
Consistent Antipredator Behavioral Responses among Populations of Red River Pupfish with Disparate Predator Communities



Abstract
1. Introduction
2. Materials and Methods
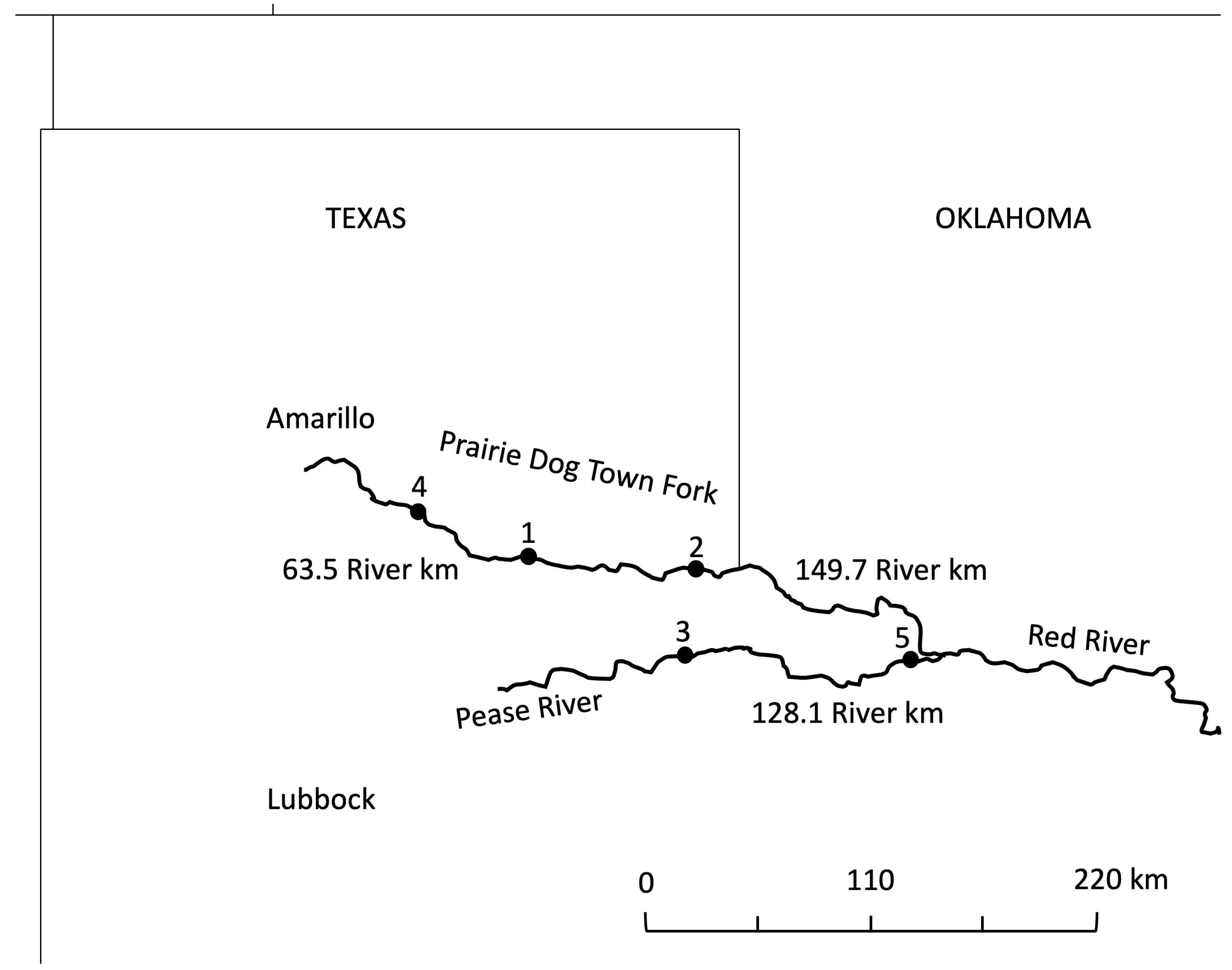
2.1. Population Selection and Fish Collection

{kind=link}
{kind=link}
{kind=link}
| Site/ Sample Size | Site Description | Location (Lat/Long) | Relative Predation Risk | Conductivity (µS/cm) | Other Fish Species Present |
|---|---|---|---|---|---|
| Site 1 n = 33 | Prairie Dog Town Fork of Red River at Hwy 256/70 | 34.628348/ −100.942 | Low | 18,159 | Plains killifish (Fundulus zebrinus) |
| Site 2 n = 22 | Prairie Dog Town Fork of Red River at Hwy 62/83 | 34.566653/ −100.196 | Low | 82,466 | Plains killifish |
| Site 3 n = 38 | Pease River 5.6 km below Hwy 62/68 | 34.194236/ −100.251 | Moderate | 24,151 | Plains killifish Red River shiner (Notropis bairdi) Plains minnow (Hypognathus placitus) |
| Site 4 n = 38 | Prairie Dog Town Fork of Red River at Hwy 207 | 34.837054/ −101.416 | Moderate | 25,403 | Plains killifish Red River shiner |
| Site 5 n = 29 | Pease River at Hwy 283 | 34.179296/ −99.2784 | High | 13,847 | Plains killifish Red River shiner Red shiner (Cyprinella lutrensis) Largemouth bass (Micropterus salmoides) Orange-spotted sunfish (Lepomis humilis) Western mosquitofish (Gambusia affinis) Bullhead minnow (Pimephales vigilax) |
2.2. Preparation of Alarm Cues and Evaluation of Fish Behavior
2.3. Data Analysis
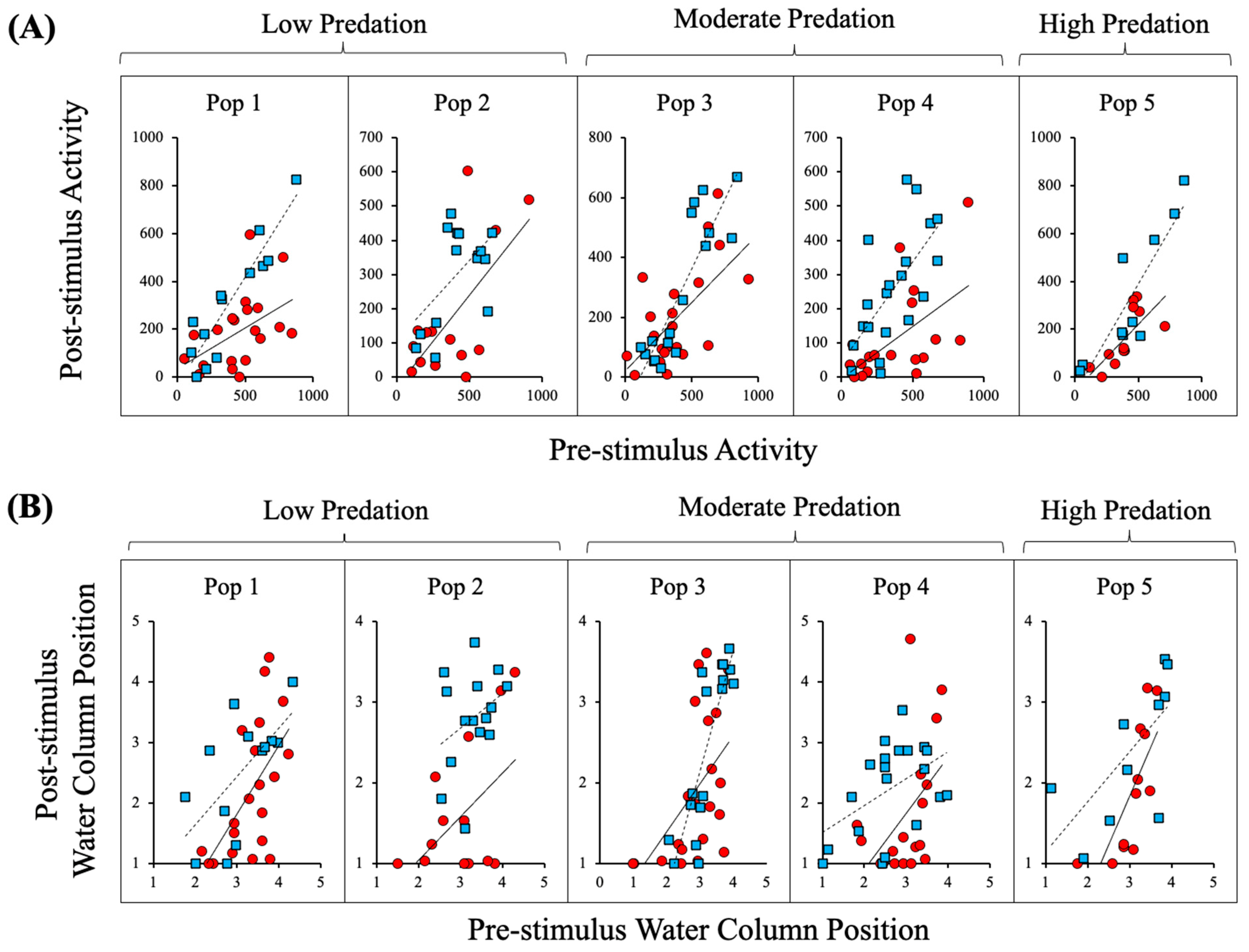
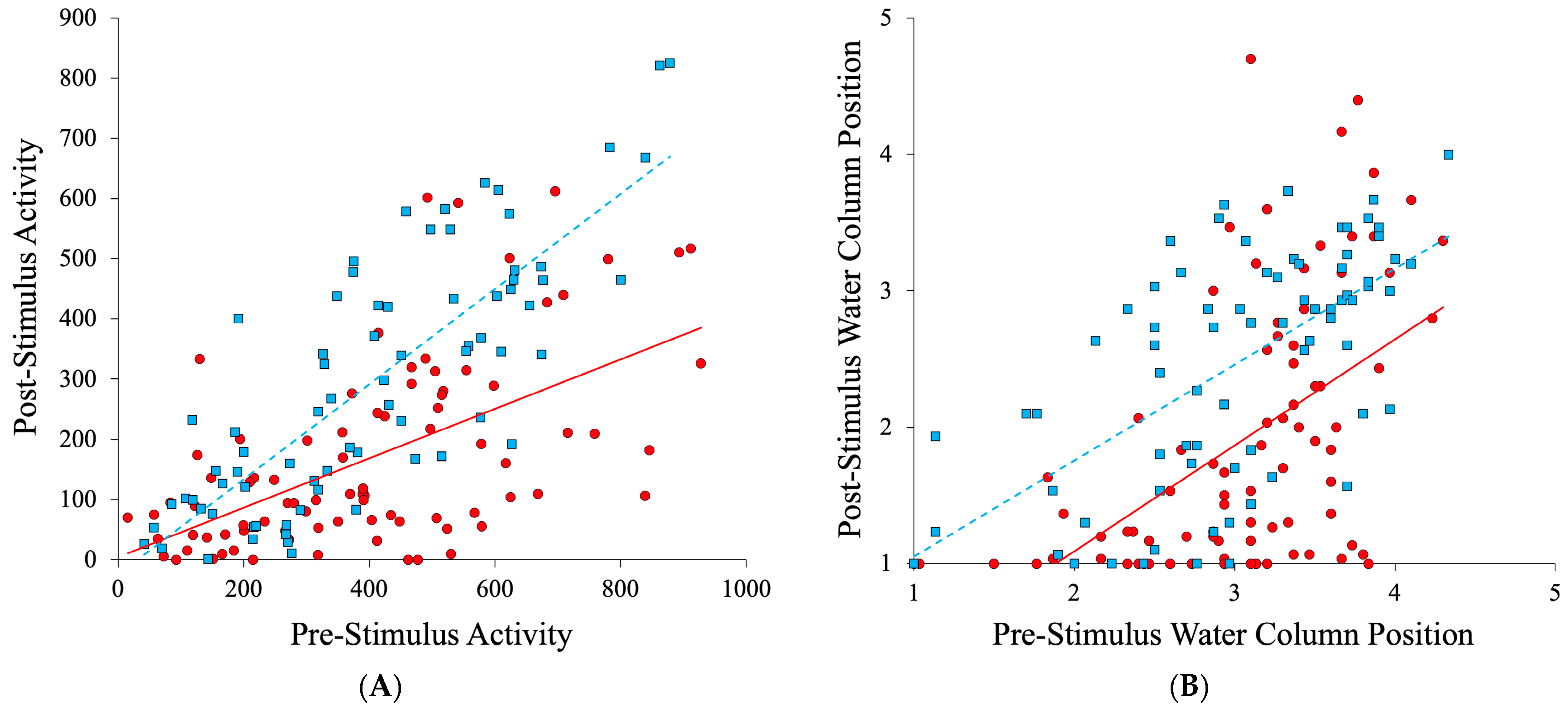
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Anton, A.; Geraldi, N.R.; Ricciardi, A.; Dick, J.T.A. Global determinants of prey naiveté to exotic predators. Proc. R. Soc. B Biol. Sci. 2020, 287, 20192978. [Google Scholar] [CrossRef] [PubMed]
