
Nano-Curcumin/Chitosan Modulates Growth, Biochemical, Immune, and Antioxidative Profiles, and the Expression of Related Genes in Nile tilapia, Oreochromis niloticus

 , , , ,
, , , ,  ,
,  ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation of Curcumin Nanoparticles
2.1.1. Synthesis of Curcumin Nanoparticles
2.1.2. Synthesis of Nano-curcumin and Chitosan Capsule (NCur/Ch)
2.1.3. Characterization of Curcumin Nanoparticles
2.2. Diets and Experimental Conditions
2.3. Growth Performance
2.4. Digestive Enzymes
2.5. Immune Parameters
2.6. Biochemical Assays
2.7. Antioxidants
2.8. Intestinal Morphometric Analysis
2.9. Real-Time PCR Analysis of igf-1 and cc5
2.10. Statistical Analysis
3. Results
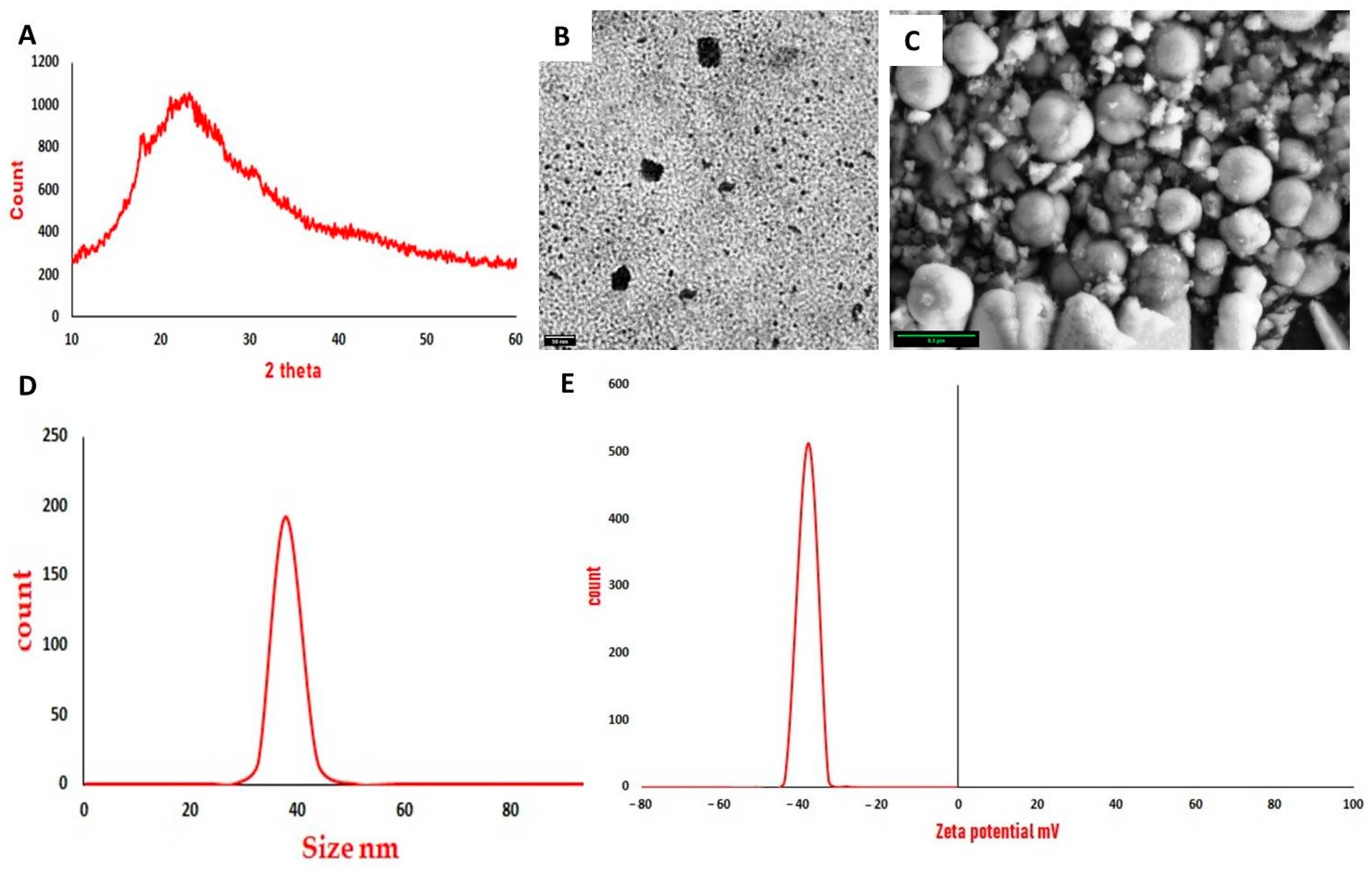
3.1. Characterization of Curcumin Nanoparticles
3.2. Growth Performance
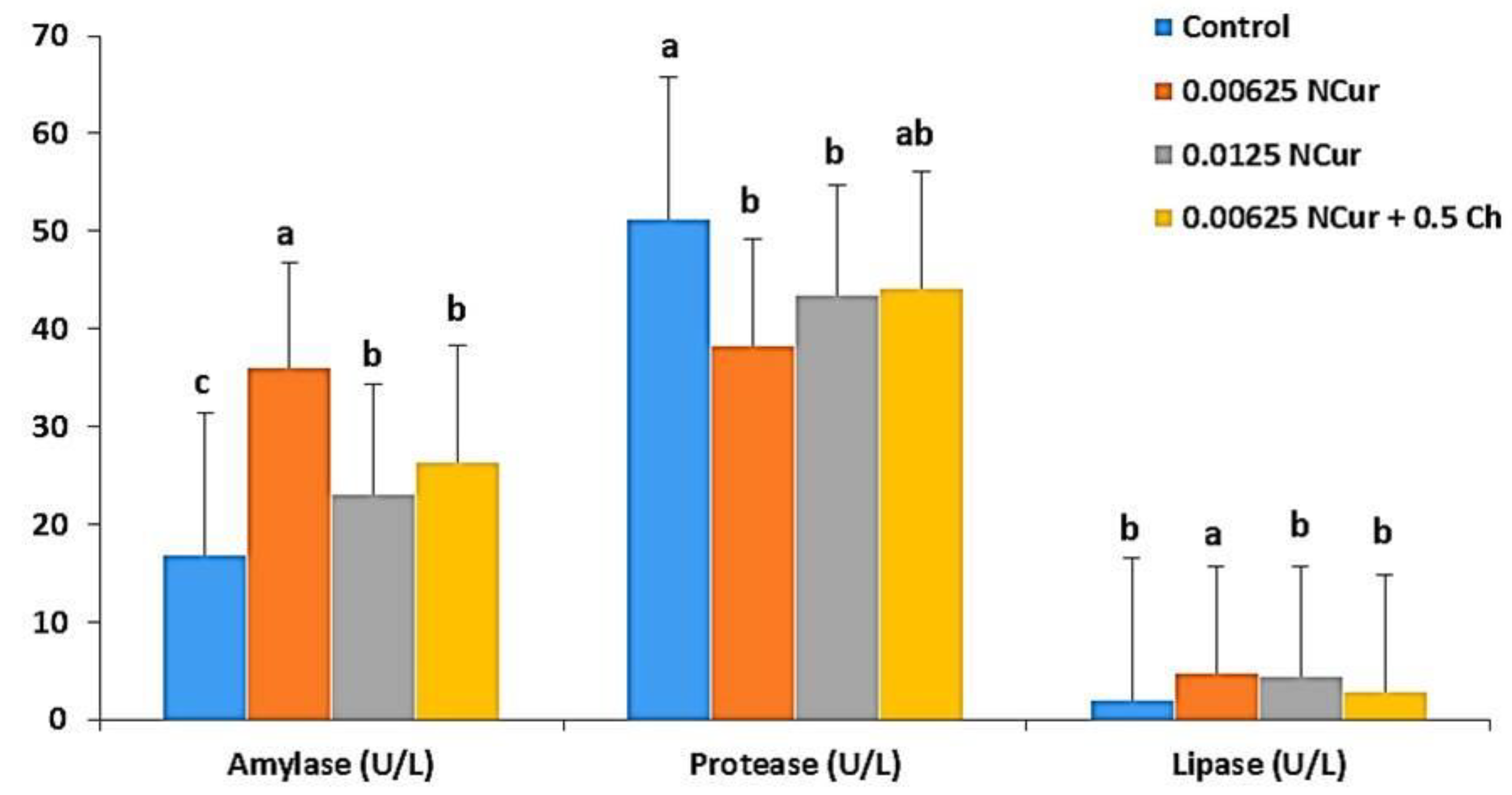
3.3. Digestive Enzymes
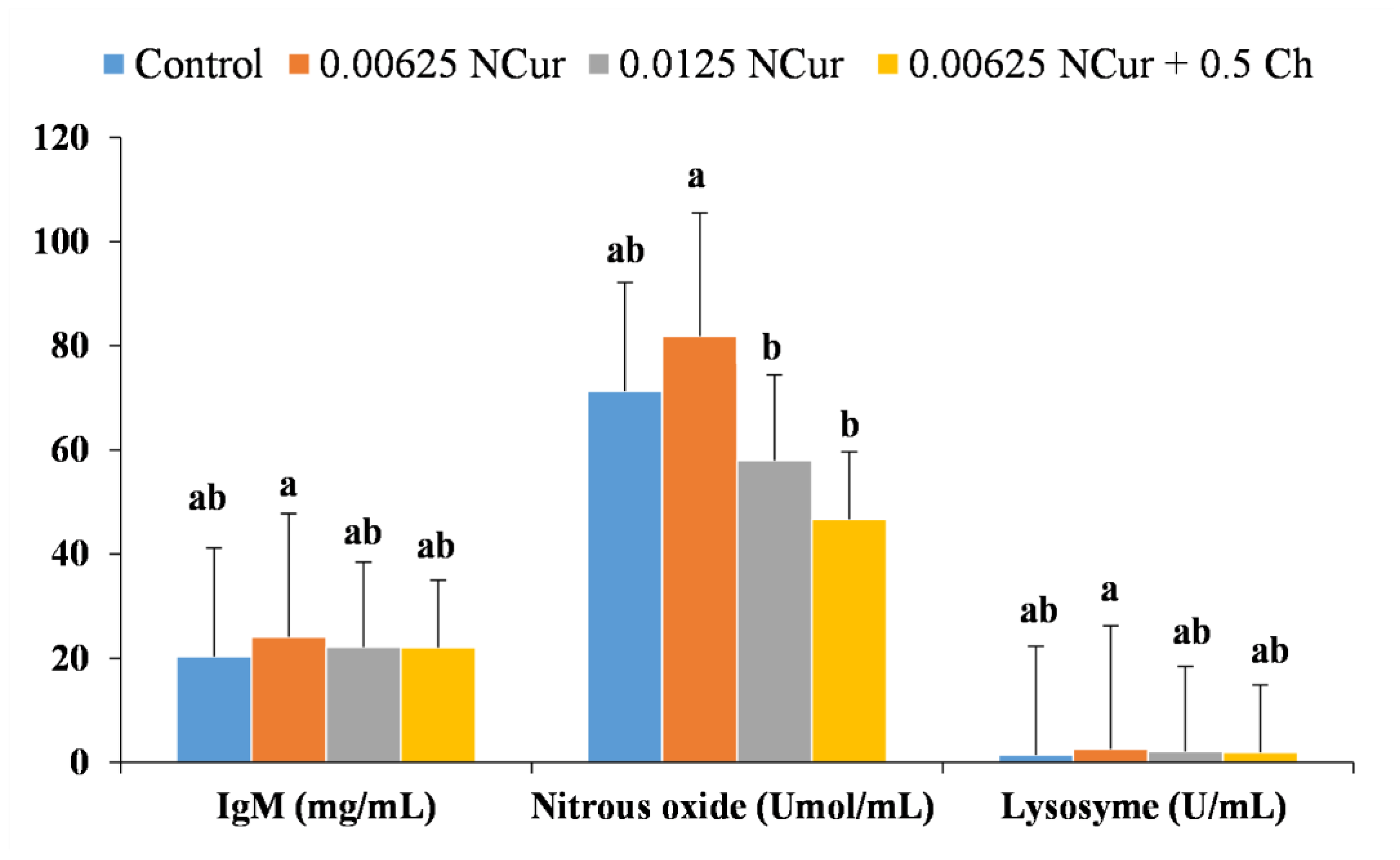
3.4. Immune Response
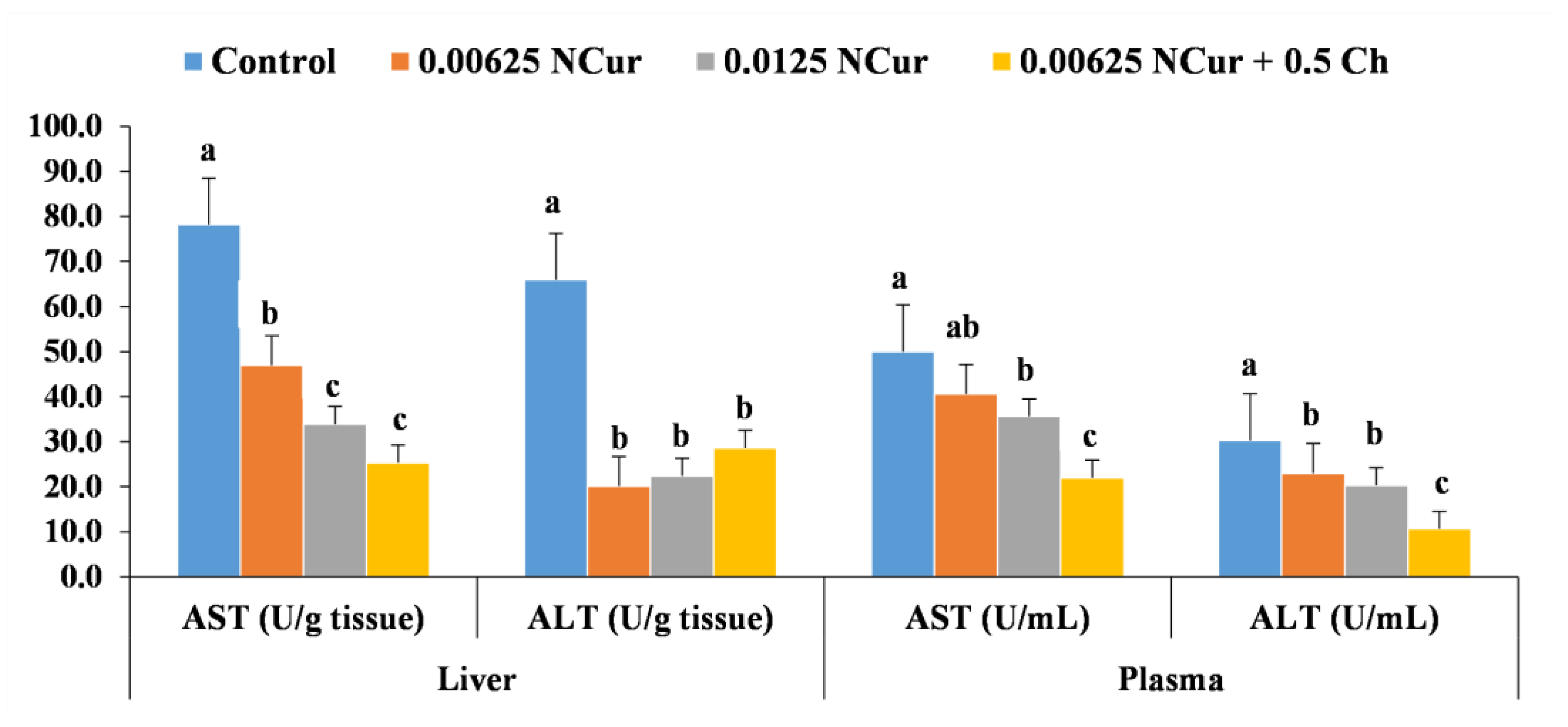
3.5. Biochemical Profile and Plasma Proteins
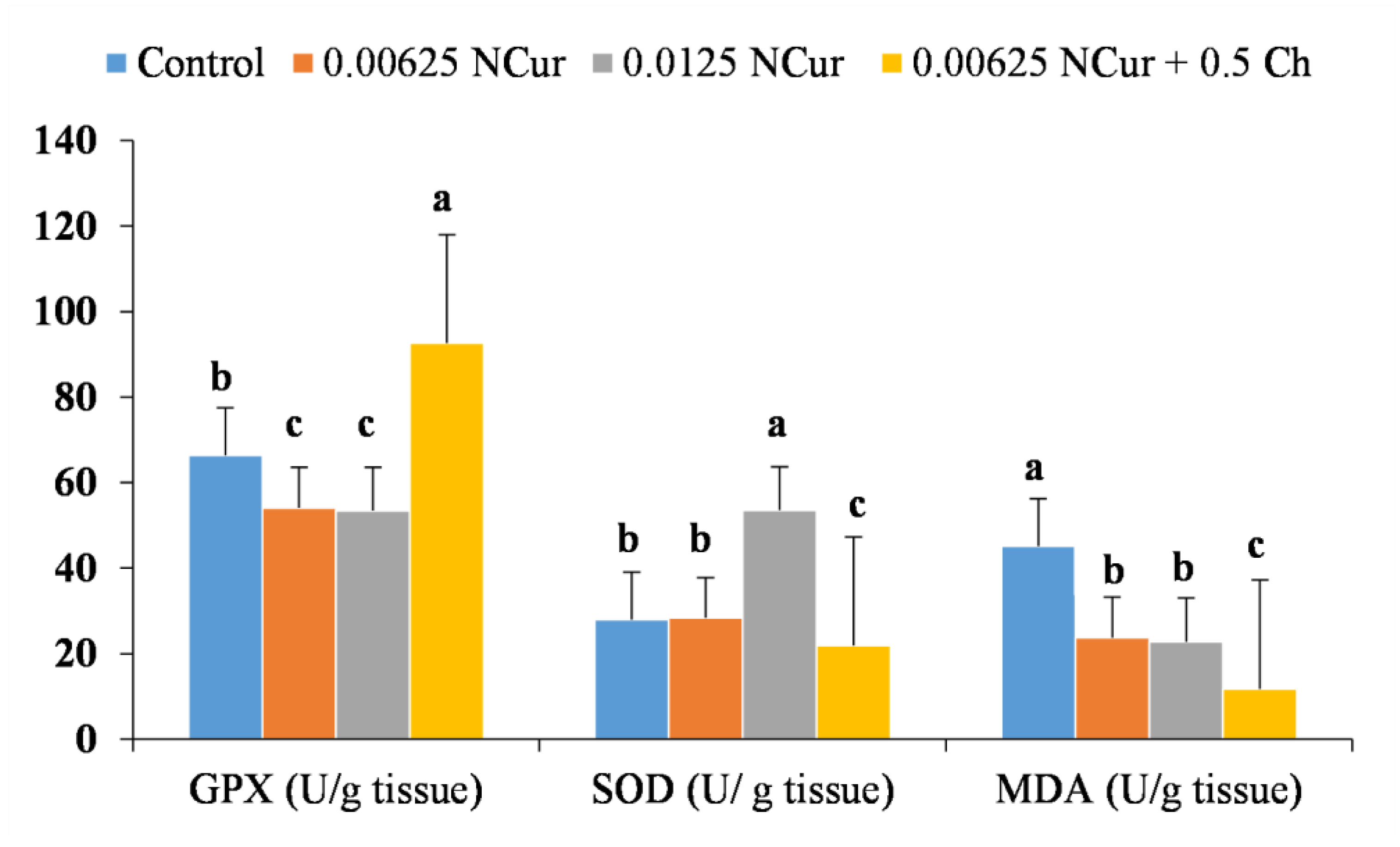
3.6. Antioxidants
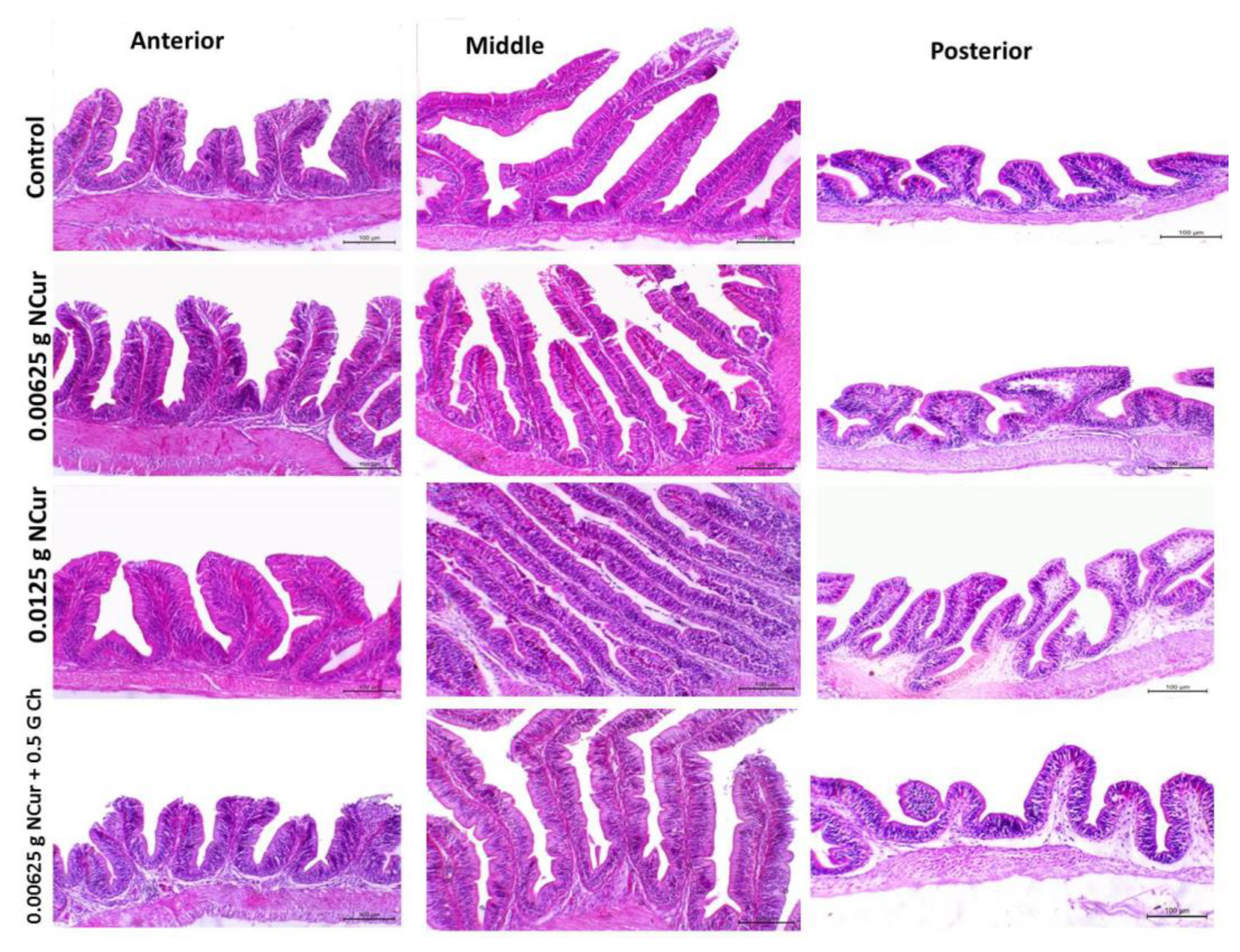
3.7. Intestinal Morphometric Analysis
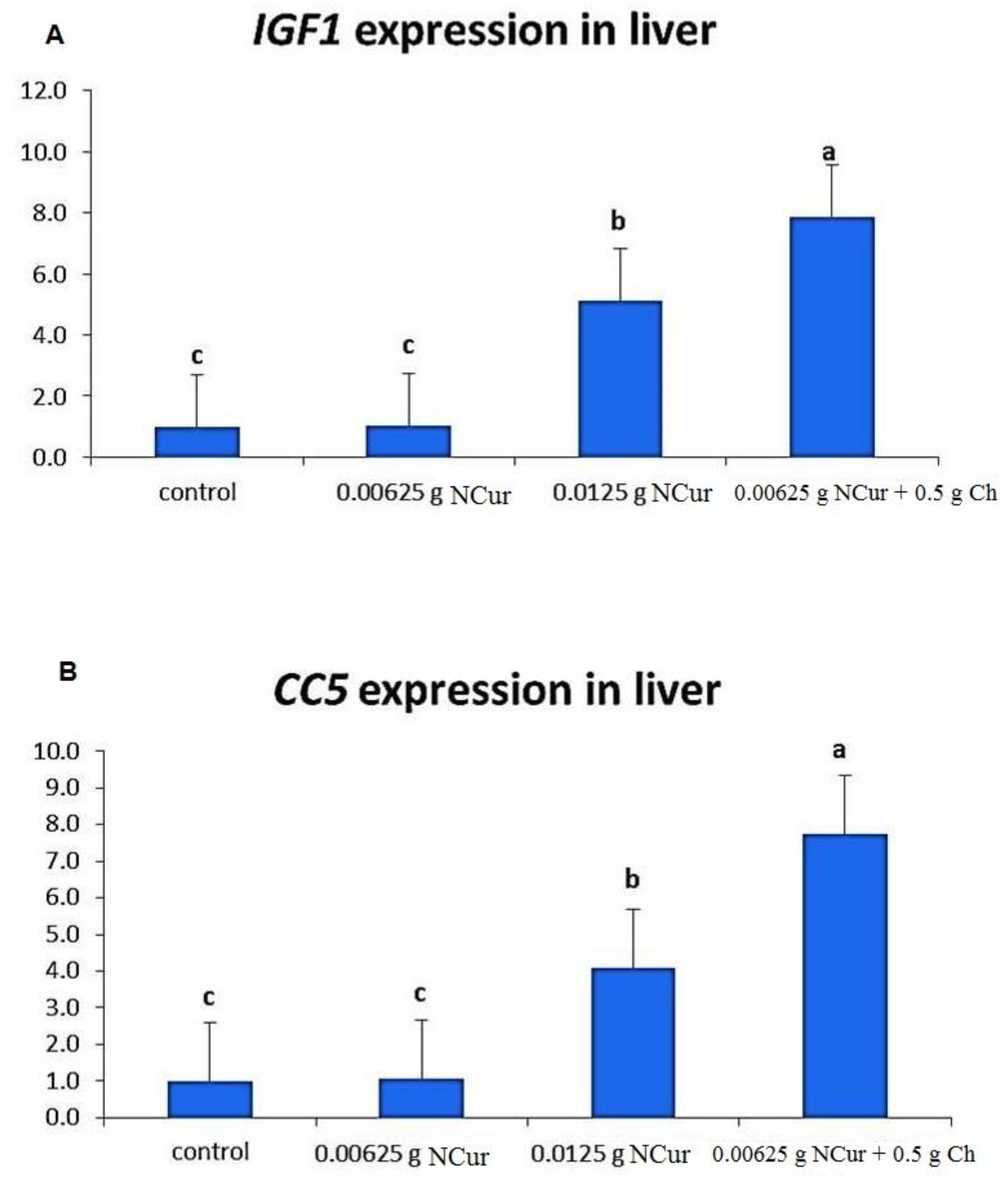
3.8. Gene Expression of igf-1 and cc5
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sallam, A.E.; Mansour, A.T.; Alsaqufi, A.S.; Salem, M.E.-S.; El-Feky, M.M. Growth performance, antioxidative status, innate immunity, and ammonia stress resistance of Siganus rivulatus fed diet supplemented with zinc and zinc nanoparticles. Aquac. Rep. 2020, 18, 100410. [Google Scholar] [CrossRef]
- Sharawy, Z.Z.; Ashour, M.; Labena, A.; Alsaqufi, A.S.; Mansour, A.T.; Abbas, E.M. Effects of dietary Arthrospira platensis nanoparticles on growth performance, feed utilization, and growth-related gene expression of Pacific white shrimp, Litopenaeus vannamei. Aquaculture 2022, 551, 737905. [Google Scholar] [CrossRef]
- Elabd, H.; Youssuf, H.; Mahboub, H.H.; Salem, S.M.; Husseiny, W.A.; Khalid, A.; El-Desouky, H.S.; Faggio, C. Growth, hemato-biochemical, immune-antioxidant response, and gene expression in Nile tilapia (Oreochromis niloticus) received nano iron oxide-incorporated diets. Fish Shellfish. Immunol. 2022, 128, 574–581. [Google Scholar] [CrossRef] [PubMed]
- Moghadam, H.; Sourinejad, I.; Johari, S.A. Growth performance, haemato-immunological responses and antioxidant status of Pacific white shrimp Penaeus vannamei fed with turmeric powder, curcumin and curcumin nanomicelles. Aquac. Nutr. 2021, 27, 2294–2306. [Google Scholar] [CrossRef]
- Rather, M.; Sharma, R.; Aklakur, M.; Ahmad, S.; Kumar, N.; Khan, M.; Ramya, V. Nanotechnology: A novel tool for aquaculture and fisheries development. A prospective mini-review. Fish. Aquac. J. 2011, 16, 3. [Google Scholar]
