
Changes in Abundance and Distribution of the Sea Pen, Funiculina quadrangularis, in the Central Adriatic Sea (Mediterranean Basin) in Response to Variations in Trawling Intensity
 , , , , and
, , , , and
Abstract

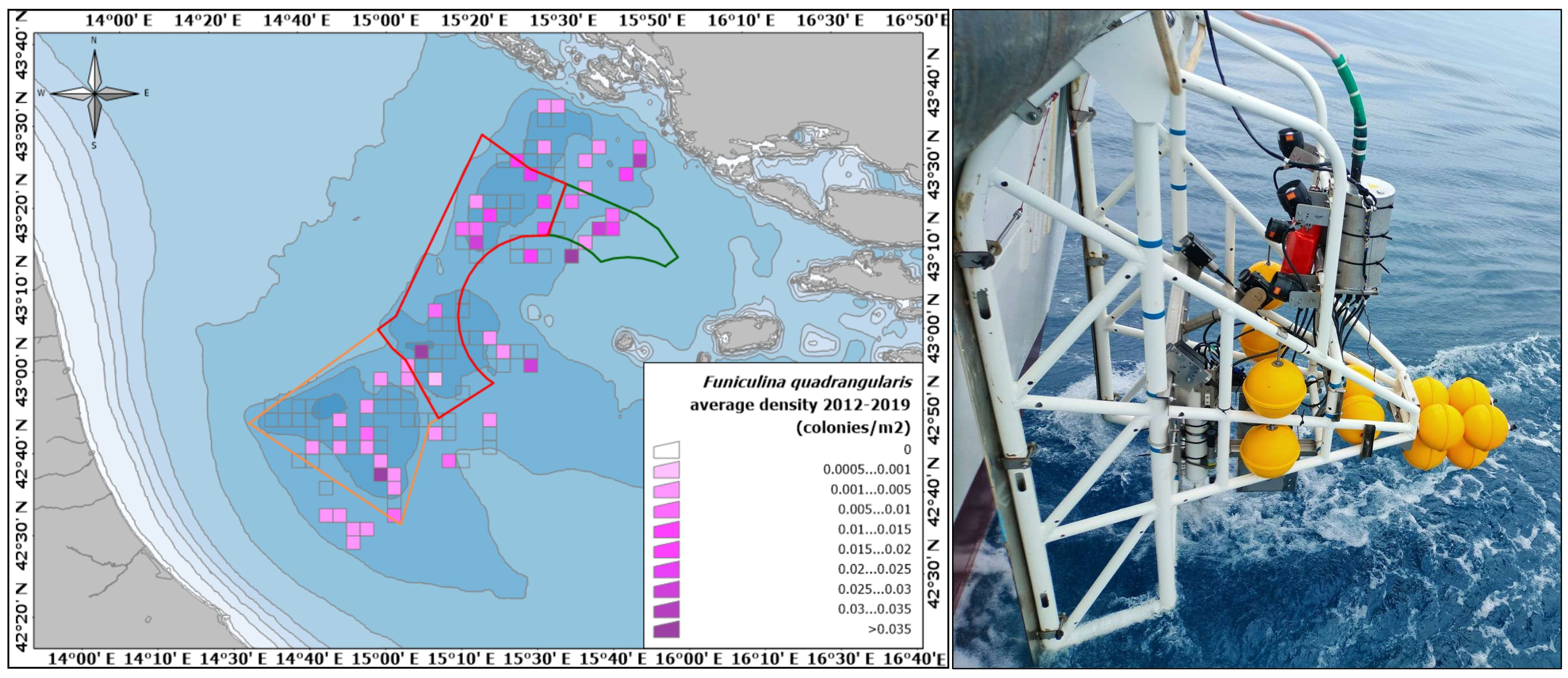{kind=link}
{kind=link}
{kind=link}
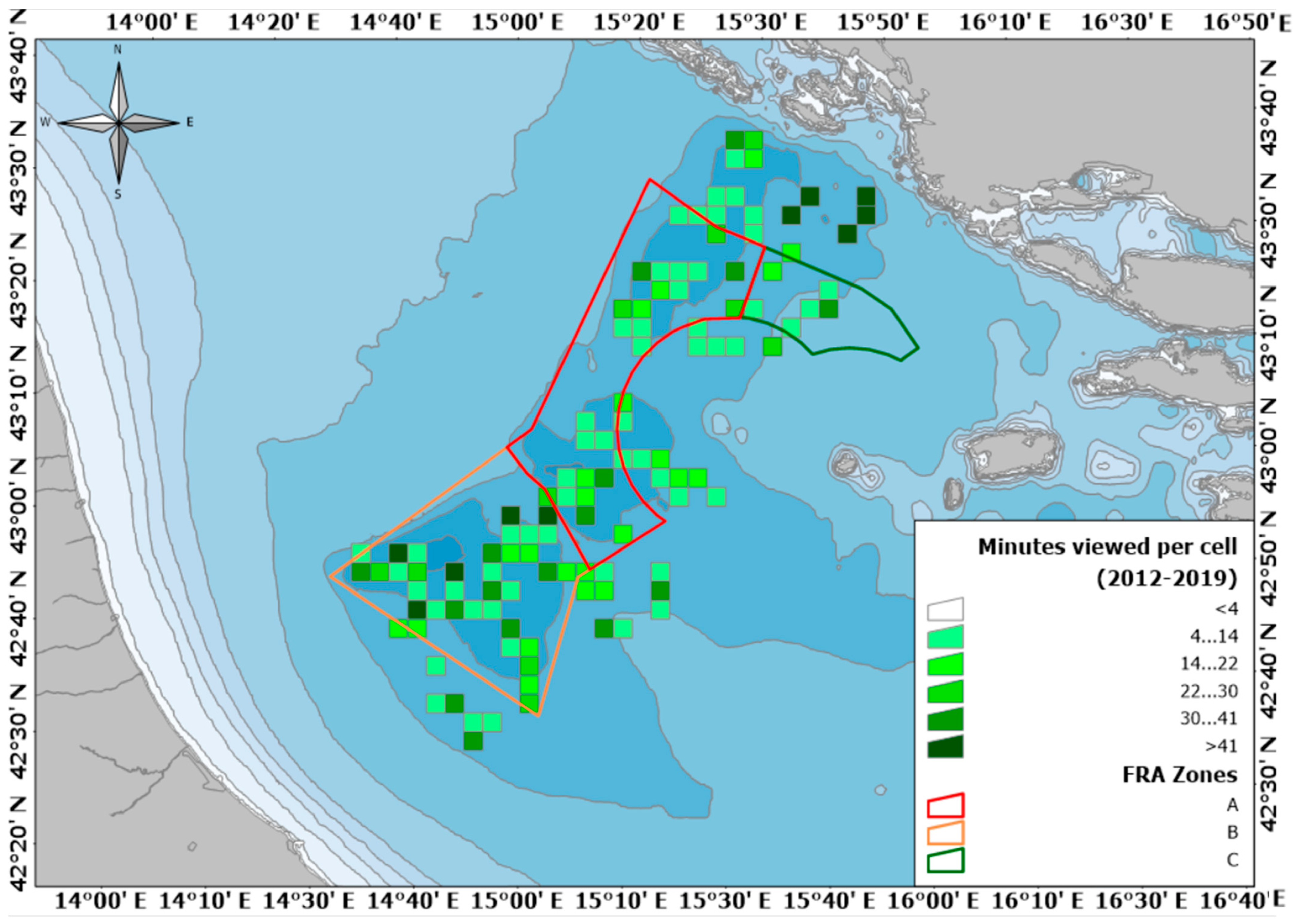{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Sample Collection and Video Analysis
2.3. Data Analysis
3. Results
3.1. Spatial Distribution of Colonies and Density by Period for Each Zone (“A”,“B”, and “C”)
3.1.1. Area “A” across the Periods
3.1.2. Area “B” across the Periods
3.1.3. Area “C” across the Periods
3.2. Persistence per Cell
4. Discussion
4.1. Effects of Different Protection Levels on Funiculina quadrangularis
4.2. Ecological Aspects and Colonies’ Persistence
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Clark, M.R.; Althaus, F.; Schlacher, T.A.; Williams, A.; Bowden, D.A.; Rowden, A.A. The Impacts of Deep-Sea Fisheries on Benthic Communities: A Review. ICES J. Mar. Sci. 2016, 73, i51–i69. [Google Scholar] [CrossRef]
- Farella, G.; Tassetti, A.N.; Menegon, S.; Bocci, M.; Ferrà, C.; Grati, F.; Fadini, A.; Giovanardi, O.; Fabi, G.; Raicevich, S.; et al. Ecosystem-Based MSP for Enhanced Fisheries Sustainability: An Example from the Northern Adriatic (Chioggia-Venice and Rovigo, Italy). Sustainability 2021, 13, 1211. [Google Scholar] [CrossRef]
- Van Denderen, P.D.; Hintzen, N.T.; Van Kooten, T.; Rijnsdorp, A.D. Temporal Aggregation of Bottom Trawling and Its Implication for the Impact on the Benthic Ecosystem. ICES J. Mar. Sci. 2015, 72, 952–961. [Google Scholar] [CrossRef]
- Pusceddu, A.; Bianchelli, S.; Martín, J.; Puig, P.; Palanques, A.; Masqué, P.; Danovaro, R. Chronic and Intensive Bottom Trawling Impairs Deep-Sea Biodiversity and Ecosystem Functioning. Proc. Natl. Acad. Sci. USA 2014, 111, 8861–8866. [Google Scholar] [CrossRef] [PubMed]
- Pusceddu, A.; Bianchelli, S.; Danovaro, R. Quantity and Biochemical Composition of Particulate Organic Matter in a Highly Trawled Area (Thermaikos Gulf, Eastern Mediterranean Sea). Adv. Oceanogr. Limnol. 2015, 6, 21–32. [Google Scholar] [CrossRef]