- Miller, R.R. Man and the changing fish fauna of the southwestern United States. Mich. Acad. Sci. Arts. Lett. 1961, 46, 365–404. [Google Scholar]
- Minckley, W.L.; Deacon, J.E. Southwestern fishes and the enigma of “endangered species”. Science 1968, 159, 1424–1432. [Google Scholar] [CrossRef] [PubMed]
- Miller, R.R.; Williams, J.D.; Williams, J.E. Extinctions of North American fishes during the past century. Fisheries 1989, 14, 22–38. [Google Scholar] [CrossRef]
- Stockwell, C.A.; Schmelzer, M.; Gillis, B.; Anderson, C.; Wisenden, B.D. Ignorance is not bliss: Evolutionary naiveté in an endangered desert fish and implications for conservation. Proc. R. Soc. B 2022, 289, 20220752. [Google Scholar] [CrossRef] [PubMed]
- Jimenez, M.; Goodchild, S.C.; Stockwell, C.A.; Lema, S.C. Characterization and phylogenetic analysis of complete mitochondrial genomes for two desert cyprinodontoid fishes, Empetrichthys latos and Crenichthys baileyi. Gene 2017, 626, 163–172. [Google Scholar] [CrossRef]
- Guadalupe, K.; (Nevada Department of Wildlife, Las Vegas, NV, USA). Personal Communication, 2018.
- Goodchild, S.C.; Stockwell, C.A. An experimental test of novel ecological communities of imperiled and invasive species. Trans. Am. Fish. Soc. 2016, 145, 264–268. [Google Scholar] [CrossRef]
- Paulson, B. Ex Situ Analysis of Non-Native Species Impacts on Imperiled Desert Fishes. Master’s Thesis, North Dakota State University, Fargo, ND, USA, 2019. [Google Scholar]
- Ferrari, M.C.O.; Wisenden, B.D.; Chivers, D.P. Chemical ecology of predator-prey interactions in aquatic ecosystems: A review and prospectus. Can. J. Zool. 2010, 88, 698–724. [Google Scholar] [CrossRef]