- Rashidi, L. Different nano-delivery systems for delivery of nutraceuticals. Food Biosci. 2021, 43, 101258. [Google Scholar] [CrossRef]
- Pirani, F.; Moradi, S.; Ashouri, S.; Johari, S.A.; Ghaderi, E.; Kim, H.P.; Yu, I.J. Dietary supplementation with curcumin nanomicelles, curcumin, and turmeric affects growth performance and silver nanoparticle toxicity in Cyprinus carpio. Environ. Sci. Pollut. Res. 2021, 28, 64706–64718. [Google Scholar] [CrossRef]
- El-Houseiny, W.; Anter, R.G.; Arisha, A.H.; Mansour, A.T.; Safhi, F.A.; Alwutayd, K.M.; Elshopakey, G.E.; Abd El-Hakim, Y.M.; Mohamed, E.M. Growth Retardation, Oxidative Stress, Immunosuppression, and Inflammatory Disturbances Induced by Herbicide Exposure of Catfish, Clarias gariepinus, and the Alleviation Effect of Dietary Wormwood, Artemisia cina. Fishes 2023, 8, 297. [Google Scholar] [CrossRef]
- Shahin, S.A.; Mansour, A.T.; Abdel-Rahim, M.M.; El-Dahhar, A.A.; El Basuini, M.F.; Elhetawy, A.I. Improving survival, growth, feed utilization, antioxidant status, and fatty acids profile of European seabass, larvae fed silymarin, supplemented weaning diet. Ann. Anim. Sci. 2022, 23, 253–264. [Google Scholar] [CrossRef]
- Elumalai, P.; Kurian, A.; Lakshmi, S.; Faggio, C.; Esteban, M.A.; Ringø, E. Herbal immunomodulators in aquaculture. Rev. Fish. Sci. Aquac. 2020, 29, 33–57. [Google Scholar] [CrossRef]
- Mansour, A.T.; Hamed, H.S.; El-Beltagi, H.S.; Mohamed, W.F. Modulatory Effect of Papaya Extract against Chlorpyrifos-Induced Oxidative Stress, Immune Suppression, Endocrine Disruption, and DNA Damage in Female Clarias gariepinus. Int. J. Environ. Res. Public Health 2022, 19, 4640. [Google Scholar] [CrossRef]
- Mansour, A.T.; Mahboub, H.H.; Amen, R.M.; El-Beltagy, M.A.; Ramah, A.; Abdelfattah, A.M.; El-Beltagi, H.S.; Shalaby, T.A.; Ghazzawy, H.S.; Ramadan, K.M.; et al. Ameliorative Effect of Quercetin against Abamectin-Induced Hemato-Biochemical Alterations and Hepatorenal Oxidative Damage in Nile Tilapia, Oreochromis niloticus. Animals 2022, 12, 3429. [Google Scholar] [CrossRef] [PubMed]
- Almarri, S.H.; Khalil, A.A.; Mansour, A.T.; El-Houseiny, W. Antioxidant, Immunostimulant, and Growth-Promoting Effects of Dietary Annona squamosa Leaf Extract on Nile Tilapia, Oreochromis niloticus, and Its Tolerance to Thermal Stress and Aeromonas sobria Infection. Animals 2023, 13, 746. [Google Scholar] [CrossRef] [PubMed]
- Tabanelli, R.; Brogi, S.; Calderone, V. Improving curcumin bioavailability: Current strategies and future perspectives. Pharmaceutics 2021, 13, 1715. [Google Scholar] [CrossRef]
