enterococcous infection caused the generation of ROS (higher MDA levels) with an impairment of antioxidant molecules (reduction of GSH, SOD, and catalase) in both the gills and brain of the diseased flathead grey mullet compared to the control one. This finding could indicate that the antioxidant systems failed to protect the gills and brain tissues from the harmful effects of ROS during Vibrio and Enterococcous infection, thereby contributing to disease pathogenesis.
The pathological lesions in naturally exposed fish to Vibrio spp. and E. cloacae were presented in this study. Here, we clearly monitored that the main pathological changes in the affected tissues of diseased M. cephalus were septicemic lesions, such as hemorrhage, oedema, hemosiderosis, and proliferative and necrotic alterations. These lesions are considered the main changes in the natural infection in the acute phase of septicemic diseases, including vibriosis [49,118]. Similar findings were stated in the internal organs and gills of seabass [118,119], thin lip grey mullet, M. capito [27], and M. cephalus [91] upon infection with V. alginolyticus. In addition, extensive inflammatory and degenerative histopathological changes in fish infected with Enterococcus species were observed in recent reports [46,47]. A recent study observed similar lesions in M. cephalus infected with Streptococcus agalactiae and Enterococcus faecalis [7]. Altogether, the obtained data of the full diagnostic techniques accompanied by immunological and histopathological alterations are considered as proof of the septicemic nature of the bacterial co-infection in M. cephalus.
5. Conclusions
The deterioration in water quality via sewage and agriculture discharges was strongly related to a high prevalence of vibriosis and Enterococcosis in flathead grey mullet farmed in the Bahar El-Baqar area, Egypt. V. alginolyticus, V. cholera, and E. cloacae are the most causative bacterial species that were identified from diseased M. cephalus via phenotypic and molecular characterization. In addition, they are associated with septicemic clinical signs, pathological, and immunological alterations. Under the conditions of this study, the obtained data revealed that the monitoring of the water for heavy metals pollution is a crucial step to diminish the severity of Vibrio serovars infections. Subsequently, further investigations are required to assess the food safety regulations and the consumer’s health in the growing aquaculture sector in Egypt.
Author Contributions
S.E., G.E.E. and M.A.M.E.-S.: methodology, formal analysis, investigation, validation, writing—original draft, writing—review and editing; S.E. and S.J.D.: conceptualization, supervision, and final revision; M.S.S., E.A.A.E., A.A.A., E.M.Y. and S.J.D.: investigation, visualization, and methodology; A.A.A., E.M.Y. and S.J.D.: Resources. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the researchers supporting project number (RSPD2023R700), King Saud University, Riyadh, Saudi Arabia.
Institutional Review Board Statement
All protocols were approved and conducted following the Medical Research Ethics Committee of Veterinary Medicine, Mansoura University, following the general guidelines of the Canadian Council on Animal Care with a code number (R/132).
Informed Consent Statement
Not applicable.
Data Availability Statement
The data that support the findings of this study are available on reasonable request from the corresponding author, S. Elbahnaswy. The data are not publicly available due to their containing information that could compromise the privacy of research participants. The sequences generated in the present study were deposited in the NCBI GenBank database under the accession numbers OM459825, OM185314, OM841535, and OM914983.
Acknowledgments
This research was funded by the researchers supporting project number (RSPD2023R700), King Saud University, Riyadh, Saudi Arabia.
Conflicts of Interest
The authors do not report any financial or personal connections with other persons or organizations that might negatively affect the contents of this publication and/or claim authorship rights to this publication.
References
- GAFRD. General Authority for Fish Resources Development; GAFRD: Nasr City, Egypt, 2019.