- Echelle, A.A.; Echelle, A.F.; Hill, L.G. Interspecific interactions and limiting factors of abundance and distribution in the Red River Pupfish, Cyprinodon rubrofluviatilis. Am. Midl. Nat. 1972, 88, 109–130. [Google Scholar] [CrossRef]
- Ruppel, D.S. Factors Influencing Community Structure of Riverine Organisms: Implications for Imperiled Species Management. Ph.D. Dissertation, Texas State University, San Marcos, TX, USA, 2019. [Google Scholar]
- Nosil, P.; Crespi, B.J. Experimental evidence that predation promotes divergence in adaptive radiation. Proc. Natl. Acad. Sci. USA 2006, 103, 9090–9095. [Google Scholar] [CrossRef]
- Langerhans, R.B.; Gifford, M.E.; Joseph, E.O. Ecological speciation in Gambusia fishes. Evolution 2007, 61, 2056–2074. [Google Scholar] [CrossRef]
- Anderson, C.M. Pupfishes as a System to Test the Predator Naiveté Hypothesis. Master’s Thesis, North Dakota State University, Fargo, ND, USA, 2022. [Google Scholar]
- Wisenden, B.D. Quantifying anti-predator responses to chemical alarm cues. In Zebrafish Behavioral Protocols; Kalueff, A.V., Hart, P., LaPorte, J., Eds.; Humana Press Springer Science: New York, NY, USA, 2011; pp. 49–60. [Google Scholar]
- Cox, J.G.; Lima, S.L. Naiveté and an aquatic-terrestrial dichotomy in the effects of introduced predators. Trends Ecol. Evol. 2006, 21, 674–680. [Google Scholar] [CrossRef]
- Carthey, A.J.; Blumstein, D.T. Predicting predator recognition in a changing world. Trends Ecol. Evol. 2018, 33, 106–115. [Google Scholar] [CrossRef]
- Mathis, A.; Smith, R.J.F. Fathead minnows, Pimephales promelas, learn to recognize northern pike, Esox lucius, as predators on the basis of chemical stimuli from minnows in the pike’s diet. Anim. Behav. 1993, 46, 645–656. [Google Scholar] [CrossRef]