- Alagawany, M.; Farag, M.R.; Abdelnour, S.A.; Dawood, M.A.; Elnesr, S.S.; Dhama, K. Curcumin and its different forms: A review on fish nutrition. Aquaculture 2021, 532, 736030. [Google Scholar] [CrossRef]
- El-abd, H.; El-latif, A.; Shaheen, A. Effect of curcumin on growth performance and antioxidant stress status of Nile tilapia (Oreochromis niloticus). Iran. J. Fish. Sci. 2021, 20, 1234–1246. [Google Scholar]
- Abdel-Tawwab, M.; Eissa, E.-S.H.; Tawfik, W.A.; Abd Elnabi, H.E.; Saadony, S.; Bazina, W.K.; Ahmed, R.A. Dietary curcumin nanoparticles promoted the performance, antioxidant activity, and humoral immunity, and modulated the hepatic and intestinal histology of Nile tilapia fingerlings. Fish Physiol. Biochem. 2022, 48, 585–601. [Google Scholar] [CrossRef]
- Kotha, R.R.; Luthria, D.L. Curcumin: Biological, pharmaceutical, nutraceutical, and analytical aspects. Molecules 2019, 24, 2930. [Google Scholar] [CrossRef]
- Karthikeyan, A.; Senthil, N.; Min, T. Nanocurcumin: A promising candidate for therapeutic applications. Front. Pharmacol. 2020, 11, 487. [Google Scholar] [CrossRef]
- Tawfik, S.M.; Azizov, S.; Elmasry, M.R.; Sharipov, M.; Lee, Y.-I. Recent advances in nanomicelles delivery systems. Nanomaterials 2020, 11, 70. [Google Scholar] [CrossRef]
- Abdel-Ghany, H.M.; Salem, M.E.S. Effects of dietary chitosan supplementation on farmed fish: A review. Rev. Aquac. 2020, 12, 438–452. [Google Scholar] [CrossRef]
- El-Naggar, M.; Medhat, F.; Taha, A. Applications of chitosan and chitosan nanoparticles in fish aquaculture. Egypt. J. Aquat. Biol. Fish. 2022, 26, 23–43. [Google Scholar] [CrossRef]
- Yan, J.; Guo, C.; Dawood, M.; Gao, J. Effects of dietary chitosan on growth, lipid metabolism, immune response and antioxidant-related gene expression in Misgurnus anguillicaudatus. Benef. Microbes 2017, 8, 439–449. [Google Scholar] [CrossRef] [PubMed]
- Abu-Elala, N.M.; Mohamed, S.H.; Zaki, M.M.; Eissa, A.E. Assessment of the immune-modulatory and antimicrobial effects of dietary chitosan on Nile tilapia (Oreochrmis niloticus) with special emphasis to its bio-remediating impacts. Fish Shellfish. Immunol. 2015, 46, 678–685. [Google Scholar] [CrossRef] [PubMed]
- Maqsood, S.; Singh, P.; Samoon, M.H.; Balange, A.K. Effect of dietary chitosan on non-specific immune response and growth of Cyprinus carpio challenged with Aeromonas hydrophila. Int. Aquat. Res. 2010, 2, 77. [Google Scholar]
- Abdel-Tawwab, M.; Razek, N.A.; Abdel-Rahman, A.M. Immunostimulatory effect of dietary chitosan nanoparticles on the performance of Nile tilapia, Oreochromis niloticus (L.). Fish Shellfish. Immunol. 2019, 88, 254–258. [Google Scholar] [CrossRef]
- Saleh, M.; Essawy, E.; Shaalan, M.; Osman, S.; Ahmed, F.; El-Matbouli, M. Therapeutic Intervention with Dietary Chitosan Nanoparticles Alleviates Fish Pathological and Molecular Systemic Inflammatory Responses against Infections. Mar. Drugs 2022, 20, 425. [Google Scholar] [CrossRef]