- Soyinka, O.O. The feeding ecology of Mugil cephalus (Linnaeus) from a high brackish tropical lagoon in South-west, Nigeria. Afr. J. Biotechnol. 2008, 7, 4192–4198. [Google Scholar]
- Plumber, A. Fishy Business: Assessing Egypt’s Growing Aquaculture Sector. Al Noor J. Middle East. Stud. 2019, 44–51. [Google Scholar]
- Sukumaran, K.; Thomas, D.; Rekha, M.; Angel, J.R.J.; Bera, A.; Mandal, B.; Subburaj, R.; Thiagarajan, G.; Makesh, M.; Ambasankar, K. Reproductive maturation and induced breeding of two geographical groups of grey mullet, Mugil cephalus Linnaeus, 1758. Aquaculture 2021, 536, 736423. [Google Scholar] [CrossRef]
- Abou El-gheit, E. Some Investigations on the role of water parameters in microbial infections of fishes. Egypt. J. Exp. Biol. 2005, 1, 9–14. [Google Scholar]
- El-Son, M.A.; Nofal, M.I.; Abdel-Latif, H.M. Co-infection of Aeromonas hydrophila and Vibrio parahaemolyticus isolated from diseased farmed striped mullet (Mugil cephalus) in Manzala, Egypt–A case report. Aquaculture 2021, 530, 735738. [Google Scholar] [CrossRef]
- Mahmoud, M.A.; Attia, M.M.; Abdelsalam, M.; Abdel-Moneam, D.A.; Zaki Ewiss, M. Ergasilus extensus and bacterial co-infection in flathead grey mullet, Mugil cephalus (Linnaeus, 1758), are associated with pathological changes and immunological gene expression alterations. Aquac. Res. 2021, 52, 6143–6151. [Google Scholar] [CrossRef]
- Nofal, M.I.; Abdel-Latif, H.M. Ectoparasites and bacterial co-infections causing summer mortalities among cultured fishes at Al-Manzala with special Reference to Water quality parameters. Life Sci. J. 2017, 14, 72–83. [Google Scholar]
- Kotob, M.H.; Menanteau-Ledouble, S.; Kumar, G.; Abdelzaher, M.; El-Matbouli, M. The impact of co-infections on fish: A review. Vet. Res. 2017, 47, 98. [Google Scholar] [CrossRef]
- Ina-Salwany, M.; Al-saari, N.; Mohamad, A.; Mursidi, F.A.; Mohd-Aris, A.; Amal, M.; Kasai, H.; Mino, S.; Sawabe, T.; Zamri-Saad, M. Vibriosis in fish: A review on disease development and prevention. J. Aquat. Anim. Health 2019, 31, 3–22. [Google Scholar] [CrossRef]
- El-Bouhy, Z.; El-Nobi, G.; El-Murr, A.; Abd El-Hakim, S. Study on Vibriosis in Mugil Capito in El-Dakahlia and Damitta Governorates, Egypt. Abbassa Int. J. Aquat. 2016, 9, 2016. [Google Scholar]
- Austin, B. Vibrios as causal agents of zoonoses. Vet. Microbiol. 2010, 140, 310–317. [Google Scholar] [CrossRef] [PubMed]
- Traoré, O.; Martikainen, O.; Siitonen, A.; Traoré, A.S.; Barro, N.; Haukka, K. Occurrence of Vibrio cholerae in fish and water from a reservoir and a neighboring channel in Ouagadougou, Burkina Faso. J. Infect. Dev. Ctries. 2014, 8, 1334–1338. [Google Scholar] [CrossRef]
- Flach, E.J.; LeNette-Dawson, D.; Greig, D.R.; Ismail Ahmed, A.; Jenkins, C.; John, S.K.; Macgregor, S.K.; Masters, N.; Stidworthy, M.F.; Zimmerman, B. Isolation and characterisation of Vibrio cholerae from fish examined postmortem at ZSL London Zoo between 2014 and 2018. Vet. Rec. 2020, 187, e86. [Google Scholar] [CrossRef] [PubMed]
- Gayathri, V. The Prevalence and Public Health Significance of Human Pathogenic Vibrio Species in Hawaii’s Diverse Tropical Coastal Water Environments; Water Resources Research Centre, University of Hawaii: Honolulu, HI, USA, 2011. [Google Scholar]
- El-Tawab, A.; El-Hofy, F.; Hasb-Elnaby, G.; El-Khayat, M.; Refaey, M. Prevalence and virulence genes of vibrio and Aeromonas species isolated from Nile tilapia and mugil fish farms in Egypt. Adv. Anim. Vet. Sci. 2021, 9, 1625–1631. [Google Scholar]
- Haenen, O.; Van Zanten, E.; Jansen, R.; Roozenburg, I.; Engelsma, M.; Dijkstra, A.; Boers, S.; Voorbergen-Laarman, M.; Möller, A. Vibrio vulnificus outbreaks in Dutch eel farms since 1996: Strain diversity and impact. Dis. Aquat. Org. 2014, 108, 201–209. [Google Scholar] [CrossRef]
- Abdelaziz, M.; Ibrahem, M.D.; Ibrahim, M.A.; Abu-Elala, N.M.; Abdel-moneam, D.A. Monitoring of different vibrio species affecting marine fishes in Lake Qarun and Gulf of Suez: Phenotypic and molecular characterization. Egypt. J. Aquat. Res. 2017, 43, 141–146. [Google Scholar] [CrossRef]