- Chivers, D.P.; Mirza, R.S. Predator diet cues and the assessment of predation risk by aquatic vertebrates: A review and prospectus. Chem. Sig. Vert. 2002, 9, 277–284. [Google Scholar] [CrossRef]
- Mitchell, M.D.; Chivers, D.P.; McCormick, M.I.; Ferrari, M.C. Learning to distinguish between predators and non-predators: Understanding the critical role of diet cues and predator odours in generalisation. Sci. Rep. 2015, 5, 13918. [Google Scholar] [CrossRef]
- Suboski, M.D. Releaser-induced recognition learning. Psychol. Rev. 1990, 97, 271. [Google Scholar] [CrossRef]
- Snider, M. Antipredator Behavior and Morphology in Isolated Cyprinodont Fishes. Master’s Thesis, North Dakota State University, Fargo, ND, USA, 2019. [Google Scholar]
- Wisenden, B.D.; Anderson, C.M.; Hanson, K.; Johnson, M.; Stockwell, C.A. A sniff and a whiff: Acquired predator recognition via epidermal alarm cues but not dietary alarm cues by isolated pupfish. Roy. Soc. Open Sci. 2023; submitted. [Google Scholar]
- Miller, R.R. The Cyprinodont Fishes of the Death Valley System of Eastern California and Southwestern Nevada; Miscellaneous Publications, University of Michigan, Museum of Zoology: Ann Arbor, MI, USA, 1948; Volume 68, pp. 1–55. [Google Scholar]
- Stoffels, R.J.; Rehwinkle, R.A.; Price, A.E.; Fagan, W.F. Dynamics of fish dispersal during river-floodplain connectivity and its implications for community assembly. Aquat. Sci. 2016, 78, 355–365. [Google Scholar] [CrossRef]
- Storfer, A.; Sih, A. Gene flow and ineffective antipredator behavior in a stream-breeding salamander. Evolution 1998, 52, 558–565. [Google Scholar] [CrossRef]
- Lenormand, T. Gene flow and the limits to natural selection. Trends Ecol. Evol. 2002, 17, 183–189. [Google Scholar] [CrossRef]