- Wu, Y.; Rashidpour, A.; Almajano, M.P.; Metón, I. Chitosan-based drug delivery system: Applications in fish biotechnology. Polymers 2020, 12, 1177. [Google Scholar] [CrossRef]
- Elabd, H.; Wang, H.-P.; Shaheen, A.; Yao, H.; Abbass, A. Anti-oxidative effects of some dietary supplements on Yellow perch (Perca flavescens) exposed to different physical stressors. Aquac. Rep. 2017, 8, 21–30. [Google Scholar] [CrossRef]
- Liu, Z.; Que, S.; Xu, J.; Peng, T. Alanine aminotransferase-old biomarker and new concept: A review. Int. J. Med. Sci. 2014, 11, 925. [Google Scholar] [CrossRef]
- Huang, X.-J.; Choi, Y.-K.; Im, H.-S.; Yarimaga, O.; Yoon, E.; Kim, H.-S. Aspartate aminotransferase (AST/GOT) and alanine aminotransferase (ALT/GPT) detection techniques. Sensors 2006, 6, 756–782. [Google Scholar] [CrossRef]
- Aebi, H. Methods in Enzymology Oxygen Radical in Biological System; Academic Press, Inc.: Cambridge, MA, USA, 1984; Volume 105, pp. 121–126. [Google Scholar]
- Suvarna, S.K.; Layton, C.; Bancroft, J.D. Theory and Practice of Histological Techniqueseighth; Elsevier: Amsterdam, The Netherlands, 2019. [Google Scholar]
- Elabd, H.; Wang, H.-P.; Shaheen, A.; Yao, H.; Abbass, A. Feeding Glycyrrhiza glabra (liquorice) and Astragalus membranaceus (AM) alters innate immune and physiological responses in yellow perch (Perca flavescens). Fish Shellfish. Immunol. 2016, 54, 374–384. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.S.; Reed, A.; Chen, F.; Stewart, C.N. Statistical analysis of real-time PCR data. BMC Bioinform. 2006, 7, 85. [Google Scholar] [CrossRef]
- Gröner, F.; Ziková, A.; Kloas, W. Effects of the pharmaceuticals diclofenac and metoprolol on gene expression levels of enzymes of biotransformation, excretion pathways and estrogenicity in primary hepatocytes of Nile tilapia (Oreochromis niloticus). Comp. Biochem. Physiol. C Toxicol. 2015, 167, 51–57. [Google Scholar] [CrossRef]
- Cruz, E.V.; Brown, C. Influence of the photoperiod on growth rate and insulin-like growth factor-I gene expression in Nile tilapia Oreochromis niloticus. J. Fish Biol. 2009, 75, 130–141. [Google Scholar] [CrossRef]
- Abdel-Tawwab, M.; Abbass, F.E. Turmeric powder, Curcuma longa L., in common carp, Cyprinus carpio L., diets: Growth performance, innate immunity, and challenge against pathogenic Aeromonas hydrophila infection. J. World Aquac. Soc. 2017, 48, 303–312. [Google Scholar] [CrossRef]
- Mahmoud, H.K.; Al-Sagheer, A.A.; Reda, F.M.; Mahgoub, S.A.; Ayyat, M.S. Dietary curcumin supplement influence on growth, immunity, antioxidant status, and resistance to Aeromonas hydrophila in Oreochromis niloticus. Aquaculture 2017, 475, 16–23. [Google Scholar] [CrossRef]