- Pommer, C.D.; Olesen, M.; Jhansen, Ø.L.S. Impact and Distribution of Bottom Trawl Fishing on Mud-Bottom Communities in the Kattegat. Mar. Ecol. Prog. Ser. 2016, 548, 47–60. [Google Scholar] [CrossRef]
- Jennings, S.; Pinnegar, J.K.; Polunin, N.V.C.; Warr, K.J. Impacts of Trawling Disturbance on the Trophic Structure of Benthic Invertebrate Communities. Mar. Ecol. Prog. Ser. 2001, 213, 127–142. [Google Scholar] [CrossRef]
- Hutchings, P. Review of the Effects of Trawling on Macrobenthic Epifaunal Communities. Mar. Freshw. Res. 1990, 41, 111–120. [Google Scholar] [CrossRef]
- Kaiser, M.J.; Collie, J.S.; Hall, S.J.; Jennings, S.; Poiner, I.R. Modification of Marine Habitats by Trawling Activities: Prognosis and Solutions. Fish Fish. 2002, 3, 114–136. [Google Scholar] [CrossRef]
- Lauria, V.; Garofalo, G.; Fiorentino, F.; Massi, D.; Milisenda, G.; Piraino, S.; Russo, T.; Gristina, M. Species Distribution Models of Two Critically Endangered Deep-Sea Octocorals Reveal Fishing Impacts on Vulnerable Marine Ecosystems in Central Mediterranean Sea. Sci. Rep. 2017, 7, 8049. [Google Scholar] [CrossRef]
- Rodríguez-Rodríguez, D.; Rodríguez, J.; Abdul Malak, D. Development and Testing of a New Framework for Rapidly Assessing Legal and Managerial Protection Afforded by Marine Protected Areas: Mediterranean Sea Case Study. J. Environ. Manage. 2016, 167, 29–37. [Google Scholar] [CrossRef]
- Bastardie, F.; Angelini, S.; Bolognini, L.; Fuga, F.; Manfredi, C.; Martinelli, M.; Nielsen, J.R.; Santojanni, A.; Scarcella, G.; Grati, F. Spatial Planning for Fisheries in the Northern Adriatic: Working toward Viable and Sustainable Fishing. Ecosphere 2017, 8, e01696. [Google Scholar] [CrossRef]
- Serrano, A.; de la Torriente, A.; Punzón, A.; Blanco, M.; Bellas, J.; Durán-Muñoz, P.; Murillo, F.J.; Sacau, M.; García-Alegre, A.; Antolínez, A.; et al. Sentinels of Seabed (SoS) Indicator: Assessing Benthic Habitats Condition Using Typical and Sensitive Species. Ecol. Indic. 2022, 140, 108979. [Google Scholar] [CrossRef]