- Snoussi, M.; Hajlaoui, H.; Noumi, E.; Zanetti, S.; Bakhrouf, A. Phenotypic and genetic diversity of Vibrio alginolyticus strains recovered from juveniles and older Sparus aurata reared in a Tunisian marine farm. Ann. Microbiol. 2008, 58, 141–146. [Google Scholar] [CrossRef]
- Marhual, N.P.; Das, B.K.; Sadique, M.; Swain, A.K.; Mishra, B.K.; Maiti, N.K.; Eknath, A.E. Molecular identification and typing of Vibrio alginolyticus and Vibrio parahaemolyticus strains isolated from black tiger shrimp Penaeus monodon. J. Aquac. Trop. 2010, 25, 25. [Google Scholar]
- Gong, Q.-y.; Yang, M.-j.; Yang, L.-f.; Chen, Z.-g.; Jiang, M.; Peng, B. Metabolic modulation of redox state confounds fish survival against Vibrio alginolyticus infection. Microb. Biotechnol. 2020, 13, 796–812. [Google Scholar] [CrossRef]
- Austin, B.; Stobie, M.; Robertson, P.; Glass, H.; Stark, J.; Mudarris, M. Vibrio alginolyticus: The cause of gill disease leading to progressive low-level mortalities among juvenile turbot, Scophthalmus maximus L., in a Scottish aquarium. J. Fish Dis. 1993, 16, 277–280. [Google Scholar] [CrossRef]
- Colorni, A.; Paperna, I.; Gordin, H. Bacterial infections in gilt-head sea bream Sparus aurata cultured at Elat. Aquaculture 1981, 23, 257–267. [Google Scholar] [CrossRef]
- Balebona, M.C.; Andreu, M.J.; Bordas, M.A.; Zorrilla, I.; Moriñigo, M.A.; Borrego, J.J. Pathogenicity of Vibrio alginolyticus for cultured gilt-head sea bream (Sparus aurata L.). Appl. Environ. Microbiol. 1998, 64, 4269–4275. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.-K. Pathogenesis studies on Vibrio alginolyticus in the grouper, Epinephelus malabaricus, Bloch et Schneider. Microb. Pathog. 1995, 19, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Santhyia, A.V.; Mulloorpeedikayil, R.G.; Kollanoor, R.J.; Jeyaseelan, P.M. Molecular variations in Vibrio alginolyticus and V. harveyi in shrimp-farming systems upon stress. Braz. J. Microbiol. 2015, 46, 1001–1008. [Google Scholar] [CrossRef] [PubMed]
- Khalil, R.; Abd El-Latif, H. Effect of Vibrio alginolyticus on Mugil Capito. J. Arab. Aquacult. Soc. 2013, 8, 193–204. [Google Scholar]
- Lutz, C.; Erken, M.; Noorian, P.; Sun, S.; McDougald, D. Environmental reservoirs and mechanisms of persistence of Vibrio cholerae. Front. Microbiol. 2013, 4, 375. [Google Scholar] [CrossRef]
- Yamanoi, H.; Muroga, K.; Takahashi, S. Physiological characteristics and pathogenicity of NAG Vibrio isolated from diseased ayu. Fish Pathol. 1980, 15, 69–73. [Google Scholar] [CrossRef]
- Kiiyukia, C.; Nakajima, A.; Nakai, T.; Muroga, K.; Kawakami, H.; Hashimoto, H. Vibrio cholerae non-O1 isolated from ayu fish (Plecoglossus altivelis) in Japan. Appl. Environ. Microbiol. 1992, 58, 3078–3082. [Google Scholar] [CrossRef]
- Kiani, S.; Naghavi, N.S.; Nazari, A. Detection of Vibrio species isolated from ornamental guppy fish in Kashan, Isfahan, Iran fish culturing pounds. Biol. J. Microorg. 2016, 4, 43–48. [Google Scholar]
- Dong, H.T.; Nguyen, V.V.; Le, H.D.; Sangsuriya, P.; Jitrakorn, S.; Saksmerprome, V.; Senapin, S.; Rodkhum, C. Naturally concurrent infections of bacterial and viral pathogens in disease outbreaks in cultured Nile tilapia (Oreochromis niloticus) farms. Aquaculture 2015, 448, 427–435. [Google Scholar] [CrossRef]
- Halpern, M.; Izhaki, I. Fish as Hosts of Vibrio cholerae. Front. Microbiol. 2017, 8, 282. [Google Scholar] [CrossRef]
- Rehulka, J.; Petras, P.; Marejkova, M.; Aldova, E. Vibrio cholerae non-O1/non-O139 infection in fish in the Czech Republic. Vet. Med. 2015, 60, 16–22. [Google Scholar] [CrossRef]
- Song, G.; Zhou, Y.; Niu, S.; Deng, X.; Qiu, J.; Li, L.; Wang, J. Nordihydroguaiaretic acid reverses the antibacterial activity of colistin against MCR-1-positive bacteria in vivo/in vitro by inhibiting MCR-1 activity and injuring the bacterial cell membrane. Phytomedicine 2022, 98, 153946. [Google Scholar] [CrossRef] [PubMed]
- Simó, C.; Fornari, T.; García-Risco, M.R.; Peña-Cearra, A.; Abecia, L.; Anguita, J.; Rodríguez, H.; García-Cañas, V. Resazurin-based high-throughput screening method for the discovery of dietary phytochemicals to target microbial transformation of l-carnitine into trimethylamine, a gut metabolite associated with cardiovascular disease. Food Funct. 2022, 13, 5640–5653. [Google Scholar] [CrossRef] [PubMed]