- Sruthi, M.; Nair, A.B.; Arun, D.; Thushara, V.; Sheeja, C.; Vijayasree, A.S.; Oommen, O.V.; Divya, L. Dietary curcumin influences leptin, growth hormone and hepatic growth factors in Tilapia (Oreochromis mossambicus). Aquaculture 2018, 496, 105–111. [Google Scholar] [CrossRef]
- Ashry, A.M.; Hassan, A.M.; Habiba, M.M.; El-Zayat, A.; El-Sharnouby, M.E.; Sewilam, H.; Dawood, M.A. The impact of dietary curcumin on the growth performance, intestinal antibacterial capacity, and haemato-biochemical parameters of gilthead seabream (Sparus aurata). Animals 2021, 11, 1779. [Google Scholar] [CrossRef]
- Jiang, J.; Wu, X.-Y.; Zhou, X.-Q.; Feng, L.; Liu, Y.; Jiang, W.-D.; Wu, P.; Zhao, Y. Effects of dietary curcumin supplementation on growth performance, intestinal digestive enzyme activities and antioxidant capacity of crucian carp Carassius auratus. Aquaculture 2016, 463, 174–180. [Google Scholar] [CrossRef]
- Fadl, S.E.; El-Gammal, G.A.; Abdo, W.S.; Barakat, M.; Sakr, O.A.; Nassef, E.; Gad, D.M.; El-Sheshtawy, H.S. Evaluation of dietary chitosan effects on growth performance, immunity, body composition and histopathology of Nile tilapia (Oreochromis niloticus) as well as the resistance to Streptococcus agalactiae infection. Aquac. Res. 2020, 51, 1120–1132. [Google Scholar] [CrossRef]
- Chen, Y.; Zhu, X.; Yang, Y.; Han, D.; Jin, J.; Xie, S. Effect of dietary chitosan on growth performance, haematology, immune response, intestine morphology, intestine microbiota and disease resistance in gibel carp (Carassius auratus gibelio). Aquac. Nutr. 2014, 20, 532–546. [Google Scholar] [CrossRef]
- Geng, X.; Dong, X.-H.; Tan, B.-P.; Yang, Q.-H.; Chi, S.-Y.; Liu, H.-Y.; Liu, X.-Q. Effects of dietary chitosan and Bacillus subtilis on the growth performance, non-specific immunity and disease resistance of cobia, Rachycentron canadum. Fish Shellfish. Immunol. 2011, 31, 400–406. [Google Scholar] [CrossRef]
- Ming, J.; Ye, J.; Zhang, Y.; Xu, Q.; Yang, X.; Shao, X.; Qiang, J.; Xu, P. Optimal dietary curcumin improved growth performance, and modulated innate immunity, antioxidant capacity and related genes expression of NF-κB and Nrf2 signaling pathways in grass carp (Ctenopharyngodon idella) after infection with Aeromonas hydrophila. Fish Shellfish. Immunol. 2020, 97, 540–553. [Google Scholar] [CrossRef]
- Yonar, M.E.; Yonar, S.M.; İspir, Ü.; Ural, M.Ş. Effects of curcumin on haematological values, immunity, antioxidant status and resistance of rainbow trout (Oncorhynchus mykiss) against Aeromonas salmonicida subsp. achromogenes. Fish Shellfish. Immunol. 2019, 89, 83–90. [Google Scholar] [CrossRef]