- Coll, M.; Piroddi, C.; Steenbeek, J.; Kaschner, K.; Lasram, F.B.R.; Aguzzi, J.; Ballesteros, E.; Bianchi, C.N.; Corbera, J.; Dailianis, T.; et al. The Biodiversity of the Mediterranean Sea: Estimates, Patterns, and Threats. PLoS ONE 2010, 5, e11842. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.; Papadopoulou, N.; Sevastou, K.; Franco, A.; Teixeira, H.; Piroddi, C.; Katsanevakis, S.; Fürhaupter, K.; Beauchard, O.; Cochrane, S.; et al. Report on Identification of Keystone Species and Processes across Regional Seas. 2014, pp. 1–103. Available online: https://hal.science/hal-01790558 (accessed on 27 April 2023).
- Degnbol, P. Indicators as a Means of Communicating Knowledge. ICES J. Mar. Sci. 2005, 62, 606–611. [Google Scholar] [CrossRef]
- European Commission. Commission Decision (EU) 2017/848 of 17 May 2017-Laying down Criteria and Methodological Standards on Good Environmental Status of Marine Waters and Specifications and Standardized Methods for Monitoring and Assessment; European Commission: Brussels, Belgium, 2017; Official Journal L 125; pp. 43–74. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX%3A32017D0848 (accessed on 27 April 2023).
- Burgos, J.M.; Buhl-Mortensen, L.; Buhl-Mortensen, P.; Ólafsdóttir, S.H.; Steingrund, P.; Ragnarsson, S.; Skagseth, Ø. Predicting the Distribution of Indicator Taxa of Vulnerable Marine Ecosystems in the Arctic and Sub-Arctic Waters of the Nordic Seas. Front. Mar. Sci. 2020, 7, 131. [Google Scholar] [CrossRef]
- Williams, G.C. The Global Diversity of Sea Pens (Cnidaria: Octocorallia: Pennatulacea). PLoS ONE 2011, 6, e22747. [Google Scholar] [CrossRef]
- Cogswell, A.; Kenchington, E.; Lirette, C.; Murillo, F.J.; Campanis, G.; Campbell, N.; Ollerhead, N. Layers Utilized by an ArcGIS Model to Approximate Commercial Coral and Sponge By-Catch in the NAFO Regulatory Area. 2011. Available online: https://archive.nafo.int/open/sc/2011/scr11-072.pdf (accessed on 26 April 2023).