- Handl, J.; Nyvltova, P.; Capek, J.; Cesla, P.; Hovsepyan, A.; Avetisyan, S.; Micankova, P.; Bruckova, L.; Stankova, P.; Knotkova, K.; et al. The comparison of biological effects of bacterial and synthetic melanins in neuroblastoma cells. Food Chem. Toxicol. 2022, 168, 113355. [Google Scholar] [CrossRef]
- Uhlig, E.; Elli, G.; Nurminen, N.; Oscarsson, E.; Canaviri-Paz, P.; Burri, S.; Rohrstock, A.-M.; Rahman, M.; Alsanius, B.; Molin, G.; et al. Comparative immunomodulatory effects in mice and in human dendritic cells of five bacterial strains selected for biocontrol of leafy green vegetables. Food Chem. Toxicol. 2022, 165, 113064. [Google Scholar] [CrossRef]
- Jonaitis, T.; Lewis, E.A.; Lourens, N.; Groot, A.; Goodman, R.E.; Mitchell, D.; Karpol, A.; Tracy, B. Subchronic feeding, allergenicity, and genotoxicity safety evaluations of single strain bacterial protein. Food Chem. Toxicol. 2022, 162, 112878. [Google Scholar] [CrossRef]
- Emery, H.; Butt, T.M.; Coates, C.J. Nutraceutical intervention protects against bacterial and chemical-induced gastrotoxicity in a non-mammalian model, Galleria mellonella. Food Chem. Toxicol. 2021, 154, 112354. [Google Scholar] [CrossRef]
- Thillai Sekar, V.; Santiago, T.; Vijayan, K.; Alavandi, S.; Stalin Raj, V.; Rajan, J.; Sanjuktha, M.; Kalaimani, N. Involvement of Enterobacter cloacae in the mortality of the fish, Mugil cephalus. Lett. Appl. Microbiol. 2008, 46, 667–672. [Google Scholar] [CrossRef]
- Nair, S.G.; Lipton, A.; De los Ríos-Escalante, P.; Ibáñez-Arancibia, E. Isolation and characterization of bacterial pathogens, Pseudomonas aeruginosa and Enterobacter cloacae from the moribund fish, Etroplus maculatus. J. Mater. Environ. Sci. 2021, 12, 1332–1349. [Google Scholar]
- Troast, J.L. Antibodies against enteric bacteria in brown bullhead catfish (Ictalurus nebulosus, LeSueur) inhabiting contaminated waters. Appl. Microbiol. 1975, 30, 189–192. [Google Scholar] [CrossRef] [PubMed]
- Hansen, G.; Raa, J.; Olafsen, J. Isolation of Enterobacter agglomerans from dolphin fish, Coryphaena hippurus L. J. Fish Dis. 1990, 13, 93–96. [Google Scholar] [CrossRef]
- Geldreich, E.E.; Clarke, N.A. Bacterial pollution indicators in the intestinal tract of freshwater fish. Appl. Microbiol. 1966, 14, 429–437. [Google Scholar] [CrossRef] [PubMed]
- Abu-Elala, N.M.; Abd-Elsalam, R.M.; Younis, N.A. Streptococcosis, Lactococcosis and Enterococcosis are potential threats facing cultured Nile tilapia (Oreochomis niloticus) production. Aquac. Res. 2020, 51, 4183–4195. [Google Scholar] [CrossRef]
- Dong, J.; Zhang, L.; Zhou, S.; Xu, N.; Yang, Q.; Liu, Y.; Ai, X. Identification of a multi-resistant Enterobacter cloacae strain from diseased crayfish (Procambarus clarkii). Aquac. Rep. 2020, 17, 100405. [Google Scholar] [CrossRef]
- Austin, B.; Austin, D.A.; Munn, C. Bacterial Fish Pathogens: Disease of Farmed and Wild Fish; Springer: Berlin/Heidelberg, Germany, 2007; Volume 26. [Google Scholar]
- El-Son, M.A.; Elbahnaswy, S.; Ibrahim, I. Molecular and histopathological characterization of Photobacterium damselae in naturally and experimentally infected Nile tilapia (Oreochromis niloticus). J. Fish Dis. 2020, 43, 1505–1517. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, S.; Haldar, S. Vibrio related diseases in aquaculture and development of rapid and accurate identification methods. J. Mar. Sci. Res. Dev. S 2012, 1, 1–7. [Google Scholar]
- Austin, B.; Austin, D.A. Photobacteriaceae Representatives. In Bacterial Fish Pathogens; Springer: Berlin/Heidelberg, Germany, 2012; pp. 327–339. [Google Scholar]
- Simões, L.N.; Lombardi, D.C.; Gomide, A.; Gomes, L.C. Efficacy of clove oil as anesthetic in handling and transportation of Nile tilapia, Oreochromis niloticus (Actinopterygii: Cichlidae) juveniles. Zoologia 2011, 28, 285–290. [Google Scholar] [CrossRef]
- Boyd, C. Water Quality in Ponds for Aquaculture; Agriculture Experiment Station, Auburn University: Auburn, AL, USA, 1990. [Google Scholar]