- Abdelkhalek, N.; El-Adl, M.; El-Ashram, A.; Othman, M.; Gadallah, H.; El-Diasty, M.; Dawood, M.A.; Almeer, R.; Abdel Daim, M. Immunological and antioxidant role of curcumin in ameliorating fipronil toxicity in Nile tilapia (Oreochromis niloticus). Aquac. Res. 2021, 52, 2791–2801. [Google Scholar] [CrossRef]
- Kohli, K.; Ali, J.; Ansari, M.; Raheman, Z. Curcumin: A natural antiinflammatory agent. Indian J. Pharmacol. 2005, 37, 141. [Google Scholar] [CrossRef]
- Gheytasi, A.; Shekarabi, S.P.H.; Islami, H.R.; Mehrgan, M.S. Feeding rainbow trout, Oncorhynchus mykiss, with lemon essential oil loaded in chitosan nanoparticles: Effect on growth performance, serum hemato-immunological parameters, and body composition. Aquac. Int. 2021, 29, 2207–2221. [Google Scholar] [CrossRef]
- Yousef, M.I.; El-Demerdash, F.M.; Radwan, F.M. Sodium arsenite induced biochemical perturbations in rats: Ameliorating effect of curcumin. Food Chem. Toxicol. 2008, 46, 3506–3511. [Google Scholar] [CrossRef] [PubMed]
- Mansour, W.A.; Abdelsalam, N.R.; Tanekhy, M.; Khaled, A.A.; Mansour, A.T. Toxicity, inflammatory and antioxidant genes expression, and physiological changes of green synthesis silver nanoparticles on Nile tilapia (Oreochromis niloticus) fingerlings. Comp. Biochem. Physiol. C Toxicol. 2021, 247, 109068. [Google Scholar] [CrossRef]
- Faheem, M.; Khaliq, S.; Abbas, R.Z.; Mansour, A.T. Moringa oleifera alleviated oxidative stress, physiological and molecular disruption induced by acute thermal stress in grass carp, Ctenopharyngodon idella. Fish Physiol. Biochem. 2022, 48, 1463–1473. [Google Scholar] [CrossRef]
- Mehta, K.; Balaraman, R.; Amin, A.; Bafna, P.; Gulati, O. Effect of fruits of Moringa oleifera on the lipid profile of normal and hypercholesterolaemic rabbits. J. Ethnopharmacol. 2003, 86, 191–195. [Google Scholar] [CrossRef]
- Ibrahim, D.; Neamat-Allah, A.N.; Ibrahim, S.M.; Eissa, H.M.; Fawzey, M.; Mostafa, D.I.; Abd El-Kader, S.A.; Khater, S.; Khater, S.I. Dual effect of Selenium loaded Chitosan Nanoparticles on growth, antioxidant, immune related genes expression, transcriptomics modulation of caspase 1, cytochrome P450 and heat shock protein and Aeromonas hydrophila resistance of Nile Tilapia (Oreochromis niloticus). Fish Shellfish Immunol. 2021, 110, 91–99. [Google Scholar]
- Ji, R.; Xiang, X.; Li, X.; Mai, K.; Ai, Q. Effects of dietary curcumin on growth, antioxidant capacity, fatty acid composition and expression of lipid metabolism-related genes of large yellow croaker fed a high-fat diet. Br. J. Nutr. 2021, 126, 345–354. [Google Scholar] [CrossRef] [PubMed]