- United Nations. Resolution 70/75: Sustainable Fisheries, Including through the 1995 Agreement for the Implementation of the Provisions of the United Nations Convention on the Law of the Sea of 10 December 1982 Relating to the Conservation and Management of Straddling Fiss; United Nations: New York, NY, USA, 2016; p. 35. Available online: https://sdgs.un.org/documents/ares7075-sustainable-fisheries-including-thr-23183 (accessed on 28 April 2023).
- Hughes, D.J. Sea Pens and Burrowing Megafauna: An Overview of Dynamics and Sensitivity Characteristics for Conservation Management of Marine SACs. Science 1998, 3, 1–105. [Google Scholar]
- Felder, D.L.; Camp, D.K. Gulf of Mexico: Origin, Waters, and Biota. 2009, Volume 1. Available online: https://decapoda.nhm.org/pdfs/31409/31409.pdf (accessed on 25 April 2023).
- Wright, E.P.; Kemp, K.; Rogers, A.D.; Yesson, C. Genetic Structure of the Tall Sea Pen Funiculina quadrangularis in NW Scottish Sea Lochs. Mar. Ecol. 2015, 36, 659–667. [Google Scholar] [CrossRef]
- Ardizzone, G.D. An Introduction to Sensitive and Essential Fish Habitats Identification and Protection in the Mediterranean Sea; STECF: Rome, Italy, 2006; p. 63. [Google Scholar]
- Greathead, C.F.; Donnan, D.W.; Mair, J.M.; Saunders, G.R. The Sea Pens Virgularia mirabilis, Pennatula phosphorea and Funiculina quadrangularis: Distribution and Conservation Issues in Scottish Waters. J. Mar. Biol. Assoc. UK 2007, 87, 1095–1103. [Google Scholar] [CrossRef]
- Rogers, A.D.; Gianni, M. The Implementation of UNGA Resolutions 61/105 and 64/72 in the Management of Deep-Sea Fisheries on the High Seas. 2010. Available online: http://www.stateoftheocean.org/pdfs/61105-Implemention-finalreport.pdf (accessed on 18 April 2023).
- FAO; GFCM. Fishery and Aquaculture Statistics. GFCM Capture Production 1970-2020 (FishStatJ); FAO Fisheries and Aquaculture Division: Rome, Italy, 2022; Available online: https://www.fao.org/fishery/en/topic/166235?lang=en (accessed on 10 April 2023).
- International Union for Conservation of Nature. The IUCN Red List of Threatened Species; IUCN Version 2022-2; IUCN: Gland, Switzerland, 2022; Available online: https://nc.iucnredlist.org/redlist/content/attachment_files/2022-2_RL_Stats_Table_1a.pdf (accessed on 25 April 2023).
- Bastari, A.; Pica, D.; Ferretti, F.; Micheli, F.; Cerrano, C. Sea Pens in the Mediterranean Sea: Habitat Suitability and Opportunities for Ecosystem Recovery. ICES J. Mar. Sci. 2018, 75, 1722–1732. [Google Scholar] [CrossRef]
- Mačić, V.; Đorđević, N.; Đurović, M.; Petović, S.; Russo, T. Improving Knowledge of Funiculina quadrangularis and Vulnerable Marine Ecosystems in the South Adriatic. Mediterr. Mar. Sci. 2022, 23, 805–816. [Google Scholar] [CrossRef]
- Edwards, D.C.B.; Moore, C.G. Reproduction in the Sea Pen Funiculina quadrangularis (Anthozoa: Pennatulacea) from the West Coast of Scotland. Estuar. Coast. Shelf Sci. 2009, 82, 161–168. [Google Scholar] [CrossRef]
- Downie, A.L.; Noble-James, T.; Chaverra, A.; Howell, K.L. Predicting Sea Pen (Pennatulacea) Distribution on the UK Continental Shelf: Evidence of Range Modification by Benthic Trawling. Mar. Ecol. Prog. Ser. 2021, 670, 75–91. [Google Scholar] [CrossRef]
- Martinelli, M.; Morello, E.B.; Isajlović, I.; Belardinelli, A.; Lucchetti, A.; Santojanni, A.; Atkinson, R.J.A.; Vrgoč, N.; Arneri, E. Towed Underwater Television towards the Quantification of Norway Lobster, Squat Lobsters and Sea Pens in the Adriatic Sea. Acta Adriat. 2013, 54, 3–12. [Google Scholar]
- Miatta, M.; Snelgrove, P.V.R. Sea Pens as Indicators of Macrofaunal Communities in Deep-Sea Sediments: Evidence from the Laurentian Channel Marine Protected Area. Deep. Res. Part I Oceanogr. Res. Pap. 2022, 182, 103702. [Google Scholar] [CrossRef]
- Vigo, M.; Navarro, J.; Aguzzi, J.; Bahamon, N.; García, J.A.; Rotllant, G.; Recasens, L.; Company, J.B. ROV-Based Monitoring of Passive Ecological Recovery in a Deep-Sea No-Take Fishery Reserve. Sci. Total Environ. 2023, 883, 163339. [Google Scholar] [CrossRef]
- ICES. Working Group on Nephrops Surveys (WGNEPS; Outputs from 2021). ICES Sci. Rep. 2022, 4, 183. [Google Scholar] [CrossRef]
- OSPAR. Meeting of the Intersessional Correspondence Group on the Implementation Follow up of Measures for the Protection and Conservation of Species and Habitats (ICG-POSH), Paris, France, 9–11 October 2018. Roadmap for the Implementation of Collective Actions W. 2018, pp. 1–8. Available online: https://www.ospar.org/site/assets/files/38906/0205a5_actionsheet13_mpashabitats.pdf (accessed on 2 May 2023).