- Hendry, A.P.; Taylor, E.B.; McPhail, J.D. Adaptive divergence and the balance between selection and gene flow: Lake and stream stickleback in the Misty system. Evolution 2007, 56, 1199–1216. [Google Scholar] [CrossRef]
- Seghers, B.H. Schooling behavior in the guppy (Poecilia reticulata): An evolutionary response to predation. Evolution 1974, 28, 486–489. [Google Scholar] [CrossRef] [PubMed]
- Kelley, J.L.; Magurran, A.E. Effects of relaxed predation pressure on visual predator recognition in the guppy. Behav. Ecol. Sociobiol. 2003, 54, 225–232. [Google Scholar] [CrossRef]
- Hoagstrom, C.W.; Osborne, M. Biogeography of Cyprinodon across the Great Plains-Chihuahuan Desert region and adjacent areas. In Proceedings of the Desert Fishes Council Proceedings 2021:20-76, St. George, UT, USA, 17–21 November 2021; Desert Fishes Council: Austin, TX, USA, 2021. [Google Scholar]
- Rogowski, D.L.; Stockwell, C.A. Assessment of potential impacts of exotic species on populations of a threatened species, White Sands pupfish, Cyprinodon tularosa. Biol. Invasions 2006, 8, 79–87. [Google Scholar] [CrossRef]
- Craig, C. Descriptions, Classifications, and Explanations of Processes and Patterns Structuring and Maintaining Inland Fish Communities. Ph.D. Dissertation, Texas State University-San Marcos, San Marcos, TX, USA, 2020. [Google Scholar]
- McCormick, M.I.; Larson, J.K. Field verification of the use of chemical alarm cues in a coral reef fish. Coral Reefs 2007, 26, 571–576. [Google Scholar] [CrossRef]



| A. Post-Stimulus Activity | ||||
| Source | DF | SS | F | p |
| Population | 4, 140 | 55,425.2 | 0.86 | 0.490 |
| Cue | 1, 140 | 587,546.8 | 36.47 | <0.001 |
| Population × Cue | 4, 140 | 56,689.5 | 0.88 | 0.478 |
| Pre-Stimulus Activity | 1, 140 | 2,210,186.8 | 137.17 | <0.001 |
| Population × Pre-Stimulus Activity | 4, 140 | 79,236.3 | 1.23 | 0.301 |