- Singare, P.; Ferns, S. Study of toxic heavy metals in Mahim Creek of Mumbai. Int. Lett. Chem. Phys. Astron. 2014, 17, 98–106. [Google Scholar] [CrossRef]
- Shakweer, M.S.; Elshopakey, G.E.; Abdelwarith, A.A.; Younis, E.M.; Davies, S.J.; Elbahnaswy, S. Comparison of Immune Response of Litopenaeus vannamei Shrimp Naturally Infected with Vibrio Species, and after Being Fed with Florfenicol. Fishes 2023, 8, 148. [Google Scholar] [CrossRef]
- Lagacé, L.; Pitre, M.; Jacques, M.; Roy, D. Identification of the bacterial community of maple sap by using amplified ribosomal DNA (rDNA) restriction analysis and rDNA sequencing. Appl. Environ. Microbiol. 2004, 70, 2052–2060. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Balouiri, M.; Sadiki, M.; Ibnsouda, S.K. Methods for in vitro evaluating antimicrobial activity: A review. J. Pharm. Anal. 2016, 6, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Zidour, M.; Chevalier, M.; Belguesmia, Y.; Cudennec, B.; Grard, T.; Drider, D.; Souissi, S.; Flahaut, C. Isolation and characterization of bacteria colonizing Acartia tonsa copepod eggs and displaying antagonist effects against Vibrio anguillarum, Vibrio alginolyticus and other pathogenic strains. Front. Microbiol. 2017, 8, 1919. [Google Scholar] [CrossRef] [PubMed]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing; Twenty-Fifth Informational Supplement M100-S25; Clinical and Laboratory Standards Institute (CLSI): Wayne, PA, USA, 2015. [Google Scholar]
- Wedemeyer, G.A.; Yasutake, W.T. Clinical Methods for the Assessment of the Effects of Environmental Stress on Fish Health; Department of the Interior, Fish and Wildlife Service: Bailey’s Crossroads, VA, USA, 1977; Volume 89.
- El-Son, M.A.; Elshopakey, G.E.; Rezk, S.; Eldessouki, E.A.; Elbahnaswy, S. Dietary mixed Bacillus strains promoted the growth indices, enzymatic profile, intestinal immunity, and liver and intestinal histomorphology of Nile tilapia, Oreochromis niloticus. Aquac. Rep. 2022, 27, 101385. [Google Scholar] [CrossRef]
- He, S.; Atkinson, C.; Qiao, F.; Cianflone, K.; Chen, X.; Tomlinson, S. A complement-dependent balance between hepatic ischemia/reperfusion injury and liver regeneration in mice. J. Clin. Investig. 2009, 119, 2304–2316. [Google Scholar] [CrossRef] [PubMed]
- Dati, F.; Lammers, M. Immunochemical methods for determination of urinary proteins (albumin and α1-microglobulin) in kidney disease. J. Int. Fed. Clin. Chem. 1989, 1, 68–77. [Google Scholar]
- Elbahnaswy, S.; Elshopakey, G.E. Differential gene expression and immune response of Nile tilapia (Oreochromis niloticus) challenged intraperitoneally with Photobacterium damselae and Aeromonas hydrophila demonstrating immunosuppression. Aquaculture 2020, 526, 735364. [Google Scholar] [CrossRef]
- Ghareghanipoora, M.; Akbary, P.; Akhlaghi, M.; Fereidouni, M. Non-specific immune responses and immune related genes expression of rainbow trout (Oncorhynchus mykiss, walbaum) fed Zataria multiflora boiss extract. Bull. Env. Pharmacol. 2014, 3, 140–146. [Google Scholar]
- Fernandez-Botran, R.; Gorantla, V.; Sun, X.; Ren, X.; Perez-Abadia, G.; Crespo, F.A.; Oliver, R.; Orhun, H.I.; Quan, E.E.; Maldonado, C. Targeting of glycosaminoglycan-cytokine interactions as a novel therapeutic approach in allotransplantation1. Transplantation 2002, 74, 623–629. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Roberts, R.J. Fish Pathology; John Wiley & Sons: Hoboken, NJ, USA, 2012. [Google Scholar]
- Boyd, C.E.; Tucker, C.S. Pond Aquaculture Water Quality Management; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2012. [Google Scholar]
- Mustapha, S.; Mustapha, M.; Brahim, B.; Nozhal, C. Characterization of Vibrio alginolyticus Trh positive from mediterranean environment of Tamouda Bay (Morocco). World Environ. 2012, 2, 76–80. [Google Scholar] [CrossRef]
- Albert, V.; Ransangan, J. Effect of water temperature on susceptibility of culture marine fish species to vibriosis. Int. J. Res. Pure Appl. Microbiol. 2013, 3, 48–52. [Google Scholar]