- EMODnet; EMODnet Bathymetry Consortium. EMODnet Digital Bathymetry (DTM), Ifremer; EMODnet: Oostende, Belgium, 2016. [Google Scholar] [CrossRef]
- Russo, T.; Morello, E.B.; Parisi, A.; Scarcella, G.; Angelini, S.; Labanchi, L.; Martinelli, M.; D’Andrea, L.; Santojanni, A.; Arneri, E.; et al. A Model Combining Landings and VMS Data to Estimate Landings by Fishing Ground and Harbor. Fish. Res. 2018, 199, 218–230. [Google Scholar] [CrossRef]
- Marini, M.; Maselli, V.; Campanelli, A.; Foglini, F.; Grilli, F. Role of the Mid-Adriatic Deep in Dense Water Interception and Modification. Mar. Geol. 2016, 375, 5–14. [Google Scholar] [CrossRef]
- Taviani, M.; Angeletti, L.; Beuck, L.; Campiani, E.; Canese, S.; Foglini, F.; Freiwald, A.; Montagna, P.; Trincardi, F. Reprint of “On and off the Beaten Track: Megafaunal Sessile Life and Adriatic Cascading Processes”. Mar. Geol. 2016, 375, 146–160. [Google Scholar] [CrossRef]
- Jennings, S.; Kaiser, M.J. The Effects of Fishing on Marine Ecosystems. Adv. Mar. Biol. 1998, 34, 201–212. [Google Scholar] [CrossRef]
- Lucey, S.M.; Nye, J.A. Shifting Species Assemblages in the Northeast US Continental Shelf Large Marine Ecosystem. Mar. Ecol. Prog. Ser. 2010, 415, 23–33. [Google Scholar] [CrossRef]
- Albouy, C.; Guilhaumon, F.; Araújo, M.B.; Mouillot, D.; Leprieur, F. Combining Projected Changes in Species Richness and Composition Reveals Climate Change Impacts on Coastal Mediterranean Fish Assemblages. Glob. Chang. Biol. 2012, 18, 2995–3003. [Google Scholar] [CrossRef] [PubMed]
- Hillebrand, H.; Soininen, J.; Snoeijs, P. Warming Leads to Higher Species Turnover in a Coastal Ecosystem. Glob. Chang. Biol. 2010, 16, 1181–1193. [Google Scholar] [CrossRef]
- Lavergne, S.; Mouquet, N.; Thuiller, W.; Ronce, O. Biodiversity and Climate Change: Integrating Evolutionary and Ecological Responses of Species and Communities. Annu. Rev. Ecol. Evol. Syst. 2010, 41, 321–350. [Google Scholar] [CrossRef]
- GFCM. Recommendation GFCM/41/2017/3 on the Establishment of a Fisheries Restricted Area in the Jabuka/Pomo Pit in the Adriatic Sea; GFCM: Rome, Italy, 2017. [Google Scholar]
- Zupanovic, S.; Jardas, I. A Contribution to the Study of Biology and Population Dynamics of the Adriatic Hake, Merluccius merluccius (L.). Acta Adriat. 1986, 27, 97–146. [Google Scholar]
- Angelini, S. Towards an Ecosystem Approach to Fisheries for Nephrops norvegicus and Merluccius merluccius Inhabiting the Central Adriatic Sea. 2016. Available online: http://amsdottorato.unibo.it/7380/1/Angelini_Silvia_tesi.pdf (accessed on 3 May 2023).