- Elsabagh, M.; Mohamed, R.; Moustafa, E.M.; Hamza, A.; Farrag, F.; Decamp, O.; Dawood, M.A.; Eltholth, M. Assessing the impact of Bacillus strains mixture probiotic on water quality, growth performance, blood profile and intestinal morphology of Nile tilapia, Oreochromis niloticus. Aquac. Nutr. 2018, 24, 1613–1622. [Google Scholar] [CrossRef]
- Giannenas, I.; Pappas, I.; Mavridis, S.; Kontopidis, G.; Skoufos, J.; Kyriazakis, I. Performance and antioxidant status of broiler chickens supplemented with dried mushrooms (Agaricus bisporus) in their diet. Poult. Sci. 2010, 89, 303–311. [Google Scholar] [CrossRef] [PubMed]
- Peterson, B.C.; Waldbieser, G.C.; Bilodeau, L. IGF-I and IGF-II mRNA expression in slow and fast growing families of USDA103 channel catfish (Ictalurus punctatus). Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2004, 139, 317–323. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | g/kg Diet |
|---|---|
| Fish meal * | 295 |
| Yellow corn | 185 |
| Soybean | 365 |
| Wheat bran | 45 |
| Vegetable oil | 35 |
| Gluten | 50 |
| Bicalcium phosphate | 15 |
| NaCl | 3 |
| Vitamin and mineral premixes ** | 7 |
| Ingredient | Approximate analysis (%) |
| Crude protein | 38.28 |
| Ether extract | 6.33 |
| Crude fiber | 8.35 |
| Ash | 9.04 |
| Digestible energy (Kcal/kg) | 3312 |
| Gene of Interest | Primer Sequences | Reference |
|---|---|---|
| EF-1α | CCTTCAACGCTCAGGTCATC | [36] |
| TGTGGGCAGTGTGGCAATC | ||
| igf-1 | TTCTCCAAAAACGAGCCTGCG | [37] |
| TCTGCTACTAACCTTGGGTGC | ||
| cc5 | GGACCCGGACCATACAACAG | [16] |
| GGGGTTTTGCAGAGATGGGA |
| Treatments | Initial Weight (g) | Final Weight (g) | Initial Length (cm) | Final Length (cm) | BMG (%) | SGR (%) | FCR | LGR (%) |
|---|---|---|---|---|---|---|---|---|
| Control | 12.5 ± 0.5 c | 26.6 ± 0.2 c | 9.2 ± 0.4 | 10.7 ± 0.2 c | 112.4 ± 0.05 c | 4.7 ± 0.01 c | 1.1 ± 0.03 c | 14.6 ± 0.03 c |
| 0.00625 NCur | 12.5 ± 0.5 b | 35.4 ± 0.5 b | 8 ± 0.5 | 11.8 ± 0.1 b | 182.6 ± 0.05 b | 5.1 ± 0.00 b | 0.8 ± 0.00 b | 47.9 ± 0.01 b |
| 0.0125 NCur | 12.5 ± 0.5 a | 46.8 ± 0.2 a | 8 ± 0.3 | 13.8 ± 0.2 a | 274.6 ± 0.04 a | 5.5 ± 0.00 a | 0.5 ± 0.00 a | 72.6 ± 0.01 a |
| 0.00625 NCur + 0.5 Ch | 12.5 ± 0.5 a | 44.8 ± 0.2 a | 8 ± 0.5 | 14.3 ± 0.2 a | 258.2 ± 0.05 a | 5.5 ± 0.01 a | 0.5 ± 0.01 a | 79.2 ± 0.03 a |
| Treatments | Cortisol (ng/mL) | Glucose (mg/dL) | Albumin (g/dL) | Globulin (g/dL) | Total Protein (g/dL) |
|---|---|---|---|---|---|
| Control | 38.0 ± 0.03 a | 60.0 ± 0.05 a | 0.7 ± 0.04 b | 1.3 ± 0.02 | 2.0 ± 0.00 c |
| 0.00625 NCur | 21.6 ± 0.01 a | 24.3 ± 0.02 a b | 0.9 ± 0.00 a | 1.6 ± 0.01 | 2.5 ± 0.00 a |
| 0.0125 NCur | 16.3 ± 0.05 b | 28.5 ± 0.00 b | 0.8 ± 0.04 a b | 1.4 ± 0.04 | 2.2 ± 0.00 b |
| 0.00625 NCur + 0.5 Ch | 12.7 ± 0.01 b | 22.8 ± 0.01 c | 0.7 ± 0.00 b | 1.3 ± 0.00 | 2.0 ± 0.00 c |
| Treatments | Anterior Intestine | |||
|---|---|---|---|---|