- Song, Y.; Yu, P.; Li, B.; Pan, Y.; Zhang, X.; Cong, J.; Zhao, Y.; Wang, H.; Chen, L. The mosaic accessory gene structures of the SXT/R391-like integrative and conjugative elements derived from Vibrio spp. isolated from aquatic products and environment in the Yangtze River Estuary, China. BMC Microbiol. 2013, 13, 214. [Google Scholar] [CrossRef]
- El-Hak, A.; Ezzat, M.; Mohamed, G. Genotyping of Vibrio Parahemolyticus Isolated from Some Marine Fish. Suez Canal Vet. Med. J. SCVMJ 2018, 23, 91–108. [Google Scholar] [CrossRef]
- Naka, H.; Liu, M.; Actis, L.A.; Crosa, J.H. Plasmid- and chromosome-encoded siderophore anguibactin systems found in marine vibrios: Biosynthesis, transport and evolution. BioMetals 2013, 26, 537–547. [Google Scholar] [CrossRef]
- Zhang, X.-H.; Austin, B. Haemolysins in Vibrio species. J. Appl. Microbiol. 2005, 98, 1011–1019. [Google Scholar] [CrossRef]
- Osman, K.M.; Al-Maary, K.S.; Mubarak, A.S.; Dawoud, T.M.; Moussa, I.M.; Ibrahim, M.D.; Hessain, A.M.; Orabi, A.; Fawzy, N.M. Characterization and susceptibility of streptococci and enterococci isolated from Nile tilapia (Oreochromis niloticus) showing septicaemia in aquaculture and wild sites in Egypt. BMC Vet. Res. 2017, 13, 357. [Google Scholar] [CrossRef]
- Rahman, M.; Rahman, M.; Deb, S.C.; Alam, M.; Islam, M. Molecular identification of multiple antibiotic resistant fish pathogenic Enterococcus faecalis and their control by medicinal herbs. Sci. Rep. 2017, 7, 3747. [Google Scholar] [CrossRef]
- Abdel-Fattah, M.; Helmy, A. Assessment of water quality of wastewaters of Bahr El-Baqar, Bilbies and El-Qalyubia drains in east delta, Egypt for irrigation purposes. Egypt. J. Soil Sci. 2015, 55, 287–302. [Google Scholar]
- Elkiki, M.H. Environmental Impact of Water Reuse of Bahr El-Baqar Drain; Civil Engineering Department, Faculty of Engineering, Port Said University: Port Said, Egypt, 2018. [Google Scholar]
- Abdel-Razek, N.; Shagar, G.; Ayoub, H. Investigation on mass mortality and bioaccumulation of heavy metals and fish pathogen in Nile tilapia (Oreochromis niloticus) in Fayoum governorate. Abbassa Int. J. Aquac. 2016, 9, 336–355. [Google Scholar]
- Chen, M.-X.; Li, H.-Y.; Li, G.; Zheng, T.-L. Distribution of Vibrio alginolyticus-like species in Shenzhen coastal waters, China. Braz. J. Microbiol. 2011, 42, 884–896. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Aziz, M.; Eissa, A.E.; Hanna, M.; Abou Okada, M. Identifying some pathogenic Vibrio/Photobacterium species during mass mortalities of cultured Gilthead seabream (Sparus aurata) and European seabass (Dicentrarchus labrax) from some Egyptian coastal provinces. Int. J. Vet. Sci. Med. 2013, 1, 87–95. [Google Scholar] [CrossRef]
- Zahran, E.; Elbahnaswy, S.; Mamdouh, A.Z.; El-Matbouli, M. Xenosteroids in aquaculture with special consideration to Lake Manzala (Northern delta lake, Egypt): Types, sources and mechanism of action. Aquac. Res. 2021, 52, 5962–5977. [Google Scholar] [CrossRef]
- Reyad, Y.A.; Elbahnaswy, S.; Zaki, V.H. Bioaccumulation of some heavy metals in water and tissues of naturally infected Oreochromis niloticus from two polluted sites in Egypt, with reference to related oxidative stress. Egypt. J. Aquat. Biol. Fish. 2021, 25, 139–157. [Google Scholar]
- Kubrak, O.I.; Husak, V.V.; Rovenko, B.M.; Poigner, H.; Mazepa, M.A.; Kriews, M.; Abele, D.; Lushchak, V.I. Tissue specificity in nickel uptake and induction of oxidative stress in kidney and spleen of goldfish Carassius auratus, exposed to waterborne nickel. Aquat. Toxicol. 2012, 118–119, 88–96. [Google Scholar] [CrossRef] [PubMed]
- Gehringer, D.B.; Finkelstein, M.E.; Coale, K.H.; Stephenson, M.; Geller, J.B. Assessing Mercury Exposure and Biomarkers in Largemouth Bass (Micropterus salmoides) from a Contaminated River System in California. Arch. Environ. Contam. Toxicol. 2013, 64, 484–493. [Google Scholar] [CrossRef]
- Anderson, J.E.; Sparling, P.F.; Cornelissen, C.N. Gonococcal transferrin-binding protein 2 facilitates but is not essential for transferrin utilization. J. Bacteriol. 1994, 176, 3162–3170. [Google Scholar] [CrossRef]
- Bullen, J. The significance of iron in infection. Rev. Infect. Dis. 1981, 3, 1127–1138. [Google Scholar] [CrossRef]