- Angelini, S.; Martinelli, M.; Santojanni, A.; Colella, S. Biological Evidence of the Presence of Different Subpopulations of Norway Lobster (Nephrops norvegicus) in the Adriatic Sea (Central Mediterranean Sea). Fish. Res. 2020, 221, 105365. [Google Scholar] [CrossRef]
- Colloca, F.; Mastrantonio, G.; Lasinio, G.J.; Ligas, A.; Sartor, P. Parapenaeus longirostris (Lucas, 1846) an Early Warning Indicator Species of Global Warming in the Central Mediterranean Sea. J. Mar. Syst. 2014, 138, 29–39. [Google Scholar] [CrossRef]
- Martinelli, M.; Angelini, S.; Belardinelli, A.; Caccamo, G.; Cacciamani, R.; Calì, F.; Canduci, G.; Chiarini, M.; Croci, C.; Domenichetti, F.; et al. Accordo Tra MIPAAF e CNR-IRBIM Ancona in Merito Alla Proposta Progettuale Relativa Alle Attività Di Monitoraggio Periodico Delle Fosse Di Pomo e All’attuazione Di Misure Che, Nel Rispetto Dei Piani Di Gestione, Comportino Il Mantenimento Delle Condizioni Ambientali Idonee Alla vita e All’accrescimento dei Molluschi Bivalvi, Ponendo in Essere Misure Supplementari Tese a Proteggere le Diverse Fasi del Ciclo Biologico Delle Specie Interessate (CUP J41F19000080001)-Parte Monitoraggio Fosse di Pomo periodo 2019-2020; Secondo interim report-Luglio 2020; CNR-IRBIM: Ancona, Italy, 2020. [Google Scholar]
- Froglia, C.; Gramitto, M.; Martinelli, M.; Betulla, M. Long Term Changes in the Decapod Crustaceans Assemblage in the Western Meso-Adriatic Depression (Pomo Pit). Crustac. Soc. Mid-Year 2017, 1, 19–22. [Google Scholar]
- Chiarini, M.; Guicciardi, S.; Zacchetti, L.; Domenichetti, F.; Canduci, G.; Angelini, S.; Belardinelli, A.; Croci, C.; Giuliani, G.; Scarpini, P.; et al. Looking for a Simple Assessment Tool for a Complex Task: Short-Term Evaluation of Changes in Fisheries Management Measures in the Pomo/Jabuka Pits Area (Central Adriatic Sea). Sustainability 2022, 14, 7742. [Google Scholar] [CrossRef]
- GFCM. GFCM-SAC Report of the Working Group on Vulnerable Marine Ecosystems and Essential Fish Habitats (WGVME-EFH); FAO: Rome, Italy, 2023; p. 41. [Google Scholar]
- Chiarini, M.; Guicciardi, S.; Angelini, S.; Tuck, I.D.; Grilli, F.; Penna, P.; Domenichetti, F.; Canduci, G.; Belardinelli, A.; Santojanni, A.; et al. Accounting for Environmental and Fishery Management Factors When Standardizing CPUE Data from a Scientific Survey: A Case Study for Nephrops norvegicus in the Pomo Pits Area (Central Adriatic Sea). PLoS ONE 2022, 17, e0270703. [Google Scholar] [CrossRef] [PubMed]
- EU. Regulation (EU) 2019/982 of the European Parliament and of the Council of 5 June 2019 Amending Regulation (EU) No 1343/2011 on Certain Provisions for Fishing in the GFCM (General Fisheries Commission for the Mediterranean) Agreement Area; 2019, Official Journal L 164, pp. 1–22. Available online: http://data.europa.eu/eli/reg/2019/982/oj (accessed on 5 May 2023).
- GFCM. Recommendation GFCM/44/2021/2 on the Establishment of a Fisheries Restricted Area in the Jabuka/Pomo Pit in the Adriatic Sea (Geographical Subarea 17), Amending Recommendation; GFCM: Rome, Italy, 2021. [Google Scholar]
- Martinelli, M.; Belardinelli, A.; Guicciardi, S.; Penna, P.; Domenichetti, F.; Croci, C.; Angelini, S.; Medvesek, D.; Froglia, C.; Scarpini, P. Support the Monitoring of Fisheries and Fisheries Resources in the Adriatic Sea” Report of Task 2 “To Perform the Appraisal of Nephrops Norvegicus in the Central Adriatic Sea (GFCM GSA 17) through Underwater Television Surveys; ISMAR: Ancona, Italy, 2017. [Google Scholar]
- Martinelli, M.; Belardinelli, A.; Guicciardi, S.; Penna, P.; Domenichetti, F.; Croci, C.; Angelini, S.; Medvesek, D.; Scarpini, P.; Micucci, D.; et al. SP2_LI1_WP1_UO05_D01-Rapporto Della Campagna 2015 (Ex SP2_WP1_AZ3_UO05_D03-Report 3° UWTV Survey–RITMARE)-RITMARE La Ricerca ITaliana per Il MARE; ISMAR: Ancona, Italy, 2016. [Google Scholar]