| Villus Length | Villus Width | Inter-Villi Space | Goblet Cells/mm2 | |
| Control | 167.0 ± 1.4 c | 104.45 ± 3.5 b | 102.7 ± 3.5 a | 19.3 ± 0.8 c |
| 0.00625 NCur | 229.67 ± 3.7 b | 100.07 ± 0.8 b | 77.3 ± 1.5 b | 27.6 ± 2.4 a b |
| 0.0125 NCur | 318.8 ± 2.5 a | 120.68 ± 2.3 a | 51.5 ± 1.5 c | 30.0 ± 2.4 a |
| 0.00625 NCur + 0.5 Ch | 275.0 ± 1.3 a | 94.1570 ± 0.9 b | 75.7 ± 1.3 b | 22.0 ± 1.5 b c |
| Middle Intestine | ||||
| Control | 312.3 ± 3.5 c | 74.2 ± 3.5 | 74.8 ± 3.5 a | 30.6 ± 1.2 b |
| 0.00625 NCur | 416.6 ± 2.5 b | 76.6 ± 2.5 | 55.2 ± 2.4 a b | 36.0 ± 1.3 b |
| 0.0125 NCur | 618.1 ± 3.5 a | 60.4 ± 3.5 | 18.5 ± 1.3 c | 57.0 ± 2.5 a |
| 0.00625 NCur + 0.5 Ch | 433.2 ± 2.7 b | 76.6 ± 2.5 | 44.5 ± 2.5 b c | 38.3 ± 1.2 b |
| Posterior Intestine | ||||
| Control | 103.7 ± 3.5 b | 87.2 ± 3.5 b | 122.9 ± 4.1 a | 7.6 ± 1.2 b |
| 0.00625 NCur | 114.1 ± 3.2 b | 130.0 ± 1.5 a | 116.4 ± 3.5 a b | 12.3 ± 1.8 b |
| 0.0125 NCur | 204.6 ± 1.5 a | 92.9 ± 2.0 b | 84.5 ± 2.4 c | 23.0 ± 2.5 a |
| 0.00625 NCur + 0.5 Ch | 165.4 ± 2.5 a | 91.2 ± 3.5 b | 95.4 ± 1.5 b c | 12.6 ± 1.2 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elabd, H.; Mahboub, H.H.; Salem, S.M.R.; Abdelwahab, A.M.; Alwutayd, K.M.; Shaalan, M.; Ismail, S.H.; Abdelfattah, A.M.; Khalid, A.; Mansour, A.T.; et al. Nano-Curcumin/Chitosan Modulates Growth, Biochemical, Immune, and Antioxidative Profiles, and the Expression of Related Genes in Nile tilapia, Oreochromis niloticus. Fishes 2023, 8, 333. https://doi.org/10.3390/fishes8070333
Elabd H, Mahboub HH, Salem SMR, Abdelwahab AM, Alwutayd KM, Shaalan M, Ismail SH, Abdelfattah AM, Khalid A, Mansour AT, et al. Nano-Curcumin/Chitosan Modulates Growth, Biochemical, Immune, and Antioxidative Profiles, and the Expression of Related Genes in Nile tilapia, Oreochromis niloticus. Fishes. 2023; 8(7):333. https://doi.org/10.3390/fishes8070333
Chicago/Turabian StyleElabd, Hiam, Heba H. Mahboub, Shimaa M. R. Salem, Abdelwahab M. Abdelwahab, Khairiah Mubarak Alwutayd, Mohamed Shaalan, Sameh H. Ismail, Abdelfattah M. Abdelfattah, A. Khalid, Abdallah Tageldein Mansour, and et al. 2023. "Nano-Curcumin/Chitosan Modulates Growth, Biochemical, Immune, and Antioxidative Profiles, and the Expression of Related Genes in Nile tilapia, Oreochromis niloticus" Fishes 8, no. 7: 333. https://doi.org/10.3390/fishes8070333
APA StyleElabd, H., Mahboub, H. H., Salem, S. M. R., Abdelwahab, A. M., Alwutayd, K. M., Shaalan, M., Ismail, S. H., Abdelfattah, A. M., Khalid, A., Mansour, A. T., Hamed, H. S., & Youssuf, H. (2023). Nano-Curcumin/Chitosan Modulates Growth, Biochemical, Immune, and Antioxidative Profiles, and the Expression of Related Genes in Nile tilapia, Oreochromis niloticus. Fishes, 8(7), 333. https://doi.org/10.3390/fishes8070333