- Mushlake, K.; Nakai, T.; Muroga, K. Lowered phagocytosis in the blood of eels exposed to copper. Fish Pathol. 1985, 20, 49–53. [Google Scholar] [CrossRef]
- El-Sharaby, S.; Abd-Elgaber, M.; Tarabees, R.; Khalil, R.; Ali, M.; El-Ballal, S. Bacteriological and histopathological studies on Vibrio species isolated from naturally infected freshwater fish in Delta Region, Egypt. Adv. Anim. Vet. Sci. 2018, 6, 17–26. [Google Scholar]
- Buller, N.B. Bacteria from Fish and Other Aquatic Animals: A Practical Identification Manual; Cabi: Wallingford, UK, 2004. [Google Scholar]
- Khalil, S.A.; Abou-Akkada, A.S.; El-Hoshy, S. Molecular studies on vibrio species isolated from imported frozen fish. Glob. Vet. 2014, 12, 782–789. [Google Scholar]
- Thompson, F.; Gevers, D.; Thompson, C.; Dawyndt, P.; Naser, S.; Hoste, B.; Munn, C.; Swings, J. Phylogeny and molecular identification of vibrios on the basis of multilocus sequence analysis. Appl. Environ. Microbiol. 2005, 71, 5107–5115. [Google Scholar] [CrossRef] [PubMed]
- Ransangan, J.; Lal, T.M.; Al-Harbi, A.H. Characterization and experimental infection of Vibrio harveyi isolated from diseased Asian seabass (Lates calcarifer). Malays. J. Microbiol. 2012, 8, 104–115. [Google Scholar] [CrossRef]
- Recknagel, R.O.; Glende, E.A., Jr.; Dolak, J.A.; Waller, R.L. Mechanisms of carbon tetrachloride toxicity. Pharmacol. Ther. 1989, 43, 139–154. [Google Scholar] [CrossRef]
- Pérez-Rostro, C.I.; Racotta, I.S.; Ibarra, A.M. Decreased genetic variation in metabolic variables of Litopenaeus vannamei shrimp after exposure to acute hypoxia. J. Exp. Mar. Biol. Ecol. 2004, 302, 189–200. [Google Scholar] [CrossRef]
- Hal, A.M.; Manal, I. Effect of Nigella sativa oil and ciprofloxacin against bacterial infection on gene expression in Nile tilapia (Oreochromis niloticus) blood. Aquaculture 2021, 532, 736071. [Google Scholar] [CrossRef]
- Souza, C.F.; Baldissera, M.D.; Vaucher, R.A.; Lopes, L.Q.; Vizzotto, B.S.; Raffin, R.P.; Santos, R.C.; da Veiga, M.L.; da Rocha, M.I.U.; Stefani, L.M. In vivo bactericidal effect of Melaleuca alternifolia essential oil against Aeromonas hydrophila: Silver catfish (Rhamdia quelen) as an experimental model. Microb. Pathog. 2016, 98, 82–87. [Google Scholar] [CrossRef]
- Chen, C.-Y.; Wooster, G.A.; Bowser, P.R. Comparative blood chemistry and histopathology of tilapia infected with Vibrio vulnificus or Streptococcus iniae or exposed to carbon tetrachloride, gentamicin, or copper sulfate. Aquaculture 2004, 239, 421–443. [Google Scholar] [CrossRef]
- Qi, Z.; Zhang, Q.; Wang, Z.; Ma, T.; Zhou, J.; Holland, J.W.; Gao, Q. Transcriptome analysis of the endangered Chinese giant salamander (Andrias davidianus): Immune modulation in response to Aeromonas hydrophila infection. Vet. Immunol. Immunopathol. 2016, 169, 85–95. [Google Scholar] [CrossRef]
- Tran, N.T.; Gao, Z.-X.; Zhao, H.-H.; Yi, S.-K.; Chen, B.-X.; Zhao, Y.-H.; Lin, L.; Liu, X.-Q.; Wang, W.-M. Transcriptome analysis and microsatellite discovery in the blunt snout bream (Megalobrama amblycephala) after challenge with Aeromonas hydrophila. Fish Shellfish Immunol. 2015, 45, 72–82. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Zhang, Y.; Wen, Y.; Ji, W.; Zhou, Y.; Ji, Y.; Liu, X.; Wang, W.; Asim, M.; Liang, X. Analysis of the transcriptomic profilings of Mandarin fish (Siniperca chuatsi) infected with Flavobacterium columnare with an emphasis on immune responses. Fish Shellfish Immunol. 2015, 43, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.; Mu, L.; Fu, S.; Wu, L.; Han, K.; Wu, H.; Bian, X.; Wei, X.; Guo, Z.; Wang, A. Expression and characterization of Nile tilapia (Oreochromis niloticus) secretory and membrane-bound IgM in response to bacterial infection. Aquaculture 2019, 508, 214–222. [Google Scholar] [CrossRef]
- Pepys, M.B.; Hirschfield, G.M. C-reactive protein: A critical update. J. Clin. Investig. 2003, 111, 1805–1812. [Google Scholar] [CrossRef] [PubMed]
- MacCarthy, E.M.; Burns, I.; Irnazarow, I.; Polwart, A.; Greenhough, T.J.; Shrive, A.K.; Hoole, D. Serum CRP-like protein profile in common carp Cyprinus carpio challenged with Aeromonas hydrophila and Escherichia coli lipopolysaccharide. Dev. Comp. Immunol. 2008, 32, 1281–1289. [Google Scholar] [CrossRef]