- Martinelli, M.; Angelini, S.; Belardinelli, A.; Chiarini, M.; Croci, C.; Domenichetti, F.; Guicciardi, S.; Scarpini, P.; Santojanni, A.; Lorenzo, Z. Final report Modulo 6. Monitoraggio Fosse Di Pomo Periodo 2017-2018 (Esteso Primavera 2019) Convenzione Tra MIPAAFT e CNR-ISMAR Ancona per Uno Studio Propedeutico Al Rinnovo Dell’affidamento Della Gestione Della Pesca Dei Molluschi Bivalvi Ai Consorzi Di Gestione–CUP J53C17000540001; IRBIM-CNR: Ancona, Italy, 2019. [Google Scholar]
- Penna, P.; Grilli, F.; Belardinelli, A.; Domenichetti, F.; Scarpini, P.; Martinelli, M. Pomo Pits Pressure/Temperature/Salinity/Oxygen Profiles Spring Dataset 2012–2021. Seanoe 2022. [Google Scholar] [CrossRef]
- ICES. Working Group on Nephrops Surveys (WGNEPS; Outputs from 2019). ICES Sci. Rep. 2020, 2, 85. [Google Scholar]
- Aguzzi, J.; Bahamon, N.; Doyle, J.; Lordan, C.; Tuck, I.D.; Chiarini, M.; Martinelli, M.; Company, J.B. Burrow Emergence Rhythms of Nephrops norvegicus by UWTV and Surveying Biases. Sci. Rep. 2021, 11, 5797. [Google Scholar] [CrossRef]
- Aguzzi, J.; Chatzievangelou, D.; Robinson, N.J.; Bahamon, N.; Berry, A.; Carreras, M.; Company, J.B.; Costa, C.; del Rio Fernandez, J.; Falahzadeh, A.; et al. Advancing Fishery-Independent Stock Assessments for the Norway Lobster (Nephrops norvegicus) with New Monitoring Technologies. Front. Mar. Sci. 2022, 9, 969071. [Google Scholar] [CrossRef]
- Martinelli, M.; Scarcella, G.; Sabatini, L.; Zacchetti, L.; Luzi, F.; Domenichetti, F. Convenzione tra ISPRA e CNR-IRBIM per la realizzazione di attività condivise, finalizzate a dare attuazione alle previsioni del d. lgs 13 ottobre 2010 n. 190, nell’ambito della Strategia Marina nel triennio 2021–2023. Modulo comunità epimegabentoniche–Mar Adriatico (GSA 17), Final report; IRBIM-CNR: Ancona, Italy, 2023. [Google Scholar]
- MIPAAF. Modalità Attuative per La Pesca Nella Fossa Di Pomo; Official Gazette of the Italian Republic No 2 of 03/01/2017; Ministero Delle Politiche Agricole Alimentari e Forestali: Rome, Italy, 2017. [Google Scholar]
- MIPAAF. Arresto Temporaneo Obbligatorio Delle Unità Autorizzate All’esercizio Della Pesca Con Il Sistema Strascico-Annualità 2015; Official Gazette of the Italian Republic No 162 of 15/07/2015; Ministero Delle Politiche Agricole Alimentari e Forestali: Rome, Italy, 2015. [Google Scholar]
- MIPAAF. Misure per La Pesca Nella Fossa Di Pomo; Ministero Delle Politiche Agricole Alimentari e Forestali: Rome, Italy, 2017; Available online: https://www.politicheagricole.it/flex/cm/pages/ServeAttachment.php/L/IT/D/9%252F0%252F4%252FD.583cb74efe7278af0dc3/P/BLOB%3AID%3D11345/E/pdf?mode=download (accessed on 12 May 2023).
- Wickham, H. ggplot2 Elegant Graphics for Data Analysis, 3rd. ed; Springer: New York, NY, USA, 2016; pp. 3–10. [Google Scholar]
- Wickham, H.; François, R.; Henry, L.; Müller, K. A Grammar of Data Manipulation [R Package Dplyr Version 1.0.0]. Media. 2020. Available online: https://cran.r-project.org/web/packages/dplyr/index.html (accessed on 10 April 2023).
- Fox, J.; Weisberg, S. An {R} Companion to Applied Regression, 3rd ed.; Sage: Thousand Oaks, CA, USA, 2019. [Google Scholar]
- Kassambara, A. Rstatix:Pipe-Friendly Framework for Basic Statistical Tests. R Package Version 0.7.1. 2022. Available online: https://CRAN.R-project.org/package=rstatix (accessed on 12 April 2023).
- Colloca, F.; Garofalo, G.; Bitetto, I.; Facchini, M.T.; Grati, F.; Martiradonna, A.; Mastrantonio, G.; Nikolioudakis, N.; Ordinas, F.; Scarcella, G.; et al. The Seascape of Demersal Fish Nursery Areas in the North Mediterranean Sea, a First Step towards the Implementation of Spatial Planning for Trawl Fisheries. PLoS ONE 2015, 10, e0119590. [Google Scholar] [CrossRef] [PubMed]
- Getis, A.; Ord, J.K. The Analysis of Spatial Association by Use of Distance Statistics. Geogr. Anal. 1992, 24, 189–206. [Google Scholar] [CrossRef]
- Mastrototaro, F.; Chimienti, G.; Acosta, J.; Blanco, J.; Garcia, S.; Rivera, J.; Aguilar, R. Isidella elongata (Cnidaria: Alcyonacea) Facies in the Western Mediterranean Sea: Visual Surveys and Descriptions of Its Ecological Role. Eur. Zool. J. 2017, 84, 209–225. [Google Scholar] [CrossRef]