| Cue × Pre-Stimulus Activity | 1, 140 | 159,458.0 | 9.90 | 0.002 |
| Population × Cue × Pre-Stimulus Activity | 4, 140 | 72,938.5 | 1.13 | 0.344 |
| B. Post-Stimulus Vertical Position | ||||
| Source | DF | SS | F | p |
| Population | 4, 140 | 0.06 | 0.03 | 0.998 |
| Cue | 1, 140 | 12.36 | 21.17 | <0.001 |
| Population × Cue | 4, 140 | 2.90 | 1.24 | 0.296 |
| Pre-Stimulus Vertical Position | 1, 140 | 38.55 | 66.01 | <0.001 |
| Population × Pre-Stimulus Vertical Position | 4, 140 | 2.40 | 1.03 | 0.395 |
| Treatment × Pre-Stimulus Vertical Position | 1, 140 | 0.23 | 0.39 | 0.533 |
| Population × Cue × Pre-Stimulus Vertical Position | 4, 140 | 4.90 | 2.10 | 0.085 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Anderson, C.M.; Wisenden, B.D.; Craig, C.A.; Stockwell, C.A. Consistent Antipredator Behavioral Responses among Populations of Red River Pupfish with Disparate Predator Communities. Fishes 2023, 8, 315. https://doi.org/10.3390/fishes8060315
Anderson CM, Wisenden BD, Craig CA, Stockwell CA. Consistent Antipredator Behavioral Responses among Populations of Red River Pupfish with Disparate Predator Communities. Fishes. 2023; 8(6):315. https://doi.org/10.3390/fishes8060315
Chicago/Turabian StyleAnderson, Cody M., Brian D. Wisenden, Cody A. Craig, and Craig A. Stockwell. 2023. "Consistent Antipredator Behavioral Responses among Populations of Red River Pupfish with Disparate Predator Communities" Fishes 8, no. 6: 315. https://doi.org/10.3390/fishes8060315
APA StyleAnderson, C. M., Wisenden, B. D., Craig, C. A., & Stockwell, C. A. (2023). Consistent Antipredator Behavioral Responses among Populations of Red River Pupfish with Disparate Predator Communities. Fishes, 8(6), 315. https://doi.org/10.3390/fishes8060315