- Hoover, G.J.; El-Mowafi, A.; Simko, E.; Kocal, T.E.; Ferguson, H.W.; Hayes, M.A. Plasma proteins of rainbow trout (Oncorhynchus mykiss) isolated by binding to lipopolysaccharide from Aeromonas salmonicida. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 1998, 120, 559–569. [Google Scholar] [CrossRef]
- Barroso, C.; Ozorio, R.O.; Afonso, A.; Moraes, J.R.; Costas, B. Immune responses and gut morphology in Senegalese sole (Solea senegalensis) fed dietary probiotic supplementation and following exposure to Photobacterium damselae subsp. piscicida. Aquac. Res. 2016, 47, 951–960. [Google Scholar] [CrossRef]
- Costas, B.; Rêgo, P.; Simões, I.; Marques, J.; Castro-Cunha, M.; Afonso, A. Cellular and humoral immune responses of Senegalese sole, Solea senegalensis (Kaup), following challenge with two Photobacterium damselae subsp. piscicida strains from different geographical origins. J. Fish Dis. 2013, 36, 543–553. [Google Scholar]
- Xia, H.; Tang, Y.; Lu, F.; Luo, Y.; Yang, P.; Wang, W.; Jiang, J.; Li, N.; Han, Q.; Liu, F. The effect of Aeromonas hydrophila infection on the non-specific immunity of blunt snout bream (Megalobrama amblycephala). Cent. Eur. J. Immunol. 2017, 42, 239. [Google Scholar] [CrossRef]
- Núñez-Díaz, J.; Fumanal, M.; Mancera, J.; Moriñigo, M.; Balebona, M. Two routes of infection with Photobacterium damselae subsp. piscicida are effective in the modulation of the transcription of immune related genes in Solea senegalensis. Vet. Immunol. Immunopathol. 2016, 179, 8–17. [Google Scholar] [CrossRef]
- Sahoo, P.; Mahapatra, K.D.; Saha, J.; Barat, A.; Sahoo, M.; Mohanty, B.; Gjerde, B.; Ødegård, J.; Rye, M.; Salte, R. Family association between immune parameters and resistance to Aeromonas hydrophila infection in the Indian major carp, Labeo rohita. Fish Shellfish Immunol. 2008, 25, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Vina, J.; Borras, C.; Gomez-Cabrera, M.C. A free radical theory of frailty. Free Radic. Biol. Med. 2018, 124, 358–363. [Google Scholar] [CrossRef]
- Baldissera, M.D.; Souza, C.F.; Parmeggiani, B.; Leipnitz, G.; Verdi, C.M.; Santos, R.V.; Stefani, L.M.; Baldisserotto, B. The disturbance of antioxidant/oxidant balance in fish experimentally infected by Aeromonas caviae: Relationship with disease pathophysiology. Microb. Pathog. 2018, 122, 53–57. [Google Scholar] [CrossRef] [PubMed]
- Souza, C.F.; Baldissera, M.D.; Guarda, N.S.; Bollick, Y.S.; Moresco, R.N.; Brusque, I.C.M.; Santos, R.C.; Baldisserotto, B. Melaleuca alternifolia essential oil nanoparticles ameliorate the hepatic antioxidant/oxidant status of silver catfish experimentally infected with Pseudomonas Aeruginosa. Microb. Pathog. 2017, 108, 61–65. [Google Scholar] [CrossRef] [PubMed]
- Souza, C.D.F.; Baldissera, M.D.; Verdi, C.M.; Santos, R.C.; Da Rocha, M.I.U.; da Veiga, M.L.; da Silva, A.S.; Baldisserotto, B. Oxidative stress and antioxidant responses in Nile tilapia Oreochromis niloticus experimentally infected by Providencia rettgeri. Microb. Pathog. 2019, 131, 164–169. [Google Scholar] [CrossRef]
- Kurhalyuk, N.; Tkachenko, H. Induction of oxidative stress and antioxidant defenses in the livers of sea trout, Salmo trutta L., with ulcerative dermal necrosis. Fish. Aquat. Life 2011, 19, 229–240. [Google Scholar] [CrossRef]
- Ganesan, S.; Baskaran, B.; Raj, M.; Mandal, A.; Shanmugam, K.; Subramanian, P.; Tabarsa, M.; You, S.G.; Narayanasamy Marimuthu, P. Vibriosis Incidents in Marine Finfish Farms: Prevalence, Diagnosis of Pathogens using 16S rRNA, Histopathology, and In Vitro Antibacterial Evaluation Against Isolated Vibrio spp. using Antibiotics and Probiotics. Thalass. Int. J. Mar. Sci. 2021, 38, 385–399. [Google Scholar] [CrossRef]
- Krupesha Sharma, S.R.; Rathore, G.; Verma, D.K.; Sadhu, N.; Philipose, K.K. Vibrio alginolyticus infection in A sian seabass (Lates calcarifer, Bloch) reared in open sea floating cages in India. Aquac. Res. 2012, 44, 86–92. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).