- Lotze, H.K.; Coll, M.; Dunne, J.A. Historical Changes in Marine Resources, Food-Web Structure and Ecosystem Functioning in the Adriatic Sea, Mediterranean. Ecosystems 2011, 14, 198–222. [Google Scholar] [CrossRef]
- Eno, N.C.; MacDonald, D.S.; Kinnear, J.A.M.; Amos, S.C.; Chapman, C.J.; Clark, R.A.; Bunker, F.S.P.D.; Munro, C. Effects of Crustacean Traps on Benthic Fauna. ICES J. Mar. Sci. 2001, 58, 11–20. [Google Scholar] [CrossRef]
- Pierdomenico, M.; Russo, T.; Ambroso, S.; Gori, A.; Martorelli, E.; D’Andrea, L.; Gili, J.M.; Chiocci, F.L. Effects of Trawling Activity on the Bamboo-Coral Isidella elongata and the Sea Pen Funiculina quadrangularis along the Gioia Canyon (Western Mediterranean, Southern Tyrrhenian Sea). Prog. Oceanogr. 2018, 169, 214–226. [Google Scholar] [CrossRef]
- Bo, M.; Canese, S.; Spaggiari, C.; Pusceddu, A.; Bertolino, M.; Angiolillo, M.; Giusti, M.; Loreto, M.F.; Salvati, E.; Greco, S.; et al. Deep Coral Oases in the South Tyrrhenian Sea. PLoS ONE 2012, 7, e49870. [Google Scholar] [CrossRef] [PubMed]
- Russo, T.; D’Andrea, L.; Parisi, A.; Cataudella, S. VMSbase: An R-Package for VMS and Logbook Data Management and Analysis in Fisheries Ecology. PLoS ONE 2014, 9, e100195. [Google Scholar] [CrossRef]
- Russo, T.; D’Andrea, L.; Parisi, A.; Martinelli, M.; Belardinelli, A.; Boccoli, F.; Cignini, I.; Tordoni, M.; Cataudella, S. Assessing the Fishing Footprint Using Data Integrated from Different Tracking Devices: Issues and Opportunities. Ecol. Indic. 2016, 69, 818–827. [Google Scholar] [CrossRef]
- Ferrà Vega, C.; Tassetti, A.N.; Grati, F.; Scarcella, S.; Fabi, G. AIS as a Useful System to Support the Identification of Fisheries Restricted Areas. Licence: CC-NC-SA 3.0 IGO. FAO Fish Forum, B. Abstr.. 2018, p. 338. Available online: https://www.fao.org/3/cb7622en/cb7622en.pdf (accessed on 10 May 2023).
- Tassetti, A.N.; Ferrà, C.; Fabi, G. Rating the Effectiveness of Fishery-Regulated Areas with AIS Data. Ocean Coast. Manag. 2019, 175, 90–97. [Google Scholar] [CrossRef]
- De Clippele, L.H.; Buhl-Mortensen, P.; Buhl-Mortensen, L. Fauna Associated with Cold Water Gorgonians and Sea Pens. Cont. Shelf Res. 2015, 105, 67–78. [Google Scholar] [CrossRef]
- Grinyó, J.; Francescangeli, M.; Santín, A.; Ercilla, G.; Estrada, F.; Mecho, A.; Fanelli, E.; Costa, C.; Danovaro, R.; Company, J.B.; et al. Megafaunal assemblages in deep-sea ecosystems of the Gulf of Cadiz, northeast Atlantic ocean. Deep Sea Res. Part I Oceanogr. Res. Pap. 2022, 183, 103738. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martinelli, M.; Zacchetti, L.; Belardinelli, A.; Domenichetti, F.; Scarpini, P.; Penna, P.; Medvešek, D.; Isajlović, I.; Vrgoč, N. Changes in Abundance and Distribution of the Sea Pen, Funiculina quadrangularis, in the Central Adriatic Sea (Mediterranean Basin) in Response to Variations in Trawling Intensity. Fishes 2023, 8, 347. https://doi.org/10.3390/fishes8070347
Martinelli M, Zacchetti L, Belardinelli A, Domenichetti F, Scarpini P, Penna P, Medvešek D, Isajlović I, Vrgoč N. Changes in Abundance and Distribution of the Sea Pen, Funiculina quadrangularis, in the Central Adriatic Sea (Mediterranean Basin) in Response to Variations in Trawling Intensity. Fishes. 2023; 8(7):347. https://doi.org/10.3390/fishes8070347
Chicago/Turabian StyleMartinelli, Michela, Lorenzo Zacchetti, Andrea Belardinelli, Filippo Domenichetti, Paolo Scarpini, Pierluigi Penna, Damir Medvešek, Igor Isajlović, and Nedo Vrgoč. 2023. "Changes in Abundance and Distribution of the Sea Pen, Funiculina quadrangularis, in the Central Adriatic Sea (Mediterranean Basin) in Response to Variations in Trawling Intensity" Fishes 8, no. 7: 347. https://doi.org/10.3390/fishes8070347
APA StyleMartinelli, M., Zacchetti, L., Belardinelli, A., Domenichetti, F., Scarpini, P., Penna, P., Medvešek, D., Isajlović, I., & Vrgoč, N. (2023). Changes in Abundance and Distribution of the Sea Pen, Funiculina quadrangularis, in the Central Adriatic Sea (Mediterranean Basin) in Response to Variations in Trawling Intensity. Fishes, 8(7), 347. https://doi.org/10.3390/fishes8070347